
Influence of Spine Curvature on the Efficacy of Transcutaneous Lumbar Spinal Cord Stimulation


 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
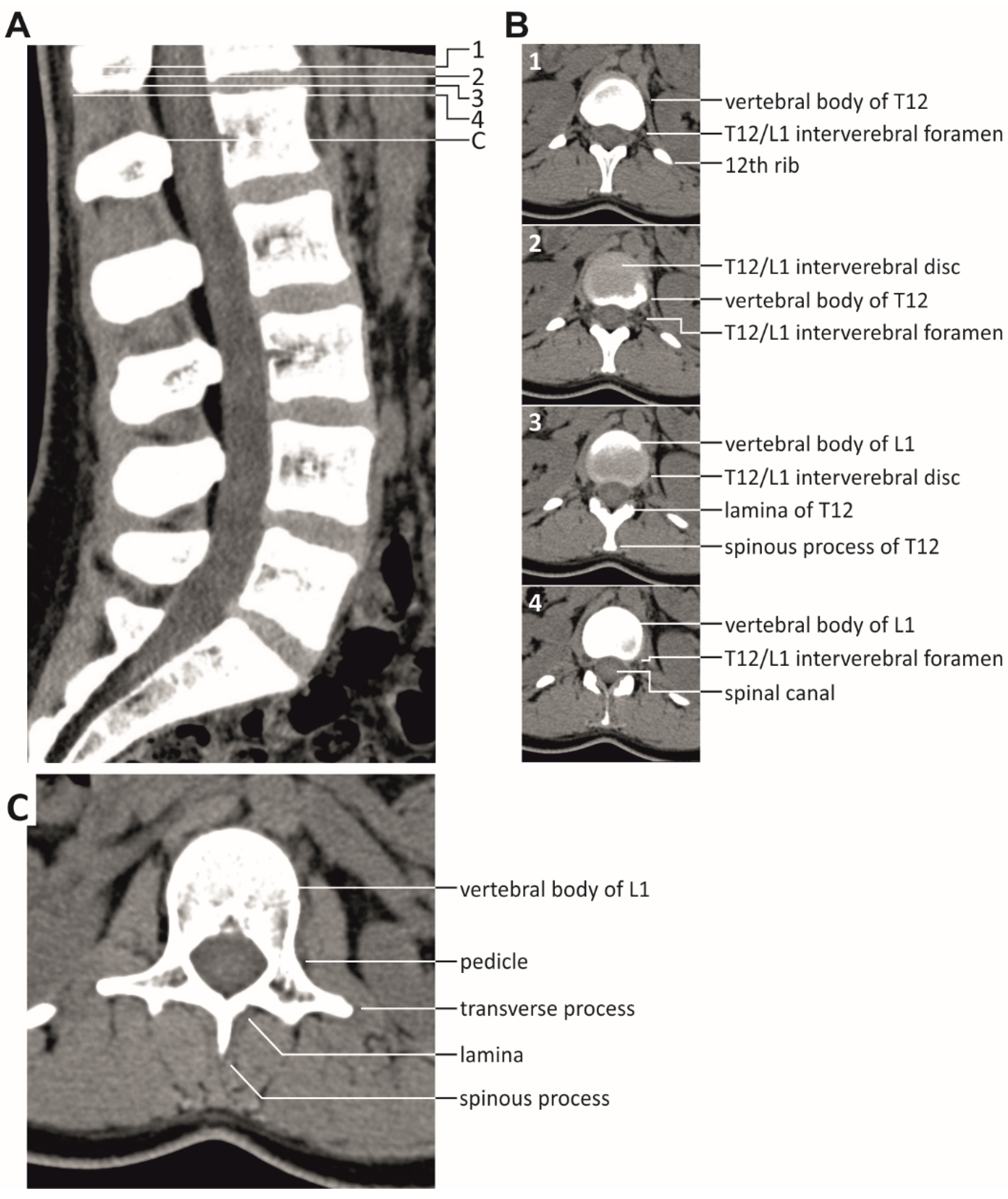
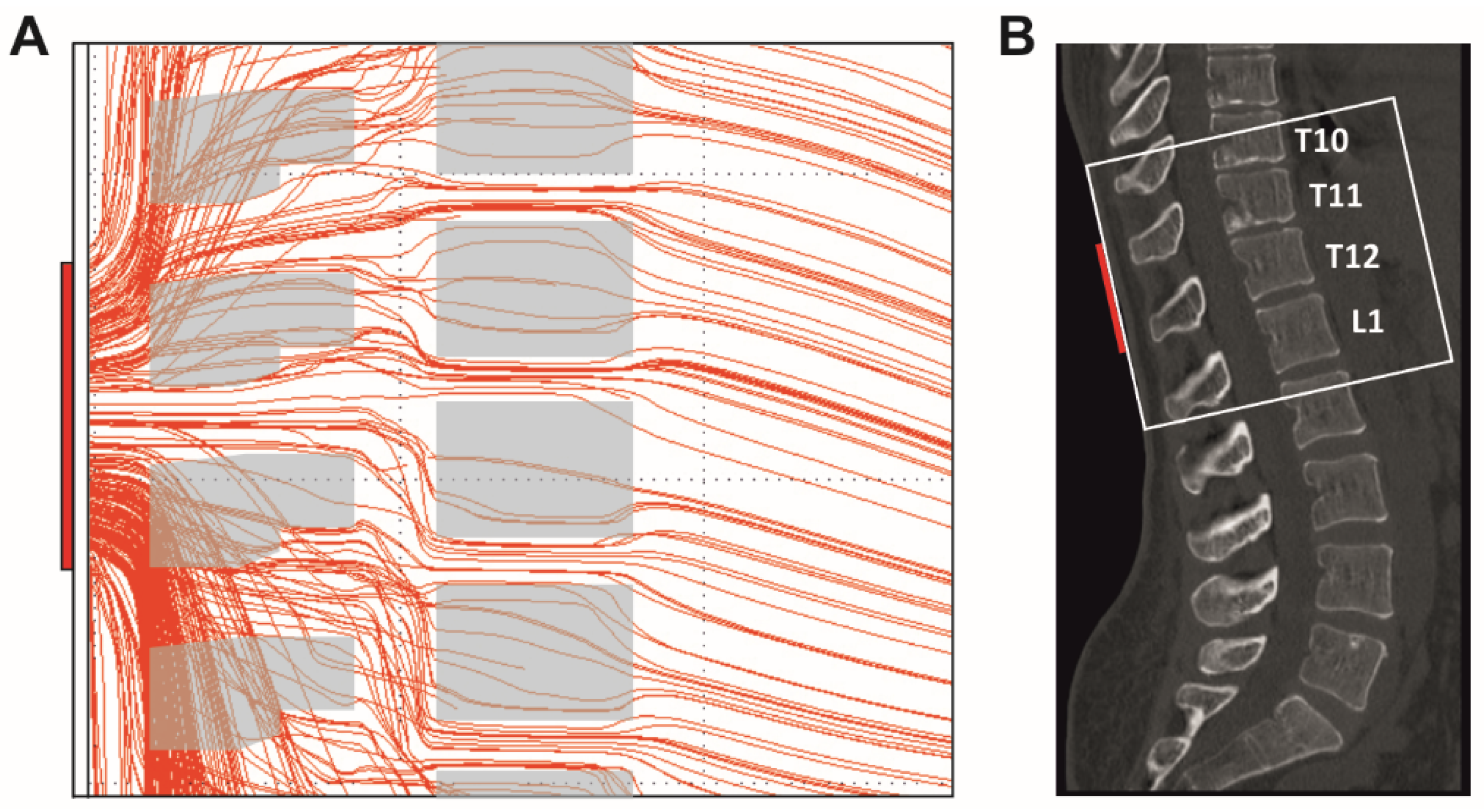
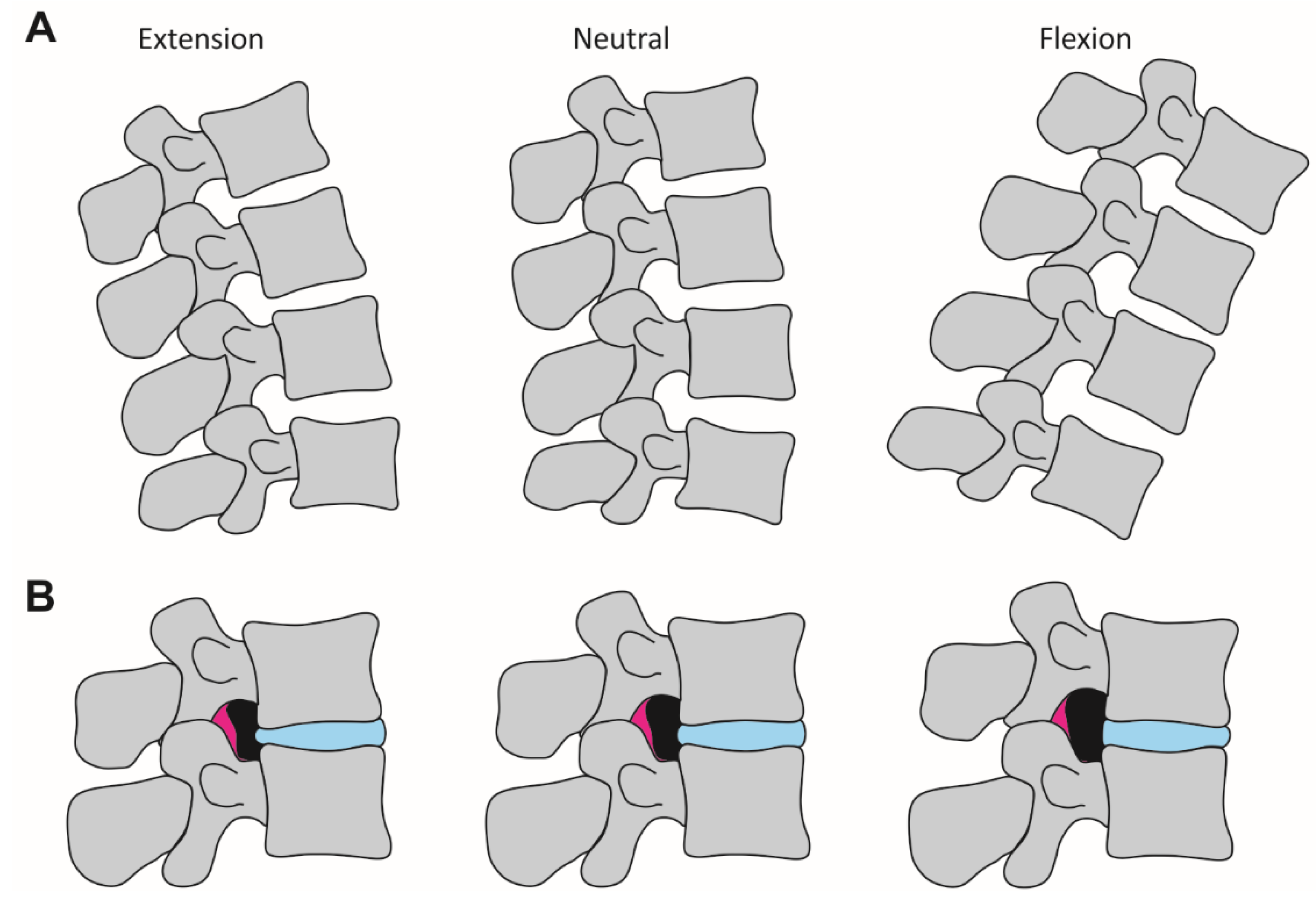
2.2. Investigated Body Positions and Spine Alignments
2.3. Electromyographic Recordings
2.4. Stimulation Procedures
2.5. Study Protocol
2.6. Data Analysis
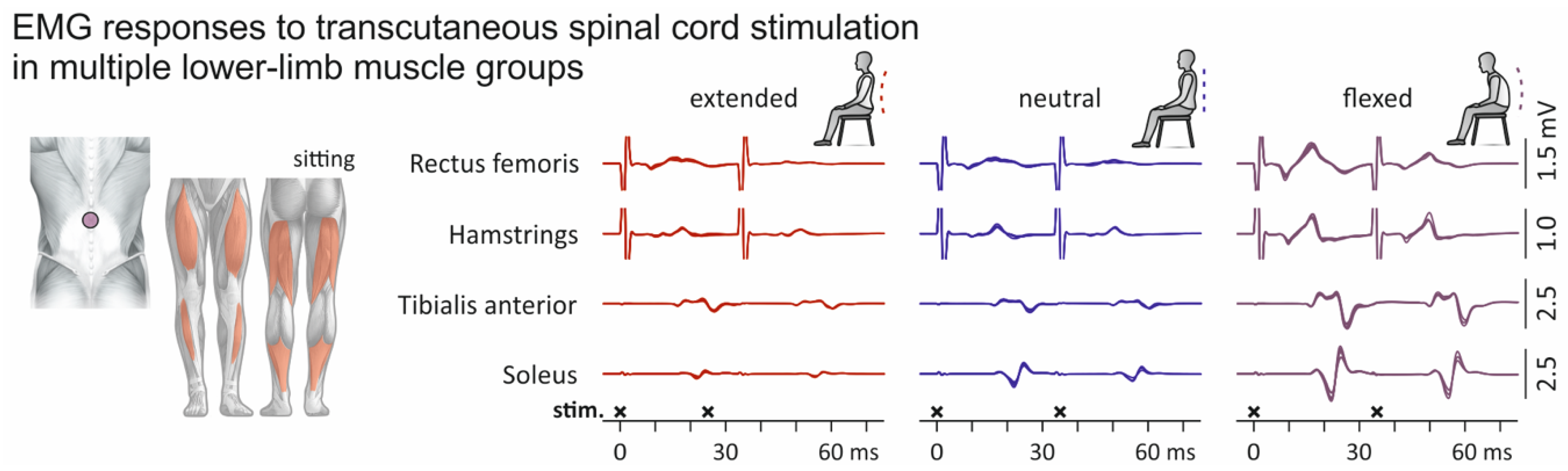
3. Results
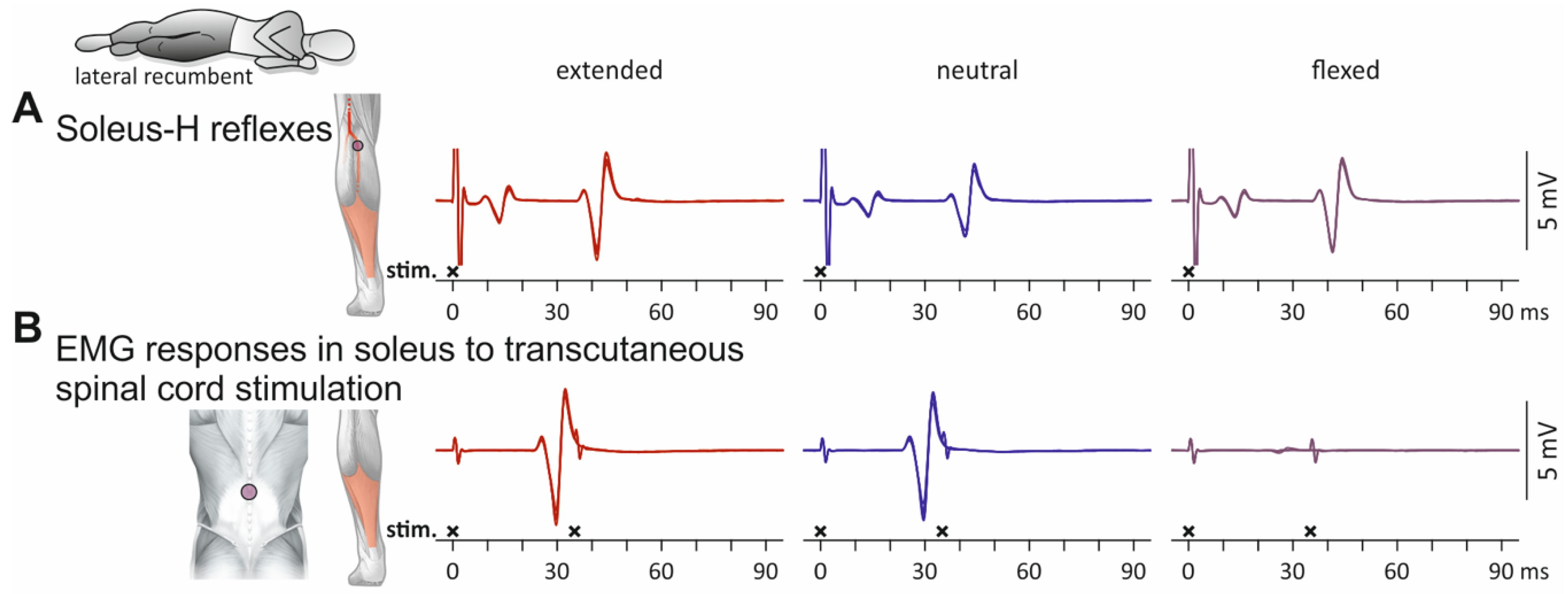
3.1. Influence of Spine Alignment Condition on H Reflexes and PRM Reflexes in Soleus
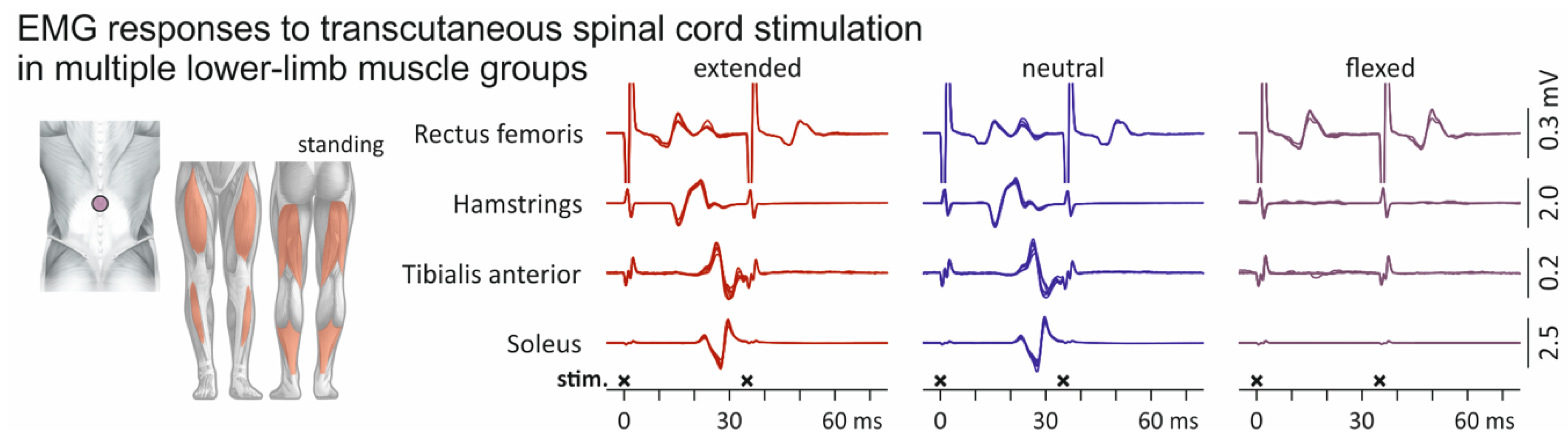
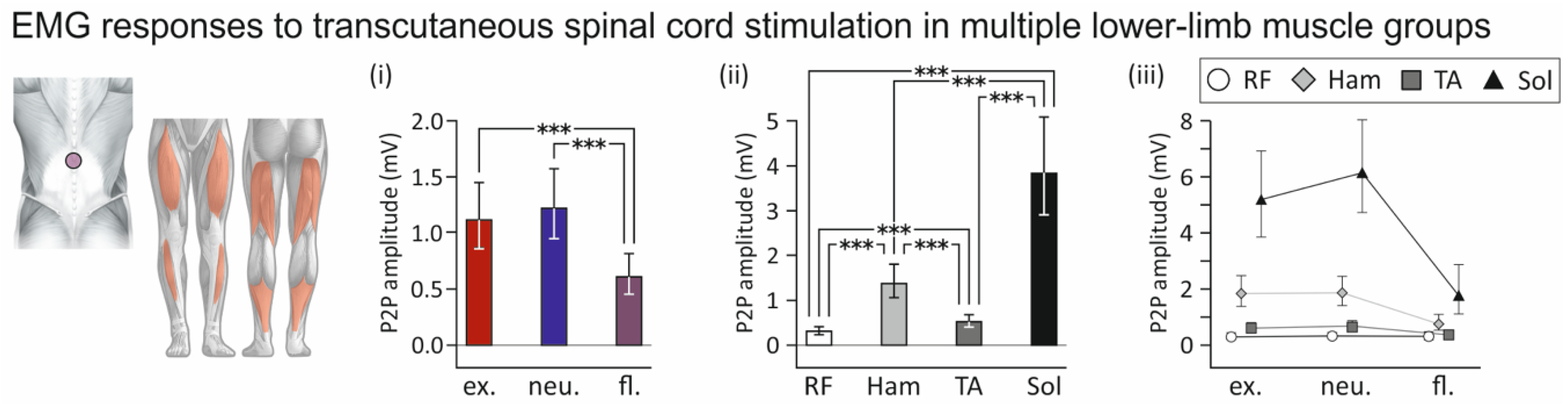
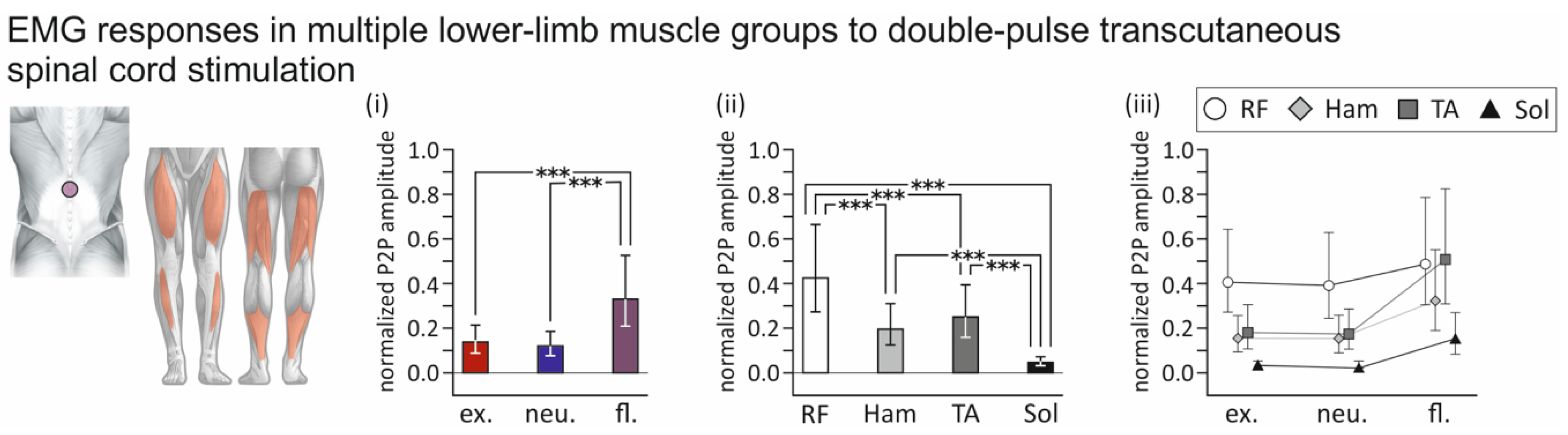
3.2. Influence of Spine Alignment Condition on Responses in Thigh and Leg Muscles Evoked by Transcutaneous Spinal Cord Stimulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Minassian, K.; Persy, I.; Rattay, F.; Dimitrijevic, M.R.; Hofer, C.; Kern, H. Posterior root-muscle reflexes elicited by transcutaneous stimulation of the human lumbosacral cord. Muscle Nerve 2007, 35, 327–336. [Google Scholar] [CrossRef]
- Minassian, K.; Freundl, B.; Hofstoetter, U.S. The posterior root-muscle reflex. In Neurophysiology in Neurosurgery; Elsevier: Amsterdam, The Netherlands, 2020; pp. 239–253. [Google Scholar]
- Hofstoetter, U.S.; Freundl, B.; Binder, H.; Minassian, K. Recovery cycles of posterior root-muscle reflexes evoked by transcutaneous spinal cord stimulation and of the H reflex in individuals with intact and injured spinal cord. PLoS ONE 2019, 14, e0227057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, J.C.; Stein, R.B.; Roy, F.D. Post-activation depression in the human soleus muscle using peripheral nerve and transcutaneous spinal stimulation. Neurosci. Lett. 2015, 589, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Kitano, K.; Koceja, D.M. Spinal reflex in human lower leg muscles evoked by transcutaneous spinal cord stimulation. J. Neurosci. Methods 2009, 180, 111–115. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Freundl, B.; Binder, H.; Minassian, K. Common neural structures activated by epidural and transcutaneous lumbar spinal cord stimulation: Elicitation of posterior root-muscle reflexes. PLoS ONE 2018, 13, e0192013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dy, C.J.; Gerasimenko, Y.P.; Edgerton, V.R.; Dyhre-Poulsen, P.; Courtine, G.; Harkema, S.J. Phase-dependent modulation of percutaneously elicited multisegmental muscle responses after spinal cord injury. J. Neurophysiol. 2010, 103, 2808–2820. [Google Scholar] [CrossRef] [Green Version]
- Saito, A.; Nakagawa, K.; Masugi, Y.; Nakazawa, K. Inter-muscle differences in modulation of motor evoked potentials and posterior root-muscle reflexes evoked from lower-limb muscles during agonist and antagonist muscle contractions. Exp. Brain Res. 2021, 239, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Hofstoetter, U.S.; Minassian, K.; Hofer, C.; Mayr, W.; Rattay, F.; Dimitrijevic, M.R. Modification of reflex responses to lumbar posterior root stimulation by motor tasks in healthy subjects. Artif. Organs 2008, 32, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Hofstoetter, U.S.; Danner, S.M.; Mayr, W.; Bruce, J.A.; McKay, W.B.; Tansey, K.E. Spinal Rhythm Generation by Step-Induced Feedback and Transcutaneous Posterior Root Stimulation in Complete Spinal Cord-Injured Individuals. Neurorehabil. Neural Repair 2016, 30, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Estes, S.; Zarkou, A.; Hope, J.M.; Suri, C.; Field-Fote, E.C. Combined Transcutaneous Spinal Stimulation and Locomotor Training to Improve Walking Function and Reduce Spasticity in Subacute Spinal Cord Injury: A Randomized Study of Clinical Feasibility and Efficacy. J. Clin. Med. 2021, 10, 1167. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Freundl, B.; Danner, S.M.; Krenn, M.J.; Mayr, W.; Binder, H.; Minassian, K. Transcutaneous Spinal Cord Stimulation Induces Temporary Attenuation of Spasticity in Individuals with Spinal Cord Injury. J. Neurotrauma 2020, 37, 481–493. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Freundl, B.; Lackner, P.; Binder, H. Transcutaneous Spinal Cord Stimulation Enhances Walking Performance and Reduces Spasticity in Individuals with Multiple Sclerosis. Brain Sci. 2021, 11, 472. [Google Scholar] [CrossRef]
- Roberts, B.W.R.; Atkinson, D.A.; Manson, G.A.; Markley, R.; Kaldis, T.; Britz, G.W.; Horner, P.J.; Vette, A.H.; Sayenko, D.G. Transcutaneous spinal cord stimulation improves postural stability in individuals with multiple sclerosis. Mult. Scler. Relat. Disord. 2021, 52, 103009. [Google Scholar] [CrossRef]
- Ladenbauer, J.; Minassian, K.; Hofstoetter, U.S.; Dimitrijevic, M.R.; Rattay, F. Stimulation of the human lumbar spinal cord with implanted and surface electrodes: A computer simulation study. IEEE Trans. Neural Syst. Rehabil. Eng. 2010, 18, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Danner, S.M.; Hofstoetter, U.S.; Ladenbauer, J.; Rattay, F.; Minassian, K. Can the human lumbar posterior columns be stimulated by transcutaneous spinal cord stimulation? A modeling study. Artif. Organs 2011, 35, 257–262. [Google Scholar] [CrossRef]
- Száva, Z.; Danner, S.M.; Minassian, K. Transcutaneous Electrical Spinal Cord Stimulation: Biophysics of a New Rehabilitation Method after Spinal Cord Injury; VDM Verlag Dr. Müller: Saarbrücken, Germany, 2011. [Google Scholar]
- Hofstoetter, U.S.; Danner, S.M.; Minassian, K. Paraspinal Magnetic and Transcutaneous Electrical Stimulation. In Encyclopedia of Computational Neuroscience; Jaeger, D., Jung, R., Eds.; Springer: New York, NY, USA, 2015; pp. 2194–2212. [Google Scholar]
- Danner, S.M.; Krenn, M.; Hofstoetter, U.S.; Toth, A.; Mayr, W.; Minassian, K. Body Position Influences Which Neural Structures Are Recruited by Lumbar Transcutaneous Spinal Cord Stimulation. PLoS ONE 2016, 11, e0147479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geddes, L.A.; Baker, L.E. The specific resistance of biological material—A compendium of data for the biomedical engineer and physiologist. Med. Biol. Eng. 1967, 5, 271–293. [Google Scholar] [CrossRef]
- White, A.A.; Panjabi, M.M. Clinical Biomechanics of the Spine; JB Lippincott: Philadelphia, PA, USA, 1990. [Google Scholar]
- Sherwood, A.M.; McKay, W.B.; Dimitrijević, M.R. Motor control after spinal cord injury: Assessment using surface EMG. Muscle Nerve 1996, 19, 966–979. [Google Scholar] [CrossRef]
- Pierrot-Deseilligny, E.; Mazevet, D. The monosynaptic reflex: A tool to investigate motor control in humans. Interest and limits. Clin. Neurophysiol. 2000, 30, 67–80. [Google Scholar] [CrossRef]
- Burke, D. Clinical uses of H reflexes of upper and lower limb muscles. Clin. Neurophysiol. Pract. 2016, 1, 9–17. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kristensen, K.; Nielsen, A.; Berg, C.; Skaug, H.; Bell, B. TMB: Automatic Differentiation and Laplace Approximation. J. Stat. Softw. 2016, 70, 1–21. [Google Scholar] [CrossRef]
- Thygesen, U.; Albertsen, C.; Berg, C.; Kristensen, K.; Nielsen, A. Validation of Ecological State Space Models Using the Laplace Approximation. Environ. Ecol. Stat. 2017, 24, 317–339. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.E.; Kristensen, K.; Benthem, K.J.V.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R Journal 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Rattay, F.; Minassian, K.; Dimitrijevic, M.R. Epidural electrical stimulation of posterior structures of the human lumbosacral cord: 2. quantitative analysis by computer modeling. Spinal Cord 2000, 38, 473–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formento, E.; Minassian, K.; Wagner, F.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Rowald, A.; Bloch, J.; Micera, S.; Capogrosso, M.; Courtine, G. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 2018, 21, 1728–1741. [Google Scholar] [CrossRef] [PubMed]
- Capogrosso, M.; Wenger, N.; Raspopovic, S.; Musienko, P.; Beauparlant, J.; Bassi Luciani, L.; Courtine, G.; Micera, S. A computational model for epidural electrical stimulation of spinal sensorimotor circuits. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 19326–19340. [Google Scholar] [CrossRef]
- Minassian, K.; Jilge, B.; Rattay, F.; Pinter, M.M.; Binder, H.; Gerstenbrand, F.; Dimitrijevic, M.R. Stepping-like movements in humans with complete spinal cord injury induced by epidural stimulation of the lumbar cord: Electromyographic study of compound muscle action potentials. Spinal Cord 2004, 42, 401–416. [Google Scholar] [CrossRef] [Green Version]
- Danner, S.M.; Hofstoetter, U.S.; Minassian, K. Finite Element Models of Transcutaneous Spinal Cord Stimulation. In Encyclopedia of Computational Neuroscience; Jaeger, D., Jung, R., Eds.; Springer: New York, NY, USA, 2015; pp. 1197–1202. [Google Scholar]
- Zhu, Y.; Starr, A.; Haldeman, S.; Chu, J.K.; Sugerman, R.A. Soleus H-reflex to S1 nerve root stimulation. Electroencephalogr. Clin. Neurophysiol. 1998, 109, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Calvert, J.S.; Gill, M.L.; Linde, M.B.; Veith, D.D.; Thoreson, A.R.; Lopez, C.; Lee, K.H.; Gerasimenko, Y.P.; Edgerton, V.R.; Lavrov, I.A.; et al. Voluntary Modulation of Evoked Responses Generated by Epidural and Transcutaneous Spinal Stimulation in Humans with Spinal Cord Injury. J. Clin. Med. 2021, 10, 4898. [Google Scholar] [CrossRef]
- Wecht, J.; Savage, W.; Famodimu, G.; Mendez, G.; Levine, J.; Maher, M.; Weir, J.; Wecht, J.; Carmel, J.; Wu, Y.; et al. Posteroanterior Cervical Transcutaneous Spinal Cord Stimulation: Interactions with Cortical and Peripheral Nerve Stimulation. J. Clin. Med. 2021, 10, 5304. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Perret, I.; Bayart, A.; Lackner, P.; Binder, H.; Freundl, B.; Minassian, K. Spinal motor mapping by epidural stimulation of lumbosacral posterior roots in humans. iScience 2020, 24, 101930. [Google Scholar] [CrossRef]
- Meyer, C.; Hofstoetter, U.S.; Hubli, M.; Hassani, R.H.; Rinaldo, C.; Curt, A.; Bolliger, M. Immediate Effects of Transcutaneous Spinal Cord Stimulation on Motor Function in Chronic, Sensorimotor Incomplete Spinal Cord Injury. J. Clin. Med. 2020, 9, 3541. [Google Scholar] [CrossRef]
- Estes, S.P.; Iddings, J.A.; Field-Fote, E.C. Priming Neural Circuits to Modulate Spinal Reflex Excitability. Front. Neurol. 2017, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Al’joboori, Y.; Massey, S.J.; Knight, S.L.; Donaldson, N.d.N.; Duffell, L.D. The Effects of Adding Transcutaneous Spinal Cord Stimulation (tSCS) to Sit-To-Stand Training in People with Spinal Cord Injury: A Pilot Study. J. Clin. Med. 2020, 9, 2765. [Google Scholar] [CrossRef] [PubMed]
- Kumru, H.; Flores, Á.; Rodríguez-Cañón, M.; Edgerton, V.R.; García, L.; Benito-Penalva, J.; Navarro, X.; Gerasimenko, Y.; García-Alías, G.; Vidal, J. Cervical Electrical Neuromodulation Effectively Enhances Hand Motor Output in Healthy Subjects by Engaging a Use-Dependent Intervention. J. Clin. Med. 2021, 10, 195. [Google Scholar] [CrossRef]
- Sandler, E.B.; Condon, K.; Field-Fote, E.C. Efficacy of Transcutaneous Spinal Stimulation versus Whole Body Vibration for Spasticity Reduction in Persons with Spinal Cord Injury. J. Clin. Med. 2021, 10, 3267. [Google Scholar] [CrossRef] [PubMed]
- Kagamihara, Y.; Hayashi, A.; Okuma, Y.; Nagaoka, M.; Nakajima, Y.; Tanaka, R. Reassessment of H-reflex recovery curve using the double stimulation procedure. Muscle Nerve 1998, 21, 352–360. [Google Scholar] [CrossRef]
- Halter, J.; Dolenc, V.; Dimitrijevic, M.R.; Sharkey, P.C. Neurophysiological assessment of electrode placement in the spinal cord. Appl. Neurophysiol. 1983, 46, 124–128. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.S.; Rattay, F. Transcutaneous lumbar posterior root stimulation for motor control studies and modification of motor activity after spinal cord injury. In Restorative Neurology of Spinal Cord Injury; Dimitrijevic, M., Kakulas, B., McKay, W., Vrbova, G., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 226–255. [Google Scholar]
- Schönström, N.; Lindahl, S.; Willén, J.; Hansson, T. Dynamic changes in the dimensions of the lumbar spinal canal: An experimental study in vitro. J. Orthop. Res. 1989, 7, 115–121. [Google Scholar] [CrossRef]
- Schmid, M.R.; Stucki, G.; Duewell, S.; Wildermuth, S.; Romanowski, B.; Hodler, J. Changes in cross-sectional measurements of the spinal canal and intervertebral foramina as a function of body position: In vivo studies on an open-configuration MR system. Am. J. Roentgenol. 1999, 172, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Weishaupt, D.; Boxheimer, L. Magnetic Resonance Imaging of the Weight-Bearing Spine. Semin. Musculoskelet. Radiol. 2003, 7, 277–286. [Google Scholar]
- Mayoux-Benhamou, M.A.; Revel, M.; Aaron, C.; Chomette, G.; Amor, B. A morphometric study of the lumbar foramen. Surg. Radiol. Anat. 1989, 11, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, Y.; Bashir, W.A.; Smith, F.W.; Magnusson, M.L.; Pope, M.H.; Takahashi, K. Postural Changes of the Dural Sac in the Lumbar Spines of Asymptomatic Individuals Using Positional Stand-Up Magnetic Resonance Imaging. Spine 2007, 32, E136–E140. [Google Scholar] [CrossRef]
- Hey, H.W.D.; Lau, E.T.-C.; Tan, K.-A.; Lim, J.L.; Choong, D.; Lau, L.-L.; Liu, K.-P.G.; Wong, H.-K. Lumbar Spine Alignment in Six Common Postures. Spine (Phila. PA, 1976). 2017, 42, 1447–1455. [Google Scholar] [CrossRef]
- Hindle, R.J.; Pearcy, M.J.; Cross, A.T.; Miller, D.H. Three-dimensional kinematics of the human back. Clin. Biomech. 1990, 5, 218–228. [Google Scholar] [CrossRef]
- Dimitrijevic, M.R.; Faganel, J.; Sherwood, A.M. Spinal cord stimulation as a tool for physiological research. Appl. Neurophysiol. 1983, 46, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Persy, I.; Rattay, F.; Pinter, M.M.; Kern, H.; Dimitrijevic, M.R. Human lumbar cord circuitries can be activated by extrinsic tonic input to generate locomotor-like activity. Hum. Mov. Sci. 2007, 26, 275–295. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, V.E.; Hofstoetter, U.S.; Rienmüller, A.; Száva, Z.; Krenn, M.J.; Minassian, K.; Danner, S.M. Influence of Spine Curvature on the Efficacy of Transcutaneous Lumbar Spinal Cord Stimulation. J. Clin. Med. 2021, 10, 5543. https://doi.org/10.3390/jcm10235543
Binder VE, Hofstoetter US, Rienmüller A, Száva Z, Krenn MJ, Minassian K, Danner SM. Influence of Spine Curvature on the Efficacy of Transcutaneous Lumbar Spinal Cord Stimulation. Journal of Clinical Medicine. 2021; 10(23):5543. https://doi.org/10.3390/jcm10235543
Chicago/Turabian StyleBinder, Veronika E., Ursula S. Hofstoetter, Anna Rienmüller, Zoltán Száva, Matthias J. Krenn, Karen Minassian, and Simon M. Danner. 2021. "Influence of Spine Curvature on the Efficacy of Transcutaneous Lumbar Spinal Cord Stimulation" Journal of Clinical Medicine 10, no. 23: 5543. https://doi.org/10.3390/jcm10235543
APA StyleBinder, V. E., Hofstoetter, U. S., Rienmüller, A., Száva, Z., Krenn, M. J., Minassian, K., & Danner, S. M. (2021). Influence of Spine Curvature on the Efficacy of Transcutaneous Lumbar Spinal Cord Stimulation. Journal of Clinical Medicine, 10(23), 5543. https://doi.org/10.3390/jcm10235543