Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview




Abstract
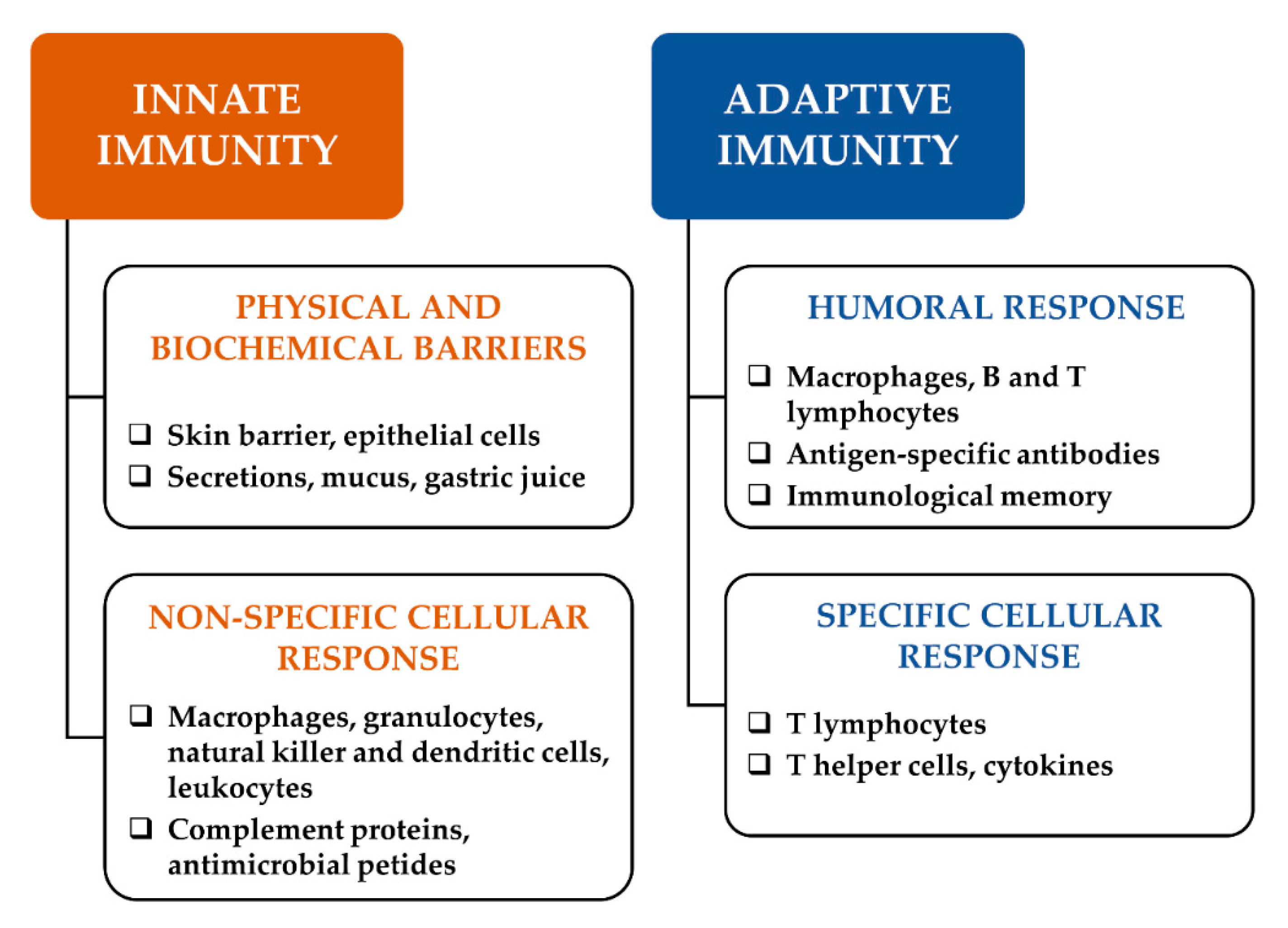
1. Introduction
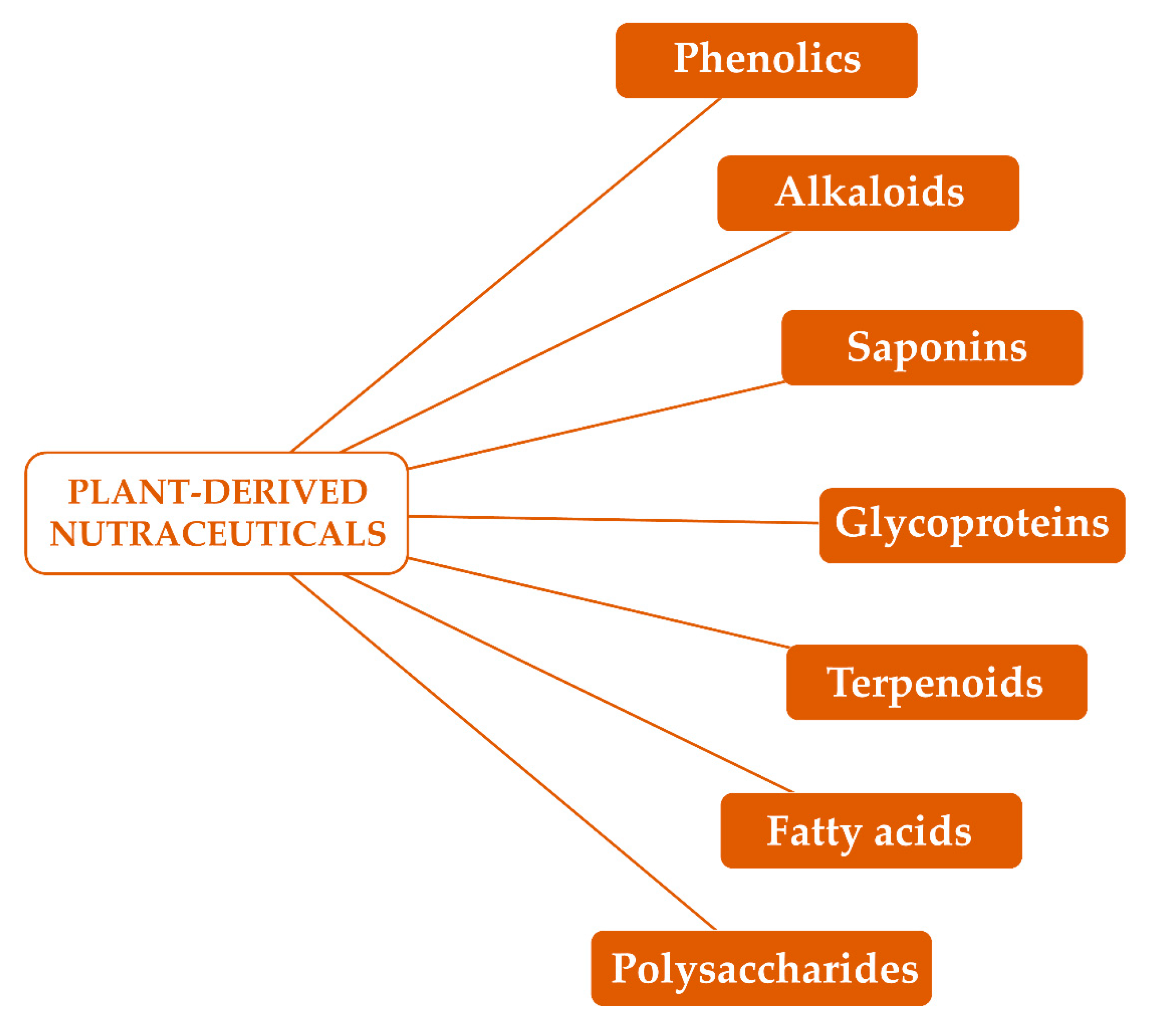
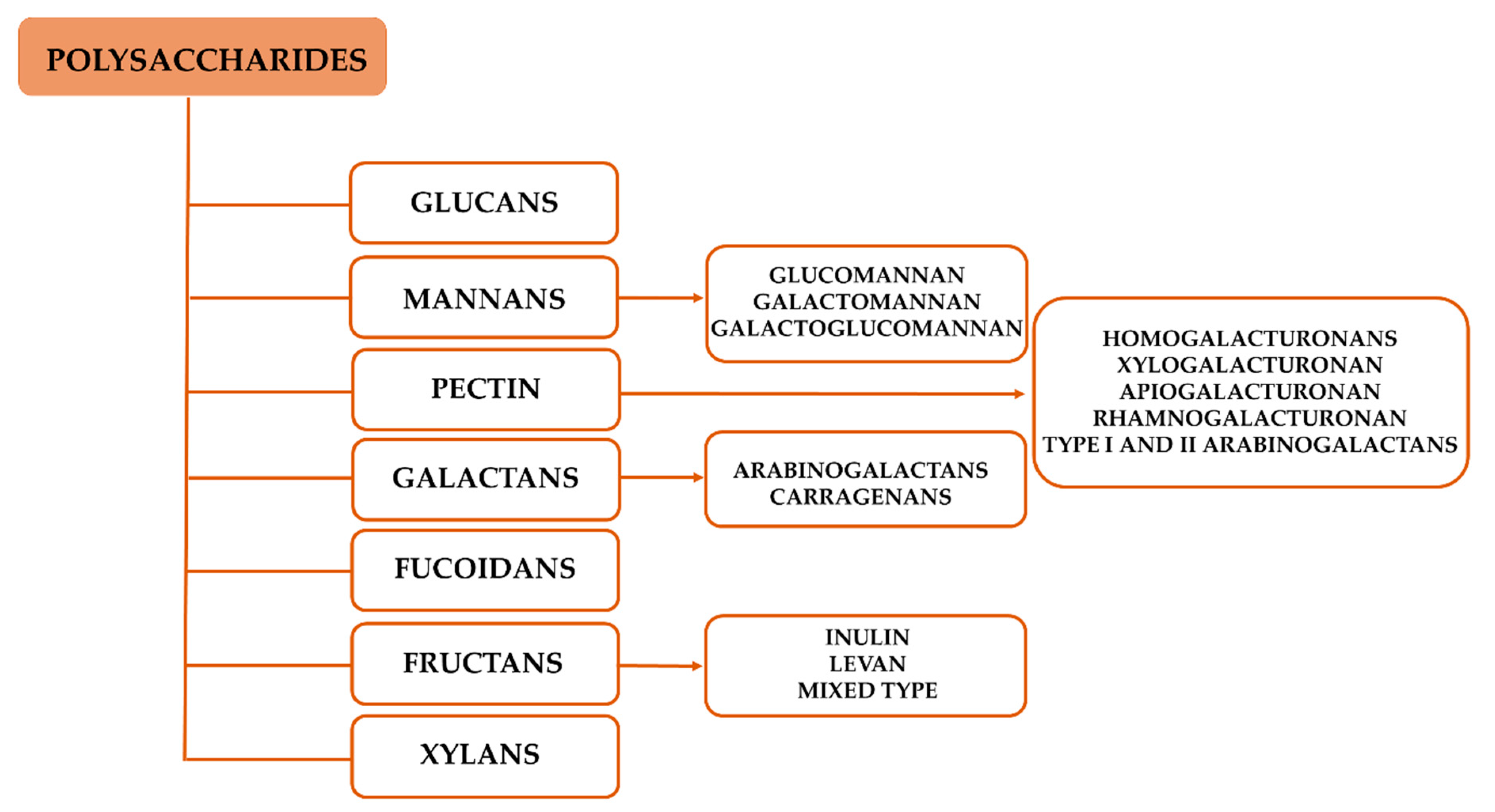
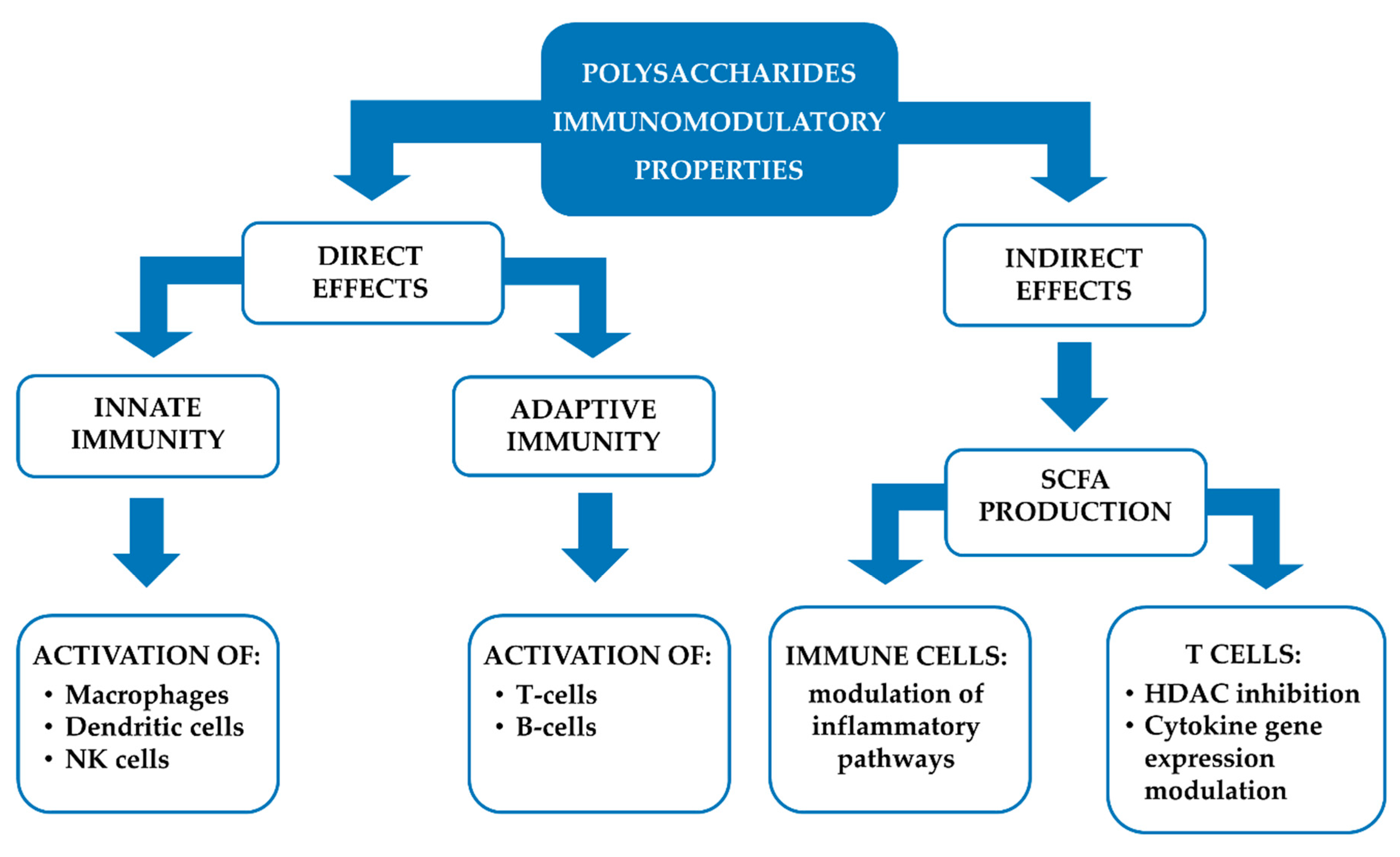
2. Polysaccharides
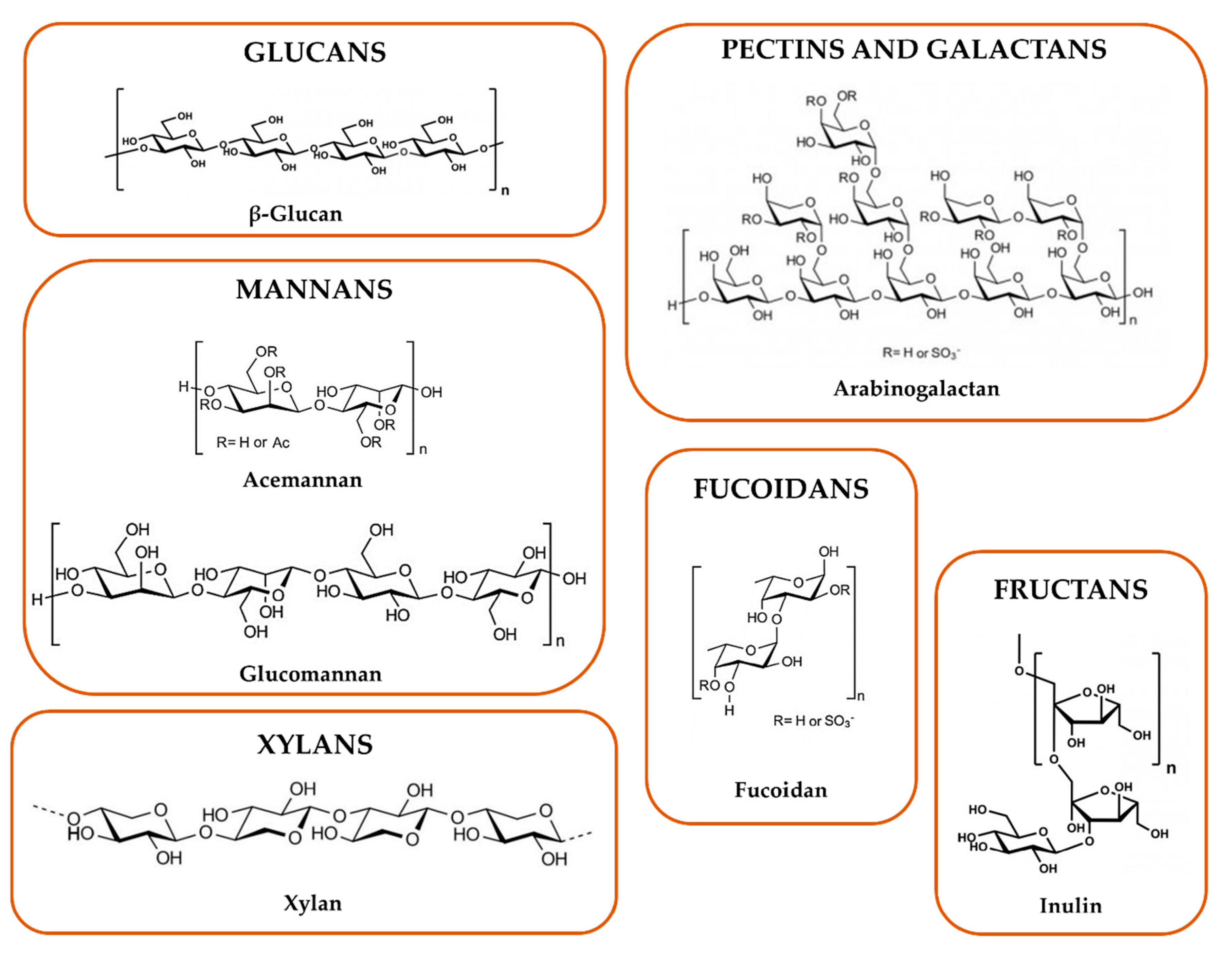
2.1. Chemical Features
2.2. Natural Occurrence
2.3. Pharmacological Properties: Preclinical Evidence
2.4. Clinical Studies
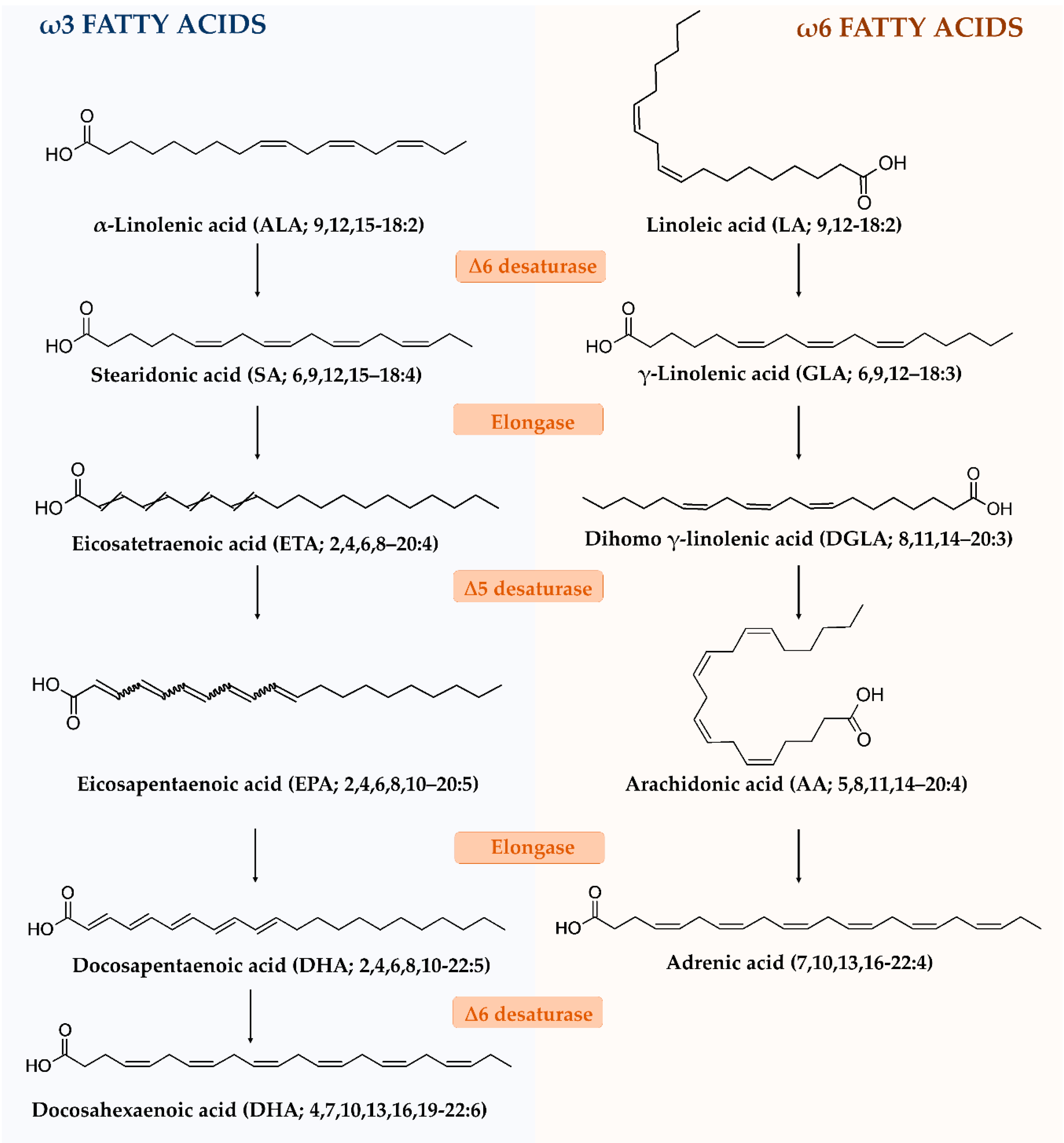
3. Fatty Acids
3.1. Chemical Features
3.2. Natural Occurrence
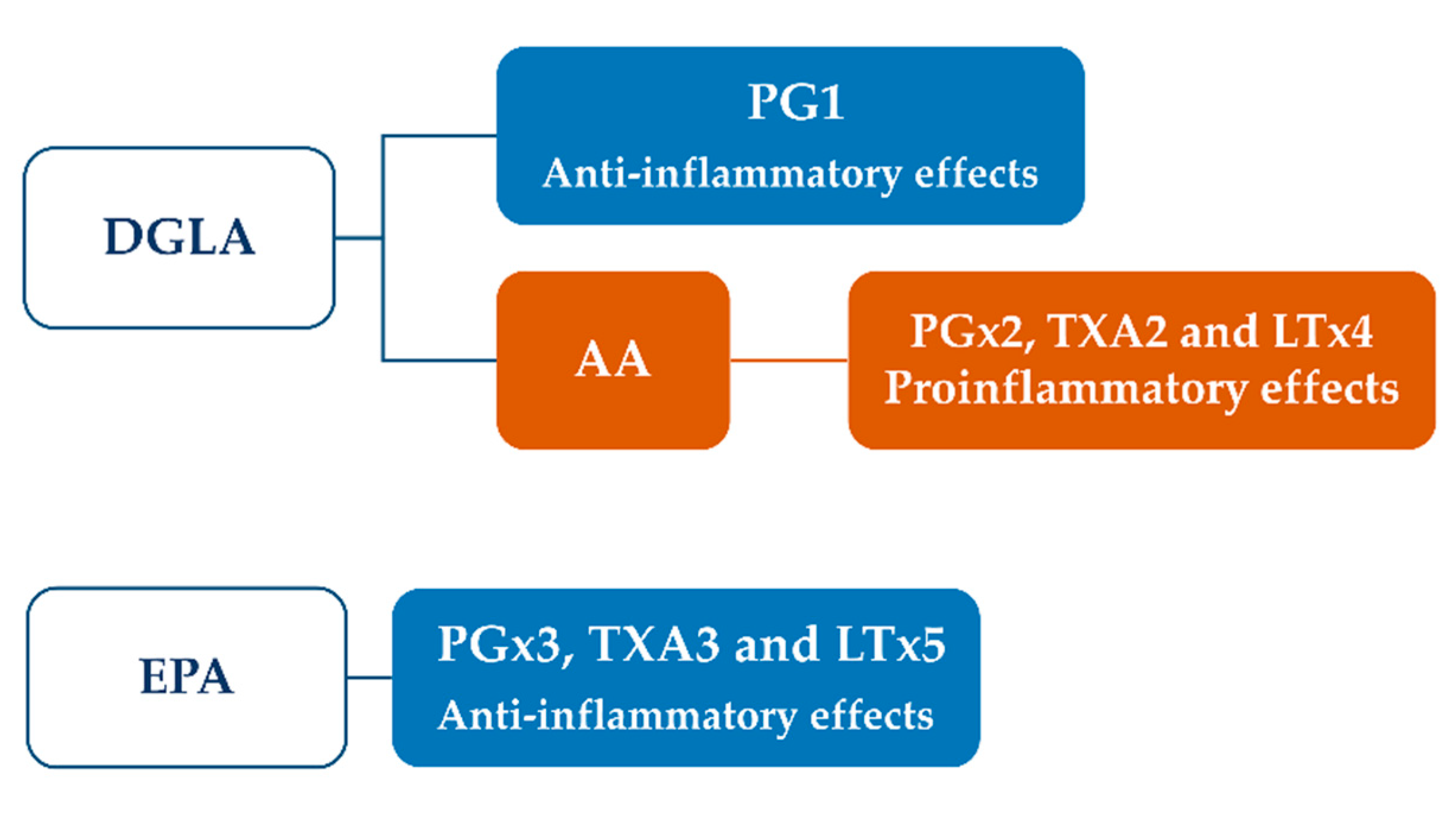
3.3. Pharmacological Properties: Preclinical Evidence
3.4. Clinical Studies
4. Labdane Diterpenes
4.1. Chemical Features
4.2. Natural Occurrence
4.3. Pharmacological Properties: Preclinical Studies
4.4. Clinical Studies
5. Further Research
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maggini, S.; Pierre, A.; Calder, P.C. Immune Function and Micronutrient Requirements Change over the Life Course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Moser, M.; Leo, O. Key concepts in immunology. Vaccine 2010, 28, C2–C13. [Google Scholar] [CrossRef]
- Kumar, D.; Romero, Y.; Schuck, K.N.; Smalley, H.; Subedi, B.; Fleming, S.D. Drivers and regulators of humoral innate immune responses to infection and cancer. Mol. Immunol. 2020, 121, 99–110. [Google Scholar] [CrossRef]
- Wen, C.C.; Chen, H.M.; Yang, N.S. Developing Phytocompounds from Medicinal Plants as Immunomodulators. Adv. Bot. Res. 2012, 62, 197–272. [Google Scholar] [PubMed]
- Durcan, L.; Petri, M. Immunomodulators in SLE: Clinical evidence and immunologic actions. J. Autoimmun. 2016, 74, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Mutgi, K.; Koo, J. Update on the role of systemic vitamin D in atopic dermatitis. Pediatr. Dermatol. 2013, 30, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Giamarellos-Bourboulis, E.J.; Domínguez-Andrés, J. Trained Immunity: A Tool for Reducing Susceptibility to and the Severity of SARS-CoV-2 Infection. Cell 2020, 181, 969–977. [Google Scholar] [CrossRef]
- Di Pasquale, A.; Preiss, S.; Tavares Da Silva, F.; Garçon, N. Vaccine Adjuvants: From 1920 to 2015 and beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef]
- Hui, D.S.; Lee, N.; Chan, P.K.; Beigel, J.H. The role of adjuvant immunomodulatory agents for treatment of severe influenza. Antiviral Res. 2018, 150, 202–216. [Google Scholar] [CrossRef]
- Temizoz, B.; Kuroda, E.; Ishii, K.J. Vaccine adjuvants as potential cancer immunotherapeutics. Int. Immunol. 2016, 28, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dranoff, G. Immunotherapy of Cancer. In Innate Immune Regulation and Cancer Immunotherapy; Wang, R., Ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Pawelec, G.; Gupta, S. Editorial: Immunology of Aging. Front. Immunol. 2019, 10, 1614. [Google Scholar] [CrossRef]
- Mohamed, S.I.A.; Jantan, I.; Haque, M.A. Naturally occurring immunomodulators with antitumor activity: An insight on their mechanisms of action. Int. Immunopharmacol. 2017, 50, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Michel, J.; Mezouar, S.; Sereme, Y.; Chanez, P.; Vitte, J.; Dubus, J.C.; Chantran, Y.; Leone, M.; Mège, J.L.; Fabre, A.; et al. Microbiome and the immune system: From a healthy steady-state to allergy associated disruption. Hum. Microbiome J. 2018, 10, 11–20. [Google Scholar]
- Mukherjee, P.K.; Nema, N.K.; Venkatesh, P.; Debnath, P.K. Changing scenario for promotion and development of Ayurveda—Way forward. J. Ethnopharmacol. 2012, 143, 424434. [Google Scholar] [CrossRef]
- Wang, C.; Hou, Y.; Lv, Y.; Chen, S.; Zhou, X.; Zhu, R.; Wang, J.; Jia, W.; Wang, X. Echinacea purpurea Extract Affects the Immune System, Global Metabolome, and Gut Microbiome in Wistar Rats. J. Agric. Sci. 2017, 9, 1. [Google Scholar] [CrossRef][Green Version]
- Sun, Y.; Chen, S.; Wei, R.; Xei, X.; Wang, C.; Fan, S.; Zhang, X.; Su, J.; Liu, J.; Jia, W.; et al. Metabolome and gut microbiota variation with long-term intake of Panax ginseng extracts on rats. Food Funct. 2018, 9, 3547–3556. [Google Scholar] [CrossRef]
- Chen, D.; Chen, G.; Ding, Y.; Wan, P.; Peng, Y.; Chen, C.; Ye, H.; Zeng, X.; Ran, L. Polysaccharides from the flowers of tea (Camellia sinensis L.) modulate gut health and ameliorate cyclophosphamide-induced immunosuppression. J. Funct. Food 2019, 61, 103470. [Google Scholar] [CrossRef]
- Stohs, S.J.; Bagchi, D. Antioxidant, Anti-inflammatory, and Chemoprotective Properties of Acacia catechu Heartwood Extracts. Phytother. Res. 2015, 29, 818–824. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wang, H.M.; Song, Y.L.; Nie, L.H.; Wang, L.F.; Liu, B.; Shen, P.P.; Liu, Y. Isolation, structure elucidation, antioxidative and immunomodulatory properties of two novel dihydrocoumarins from Aloe vera. Bioorg. Med. Chem. Lett. 2016, 16, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, Y.; Pi, F.; Cheng, Y.; Guo, Y.; Qian, H. Extraction, Purification, Structural Characteristics, Biological Activities and Pharmacological Applications of Acemannan, a Polysaccharide from Aloe vera: A Review. Molecules 2019, 24, 1554. [Google Scholar] [CrossRef] [PubMed]
- Pugh, N.; Ross, S.A.; El Sohly, M.A.; Pasco, D.S. Characterization of Aloeride, a new high-molecular-weight polysaccharide from Aloe vera with potent immunostimulatory activity. J. Agric. Food Chem. 2001, 49, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, J.; Dong, S.F.; Liu, C.H.; Italiani, P.; Sun, S.H.; Xu, J.; Boraschi, D.; Ma, S.P.; Qu, D. Immunomodulatory activity of andrographolide on macrophage activation and specific antibody response. Acta Pharmacol. Sin. 2010, 31, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Adorisio, S.; Fierabracci, A.; Muscari, I. Artocarpus tonkinensis Protects Mice Against Collagen-Induced Arthritis and Decreases Th17 Cell Function. Front. Pharmacol. 2019, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ren, W.; Zhang, L.; Zhang, Y.; Liu, D.; Liu, Y. A Review of the Pharmacological Action of Astragalus Polysaccharide. Front. Pharmacol. 2020, 11, 349. [Google Scholar] [CrossRef]
- Sharma, M.L.; Kaul, A.; Khajuria, A. Immunomodulatory activity of Boswellic acids (Pentacyclic Triterpene Acids) from Boswellia serrata. Phytother. Res 1996, 10, 107–112. [Google Scholar] [CrossRef]
- Rahayu, R.P.; Prasetyo, R.A.; Purwanto, D.A.; Kresnoadi, U.; Iskandar, R.P.D.; Rubianto, M. The immunomodulatory effect of green tea (Camellia sinensis) leaves extract on immunocompromised Wistar rats infected by Candida albicans. Vet. World 2018, 11, 765–770. [Google Scholar] [CrossRef]
- Gohil, K.J.; Patel, J.A.; Gajjar, A.K. Pharmacological Review on Centella asiatica: A Potential Herbal Cure-all. Ind. J. Pharm. Sci. 2010, 72, 546–556. [Google Scholar] [CrossRef]
- Mollazadeh, H.; Cicero, A.F.G.; Blesso, C.N.; Pirro, M.; Majeed, M.; Sahebkar, A. Immune modulation by curcumin: The role of interleukin-10. Crit. Rev. Food Sci. Nutr. 2019, 59, 89–101. [Google Scholar] [CrossRef]
- Guo, A.; He, D.; Xu, H.; Geng, C.; Zhao, J. Promotion of regulatory T cell induction by immunomodulatory herbal medicine licorice and its two constituents. Sci. Rep. 2016, 5, 14046. [Google Scholar] [CrossRef] [PubMed]
- Matyanga, C.M.J.; Morse, G.D.; Gundidza, M.; Nbachi, C.F.B. African potato (Hypoxis hemerocallidea): A systematic review of its chemistry, pharmacology and ethno medicinal properties. BMC Complement. Med. Ther. 2020, 20, 182. [Google Scholar] [CrossRef] [PubMed]
- Jeba, R.C.; Vaidyanathan, R.A.M.A.; Rameshkumar, G. Efficacy of Ocimum basilicum for immunomodulatory activity in Wistar albino rat. Int. J. Pharm. Pharm. Sci. 2011, 3, 199–203. [Google Scholar]
- Wagner, H.; Nörr, H.; Winterhoff, H. Plant daptogens. Phytomedicine 1994, 1, 63–76. [Google Scholar] [CrossRef]
- Riaz, M.; Rahman, N.U.; Zia-Ul-Haq, M.; Jaffar, H.Z.E.; Manea, R. Ginseng: A dietary supplement as immune-modulator in various diseases. Trends Food Sci. Tech. 2019, 83, 12–30. [Google Scholar] [CrossRef]
- Wael, S.; Watuguly, T.W.; Arini, I.; Smit, A.; Matdoan, N.; Prihati, D.R.; Sari, A.B.; Wahyudi, D.; Nuringtyas, T.R.; Wijayanti, N.; et al. Potential of Syzygium aromaticum (Clove) Leaf Extract on Immune Proliferation Response in Balb/c Mice Infected with Salmonella typhimurium. Clin. Med. Case Rep. 2018, 7, 613–627. [Google Scholar] [CrossRef]
- Haque, M.A.; Jantan, I.; Abbas Bukhari, S.N. Tinospora species: An overview of their modulating effects on the immune system. J. Ethnopharmacol. 2017, 207, 67–85. [Google Scholar] [CrossRef]
- Chandran, U.; Patwardhan, B. Network ethnopharmacological evaluation of the immunomodulatory activity of Withania somnifera. J. Ethnopharmacol. 2017, 197, 250–256. [Google Scholar] [CrossRef]
- Kalra, R.; Kaushik, N. Withania somnifera (Linn.) Dunal: A review of chemical and pharmacological diversity. Phytochem. Rev. 2017, 16, 953–987. [Google Scholar] [CrossRef]
- Ali, B.H.; Blunden, G.; Tanira, M.O.; Nemmar, A. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol. 2008, 46, 409–420. [Google Scholar] [CrossRef]
- Catanzaro, M.; Corsini, E.; Rosini, M.; Racchi, M.; Lanni, C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules 2018, 23, 2778. [Google Scholar] [CrossRef]
- Pillai, S.; Pillai, C.; Mitscher, L.A.; Cooper, R. Use of quantitative flow cytometry to measure ex vivo immunostimulant activity of Echinacea: The case for polysaccharides. J. Altern. Complement. Med. 2007, 13, 625–634. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Assessment Report on Echinacea purpurea (L.) Moench, Radix; EMA/HMPC/424584/2016; European Medicines Agency: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Kaur, P.; Makanjuola, V.O.; Arora, R.; Singh, B.; Arora, S. Immunopotentiating significance of conventionally used plant adaptogens as modulators in biochemical and molecular signalling pathways in cell mediated processes. Biomed. Pharmacother. 2017, 95, 1815–1829. [Google Scholar] [CrossRef] [PubMed]
- Bachiega, T.F.; Orsatti, C.L.; Pagliarona, A.C.; Missima, F.; Sousa, J.P.B.; Bastos, J.K.; Sforcin, J.M. Th1/Th2 Cytokine Production by Clove-Treated Mice. Nat. Prod. Res. 2009, 23, 1552–1558. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S. Azadirachta indica A. Juss. (Meliaceae). In Handbook of 200 Medicinal Plants; Akbar, S., Ed.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Cundell, R.D. Herbal Phytochemicals as Immunomodulators. Curr. Immunol. Rev. 2014, 10, 64–81. [Google Scholar] [CrossRef]
- Radzikowska, U.; Rinaldi, A.O.; Çelebi Sözener, Z.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef]
- Hoang, M.L.; Chen, C.H.; Chen, P.C.; Roberts, N.J.; Dickman, K.G.; Yun, B.H.; Turesky, R.J.; Pu, Y.S.; Vogelstein, B.; Papadopoulus, N.; et al. Aristolochic acid in the etiology of renal cell carcinoma. Cancer Epidemiol. Prev. Biomarkers 2016, 25, 1600–1608. [Google Scholar] [CrossRef]
- Jantan, I.; Ahmad, W.; Bukhari, S.N.A. Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Front. Plant Sci. 2015, 25, 655. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health Benefits and Molecular Mechanisms of Resveratrol: A Narrative Review. Foods 2020, 9, 340. [Google Scholar] [CrossRef]
- Kuban-Jankowska, A.; Sahu, K.K.; Gorska, M.; Tuszynski, J.A.; Wozniak, M. Chicoric acid binds to two sites and decreases the activity of the YopH bacterial virulence factor. Oncotarget 2016, 7, 2229–2238. [Google Scholar] [CrossRef]
- Tuyaerts, S.; Rombauts, K.; Everaert, T.; Van Nuffel, A.M.T.; Amant, F. A Phase 2 Study to Assess the Immunomodulatory Capacity of a Lecithin-based Delivery System of Curcumin in Endometrial Cancer. Front. Nutr. 2019, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the ‘fiber gap’. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure-function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef]
- Lee, J.S.; Kwon, J.S.; Yun, J.S.; Pahk, J.W.; Shin, W.C.; Lee, S.Y.; Hong, E.K. Structural characterization of immunostimulating polysaccharide from cultured mycelia of Cordyceps militaris. Carbohyd. Polym. 2010, 80, 1011–1017. [Google Scholar] [CrossRef]
- Simões, J.; Madureira, P.; Nunes, F.M.; Domingues, M.D.R.; Vilanova, M.; Coimbra, M.A. Immunostimulatory properties of coffee mannans. Mol. Nutr. Food Res. 2009, 53, 1036–1043. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, Z.-H.; Lee, C.G.; Synytsya, A.; Jo, H.S.; Kim, S.O.; Park, J.W.; Park, Y.I. Characterization and immunostimulating activity of a water-soluble polysaccharide isolated from Haematococcus lacustris. Biotechnol. Bioproc. E 2011, 16, 1090–1098. [Google Scholar] [CrossRef]
- Ohta, Y.; Lee, J.B.; Hayashi, K.; Fujita, A.; Park, D.K.; Hayashi, T. In vivo anti influenza virus activity of an immunomodulatory acidic polysaccharide isolated from Cordyceps militaris grown on germinated soybeans. J. Agric. Food Chem. 2007, 55, 10194–10199. [Google Scholar] [CrossRef]
- Xie, G.; Schepetkin, I.A.; Siemsen, D.W.; Kirpotina, L.N.; Wiley, J.A.; Quinn, M.T. Fractionation and characterization of biologically-active polysaccharides from Artemisia tripartita. Phytochemistry 2008, 69, 1359–1371. [Google Scholar] [CrossRef]
- Popov, S.V.; Ovodov, Y.S. Polypotency of the immunomodulatory effect of pectins. Biochemistry 2013, 78, 823–835. [Google Scholar] [CrossRef]
- Yin, J.Y.; Chan, B.C.L.; Yu, H.; Lau, I.Y.K.; Han, X.Q.; Cheng, S.W.; Wong, C.K.; Lau, C.B.S.; Xie, M.Y.; Fung, K.P.; et al. Separation, structure characterization, conformation and immunomodulating effect of a hyperbranched heteroglycan from Radix Astragali. Carbohydr. Polym. 2012, 87, 667–675. [Google Scholar] [CrossRef]
- Fang, X.; Chen, X. Structure elucidation and immunological activity of a novel pectic polysaccharide from the stems of Avicennia marina. Eur. Food Res. Technol. 2013, 236, 243–248. [Google Scholar] [CrossRef]
- Stephanie, B.; Eric, D.; Sophie, F.M.; Christian, B.; Yu, G. Carrageenan from Solieria chordalis (Gigartinales): Structural analysis and immunological activities of the low molecular weight fractions. Carbohydr. Polym. 2010, 81, 448–460. [Google Scholar] [CrossRef]
- Bhatia, S.; Rathee, P.; Sharma, K.; Chaugule, B.B.; Kar, N.; Bera, T. Immunomodulation effect of sulphated polysaccharide (porphyran) from Porphyra vietnamensis. Int. J. Biol. Macromol. 2013, 57, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Chlubnová, I.; Sylla, B.; Nugier-Chauvin, C.; Daniellou, R.; Legentil, L.; Kralová, B.; Ferrières, V. Natural glycans and glycoconjugates as immunomodulating agents. Nat. Prod. Rep. 2011, 28, 937–952. [Google Scholar] [CrossRef] [PubMed]
- Benkeblia, N. Fructooligosaccharides and fructans analysis in plants and food crops. J. Chromatogr. A 2013, 1313, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Dai, H.; Huang, L.; Gao, X.; Tsim, K.W.K.; Tu, P. A fructan, from Radix ophiopogonis, stimulates the proliferation of cultured lymphocytes: Structural and functional analyses. J. Nat. Prod. 2006, 69, 1257–1260. [Google Scholar] [CrossRef]
- Akhtar, M.; Tariq, A.F.; Awais, M.M.; Iqbal, Z.; Muhammad, F.; Shahid, M.; Hiszczynska-Sawicka, E. Studies on wheat bran Arabinoxylan for its immunostimulatory and protective effects against avian coccidiosis. Carbohydr. Polym. 2012, 90, 333–339. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, X.; Guo, Y.; Wang, Q.; Peng, D.; Cao, L. Comparison of the immunological activities of arabinoxylans from wheat bran with alkali and xylanase-aided extraction. Carbohydr. Polym. 2010, 81, 784–789. [Google Scholar] [CrossRef]
- Albuquerque, P.B.S.; de Oliveira, W.F.; Dos Santos Silva, P.M.; Dos Santos Correia, M.T.; Kennedy, J.F.; Coelho, L.C.B.B. Epiphanies of well-known and newly discovered macromolecular carbohydrates—A review. Int. J. Biol. Macromol. 2020, 156, 51–66. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Korolenko, T.A.; Bgatova, N.P.; Vetvicka, V. Glucan and Mannan-Two Peas in a Pod. Int. J. Mol. Sci. 2019, 20, 3189. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, M.; Luo, Y.; Wu, W. Isolation and structural characterization of an immunostimulating polysaccharide from fuzi, Aconitum carmichaeli. Carbohydr. Res. 2006, 341, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, P.M.; Prashanth, K.V.H.; Venkatesh, Y.P. Isolation, structural elucidation and immunomodulatory activity of fructans from aged garlic extract. Phytochemistry 2011, 72, 255–264. [Google Scholar] [CrossRef]
- Simoes, J.; Nunes, F.M.; Domingues, P.; Coimbra, M.A.; Domingues, M.R. Mass spectrometry characterization of an Aloe vera mannan presentin immunostimulatory activity. Carbohydr. Polym. 2012, 90, 229–236. [Google Scholar] [CrossRef]
- Mikkonen, K.S.; Tenkanen, M. Sustainable food packaging materials based on future biorefinery products: Xylans and mannans. Trends Food. Sci. Technol. 2012, 28, 90–102. [Google Scholar] [CrossRef]
- Moretão, M.P.; Zampronio, A.R.; Gorin, P.A.J.; Iacomini, M.; Oliveira, M.B.M. Induction of secretory and tumoricidal activities in peritoneal macrophages activated by an acidic heteropolysaccharide (ARAGAL) from the gum of Anadenanthera colubrina (Angico branco). Immunol. Lett. 2004, 93, 189–197. [Google Scholar] [CrossRef]
- Petkowicz, C.D.O.; Reicher, F.; Chanzy, H.; Taravel, F.R.; Vuong, R. Linear mannan in the endosperm of Schizolobium amazonicum. Carbohydr. Polym. 2001, 44, 107–112. [Google Scholar] [CrossRef]
- Wang, X.S.; Liu, L.; Fang, J.N. Immunological activities and structure of pectin from Centella asiatica. Carbohydr. Polym. 2005, 60, 95–101. [Google Scholar] [CrossRef]
- Sachslehner, A.; Foidl, G.; Foidl, N.; Gübitz, G.; Haltrich, D. Hydrolysis of isolated coffee mannan and coffee extract by mannanases of Sclerotium rolfsii. J. Biotechnol. 2000, 80, 127–134. [Google Scholar] [CrossRef]
- Mikkonen, K.S.; Tenkanen, M.; Cooke, P.; Xu, C.; Rita, H.; Willför, S.; Holmbom, B.; Hicks, K.B.; Yadav, M.P. Mannans as stabilizers of oil-in-water beverage emulsions. LWT Food Sci. Technol. 2009, 42, 849–855. [Google Scholar] [CrossRef]
- Zha, X.Q.; Luo, J.P.; Luo, S.Z.; Jiang, S.T. Structure identification of a new immunostimulating polysaccharide from the stems of Dendrobium huoshanense. Carbohydr. Polym. 2007, 69, 86–93. [Google Scholar] [CrossRef]
- Luettig, B.; Steinmuller, C.; Gifford, G.E.; Wagner, H.; Lohmann-Matthes, M.-L. Macrophage activation by the polysaccharide arabinogalactan isolated from plant cell cultures of Echinacea purpurea. J. Natl. Cancer Inst. 1989, 81, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Stuppner, H.; Schäfer, W.; Zenk, M. Immunologically active polysaccharides of Echinacea purpurea cell cultures. Phytochemistry 1988, 27, 119–126. [Google Scholar] [CrossRef]
- Holderness, J.; Schepetkin, I.A.; Freedman, B.; Kirpotina, L.N.; Quinn, M.T.; Hedges, J.F.; Jutila, M.A. Polysaccharides isolated from acaí fruit induce innate immune responses. PLoS ONE 2011, 6, e17301. [Google Scholar] [CrossRef] [PubMed]
- Skyberg, J.A.; Rollins, M.F.; Holderness, J.S.; Marlenee, N.L.; Schepetkin, I.A.; Goodyear, A.; Dow, S.W.; Jutila, M.A.; Pascual, D.W. Nasal acai polysaccharides potentiate innate immunity to protect against pulmonary Francisella tularensis and Burkholderia pseudomallei infections. PLoS Pathog. 2012, 8, e1002587. [Google Scholar] [CrossRef] [PubMed]
- Samuelsen, A.B.; Rieder, A.; Grimmer, S.; Michaelsen, T.E.; Knutsen, S.H. Immunomodulatory activity of dietary fiber: Arabinoxylan and mixed-linked Beta-glucan isolated from barley show modest activities in vitro. Int. J. Mol. Sci. 2011, 12, 570–587. [Google Scholar] [CrossRef]
- Zhao, G.H.; Kan, J.Q.; Li, Z.X.; Chen, Z.D. Characterization and immunostimulatory activity of an (1→6)- α-d-glucan from the root of Ipomoea batatas. Int. Immunopharmacol. 2005, 5, 1436–1445. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Faulkner, C.L.; Nelson-Overton, L.K.; Wiley, J.A.; Quinn, M.T. Macrophage immunomodulatory activity of polysaccharides isolated from Juniperus scopolorum. Int. Immunopharmacol. 2005, 5, 1783–1799. [Google Scholar] [CrossRef]
- Peng, X.M.; Huang, L.J.; Qi, C.H.; Zhang, Y.X.; Tian, G.Y. Studies on chemistry and immunomodulating mechanism of aglycoconjugate from Lycium barbarum L. Chin. J. Chem. 2001, 19, 1190–1197. [Google Scholar] [CrossRef]
- Sun, L.; Peng, X.X.; Sun, P.; Shi, J.H.; Yuan, X.W.; Zhu, J.J.; Tai, G.; Zhou, Y. Structural characterization and immunostimulatory activity of a novel linear α-(1→6)- d-glucan isolated from Panax ginseng C. A. Meyer. Glycoconj. J. 2012, 29, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Willför, S.; Sundberg, K.; Tenkanen, M.; Holmbom, B. Spruce-derived mannans—A potential raw material for hydrocolloids and novel advanced natural materials. Carbohydr. Polym. 2008, 72, 197–210. [Google Scholar] [CrossRef]
- Dourado, F.; Madureira, P.; Carvalho, V.; Coelho, R.; Coimbra, M.A.; Vilanova, M.; Mota, M.; Gama, F.M. Purification, structure and immunobiological activity of an arabinan-rich pectic polysaccharide from the cell walls of Prunus dulcis seeds. Carbohydr. Res. 2004, 339, 2555–2566. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ai, L.; Cao, N.; Ma, J.; Wu, Y.; Wu, J.; Sun, X. Physicochemical properties and structural characterization of a galactomannan from Sophora alopecuroides L. seeds. Carbohydr. Polym. 2016, 140, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Schepetkin, I.A.; Quinn, M.T. Immunomodulatory activity of acidic polysaccharides isolated from Tanacetum vulgare L. Int. Immunopharmacol. 2007, 7, 1639–1650. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.K.R.; Rodriguez, S.; Ramachandran, R.; Alamo, A.; Melnick, S.J.; Escalon, E.; Garcia, P.I., Jr.; Wnuk, S.F.; Ramachandran, C. Immune stimulating properties of a novel polysaccharide from the medicinal plant Tinospora cordifolia. Int. Immunopharmacol. 2004, 4, 1645–1659. [Google Scholar] [CrossRef]
- Ramesh, H.P.; Yamaki, K.; Tsushida, T. Effect of fenugreek (Trigonella foenum-graecum L.) galactomannan fractions on phagocytosis in rat macrophages and on proliferation and IgM secretion in HB4C5 cells. Carbohydr. Polym. 2002, 50, 79–83. [Google Scholar] [CrossRef]
- Yin, M.; Zhang, Y.; Li, H. Advances in Research on Immunoregulation of Macrophages by Plant Polysaccharides. Front. Immunol. 2019, 10, 145. [Google Scholar] [CrossRef]
- Yao, J.F.; Wang, Z.X.; Zhang, X.Y.; Zhang, R.F. Study on immunomodulatory effect of Astragalus Polysaccharide on mice peritoneal macrophage. J. Henan Univ. 2005, 24, 34–36. [Google Scholar]
- Lee, K.Y.; Jeon, Y.J. Macrophage activation by polysaccharide isolated from Astragalus membranaceus. Int. Immunopharm. 2005, 5, 1225–1233. [Google Scholar] [CrossRef]
- Eman, Z.; Engy, R.; Fatma, A.; Hebata, A.M.; Tarek, I. Effects of dietary Astragalus polysaccharides (APS) on growth performance, immunological parameters, digestive enzymes, and intestinal morphology of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2014, 38, 149–157. [Google Scholar]
- Qin, Q.; Niu, J.; Wang, Z.; Xu, W.; Qiao, Z.; Gu, Y. Astragalus membranaceus extract activates immune response in macrophages via heparanase. Molecules 2012, 17, 7232–7240. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.S.; Park, S.B.; Shin, K.S. Molecular mechanisms of immunomodulatory activity by polysaccharide isolated from the peels of Citrus unshiu. Int. J. Biol. Macromol. 2018, 112, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary fiber pectin directly blocks toll-like receptor 2-1 and prevents doxorubicin-induced ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Folkerts, J.; Folkerts, G.; Maurer, M.; Braber, S. Microbiota-dependent and -independent effects of dietary fibre on human health. Br. J. Pharmacol. 2020, 177, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Wu, Y.; Wang, B.; Chen, X.; Xu, X.; Chen, H.; Li, W.; Xu, X. Echinacea purpurea extracts promote murine dendritic cell maturation by activation of JNK, p38 MAPK and NF-κB pathways. Dev. Comp. Immunol. 2017, 73, 21–26. [Google Scholar] [CrossRef]
- Li, R.; Chen, W.; Wang, W.; Tian, W.; Zhang, X. Extraction, characterization of Astragalus polysaccharides and its immune modulating activities in rats with gastric cancer. Carbohydr. Polym. 2009, 78, 738–742. [Google Scholar] [CrossRef]
- Li, J.; Zhong, Y.; Li, H.; Zhang, N.; Ma, W.; Cheng, G.; Liu, F.; Liu, F.; Xu, J. Enhancement of Astragalus polysaccharide on the immune responses in pigs inoculated with foot-and-mouth disease virus vaccine. Int. J. Biol. Macromol. 2011, 49, 362–368. [Google Scholar] [CrossRef]
- Liu, W.H.; Zhang, J.Z.; Liu, S.J.; Qiu, H.N.; Guo, J.X.; Liu, B. Effect of radix astragali drug serum on NK cell activity and expression of KLRK1. Prog. Mod. Biomed. 2014, 14, 4637–4640. [Google Scholar]
- Cao, L.; Liu, X.; Qian, T.; Sun, G.; Guo, Y.; Chang, F.; Zhou, S.; Sun, X. Antitumor andimmunomodulatory activity of arabinoxylans: A major constituent of wheatbran. Int. J. Biol. Macromol. 2011, 48, 160–164. [Google Scholar] [CrossRef]
- Ghoneum, M.; Abedi, S. Enhancement of natural killer cell activity of agedmice by modified arabinoxylan rice bran (MGN-3/Biobran). J. Pharm. Pharmacol. 2004, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.M.; Xu, W.; Dai, H.; Tu, P.; Li, Z.; Gao, X.M. A study on the immune receptors for polysaccharides from the roots of Astragalus membranaceus, a Chinese medicinal herb. Biochem. Biophys. Res. Commun. 2004, 320, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Hu, Y.; Wang, D.; Liu, J.; Zhang, J.; Zhao, X.; Liu, X.; Liu, C.; Yuan, J.; Ruan, S. Effects of Astragalus polysaccharide liposome on lymphocyte proliferation in vitro and adjuvanticity in vivo. Carbohydr. Polym. 2012, 88, 68–74. [Google Scholar] [CrossRef]
- Hong, H.; Kim, J.; Lim, T.; Song, Y.; Cho, C.; Jang, M. Mixing ratio optimization for functional complex extracts of Rhodiola crenulata, Panax quinquefolius, and Astragalus membranaceus using mixture design and verification of immune functional efficacy in animal models. J. Funct. Foods 2018, 40, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Abuelsaad, A.S. Supplementation with Astragalus polysaccharides alters Aeromonas-induced tissue-specific cellular immune response. Microb. Pathog. 2014, 66, 48–56. [Google Scholar] [CrossRef]
- Novak, M.; Vetvicka, V. Beta-glucans, history, and the present: Immunomodulatory aspects and mechanisms of action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef]
- Aguilar, J.C.; Rodriguez, E.G. Vaccine adjuvants revisited. Vaccine 2007, 25, 3752–3762. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Saade, F.; Petrovsky, N. AdvaxTM, a polysaccharide adjuvant derived from delta inulin, provides improved influenza vaccine protection through broad-based enhancement of adaptive immune responses. Vaccine 2012, 30, 5373–5381. [Google Scholar] [CrossRef]
- Saade, F.; Honda-Okubo, Y.; Trec, S.; Petrovsky, N. A novel hepatitis B vaccine containing AdvaxTM, a polysaccharide adjuvant derived from delta inulin, induces robust humoral and cellular immunity with minimal reactogenicity in preclinical testing. Vaccine 2013, 31, 1999–2007. [Google Scholar] [CrossRef]
- Du, X.; Zhao, B.; Li, J.; Cao, X.; Diao, M.; Feng, H.; Chen, X.; Chen, Z.; Zeng, X. Astragalus polysaccharides enhance immune responses of HBV DNA vaccination via promoting the dendritic cell maturation and suppressing Treg frequency in mice. Int. Immunopharmacol. 2012, 14, 463–470. [Google Scholar] [CrossRef]
- Vassilaros, S.; Tsibanis, A.; Tsikkinis, A.; Pietersz, G.A.; McKenzie, I.F.; Apostolopoulos, V. Up to 15-year clinical follow-up of a pilot Phase III immunotherapy study in stage II breast cancer patients using oxidized mannan-MUC1. Immunotherapy 2013, 5, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Kjaerup, R.M.; Dalgaard, T.S.; Norup, L.R.; Bergman, I.M.; Sørensen, P.; Juul-Madsen, H.R. Adjuvant effects of mannose-binding lectin ligands on the immune response to infectious bronchitis vaccine in chickens with high or low serum mannose-binding lectin concentrations. Immunobiology 2014, 219, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Luu, M.; Visekruna, A. Short-chain fatty acids: Bacterial messengers modulating the immunometabolism of T cells. Eur. J. Immunol. 2019, 9, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; Raingeard de la Bletiere, D.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M.; Galmiche, J.P. Butyrate inhibits inflammatory responses through NFkappaB inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Shortchain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef]
- Jiang, C.M.; Zhang, M.; Sun, Z. Addition of astragalus into peritoneal dialysate improve peritoneal macrophages function in CAPD patients. J. Med. Postgrad. 2005, 18, 135–138. [Google Scholar]
- Ji, X.Q.; Du, J.F.; Chen, G. Comparison of perioperative immune function status in colorectal cancer patients and immune regulatory function of drugs. Pract. J. Cancer 2011, 26, 331–334. [Google Scholar]
- Shi, X.S.; Sun, Z.X. Influences of astragalus Injection on clinical efficacy and cellular immune function in patients of bronchial asthma with acute exacerbation. Chin. J. Exp. Trad. Med. Form. 2012, 18, 293–296. [Google Scholar]
- Schwarz, E.; Parlesak, A.; Henneicke-von Zepelin, H.H.; Bode, J.C.; Bode, C. Effect of oral administration of freshly pressed juice of Echinacea purpurea on the number of various subpopulations of B- and T-lymphocytes in healthy volunteers: Results of a double-blind, placebo-controlled cross-over study. Phytomedicine 2005, 12, 625–631. [Google Scholar] [CrossRef]
- Brinkeborn, R.M.; Shah, D.V.; Degenring, F.H. Echinaforce and other Echinacea fresh plant preparations in the treatment of the common cold. A randomized, placebo controlled, double-blind clinical trial. Phytomedicine 1999, 6, 1–6. [Google Scholar] [CrossRef]
- Karsch-Völk, M.; Barrett, B.; Kiefer, D.; Bauer, R.; Ardjomand-Woelkart, K.; Linde, K. Echinacea for preventing and treating the common cold. Cochrane Database Syst. Rev. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.; Svozil, V.; Kral, V.; Rajnohova Dobiasova, L.; Stiborova, I.; Vetvicka, V. Clinical trials of yeast-derived beta-(1,3) glucan in children: Effects on innate immunity. Ann. Transl. Med. 2014, 2, 15. [Google Scholar] [PubMed]
- Richter, J.; Svozil, V.; Kral, V.; Rajnohova Dobiasova, L.; Vetvicka, V. beta-glucan affects mucosal immunity in children with chronic respiratory problems under physical stress: Clinical trials. Ann. Transl. Med. 2015, 3, 52. [Google Scholar]
- Vetvicka, V.; Richter, J.; Svozil, V.; Rajnohova Dobiasova, L.; Kral, V. Placebo-driven clinical trials of yeast-derived beta-(1-3) glucan in children with chronic respiratory problems. Ann. Transl. Med. 2013, 1, 26. [Google Scholar]
- Varricchio, A.M.; Capasso, M.; Della Volpe, A.; Malafronte, L.; Mansi, N.; Varricchio, A.; Ciprandi, G. Resveratrol plus carboxymethyl-β-glucan in children with recurrent respiratory infections: A preliminary and real-life experience. Ital. J. Pediatr. 2014, 40, 93. [Google Scholar] [CrossRef]
- Baldassarre, M.E.; Di Mauro, A.; Labellarte, G.; Pignatelli, M.; Fanelli, M.; Schiavi, E.; Mastromarino, P.; Capozza, M.; Panza, R.; Laforgia, N. Resveratrol plus carboxymethyl-β-glucan in infants with common cold: A randomized double-blind trial. Heliyon 2020, 6, e03814. [Google Scholar] [CrossRef]
- Miraglia Del Giudice, M.; Maiello, N.; Capristo, C.; Alterio, E.; Capasso, M.; Perrone, L.; Ciprandi, G. Resveratrol plus carboxymethyl-β-glucan reduces nasal symptoms in children with pollen-induced allergic rhinitis. Curr. Med. Res. Opin. 2014, 30, 1931–1935. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, 9–14. [Google Scholar] [CrossRef]
- Chow, C. Fatty Acids in Foods and Their Health Implications, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–17. [Google Scholar]
- Feng, W.; Ao, H.; Peng, C. Gut Microbiota, Short-Chain Fatty Acids, and Herbal Medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Oteng, A.B.; Kersten, S. Mechanisms of Action of trans Fatty Acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C. Polyunsaturated fatty acid biosynthesis and metabolism in adult mammals. In Polyunsaturated Fatty Acid Metabolism, 1st ed.; Burdge, G.C., Ed.; Elsevier Inc.: New York, NY, USA, 2018; pp. 15–30. [Google Scholar]
- Hageman, J.H.; Danielsen, M.; Nieuwenhuizen, A.G.; Feitsma, A.L.; Dalsgaard, T.K. Comparison of bovine milk fat and vegetable fat for infant formula: Implications for infant health. Int. Dairy J. 2019, 92, 37–49. [Google Scholar] [CrossRef]
- Arkcoll, D. Laurie oil resources. Econ. Bot. 1988, 42, 195–205. [Google Scholar] [CrossRef]
- Fattore, E.; Fanelli, R. Palm oil and palmitic acid: A review on cardiovascular effects and carcinogenicity. Int. J. Food Sci. Nutr. 2013, 64, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Aruna, P.; Venkataramanamma, D.; Singh, A.K.; Singh, R.P. Health benefits of punicic acid: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 16–27. [Google Scholar] [CrossRef]
- Servili, M.; Sordini, B.; Esposto, S.; Urbani, S.; Veneziani, G.; Di Maio, I.; Selvaggini, R.; Taticchi, A. Biological Activities of Phenolic Compounds of Extra Virgin Olive Oil. Antioxidants 2013, 3, 1–23. [Google Scholar] [CrossRef]
- Choi, S.G.; Won, S.R.; Rhee, H.I. Oleic Acid and Inhibition of Glucosyltransferase. In Olives and Olive Oil in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2010; pp. 1375–1383. [Google Scholar]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Lachman, J.; Hejtmánková, A.; Taborsky, J.; Kotíková, Z.; Pivec, V.; Střalková, R.; Vollmannová, A.; Bojňanská, T.; Dědina, M. Evaluation of oil content and fatty acid composition in the seed of grapevine varieties. LWT Food Sci.Technol. 2015, 63, 620–625. [Google Scholar] [CrossRef]
- Nath, U.K.; Kim, H.-T.; Khatun, K.; Park, J.I.; Kang, K.K.; Nou, I.S. Modification of fatty acid profiles of rapeseed (Brassica napus L.) oil for using as food, industrial feed-stock and biodiesel. Plant Breed. Biotechnol. 2016, 4, 123–134. [Google Scholar] [CrossRef]
- Yoshime, L.T.; de Melo, I.L.P.; Sattler, J.A.G.; de Carvalho, E.B.T.; Mancini-Filho, J. Bitter gourd (Momordica charantia L.) seed oil as a naturally rich source of bioactive compounds for nutraceutical purposes. Nutrire 2016, 41, 12. [Google Scholar] [CrossRef]
- Parikh, M.; Netticadan, T.; Pierce, G.N. Flaxseed: Its bioactive components and their cardiovascular benefits. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H146–H159. [Google Scholar] [CrossRef] [PubMed]
- Asif, M. Health effects of omega-3,6,9 fatty acids: Perilla frutescens is a good example of plant oils. Orient. Pharm. Exp. Med. 2011, 11, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Guil-Guerrero, J.L.; Gómez-Mercado, F.; Ramos-Bueno, R.P.; González-Fernández, M.J.; Urrestarazu, M.; Jiménez-Becker, S.; de Bélair, G. Fatty acid profiles and sn -2 fatty acid distribution of γ-linolenic acid-rich Borago species. J. Food Compos. Anal. 2018, 66, 74–80. [Google Scholar] [CrossRef]
- Timoszuk, M.; Bielawska, K.; Skrzydlewska, E. Evening Primrose (Oenothera biennis) Biological Activity Dependent on Chemical Composition. Antioxidants 2018, 7, 108. [Google Scholar] [CrossRef]
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The Seed of Industrial Hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients 2020, 12, 1935. [Google Scholar] [CrossRef]
- Traitler, H.; Winter, H.; Richli, U.; Ingenbleek, Y. Characterization of gamma-linolenic acid in Ribes seed. Lipids 1984, 19, 923–928. [Google Scholar] [CrossRef]
- Kim, D.E.; Shang, X.; Assefa, A.D.; Keum, Y.S.; Saini, R.K. Metabolite profiling of green, green/red, and red lettuce cultivars: Variation in health beneficial compounds and antioxidant potential. Food Res. Int. 2018, 105, 361–370. [Google Scholar] [CrossRef]
- Yaqoob, P. Monounsaturated fatty acids and immune function. Eur. J. Clin. Nutr. 2002, 56, S9–S13. [Google Scholar] [CrossRef]
- Jeffery, N.M.; Cortina, M.; Newsholme, E.A.; Calder, P.C. Effects of variations in the proportions of saturated, monounsaturated and polyunsaturated fatty acids in the rat diet on spleen lymphocyte functions. Br. J. Nutr. 1997, 77, 805–823. [Google Scholar] [CrossRef]
- Mastrangelo, A.M.; Jeitner, T.M.; Eaton, J.W. Oleic acid increases cell surface expression and activity of CD11b on human neutrophils. J. Immunol. 1998, 161, 4268–4275. [Google Scholar]
- Ferrante, A.; Goh, D.; Harvey, D.P.; Robinson, B.S.; Hii, C.S.; Bates, E.J.; Hardy, S.J.; Johnson, D.W.; Poulos, A. Neutrophil migration inhibitory properties of polyunsaturated fatty acids. The role of fatty acid structure, metabolism, and possible second messenger systems. J. Clin. Investig. 1994, 93, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Llado, V.; Gutierrez, A.; Martinez, J.; Casas, J.; Teres, S.; Higuera, M.; Galmés, A.; Saus, C.; Besalduch, J.; Busquets, X.; et al. Minerval induces apoptosis in Jurkat and other cancer cells. J. Cell. Mol. Med. 2010, 14, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Cury-Boaventura, M.F.; Pompeia, C.; Curi, R. Comparative toxicity of oleic acid and linoleic acid on Jurkat cells. Clin. Nutr. 2004, 23, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Cury-Boaventura, M.F.; Pompeia, C.; Curi, R. Comparative toxicity of oleic acid and linoleic acid on Raji cells. Nutrition 2005, 21, 395–405. [Google Scholar] [CrossRef]
- Kim, D.H.; Cho, Y.M.; Lee, K.H.; Jeong, S.W.; Kwon, O.J. Oleate protects macrophages from palmitate-induced apoptosis through the downregulation of CD36 expression. Biochem. Biophys. Res. Commun. 2017, 488, 477–482. [Google Scholar] [CrossRef]
- Wu, H.; Weidinger, C.; Schmidt, F.; Keye, J.; Friedrich, M.; Yerinde, C.; Willimsky, G.; Qin, Z.; Siegmund, B.; Glauben, R. Oleate but not stearate induces the regulatory phenotype of myeloid suppressor cells. Sci. Rep. 2017, 7, 7498. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Hanzelmann, D.; Härtner, T.; Peschel, A.; Götz, F. Skin-Specific Unsaturated Fatty Acids Boost the Staphylococcus aureus Innate Immune Response. Infect. Immun. 2015, 84, 205–215. [Google Scholar] [CrossRef]
- Viladomiu, M.; Hontecillas, R.; Lu, P.; Bassaganya-Riera, J. Preventive and prophylactic mechanisms of action of pomegranate bioactive constituents. Evid. Based Complement. Alternat. Med. 2013, 2013, 789764. [Google Scholar] [CrossRef]
- Yuan, G.; Chen, X.; Li, D. Modulation of peroxisome proliferator-activated receptor gamma(PPAR γ) by conjugated fatty acid in obesity and inflammatory bowel disease. J. Agric. Food Chem. 2015, 25, 1883–1895. [Google Scholar] [CrossRef]
- Boussetta, T.; Raad, H.; Lettéron, P.; Gougerot-Pocidalo, M.A.; Marie, J.C.; Driss, F.; El-Benna, J. Punicic acid a conjugated linolenic acid inhibits TNFalpha-induced neutrophil hyperactivation and protects from experimental colon inflammation in rats. PLoS ONE 2009, 4, e6458. [Google Scholar] [CrossRef]
- Lorente-Cebrián, S.; Costa, A.G.V.; Navas-Carretero, S.; Zabala, M.; Martínez, J.A.; Moreno-Aliaga, M.J. Role of omega-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases: A review of the evidence. J. Physiol. Biochem. 2013, 69, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polyunsaturated fatty acids and inflammation. Biochem. Soc. Trans. 2005, 33, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, F.; Ortiz, M.; Valenzuela, R.; Videla, L.A. Long-chain polyunsaturated fatty acids regulation of PPARs, signaling: Relationship to tissue development and aging. Prostaglandins Leukot. Essent. Fat Acids. 2016, 114, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.G.; Contaifer, D.; Madurantakam, P.; Carbone, S.; Price, E.T.; Van Tassell, B.; Brophy, D.F.; Wijesinghe, D.S. Dietary Bioactive Fatty Acids as Modulators of Immune Function: Implications on Human Health. Nutrients 2019, 11, 2974. [Google Scholar] [CrossRef] [PubMed]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.R.; Severino, P.; Ferreira, C.S.; Zielinska, A.; Santini, A.; Souto, S.B.; Souto, E.B. Linseed Essential Oil—Source of Lipids as Active Ingredients for Pharmaceuticals and Nutraceuticals. Curr. Med. Chem. 2019, 26, 4537–4558. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Samani, M.; Bahmani, M.; Rafieian-Kopaei, M. The chemical composition, botanical characteristic and biological activities of Borago officinalis: A review. Asian Pac. J. Trop. Med. 2014, 7, S22–S28. [Google Scholar] [CrossRef]
- Kasote, D.M.; Zanwar, A.A.; Devkar, S.T.; Hegde, M.V.; Deshmukh, K.K. Immunomodulatory activity of ether insoluble phenolic components of n-butanol fraction (EPC-BF) of flaxseed in rat. Asian Pac. J. Trop. Biomed. 2012, 2, S623–S626. [Google Scholar] [CrossRef]
- Park, H.J.; Park, J.S.; Hayek, M.G.; Reinhart, G.A.; Chew, B.P. Dietary fish oil and flaxseed oil suppress inflammation and immunity in cats. Vet. Immunol. Immunopathol. 2011, 141, 301–306. [Google Scholar] [CrossRef]
- Che, L.; Zhou, Q.; Liu, Y.; Hu, L.; Peng, X.; Wu, C.; Zhang, R.; Tang, J.; Wu, F.; Fang, Z.; et al. Flaxseed oil supplementation improves intestinal function and immunity, associated with altered intestinal microbiome and fatty acid profile in pigs with intrauterine growth retardation. Food Funct. 2019, 10, 8149–8160. [Google Scholar] [CrossRef]
- Hamburger, M.; Riese, U.; Graf, H.; Melzig, M.F.; Ciesielski, S.; Baumann, D.; Dittmann, K.; Wegner, C. Constituents in evening primrose oil with radical scavenging, cyclooxygenase, and neutrophil elastase inhibitory activities. J. Agric. Food Chem. 2002, 50, 5533–5538. [Google Scholar] [CrossRef] [PubMed]
- De La Puerta Vazquez, R.; Martinez-Dominguez, E.; Sanchez Perona, J.; Ruiz-Gutierrez, V. Effects of different dietary oils on inflammatory mediator generation and fatty acid composition in rat neutrophils. Metabolism 2004, 53, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Inhibition of natural killer cell activity by dietary lipids. Immunol. Lett. 1994, 41, 241–247. [Google Scholar] [CrossRef]
- Dirks, J.; van Aswegen, C.H.; du Plessis, D.J. Cytokine levels affected by gamma-linolenic acid. Prostaglandins Leukot. Essent. Fatty Acids 1998, 59, 273–277. [Google Scholar] [CrossRef]
- Calder, P.C. The effects of fatty acids on lymphocyte functions. Braz. J. Med. Biol. Res. 1993, 26, 901–917. [Google Scholar] [PubMed]
- Harbige, L.S. Fatty acids, the immune response, and autoimmunity: A question of n-6 essentiality and the balance between n-6 and n-3. Lipids 2003, 38, 323–341. [Google Scholar] [CrossRef]
- Vangaveti, V.N.; Jansen, H.; Kennedy, R.L.; Malabu, U.H. Hydroxyoctadecadienoic acids: Oxidised derivatives of linoleic acid and their role in inflammation associated with metabolic syndrome and cancer. Eur. J. Pharmacol. 2016, 785, 70–76. [Google Scholar] [CrossRef]
- Shewale, S.V.; Boudyguina, E.; Zhu, X.; Shen, L.; Hutchins, P.M.; Barkley, R.M.; Murphy, R.C.; Parks, J.S. Botanical oils enriched in n-6 and n-3 FADS2 products are equally effective in preventing atherosclerosis and fatty liver. J. Lipid Res. 2015, 56, 1191–1205. [Google Scholar] [CrossRef]
- Proudman, S.M.; James, M.J.; Spargo, L.D.; Metcalf, R.G.; Sullivan, T.R.; Rischmueller, M.; Flabouris, K.; Wechalekar, M.D.; Lee, A.T.; Cleland, L.G. Fish oil in recent onset rheumatoid arthritis: A randomised, double-blind controlled trial within algorithm-based drug use. Ann. Rheum. Dis. 2015, 74, 89–95. [Google Scholar] [CrossRef]
- Vadell, A.K.E.; Bärebring, L.; Hulander, E.; Gjertsson, I.; Lindqvist, H.M.; Winkvist, A. Anti-inflammatory Diet In Rheumatoid Arthritis (ADIRA)—A randomized, controlled crossover trial indicating effects on disease activity. Am. J. Clin. Nutr. 2020, 111, 1203–1213. [Google Scholar] [CrossRef]
- Molfino, A.; Amabile, M.I.; Monti, M.; Muscaritoli, M. Omega-3 Polyunsaturated Fatty Acids in Critical Illness: Anti-Inflammatory, Proresolving, or Both? Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.L.; Park, Y. The association between n-3 polyunsaturated fatty acid levels in erythrocytes and the risk of rheumatoid arthritis in Korean women. Ann. Nutr. Metab. 2013, 63, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Sand, I.K. The role of diet in multiple sclerosis: Mechanistic connections and current evidence. Curr. Nutr. Rep. 2018, 7, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fatty Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Pischon, T.; Hankinson, S.E.; Hotamisligil, G.S.; Rifai, N.; Willett, W.C.; Rimm, E.B. Habitual dietary intake of n-3 and n-6 fatty acids in relation to inflammatory markers among US men and women. Circulation 2003, 108, 155–160. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K.; Demarco, A.C.; Raha, S.K.; Shay, J.; Garvey, M.; Horrobin, D.F. Effects of linoleic and gamma-linolenic acids (efamol evening primrose oil) on fatty acid-binding proteins of rat liver. Mol. Cell. Biochem. 1990, 98, 177–182. [Google Scholar] [CrossRef]
- Simon, D.; Eng, P.A.; Borelli, S.; Kägi, R.; Zimmermann, C.; Zahner, C.; Drewe, J.; Hess, L.; Ferrari, G.; Lautenschlager, S.; et al. Gamma-linolenic acid levels correlate with clinical efficacy of evening primrose oil in patients with atopic dermatitis. Adv. Ther. 2014, 31, 180–188. [Google Scholar] [CrossRef]
- Veale, D.J.; Torley, H.I.; Richards, I.M.; O’Dowd, A.; Fitzsimons, C.; Belch, J.J.; Sturrock, R.D. A double-blind placebo controlled trial of Efamol Marine on skin and joint symptoms of psoriatic arthritis. Br. J. Rheumatol. 1994, 33, 954–958. [Google Scholar] [CrossRef]
- Tomic-Smiljanic, M.; Vasiljevic, D.; Lucic-Tomic, A.; Andjelkovic, N.; Jakovljevic, V.; Bolovich, S.; Veselinovic, M. Influence of different supplementation on platelet aggregation in patients with rheumatoid arthritis. Clin. Rheumatol. 2019, 38, 2443–2450. [Google Scholar] [CrossRef]
- Cameron, M.; Gagnier, J.J.; Chrubasik, S. Herbal therapy for treating rheumatoid arthritis. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef]
- Bamford, J.T.; Ray, S.; Musekiwa, A.; van Gool, C.; Humphreys, R.; Ernst, E. Oral evening primrose oil and borage oil for eczema. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.H.; Hardy, G.; Alany, R.G. Borage oil in the treatment of atopic dermatitis. Nutrition 2010, 26, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Ursoniu, S.; Sahebkar, A.; Serban, M.C.; Pinzaru, I.; Dehelean, C.; Noveanu, L.; Rysz, J.; Banach, M. Lipid and Blood Pressure Meta-Analysis Collaboration (LBPMC) Group. A systematic review and meta-analysis of clinical trials investigating the effects of flaxseed supplementation on plasma C-reactive protein concentrations. Arch. Med. Sci. 2019, 15, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Reuben, J.P. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar]
- Dai, Y.; Chen, S.R.; Chai, L.; Zhao, J.; Wang, Y.; Wang, Y. Overview of pharmacological activities of Andrographis paniculata and its major compound andrographolide. Crit. Rev. Food Sci. Nutr. 2018, 59, S17–S29. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, J.K.; Saikia, D.; Shanker, K.; Thakur, J.P.; Negi, A.S.; Banerjee, S. A bioactive labdane diterpenoid from Curcuma amada and its semisynthetic analogues as antitubercular agents. Eur. J. Med. Chem. 2010, 45, 4379–4382. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Q.; Peng, W.; Zhan, R.; Chen, Y. A new 12,17-cyclo-labdane diterpenoid from the twigs of Dacrycarpus imbricatus. Nat Prod Res. 2018, 32, 1669–1675. [Google Scholar] [CrossRef]
- Kobayashi, J.; Sekiguchi, M.; Shigemori, H.; Ohsaki, A. Chapecoderins A-C, new labdane-derived diterpenoids from Echinodorus macrophyllus. J. Nat. Prod. 2000, 63, 375–377. [Google Scholar] [CrossRef]
- Qiao, Y.; Khutsishvili, M.; Alizade, V.; Atha, D.; Borris, R.P. Labdane and Abietane Diterpenoids from Juniperus oblonga and Their Cytotoxic Activity. Molecules 2019, 24, 1561. [Google Scholar] [CrossRef]
- Zhang, R.-H.; Liu, Z.-K.; Yang, D.-S.; Zhang, X.-J.; Sun, H.-D.; Xiao, W.-L. Phytochemistry and pharmacology of the genus Leonurus: The herb to benefit the mothers and more. Phytochemistry 2018, 147, 167–183. [Google Scholar] [CrossRef]
- Tran, Q.T.N.; Wong, W.S.F.; Chai, C.L.L. Labdane diterpenoids as potential anti-inflammatory agents. Pharmacol. Res. 2017, 124, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.K.; Thoa, N.T.K.; Linh, T.M.; Trang, D.T.; Van Kiem, P.; Nhiem, N.X.; Tai, B.H.; Van Minh, C.; Song, J.-H.; Ko, H.-J.; et al. Labdane-type diterpenoids from Vitex limonifolia and their antivirus activities. J. Nat. Med. 2018, 72, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Y.; Wu, R.H.; Logue, M.; Blondel, C.; Lai, L.; Stuart, B.; Flower, A.; Fei, Y.T.; Moore, M.; Shepherd, J.; et al. Andrographis paniculata (Chuān Xīn Lián) for symptomatic relief of acute respiratory tract infections in adults and children: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0181780. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Islam, M.A.; Shaw, S.; Khan, I.N.; Saravi, S.; Ahmad, S.; Rehman, S.; Gupta, V.K.; et al. Andrographolide, a diterpene lactone from Andrographis paniculata and its therapeutic promises in cancer. Cancer Lett. 2018, 420, 129–145. [Google Scholar] [CrossRef]
- Vaz, M.S.M.; Da Silva, M.S.V.; Oliveira, R.J.; Mota, J.D.S.; Brait, D.R.H.; De Carvalho, L.N.B.; Vani, J.M.; Berno, C.R.; Araújo, F.H.S.; De Barros, M.E. Evaluation of the toxicokinetics and apoptotic potential of ethanol extract from Echinodorus macrophyllus leaves in vivo. Regul. Toxicol. Pharmacol. 2016, 82, 32–38. [Google Scholar] [CrossRef]
- Da Silva, G.P.; Fernandes, D.C.; Vigliano, M.V.; da Fonseca, E.N.; Santos, S.V.; Marques, P.R.; Justo, M.D.; Sabino, K.C.; Coelho, M.G. Flavonoid-enriched fraction from Echinodorus macrophyllus aqueous extract exhibits high in-vitro and in-vivo anti-inflammatory activity. J. Pharm. Pharmacol. 2016, 68, 1584–1596. [Google Scholar] [CrossRef]
- Pinto, A.C.; Rego, G.C.G.; Siqueira, A.M.; Cardoso, C.C.; Reis, P.A.; Marques, E.A.; Coelho, M.G.; Sabino, K.G.C. Immunosuppressive effects of Echinodorus macrophyllus aqueous extract. J. Ethnopharmacol. 2007, 111, 435–439. [Google Scholar] [CrossRef]
- Heinrich, M.; Appendino, G.; Efferth, T.; Fürst, R.; Izzo, A.A.; Kayser, O.; Pezzuto, J.M.; Viljoen, A. Best practice in research—Overcoming common challenges in phytopharmacological research. J. Ethnopharmacol. 2020, 246, 112230. [Google Scholar] [CrossRef]
- Izzo, A.A.; Teixeira, M.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Insel, P.A.; Ji, Y.; Kendall, D.A.; Panattieri, R.A.; et al. A practical guide for transparent reporting of research on natural products in the British Journal of Pharmacology: Reproducibility of natural product research. Br. J. Pharmacol. 2020, 177, 2169–2178. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Int. J. Surg. 2010, 8, 336–341. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medicinal Plants | Phytochemicals | Effect on Immune Function/Type of Immunomodulation | References |
|---|---|---|---|
| Acacia catechu Willd. heartwood | Flavonoids, phenolic acids, catechins | Antinflammatory activity/Immunoadjuvant | [21] |
| Aloe vera (L.) Burm.f. | Acemannan, dihydrocoumarins | Adaptive immunity activation/Immunostimulant | [22,23,24] |
| Andrographis paniculata (Burm.f.) Nees | Diterpene lactones (andrographolide) | Modulation of innate and adaptive immunity/Immunosuppressor | [25] |
| Artocarpus tonkinensis A. Chev. Ex Gagnep | Auronol glycosides (maesopsin 4-O-glucoside and alphitonin-4-O-glucoside) | Inhibition of humoral and cellular adaptive immunity/Immunosuppressor | [26] |
| Astragalus membranaceous (Fisch.) Bge. | Polysaccharides | Activation of cellular immunity/Immunostimulant | [27] |
| Boswellia serrata Roxb. ex Colebr | Boswellic acid | Anti-anaphylactic and mast cell stabilization/Immunosuppressor | [28] |
| Camellia sinensis (L.) Kuntze | Polysaccharides | Activation of immunoreactivity through the modulation of gut microbiome/Immunostimulant effect | [20,29] |
| Centella asiatica (L.) Urban | Madecassoside | Regulation the abnormal humoral and cellular immunity/Immunosuppressor | [30] |
| Curcuma longa L. | Curcuminoids | IL-10-mediated anti-inflammatory and immunosuppressive activity/Immunosuppressor | [31] |
| Echinacea purpurea (L.) Moench | Alkylamides, glycoproteins, polysaccharides | Activation of cellular immunity and modulation of gut microbiome | [18] |
| Glycyrrhiza glabra L. | Triterpene saponins (glycyrrhizin) | Enhanced cellular immunity and antinflammatory activity/Immunostimulant | [32] |
| Hypoxis rooperi T. Moore | Phenolic glucosides (hypoxoside) | Antinflammatory properties/Immunoadjuvant | [33] |
| Ocimum sanctum L. | Monoterpenes (eugenol and methyleugenol), sesquiterpenes (β-caryophyllene) | Activation of innate and adaptive immunity/Immunostimulant | [34] |
| Panax ginseng C.A. Meyer | Triterpene saponins (ginsenosides) | Adaptogen effects, stimulation of immune systems via cytokine activation, modulation of gut microbiome | [19,35,36] |
| Syzigium aromaticum (L.) Merr. and L.M. Perry | Monoterpenes (eugenol), sesquiterpenes (β-caryophyllene) | Activation of adaptive humoral immunity/Immunostimulant | [37] |
| Tinospora cordifolia (Thunb.) Miers | Arabinogalactan | Activation of adaptive immunity/Immunostimulant | [38] |
| Withania somnifera (L.) Dunal | Steroidal lactones (withaferin A) | Adaptogen effects, stimulation of adaptive humoral and cellular immunity/Immunostimulant | [39,40] |
| Zingiber officianalis Roscoe | Phenolics (gingerols) | Activation of adaptive humoral immunity/Immunostimulant | [41] |
| Plants | Family | Plant Part of Biological Interest | References |
|---|---|---|---|
| Aconitum carmichaeli Debx. | Ranunculaceae | Roots | [76] |
| Allium sativum L. | Liliaceae | Bulb | [77] |
| Aloe vera L. | Liliaceae | Leaves | [78] |
| Amorphallus konjac Koch | Araceae | Tubers | [79] |
| Anadenanthera colubrina (Vell.) Brenan | Fabaceae | Gum | [80] |
| Astragalus membranaceus (Fisch.) Bge. | Fabacee | Root | [64] |
| Carum carvi L. | Apiaceae | Seeds | [81] |
| Centella asiatica L. Urb. | Apiaceae | Aerial parts | [82] |
| Coffea arabica L. and C. robusta L. | Rubiaceae | Beans | [83] |
| Cyamopsis tetragonolobus L. | Fabaceae | Seeds | [84] |
| Dendrobium huoshanense C.Z. Tang et S.J. Cheng | Orchidaceae | Stems | [85] |
| Echinacea purpurea L. (Moench) | Asteracee | Aerial parts | [86,87] |
| Euterpe oleracea Mart. | Arecaceae | Fruits | [88,89] |
| Hordeum vulgare var. Tyra | Poaceae | Stems | [90] |
| Ipomoea batatas L. | Convolvulaceae | Roots | [91] |
| Juniperus scopolorum Sarg. | Cupressaceae | Cones | [92] |
| Lycium barbarian L. | Solanaceae | Fruit | [93] |
| Panax ginseng C.A. Meyer | Araliaceae | Roots | [94] |
| Picea abies L. | Pinaceae | Softwoods | [95] |
| Prunus dulcis (Miller) D. A. Webb. | Rosacee | Seeds | [96] |
| Sophora alopecuroides L. | Fabaceae | Seeds | [97] |
| Tanacetum vulgare L. | Asteraceae | Florets | [98] |
| Tinospora cordifolia (Thunb.) Miers | Menispermaceae | Aerial parts | [99] |
| Trigonella foenum-graecum L. | Fabaceae | Seeds | [100] |
| Fatty Acid | Class of Long-Chain Fatty Acid | IUPAC Name | Abbreviation | |
|---|---|---|---|---|
| Delta (∆) Nomenclature | n- or ω-Nomenclature | |||
| Palmitic acid | SFA | Hexadecanoic acid | 16:0 | 16:0 |
| Stearic acid | SFA | Octadecanoic acid | 18:0 | 18:0 |
| Oleic acid | MUFA | (9E)-Octadec-9-enoic acid | 18:1∆9 | 18:1n-9 or 18:1ω-9 |
| Punicic acid | Conjugated PUFA | (9Z,11E,13Z)-9,11,13-Octadecatrienoic acid | 18:3∆9,9,11,13 | 18:3n-5 or 18:3 ω-5 |
| α-Linolenic acid (ALA) | PUFA | (9Z,12Z,15Z)-Octadeca-9,12,15-trienoic acid | 18:3∆9,12,15 | 18:3n-3 or 18:3ω-3 |
| Stearidonic acid (SA) | PUFA | (6Z,9Z,12Z,15Z)-Octadeca-6,9,12,15-tetraenoic acid | 18:4∆6,9,12,15 | 18:4n-3 or 18:4ω-3 |
| Eicosatetraenoic acid (ETA) | PUFA | (2Z,4Z,5Z,8Z)-2,4,6,8-Icosatetraenoic acid | 20:4∆2,4,6,8 | 20:4n-3 or 20:4ω-3 |
| Eicosapentaenoic acid (EPA) | PUFA | (2E,4E,6E,8E,10E)-2,4,6,8,10-Icosapentaenoic acid | 20:5∆2,4,6,8,10 | 20:5n-3 or 20:5ω-3 |
| Docosahexaenoic acid (DHA) | PUFA | (4Z,7Z,10Z,13Z,16Z,19Z)-Docosa-4,7,10,13,16,19-hexaenoic acid | 22:6∆4,7,10,13,16,19 | 22:6n-3 or 20:6ω-3 |
| Linoleic acid (LA) | PUFA | (9Z,12Z)-Octadeca-9,12-dienoic acid | 18:2∆9,12 | 18:2n-6 or 18:2ω-6 |
| γ-Linolenic acid (GLA) | PUFA | (6Z,9Z,12Z)-Octadeca-6,9,12-trienoic acid | 18:3∆6,9,12 | 18:3n-6 or 18:3ω-6 |
| Dihomo γ-Linolenic acid (DGLA) | PUFA | (8Z,11Z,14Z)-Icosa-8,11,14-trienoic acid | 20:3∆8,11,14 | 20:3n-6 or 20:3ω-6 |
| Arachidonic acid (AA) | PUFA | (5E,8E,11E,14E)-Icosa-5,8,11,14-tetraenoic acid | 20:4∆5,8,11,14 | 20:4n-6 or 20:4ω-6 |
| Docosapentaenoic acid (DPA) | PUFA | (7Z,10Z,13Z,16Z,19Z)-Docosa-7,10,13,16,19-pentaenoic acid | 22:5∆7,10,13,16,19 | 22:5n-6 or 22:5ω-6 |
| Species (Byproduct) | Amount (%) | References | |
|---|---|---|---|
| MUFA | |||
| Oleic acid | Olea europea L. (fruit oil) | 56–84% a | [151] |
| Vitis vinifera L. (grapeseed oil) | 9–17% | [154] | |
| Brassica spp. (rapeseed oil) | 60–80% | [155] | |
| Conjugated PUFA | |||
| Punicic acid | Punica granatum L. (pomegranate seed oil) | 70–85% | [150] |
| Momordica charantia L. (bitter gourds seed oils) | 1.5–16% | [156] | |
| Essential ω3-PUFA | |||
| α-Linolenic acid (ALA) | Linum usitatissimum L. (flaxseed oil) | 55–57% | [153,157] |
| Salvia hispanica L. (chia seeds) | 17.8% | [153] | |
| Perilla frutescens var. frutescens L. (seeds) | 54–64% | [158] | |
| EPA, DHA | Fish oils | 4–13% | [153] |
| Essential ω6-PUFA | |||
| Linoleic acid (LA) | Borago sp. (seed oil) | 26.8–37.9% | [159] |
| Oenothera biennis L. (seed oil) | 70–74% | [160] | |
| Cannabis sativa L. (hamp seed oil) | 27–58% | [153,161] | |
| Glycine max (L.) Merr. (soybean) | 50.4% | [153,161] | |
| Bertholletia excelsa (Dried brazilnuts) | 23.8% | [153,161] | |
| Juglans regia L. (dried walnuts) | 33.8% | [153,161] | |
| Zea mays L. (corn) | 53.2% | [153,161] | |
| γ-Linolenic acid (GLA) | Borago sp. (seed oil) | 9.6–39.8% | [159] |
| Oenothera biennis L. (seed oil) | 8–10% | [160] | |
| Ribes nigrum | 15–19% a | [162] | |
| (seed oil) | |||
| Echium spp. (seed oil) | 31.2–47.1% | [159] | |
| Ranunculus spp. (seed oil) | 37.9–39.7% | [159] |
| Plants | Family | Part of the Plant of Biological Interest | References |
|---|---|---|---|
| Andrographis paniculata (Burm.f.) Nees | Acanthaceae | Aerial parts, roots | [211,219] |
| Curcuma amada Roxb | Zingiberaceae | Rizome | [212] |
| Dacricarpus imbricatus (Blume) de Laub | Podocarpaceae | Bark | [213] |
| Echinodorus macrophyllus (Kunth) Micheli | Alismataceae | Leaves | [214] |
| Juniperus oblonga M. Bieb | Cupressaceae | Berries | [215] |
| Leonurus japonicas Houtt | Lamiaceae | Aerial parts | [216] |
| Marrubium L. Spp. | Lamiaceae | Aerial parts | [217] |
| Vitex limonifolia Wall. ex C.B.Clarke | Lamiaceae | Leaves | [218] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Sotto, A.; Vitalone, A.; Di Giacomo, S. Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines 2020, 8, 468. https://doi.org/10.3390/vaccines8030468
Di Sotto A, Vitalone A, Di Giacomo S. Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines. 2020; 8(3):468. https://doi.org/10.3390/vaccines8030468
Chicago/Turabian StyleDi Sotto, Antonella, Annabella Vitalone, and Silvia Di Giacomo. 2020. "Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview" Vaccines 8, no. 3: 468. https://doi.org/10.3390/vaccines8030468
APA StyleDi Sotto, A., Vitalone, A., & Di Giacomo, S. (2020). Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines, 8(3), 468. https://doi.org/10.3390/vaccines8030468