
Vaccine Based on Recombinant Fusion Protein Combining Hepatitis B Virus PreS with SARS-CoV-2 Wild-Type- and Omicron-Derived Receptor Binding Domain Strongly Induces Omicron-Neutralizing Antibodies in a Murine Model

 , , , , , , , ,
, , , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Expression and Purification of Recombinant Proteins
2.2. Matrix-Assisted Laser Desorption and Ionization Time-of-Flight Mass Spectrometry
2.3. Thermal Denaturation, Renaturation and Determination of the Melting Temperature via Circular Dichroism (CD)
2.4. Immunization of Mice, Blood Sampling and Manipulations
2.5. Measurements of Specific Antibodies
2.6. Virus Neutralization Assays
2.7. Measurement of Specific Cytokine Production
2.8. Histological Examination
2.9. Statistics
3. Results
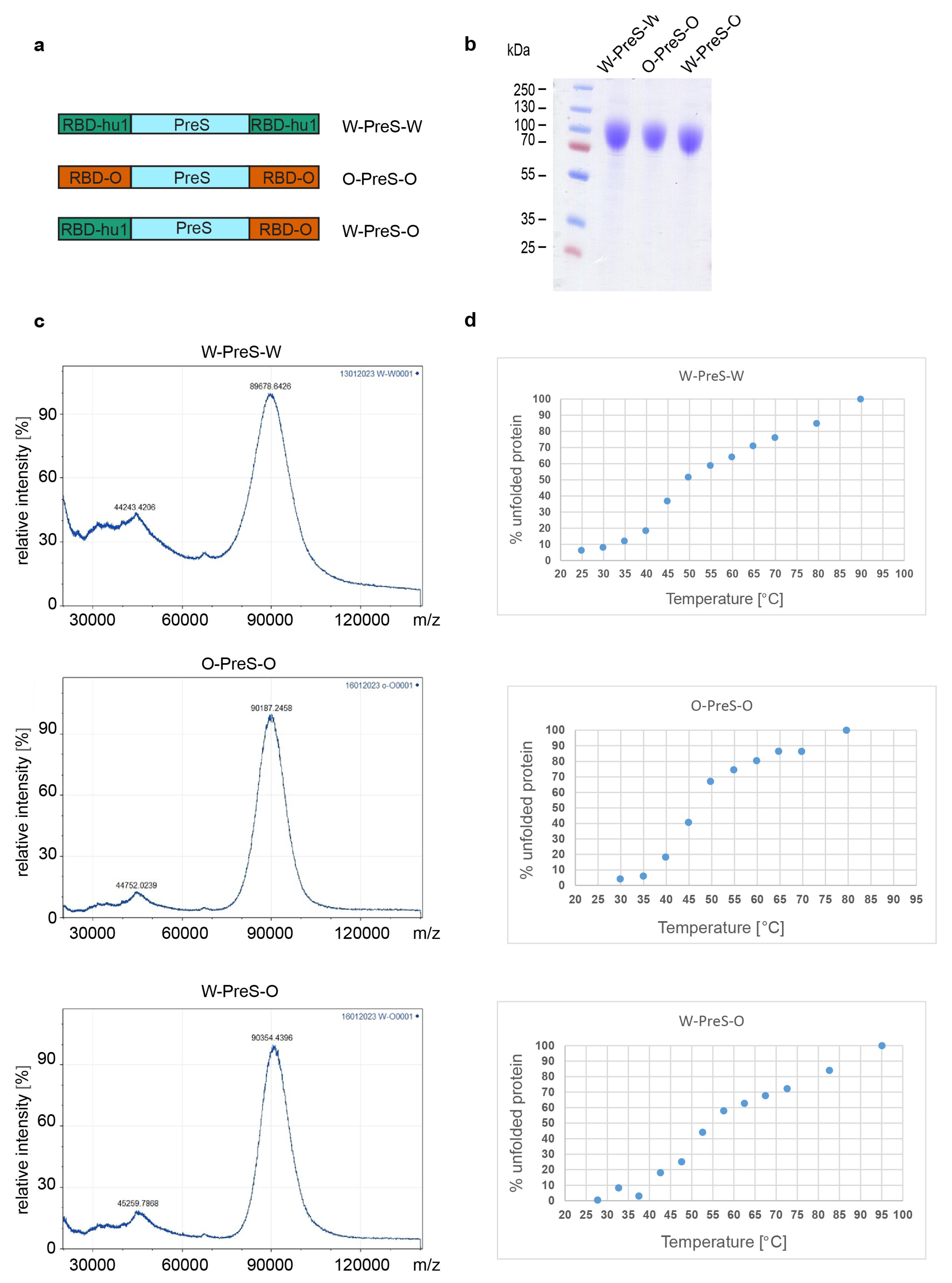
3.1. Recombinant Vaccine Antigens Represent Defined, Folded and Stable Proteins
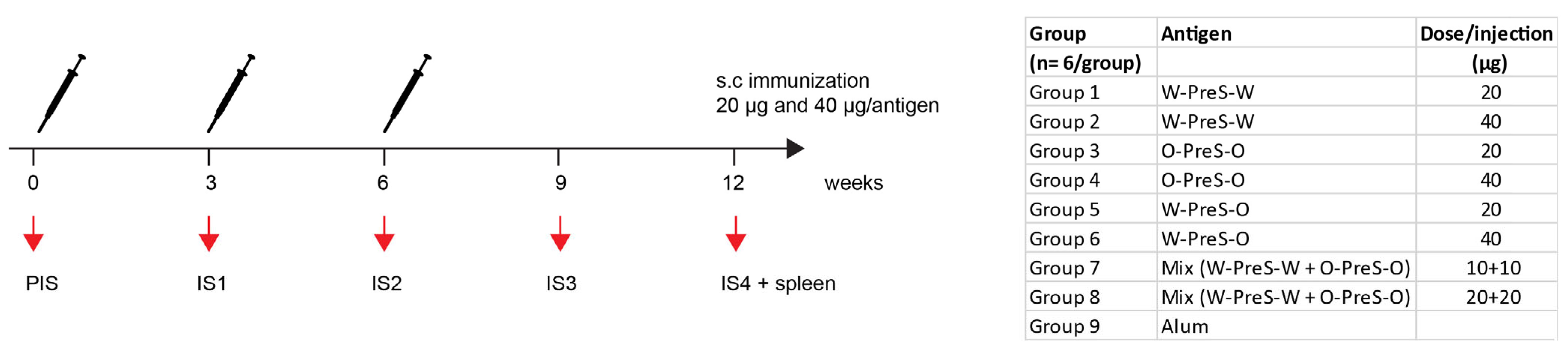
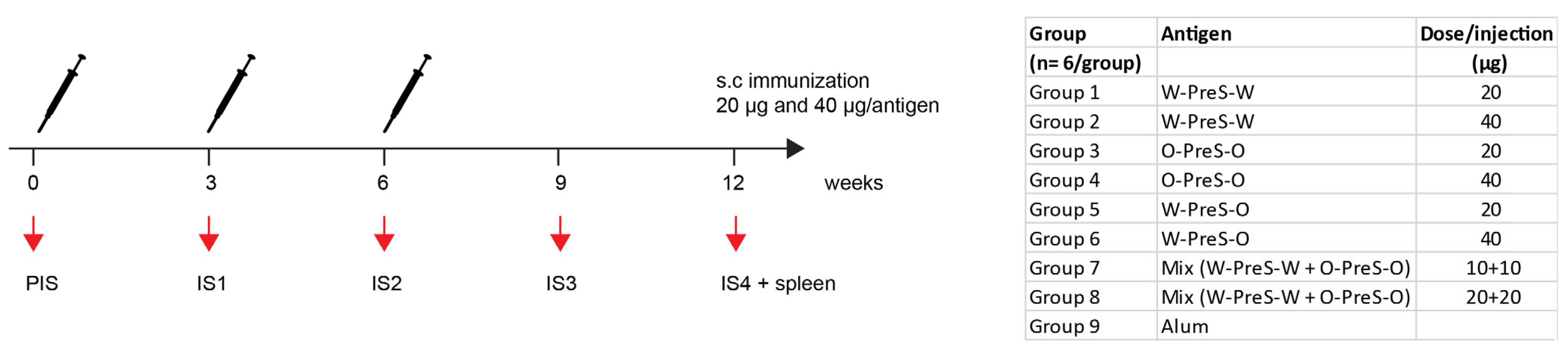
3.2. Formulation of the Subunit Vaccines and Immunization Schedule
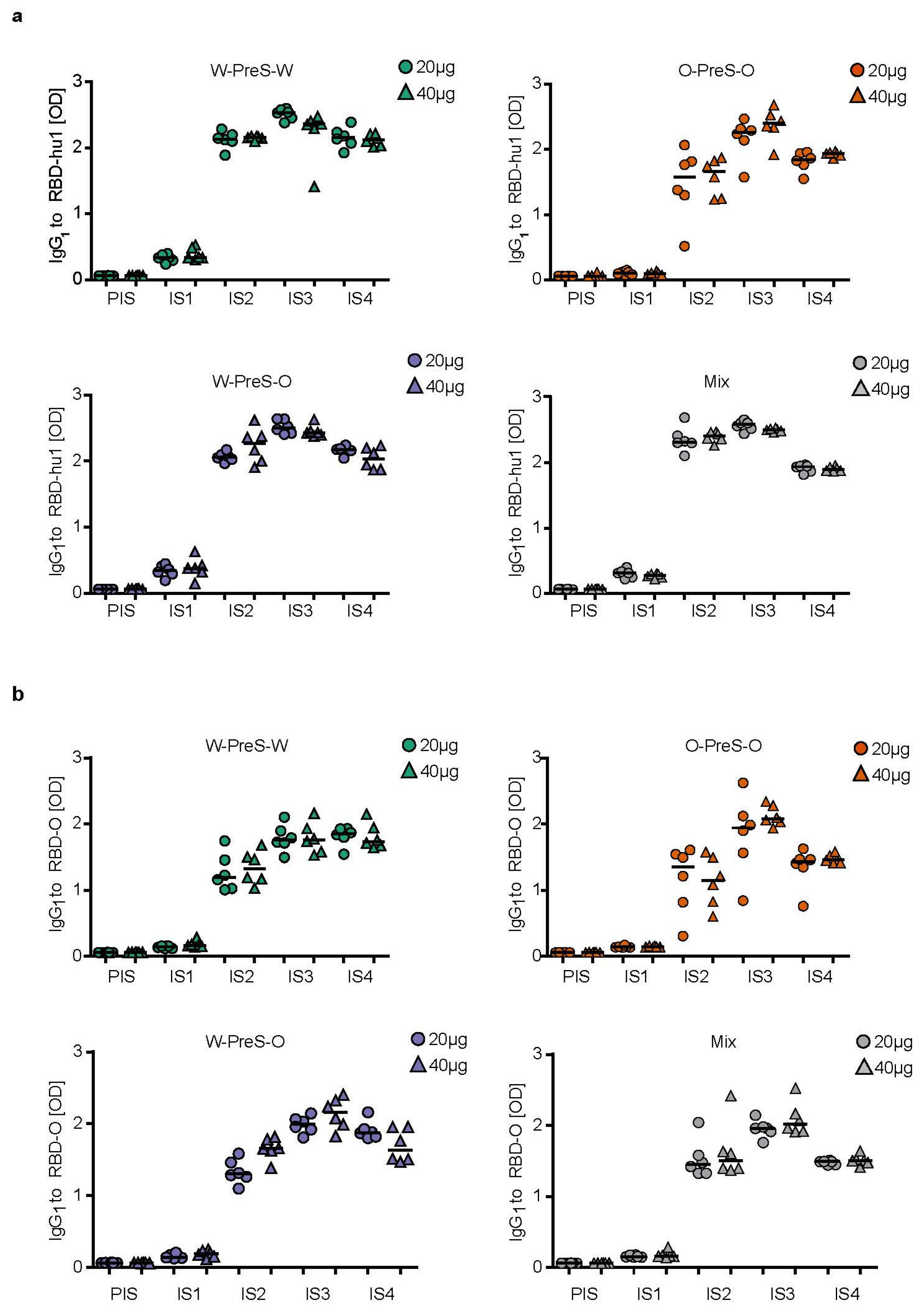
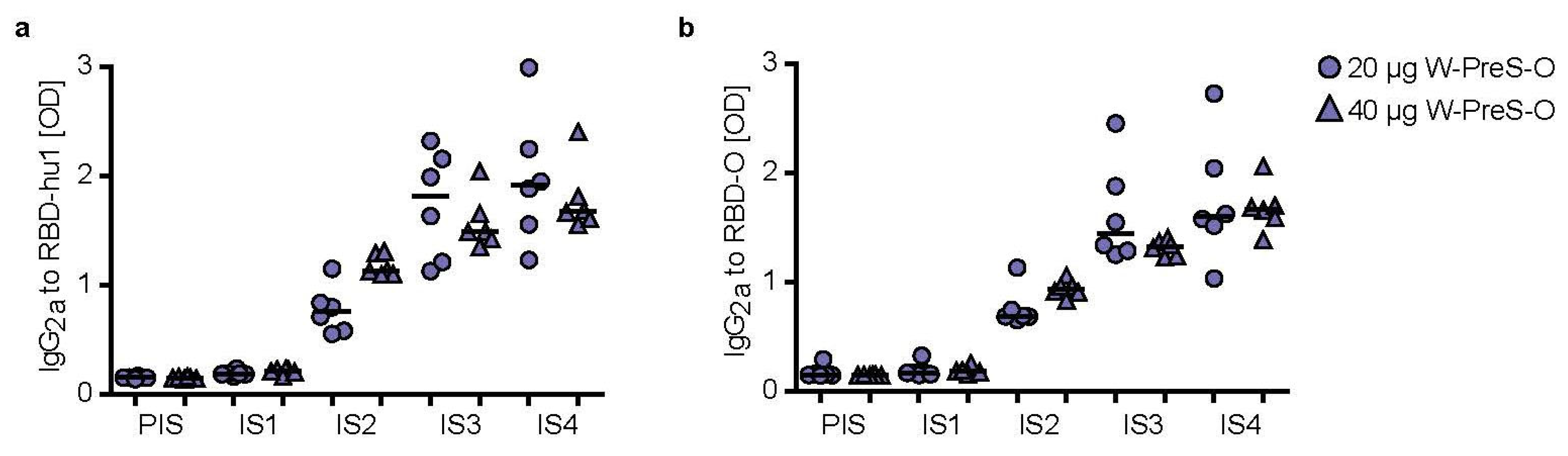
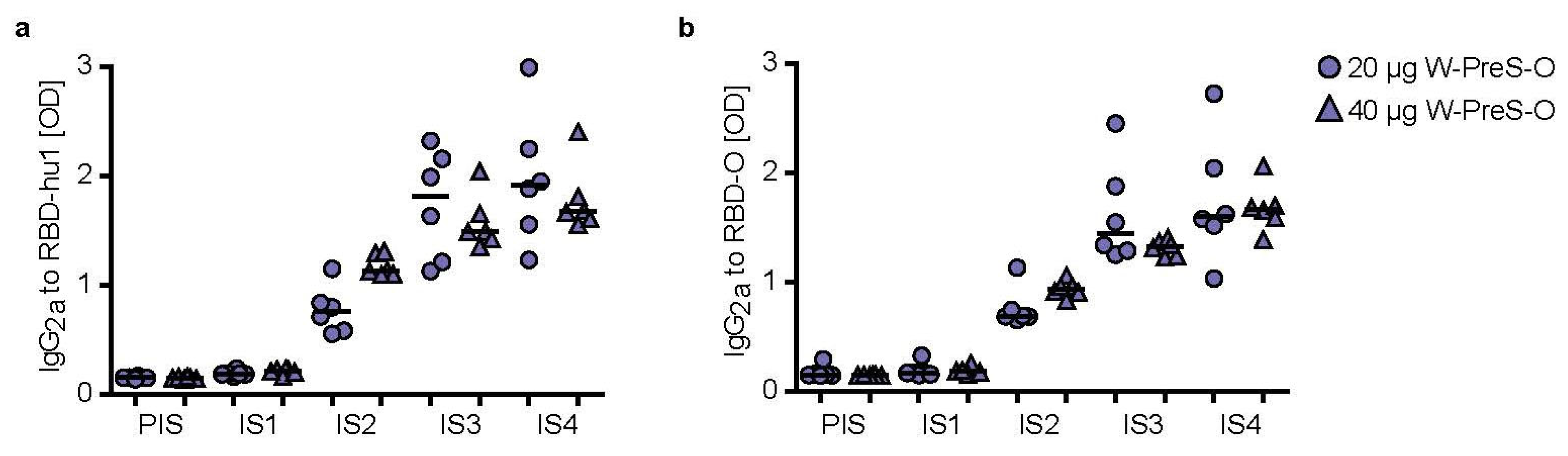
3.3. Immunization with W-PreS-O Induces High IgG Antibody Levels Specific for Wild-Type and Omicron RBD
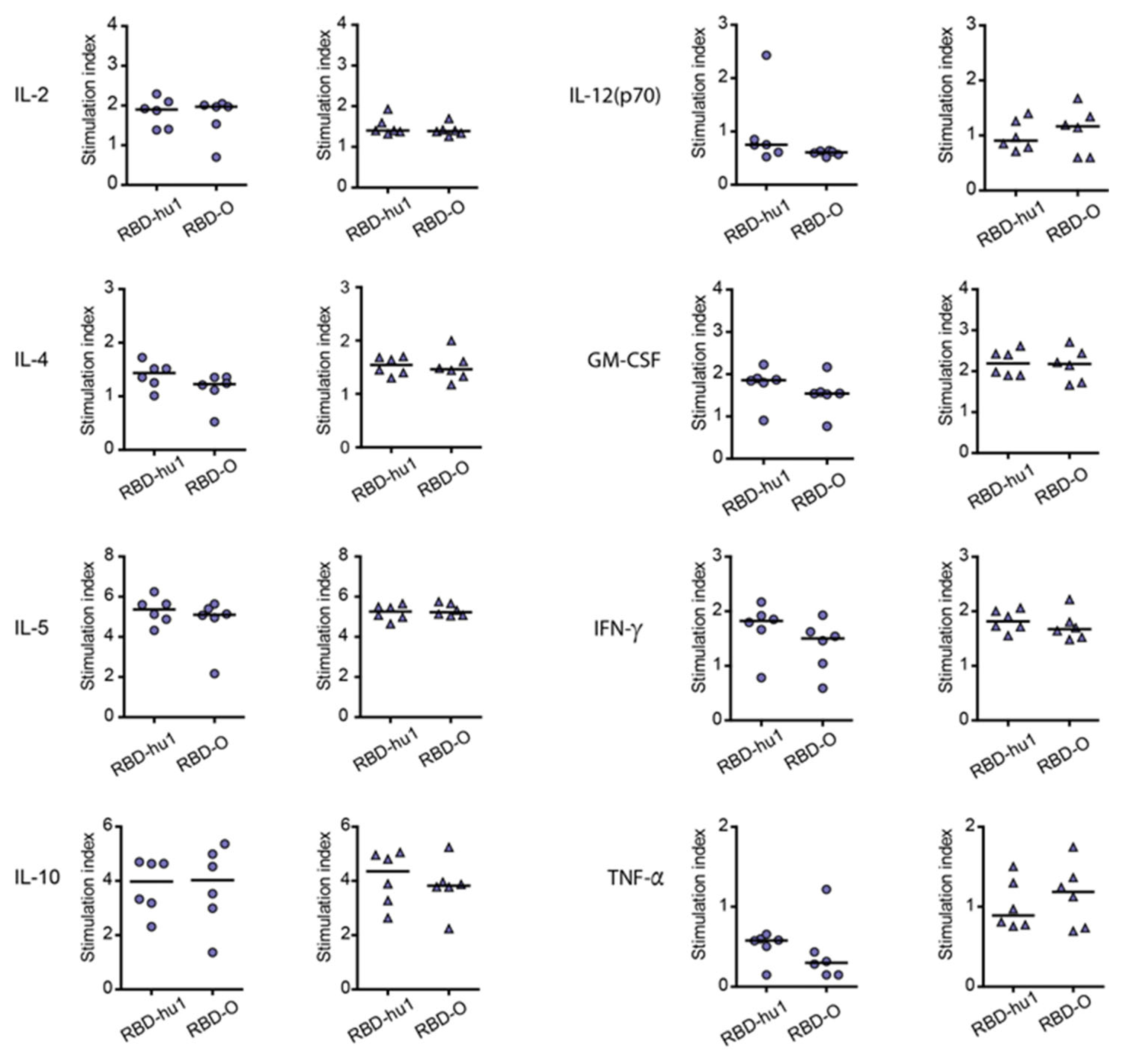
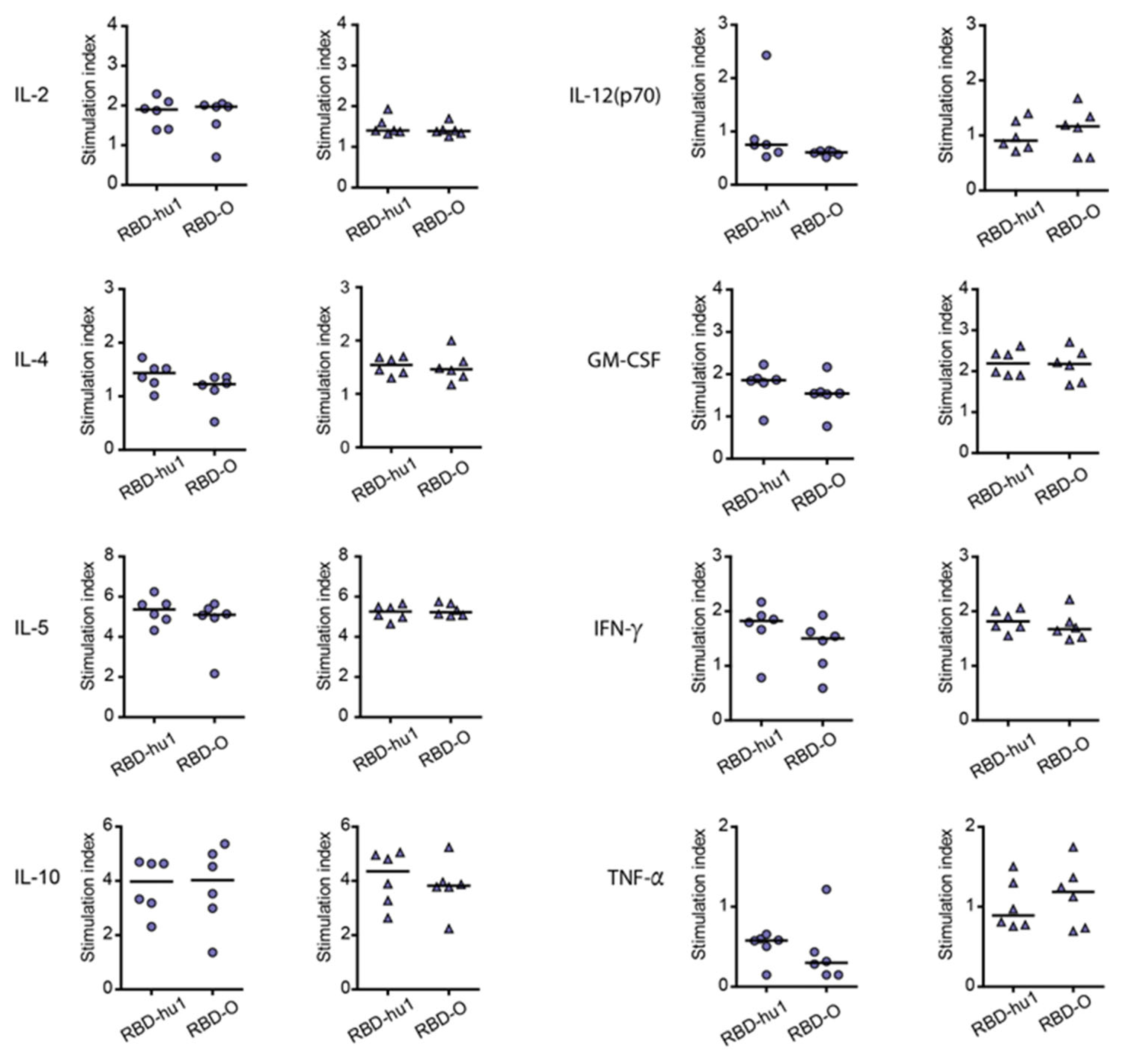
3.4. The Aluminum-Hydroxide Adsorbed W-PreS-O Vaccine Induces a Mixed Th1/Th2 Response Specific for RBD
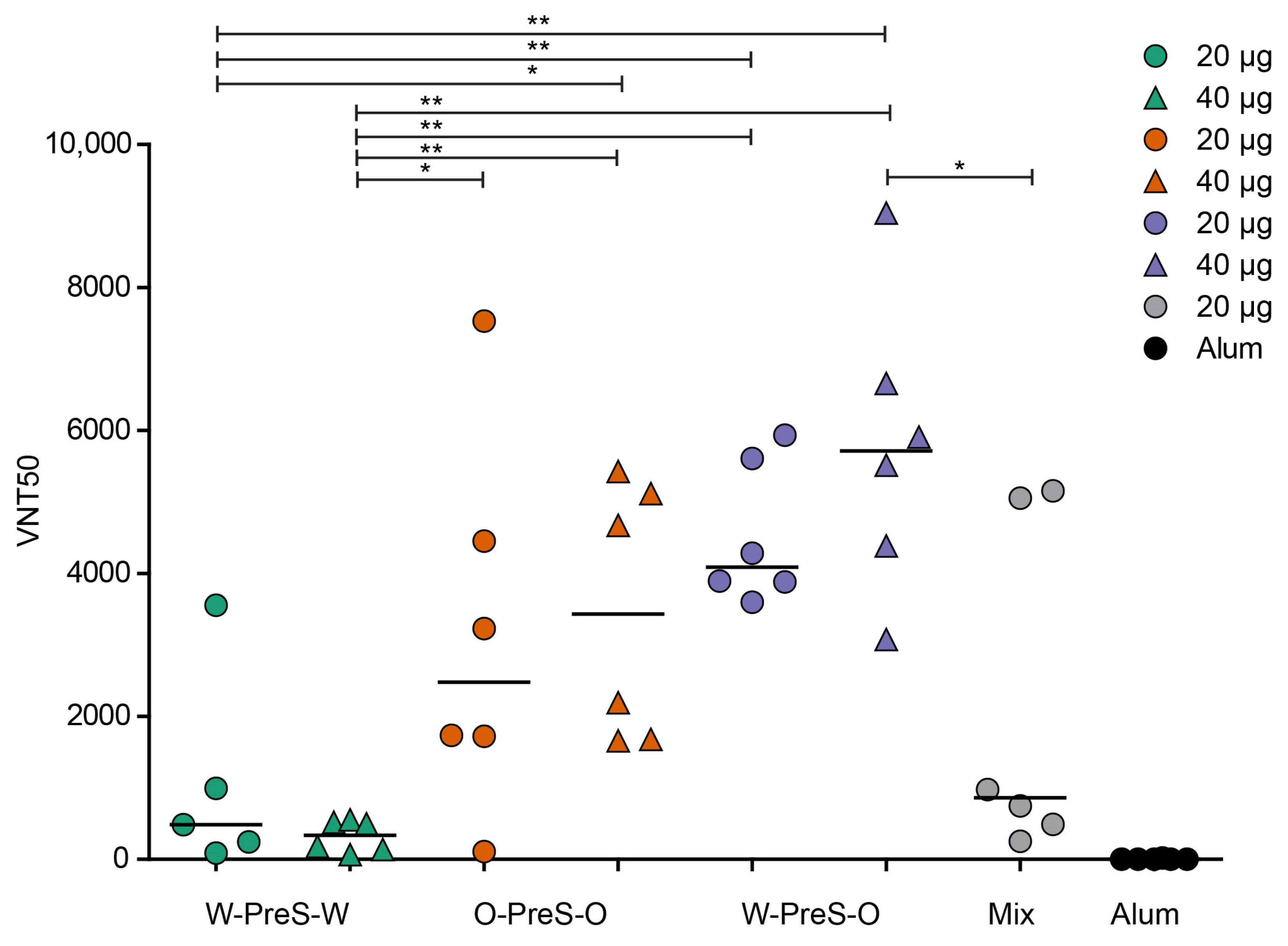
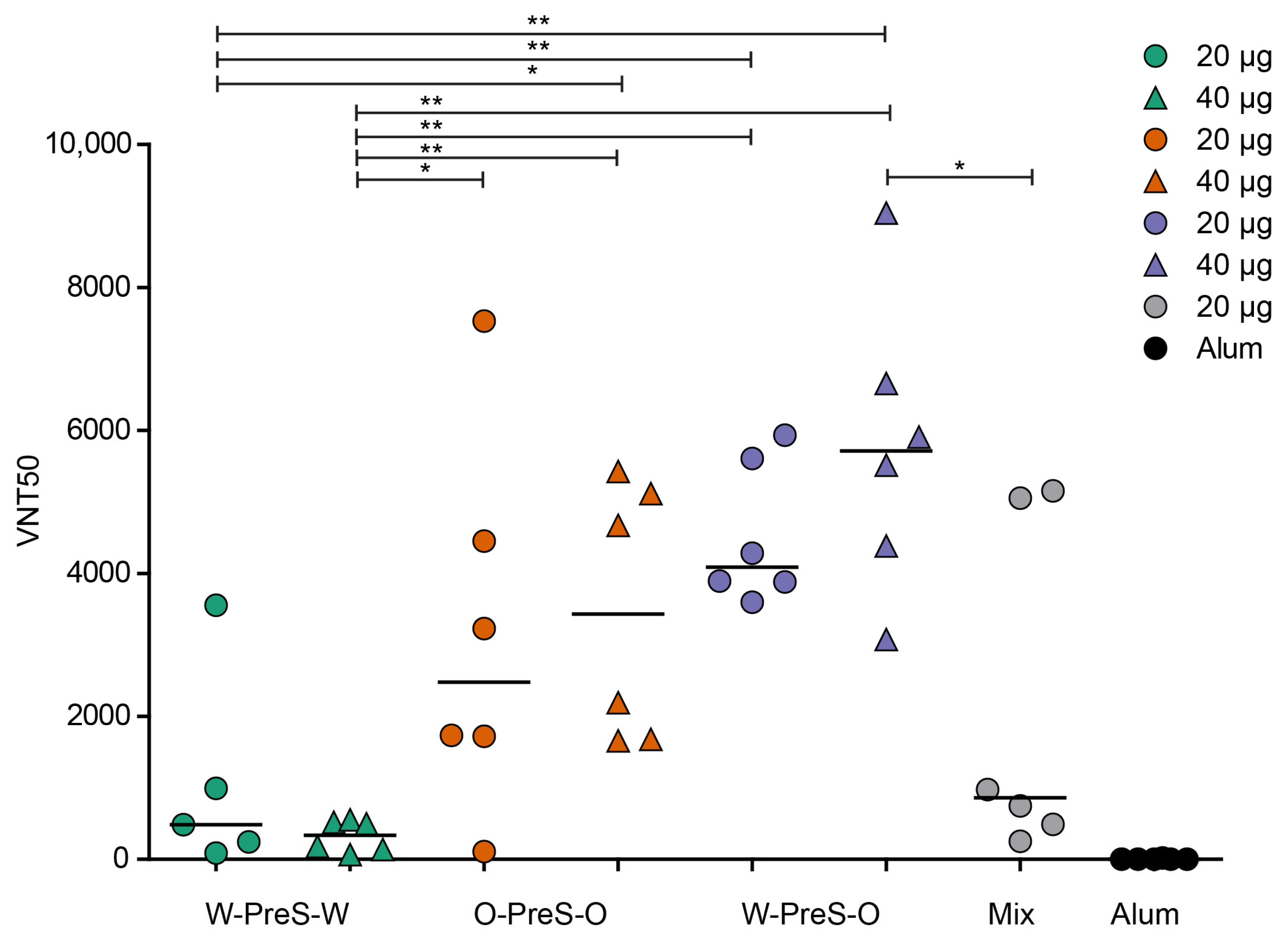
3.5. W-PreS-O Induced the Highest Virus Neutralization Titers against Omicron
3.6. Lack of Association of W-PreS-O-Induced Omicron-Specific Antibody Levels with Neutralization of Omicron
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Roncon, L.; Zuin, M.; Barco, S.; Valerio, L.; Zuliani, G.; Zonzin, P.; Konstantinides, S.V. Incidence of acute pulmonary embolism in COVID-19 patients: Systematic review and meta-analysis. Eur. J. Intern. Med. 2020, 82, 29–37. [Google Scholar] [CrossRef]
- Liu, G.; Chen, T.; Zhang, X.; Hu, B.; Shi, H. Causal relationship between COVID-19 and myocarditis or pericarditis risk: A bidirectional Mendelian randomization study. Front. Cardiovasc. Med. 2023, 10, 1271959. [Google Scholar] [CrossRef] [PubMed]
- Khoshdel-Rad, N.; Zahmatkesh, E.; Shpichka, A.; Timashev, P.; Vosough, M. Outbreak of chronic renal failure: Will this be a delayed heritage of COVID-19? J. Nephrol. 2021, 34, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.C.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune response to SARS-CoV-2 and mechanisms of immunopathological changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Xu, Y.; Wang, T.; Xie, F. Innate and adaptive immune response in SARS-CoV-2 infection-Current perspectives. Front. Immunol. 2022, 13, 1053437. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: Knowns and unknowns of antigen presentation and modes of action. NPJ Vaccines 2021, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.C.; Adams, A.C.; Hufford, M.M.; de la Torre, I.; Winthrop, K.; Gottlieb, R.L. Neutralizing monoclonal antibodies for treatment of COVID-19. Nat. Rev. Immunol. 2021, 21, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Monteil, V.; Kwon, H.; Prado, P.; Hagelkrüys, A.; Wimmer, R.A.; Stahl, M.; Leopoldi, A.; Garreta, E.; Hurtado Del Pozo, C.; Prosper, F.; et al. Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2. Cell 2020, 181, 905–913.e7. [Google Scholar] [CrossRef]
- Rocha, V.P.C.; Quadros, H.C.; Fernandes, A.M.S.; Gonçalves, L.P.; Badaró, R.; Soares, M.B.P.; Machado, B.A.S. An Overview of the Conventional and Novel Methods Employed for SARS-CoV-2 Neutralizing Antibody Measurement. Viruses 2023, 15, 1504. [Google Scholar] [CrossRef]
- Gattinger, P.; Ohradanova-Repic, A.; Valenta, R. Importance, Applications and Features of Assays Measuring SARS-CoV-2 Neutralizing Antibodies. Int. J. Mol. Sci. 2023, 24, 5352. [Google Scholar] [CrossRef]
- Rössler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 Omicron Variant Neutralization in Serum from Vaccinated and Convalescent Persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Cicchitto, G.; Cardillo, L.; de Martinis, C.; Sabatini, P.; Marchitiello, R.; Abate, G.; Rovetti, A.; Cavallera, A.; Apuzzo, C.; Ferrigno, F.; et al. Effects of Casirivimab/Imdevimab Monoclonal Antibody Treatment among Vaccinated Patients Infected by SARS-CoV-2 Delta Variant. Viruses 2022, 14, 650. [Google Scholar] [CrossRef] [PubMed]
- Gattinger, P.; Tulaeva, I.; Borochova, K.; Kratzer, B.; Trapin, D.; Kropfmüller, A.; Pickl, W.F.; Valenta, R. Omicron: A SARS-CoV-2 variant of real concern. Allergy 2022, 77, 1616–1620. [Google Scholar] [CrossRef]
- Shah, M.; Woo, H.G. Omicron: A Heavily Mutated SARS-CoV-2 Variant Exhibits Stronger Binding to ACE2 and Potently Escapes Approved COVID-19 Therapeutic Antibodies. Front. Immunol. 2021, 12, 830527. [Google Scholar] [CrossRef]
- Wolter, N.; Jassat, W.; Walaza, S.; Welch, R.; Moultrie, H.; Groome, M.; Amoako, D.G.; Everatt, J.; Bhiman, J.N.; Scheepers, C.; et al. Early assessment of the clinical severity of the SARS-CoV-2 omicron variant in South Africa: A data linkage study. Lancet 2022, 399, 437–446. [Google Scholar] [CrossRef]
- Menni, C.; Valdes, A.M.; Polidori, L.; Antonelli, M.; Penamakuri, S.; Nogal, A.; Louca, P.; May, A.; Figueiredo, J.C.; Hu, C.; et al. Symptom prevalence, duration, and risk of hospital admission in individuals infected with SARS-CoV-2 during periods of omicron and delta variant dominance: A prospective observational study from the ZOE COVID Study. Lancet 2022, 399, 1618–1624. [Google Scholar] [CrossRef]
- Focosi, D.; Casadevall, A. A Critical Analysis of the Use of Cilgavimab plus Tixagevimab Monoclonal Antibody Cocktail (Evusheld™) for COVID-19 Prophylaxis and Treatment. Viruses 2022, 14, 1999. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Dcosta, B.M.; Landau, N.R.; Tada, T. Resistance of SARS-CoV-2 Omicron BA.1 and BA.2 Variants to Vaccine-Elicited Sera and Therapeutic Monoclonal Antibodies. Viruses 2022, 14, 1334. [Google Scholar] [CrossRef] [PubMed]
- Breznik, J.A.; Rahim, A.; Zhang, A.; Ang, J.; Stacey, H.D.; Bhakta, H.; Clare, R.; Liu, L.M.; Kennedy, A.; Hagerman, M.; et al. Early Omicron infection is associated with increased reinfection risk in older adults in long-term care and retirement facilities. eClinicalMedicine 2023, 63, 102148. [Google Scholar] [CrossRef]
- Nevejan, L.; Ombelet, S.; Laenen, L.; Keyaerts, E.; Demuyser, T.; Seyler, L.; Soetens, O.; Van Nedervelde, E.; Naesens, R.; Geysels, D.; et al. Severity of COVID-19 among Hospitalized Patients: Omicron Remains a Severe Threat for Immunocompromised Hosts. Viruses 2022, 14, 2736. [Google Scholar] [CrossRef]
- Nadesalingam, A.; Cantoni, D.; Aguinam, E.T.; Chan, A.C.; Paloniemi, M.; Ohlendorf, L.; George, C.; Carnell, G.; Lyall, J.; Ferrari, M.; et al. Vaccination and protective immunity to SARS-CoV-2 omicron variants in people with immunodeficiencies. Lancet Microbe 2023, 4, e58–e59. [Google Scholar] [CrossRef]
- Gattinger, P.; Niespodziana, K.; Stiasny, K.; Sahanic, S.; Tulaeva, I.; Borochova, K.; Dorofeeva, Y.; Schlederer, T.; Sonnweber, T.; Hofer, G.; et al. Neutralization of SARS-CoV-2 requires antibodies against conformational receptor-binding domain epitopes. Allergy 2022, 77, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Gattinger, P.; Kratzer, B.; Tulaeva, I.; Niespodziana, K.; Ohradanova-Repic, A.; Gebetsberger, L.; Borochova, K.; Garner-Spitzer, E.; Trapin, D.; Hofer, G.; et al. Vaccine based on folded receptor binding domain-PreS fusion protein with potential to induce sterilizing immunity to SARS-CoV-2 variants. Allergy 2022, 77, 2431–2445. [Google Scholar] [CrossRef] [PubMed]
- mMass Software, Open Source Mass Spectrometry Tool. Available online: http://www.mmass.org/ (accessed on 1 December 2023).
- Kemmer, G.; Keller, S. Nonlinear least-squares data fitting in Excel spreadsheets. Nat. Protoc. 2010, 5, 267–281. [Google Scholar] [CrossRef]
- Wagner, A.; Ohradanova-Repic, A.; Gebetsberger, L.; Tajti, G.; Kundi, M.; Stockinger, H.; Wiedermann, U.; Grabmeier-Pfistershammer, K. Full seroconversion in initial non-responders with higher antibody levels after heterologous COVID-19 vaccination schedule. Immunol. Lett. 2022, 250, 1–6. [Google Scholar] [CrossRef]
- Akinfenwa, O.; Huang, H.J.; Linhart, B.; Focke-Tejkl, M.; Vrtala, S.; Poroshina, A.; Nikonova, A.; Khaitov, M.; Campion, N.J.; Eckl-Dorna, J.; et al. Preventive Administration of Non-Allergenic Bet v 1 Peptides Reduces Allergic Sensitization to Major Birch Pollen Allergen, Bet v 1. Front. Immunol. 2021, 12, 744544. [Google Scholar] [CrossRef]
- GINGER tool Version 1.2. Available online: https://clinicalbiometrics.shinyapps.io/GINGER (accessed on 1 December 2023).
- Zieglmayer, P.; Focke-Tejkl, M.; Schmutz, R.; Lemell, P.; Zieglmayer, R.; Weber, M.; Kiss, R.; Blatt, K.; Valent, P.; Stolz, F.; et al. Mechanisms, safety and efficacy of a B cell epitope-based vaccine for immunotherapy of grass pollen allergy. eBioMedicine 2016, 11, 43–57. [Google Scholar] [CrossRef]
- Ullman-Culleré, M.H.; Foltz, C.J. Body condition scoring: A rapid and accurate method for assessing health status in mice. Lab. Anim. Sci. 1999, 49, 319–323. [Google Scholar] [PubMed]
- Souiri, A.; Lemriss, S.; El Maliki, B.; Falahi, H.; El Fahime, E.; El Kabbaj, S. SARS-CoV-2-Neutralizing Antibody Response and Correlation of Two Serological Assays with Microneutralization. Vaccines 2023, 11, 590. [Google Scholar] [CrossRef]
- Schwarze, M.; Krizsan, A.; Brakel, A.; Pohl, F.; Volke, D.; Hoffmann, R. Cross-Reactivity of IgG Antibodies and Virus Neutralization in mRNA-Vaccinated People Against Wild-Type SARS-CoV-2 and the Five Most Common SARS-CoV-2 Variants of Concern. Front. Immunol. 2022, 13, 915034. [Google Scholar] [CrossRef]
- Hueda-Zavaleta, M.; Gómez de la Torre, J.C.; Cáceres-DelAguila, J.A.; Muro-Rojo, C.; De La Cruz-Escurra, N.; Copaja-Corzo, C.; Aragón-Ayala, C.J.; Benítes-Zapata, V.A. Neutralizing Antibodies as Predictors of Vaccine Breakthrough Infection in Healthcare Workers Vaccinated with or without a Heterologous Booster Dose: A Cohort Study during the Third COVID-19 Wave in Peru. Vaccines 2023, 11, 447. [Google Scholar] [CrossRef]
- Seekircher, L.; Bánki, Z.; Kimpel, J.; Rössler, A.; Schäfer, H.; Falkensammer, B.; Bante, D.; Forer, L.; Schönherr, S.; Harthaller, T.; et al. Immune response after two doses of the BNT162b2 COVID-19 vaccine and risk of SARS-CoV-2 breakthrough infection in Tyrol, Austria: An open-label, observational phase 4 trial. Lancet Microbe 2023, 4, e612–e621. [Google Scholar] [CrossRef]
- Siskind, G.W.; Paul, W.E.; Benacerraf, B. Studies on the effect of the carrier molecule on antihapten antibody synthesis. I. Effect of carrier on the nature of the antibody synthesized. J. Exp. Med. 1966, 123, 673–688. [Google Scholar] [CrossRef]
- McDonald, J.U.; Zhong, Z.; Groves, H.T.; Tregoning, J.S. Inflammatory responses to influenza vaccination at the extremes of age. Immunology 2017, 151, 451–463. [Google Scholar] [CrossRef]
- Ramakrishnan, A.; Althoff, K.N.; Lopez, J.A.; Coles, C.L.; Bream, J.H. Differential serum cytokine responses to inactivated and live attenuated seasonal influenza vaccines. Cytokine 2012, 60, 661–666. [Google Scholar] [CrossRef]
- Martin, J.M.; Avula, R.; Nowalk, M.P.; Lin, C.J.; Horne, W.T.; Chandran, U.R.; Nagg, J.P.; Zimmerman, R.K.; Cole, K.S.; Alcorn, J.F. Inflammatory Mediator Expression Associated with Antibody Response Induced by Live Attenuated vs Inactivated Influenza Virus Vaccine in Children. Open Forum Infect. Dis. 2018, 5, ofy277. [Google Scholar] [CrossRef] [PubMed]
- Alt, F.W.; Ferrier, P.; Malynn, B.; Lutzker, S.; Rothman, P.; Berman, J.; Blackwell, K.; Mellis, S.; Pollock, R.; Furley, A.; et al. Control of recombination events during lymphocyte differentiation. Heavy chain variable region gene assembly and heavy chain class switching. Ann. N. Y. Acad. Sci. 1988, 546, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.B.; Dantas, W.M.; do Nascimento, J.C.F.; da Silva, M.V.; de Oliveira, R.N.; Pena, L.J. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses 2021, 13, 379. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Chan, J.F.; Yuen, K.Y. Animal models in SARS-CoV-2 research. Nat. Methods 2022, 19, 392–394. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gattinger, P.; Kratzer, B.; Sehgal, A.N.A.; Ohradanova-Repic, A.; Gebetsberger, L.; Tajti, G.; Focke-Tejkl, M.; Schaar, M.; Fuhrmann, V.; Petrowitsch, L.; et al. Vaccine Based on Recombinant Fusion Protein Combining Hepatitis B Virus PreS with SARS-CoV-2 Wild-Type- and Omicron-Derived Receptor Binding Domain Strongly Induces Omicron-Neutralizing Antibodies in a Murine Model. Vaccines 2024, 12, 229. https://doi.org/10.3390/vaccines12030229
Gattinger P, Kratzer B, Sehgal ANA, Ohradanova-Repic A, Gebetsberger L, Tajti G, Focke-Tejkl M, Schaar M, Fuhrmann V, Petrowitsch L, et al. Vaccine Based on Recombinant Fusion Protein Combining Hepatitis B Virus PreS with SARS-CoV-2 Wild-Type- and Omicron-Derived Receptor Binding Domain Strongly Induces Omicron-Neutralizing Antibodies in a Murine Model. Vaccines. 2024; 12(3):229. https://doi.org/10.3390/vaccines12030229
Chicago/Turabian StyleGattinger, Pia, Bernhard Kratzer, Al Nasar Ahmed Sehgal, Anna Ohradanova-Repic, Laura Gebetsberger, Gabor Tajti, Margarete Focke-Tejkl, Mirjam Schaar, Verena Fuhrmann, Lukas Petrowitsch, and et al. 2024. "Vaccine Based on Recombinant Fusion Protein Combining Hepatitis B Virus PreS with SARS-CoV-2 Wild-Type- and Omicron-Derived Receptor Binding Domain Strongly Induces Omicron-Neutralizing Antibodies in a Murine Model" Vaccines 12, no. 3: 229. https://doi.org/10.3390/vaccines12030229
APA StyleGattinger, P., Kratzer, B., Sehgal, A. N. A., Ohradanova-Repic, A., Gebetsberger, L., Tajti, G., Focke-Tejkl, M., Schaar, M., Fuhrmann, V., Petrowitsch, L., Keller, W., Högler, S., Stockinger, H., Pickl, W. F., & Valenta, R. (2024). Vaccine Based on Recombinant Fusion Protein Combining Hepatitis B Virus PreS with SARS-CoV-2 Wild-Type- and Omicron-Derived Receptor Binding Domain Strongly Induces Omicron-Neutralizing Antibodies in a Murine Model. Vaccines, 12(3), 229. https://doi.org/10.3390/vaccines12030229