Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues

 ,
, 
 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Treatments, and Sample Collection
2.2. Sample Preparation
2.3. Determination of the Total Phenolic Content (TPC) and the Total Anthocyanin Content (TAC)
2.4. Determination of the Total Tannin Content (TTC)
2.5. Determination of the Antioxidant Activity Using the DPPH, CUPRAC, and FRAP Assays
2.5.1. Sample Preparation
2.5.2. DPPH (2,2-diphenyl-1-picrylhydrazyl) Assay
2.5.3. FRAP (Ferric Reducing Antioxidant Power) Assay
2.5.4. CUPRAC (CUPric Reducing Antioxidant Capacity) Assay
2.6. RNA Extraction, cDNA Synthesis, and qRT-PCR Analysis
2.7. Statistical Analyses
3. Results
3.1. Effect of Chitosan Application on Polyphenols in Grapevine
3.2. Effect of Chitosan Application on Anthocyanins in Grapevine
3.3. Effect of Chitosan Application on Tannins in Grapevine
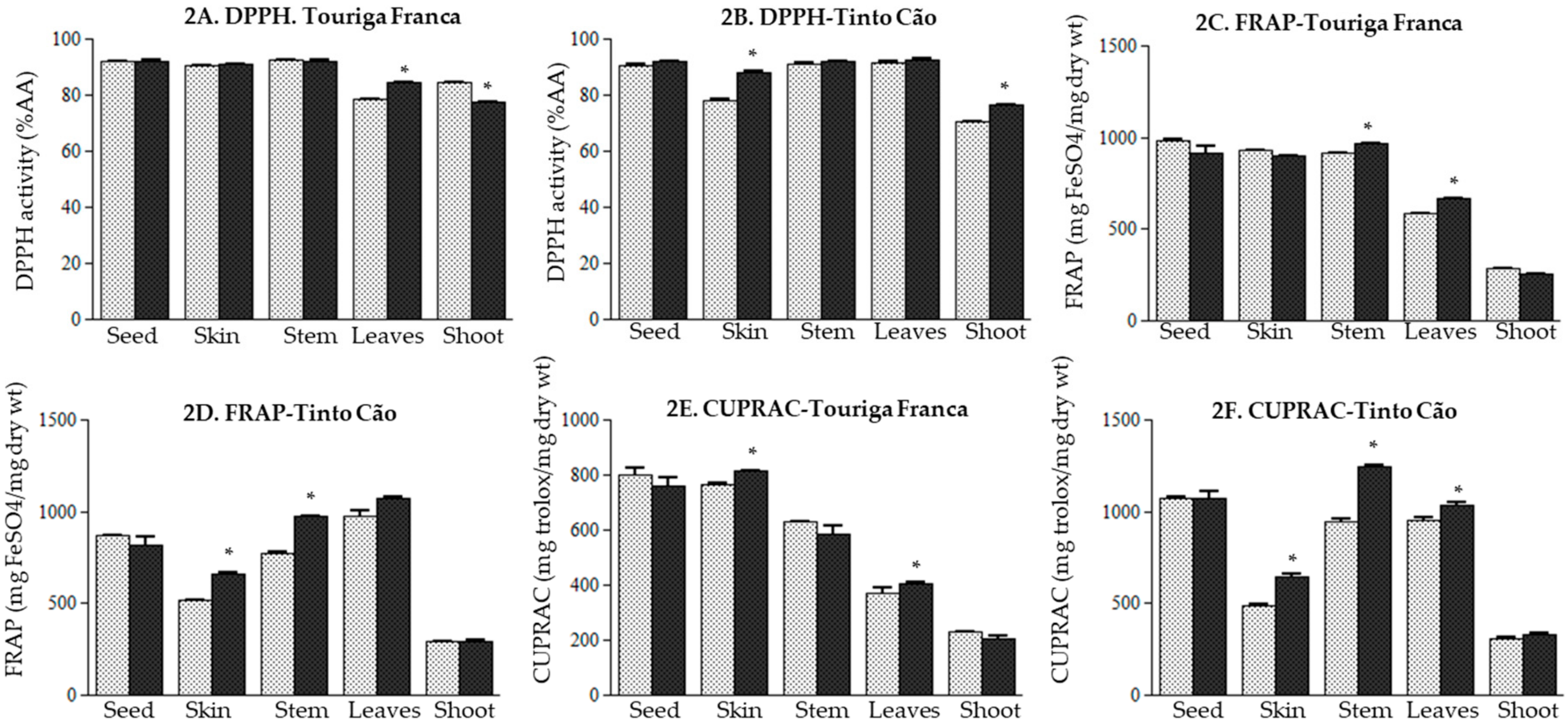
3.4. Effect of Chitosan Application on the Antioxidant Potential of Grapevine
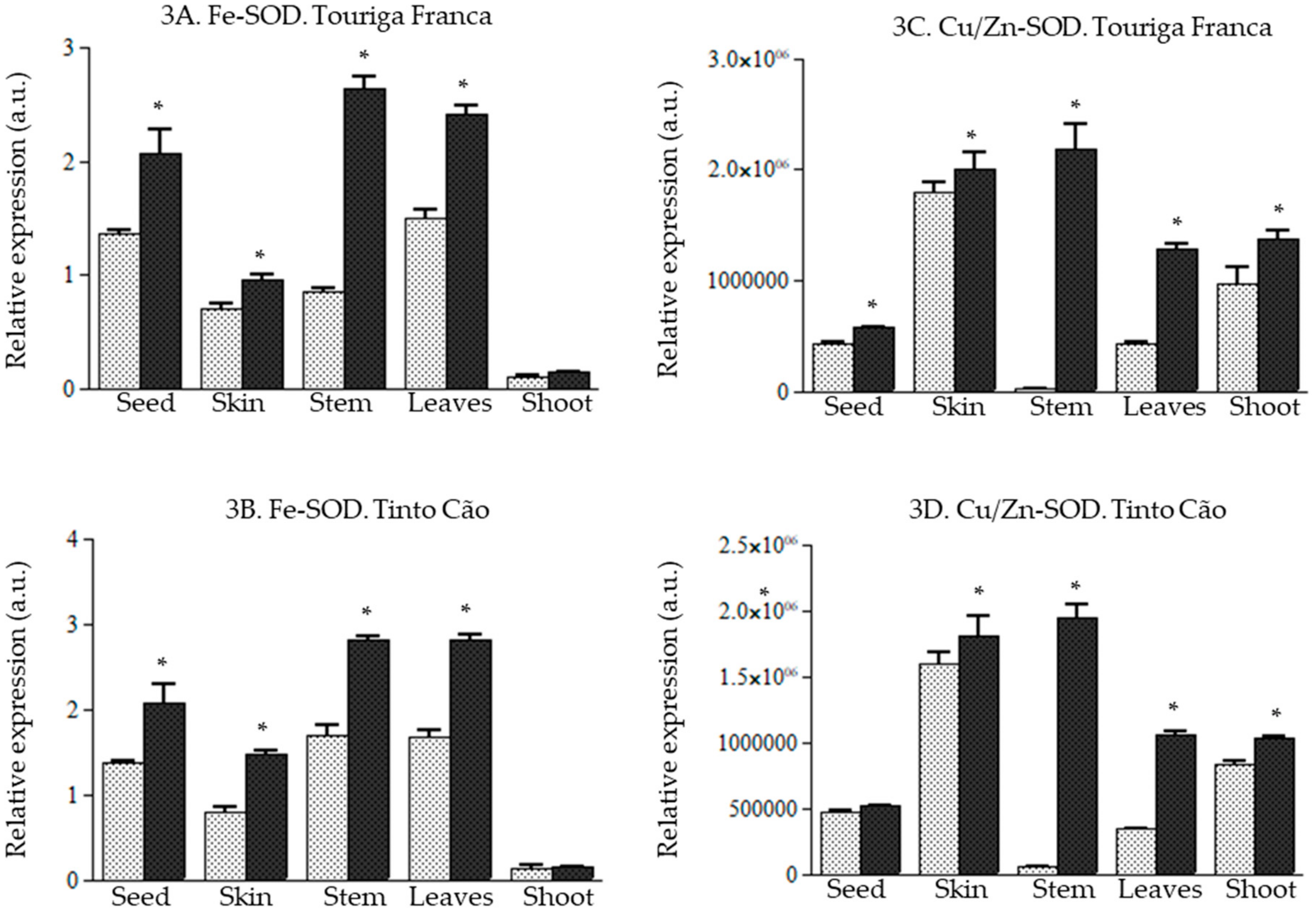
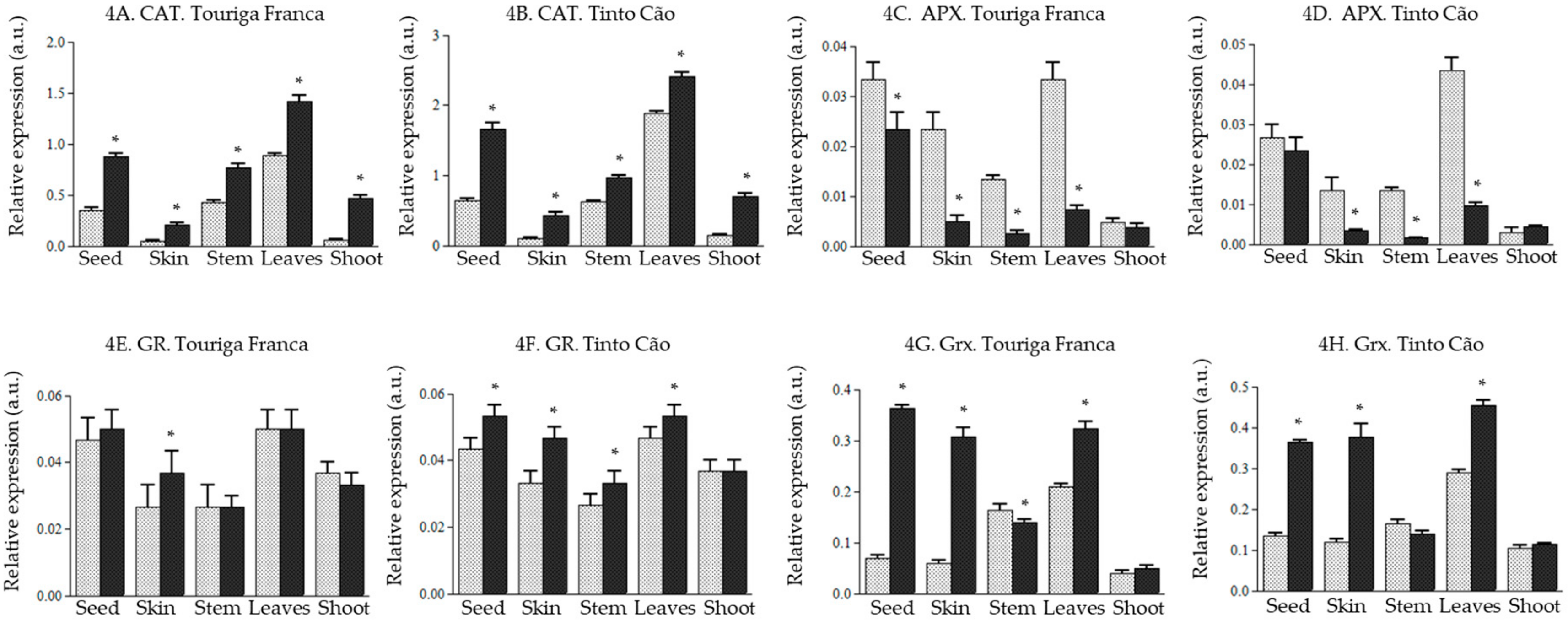
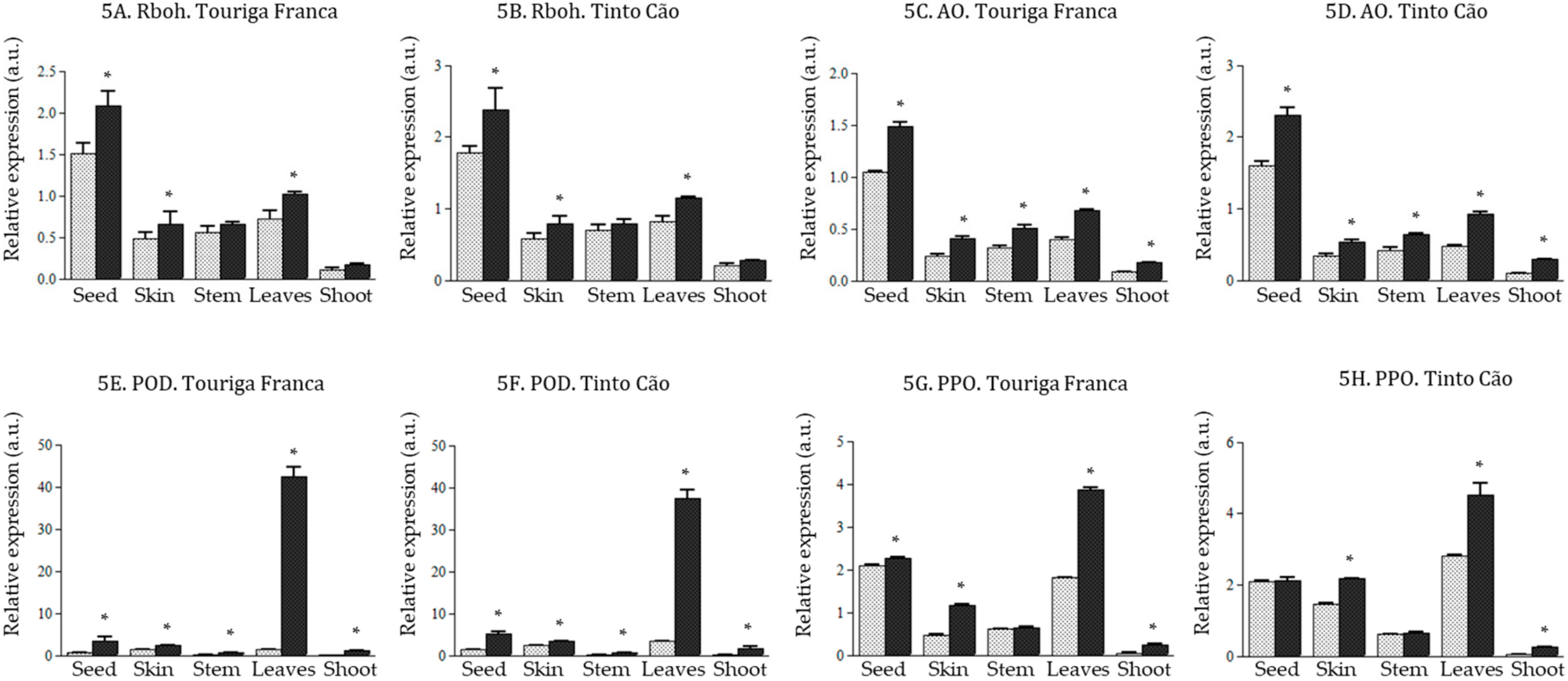
3.5. Effect of Chitosan Application of the Expression of Genes of the ROS Producing and Scavenging Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saw, N.; Riedel, H.; Cai, Z.; Kütük, O.; Smetanska, I. Stimulation of anthocyanin synthesis in grape (Vitis vinifera) cell cultures by pulsed electric fields and ethephon. Plant Cell Tiss. Organ. Cult. 2012, 108, 47–54. [Google Scholar] [CrossRef]
- Mattivi, F.; Zulian, C.; Nicolini, G.; Valenti, L. Wine, biodiversity, technology, and antioxidants. Ann. N.Y. Acad. Sci. 2002, 957, 37–56. [Google Scholar] [CrossRef]
- Lingua, M.S.; Fabani, M.P.; Wunderlin, D.A.; Baroni, M.V. From grape to wine: Changes in phenolic composition and its influence on antioxidant activity. Food Chem. 2016, 208, 228–238. [Google Scholar] [CrossRef]
- Mazza, G. Anthocyanins in grapes and grape products. Crit. Rev. Food Sci. Nutr. 1995, 35, 341–371. [Google Scholar] [CrossRef]
- Tudo, J.A.; Buica, A.; Nieuwoudt, H.; Aleixandre, J.L.; du Toit, W. Spectrophotometric analysis of phenolic compounds in grapes and wines. J. Agric. Food Chem. 2017, 65, 4009–4026. [Google Scholar] [CrossRef] [PubMed]
- Neves, G.G.; Gil, G.; Ferrer, M. Effect of different vineyard treatments on the phenolic contents in tannat (Vitis vinifera L.) grapes and their respective wines. Food Sci. Technol. Int. 2002, 8, 315–321. [Google Scholar] [CrossRef]
- Fanzone, M.; Zamora, F.; Jofre, V.; Assof, M.; Neira, P.A. Phenolic composition of malbec grape skins and seeds from Valle de Uco (Mendoza, Argentina) during Ripening: Effect of cluster thinning. J. Agric. Food Chem. 2011, 59, 6120–6136. [Google Scholar] [CrossRef] [PubMed]
- Soufleros, E.H.; Stavridou, K.; Dagkli, V. The effect of cluster thinning on phenolic maturity of Vitis vinifera cv. Xinomavro grapes. Int. J. Vine Wine Sci. 2011, 45, 171–179. [Google Scholar] [CrossRef]
- Lamela, P.C.; Falcon, G.M.S.; Gándara, S.J.; Fernández, O.I. Influence of grape variety, vine system and enological treatments on the colour stability of young red wines. Food Chem. 2007, 101, 601–606. [Google Scholar] [CrossRef]
- Koundouras, S.; Hatzidimitriou, E.; Karamolegkou, M.; Dimopoulou, E.; Kallithraka, S.; Tsialtas, J.T.; Zioziou, E.; Nikolaou, N.; Kotseridis, Y. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agric. Food Chem. 2009, 57, 7805–7813. [Google Scholar] [CrossRef]
- Romero, P.; Fernandez, F.J.I.; Cutillas, M.A. Physiological thresholds for efficient regulated deficit-irrigation management in wine grapes grown under semiarid conditions. Am. J. Enol. Vitic. 2010, 61, 300–312. [Google Scholar]
- Basile, B.; Marsal, J.; Mata, M.; Vallverdú, X.; Bellvert, J.; Girona, J. Phenological sensitivity of Cabernet Sauvignon to water stress: Vine physiology and berry composition. Am. J. Enol. Vitic. 2011, 62, 452–461. [Google Scholar] [CrossRef]
- Karla, R.E.; Limon, H.V.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusido, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar]
- Arenzana, G.L.; Portu, J.; López, R.; Garijo, P.; Cerdán, G.T.; Alfaro, L.I. Phenylalanine and urea foliar application: Effect on grape and must microbiota. Int. J. Food Microbiol. 2017, 245, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Ruggiero, A.; Rapparini, F.; Neri, L.; Tonni, M.; Iriti, M. The application of chitosan and benzothiadiazole in vineyard (Vitis vinifera L. cv Groppello Gentile) changes the aromatic profile and sensory attributes of wine. Food Chem. 2014, 162, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Cerdán, G.T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 15, 213–221. [Google Scholar] [CrossRef]
- Gonzalez, L.V.; Neira, P.A.; Ibanez, F.; Pastenes, C. Long term effect of abscisic acid (ABA) on the grape berry phenylpropanoids pathway: Gene expression and metabolic content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Tommaso, G.D.; Amico, S.D.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Simmons, W.M.; Hadwiger, L.; Ryan, C.A. Chitosans and pectic polysaccharides both induce the accumulation of the antifungal phytoalexin pisatin in pea pods and antinutrient proteinase inhibitors in tomato leaves. Biochem. Biophys. Res. Commun. 1983, 110, 194–199. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Akila, G.; Einstein, C.R. Chitosan-induced defence responses in tomato plants against early blight disease caused by Alternaria solani (Ellis and Martin) Sorauer. Arch. Phytopathol. Plant Protect. 2014, 47, 1777–1787. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Charles, R.E. Fungal cell wall polymer based nanoparticles in protection of tomato plants from wilt disease caused by Fusarium oxysporumf.sp. lycopersici. Carbohydr. Polym. 2015, 133, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Eullaffroy, P.; Clement, C.; Vernet, G. Chitosan improves development, and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell Rep. 2004, 22, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Malerba, M.; Cerana, R. Chitosan effects on plant systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef] [PubMed]
- Muxika, A.; Etxabide, A.; Uranga, J.; Guerrero, P.; Caba, K. Chitosan as a bioactive polymer: Processing, properties and applications. Int. J. Biol. Macromol. 2017, 105, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Deepmala, K.; Hemantranjan, A.; Singh, B.; Bhanu, A.N. A future perspective in crop protection: Chitosan and its oligosaccharides. Adv. Plants Agric. Res. 2014, 1, 23–30. [Google Scholar]
- Li, Z.; Zhang, Y.; Zhang, X.; Merewitz, E.; Peng, Y.; Ma, X.; Huang, L.; Yan, Y. Metabolic pathways regulated chitosan contributing to drought resistance in white clover. J. Proteome Res. 2017, 16, 3039–3052. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2018, 25, 313–326. [Google Scholar] [CrossRef]
- Sharif, R.; Mujtaba, M.; Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A Review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef]
- Miniawy, S.M.; Ragab, M.; Youssef, S.; Metwally, A. Response of strawberry plants to foliar spraying of chitosan. Res. J. Agric. Biol. Sci. 2013, 9, 366–372. [Google Scholar]
- Kaya, M.; Cesoniene, L.; Daubaras, R.; Leskauskaite, D.; Zabulione, D. Chitosan coating of red kiwifruit (Actinidia melanandra) for extending of the shelf life. Int. J. Biol. Macromol. 2016, 85, 355–360. [Google Scholar] [CrossRef]
- Meng, X.; Tian, S. Effects of preharvest application of antagonistic yeast combined with chitosan on decay and quality of harvested table grape fruit. J. Sci. Food Agric. 2009, 89, 1838–1842. [Google Scholar] [CrossRef]
- Cynkar, W.U.; Cozzolino, D.; Dambergs, R.G.; Janik, L.; Gishen, M. The effects of homogenisation method and freezing on the determination of quality parameters in red grape berries of Vitis vinifera. Aust. J. Grape Wine Res. 2004, 10, 236–242. [Google Scholar] [CrossRef]
- Mercurio, M.D.; Dambergs, R.G.; Herderich, M.J.; Smith, P.A. High throughput analysis of red wine and grape phenolics—adaptation and validation of methyl cellulose precipitable tannin assay and modified Somers color assay to a rapid 96 well plate format. J. Agric. Food Chem. 2007, 55, 4651–4657. [Google Scholar] [CrossRef]
- Sarneckis, C.J.; Dambergs, R.G.; Jones, P.; Mercurio, M.; Herderich, M.J.; Smith, P.A. Quantification of condensed tannins by precipitation with methyl cellulose: Development and validation of an optimised tool for grape and wine analysis. Aust. J. Grape Wine Res. 2006, 12, 39–49. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Moreno, C.S.; Larrauri, J.A.; Calixto, F.S. Procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 270, 270–276. [Google Scholar] [CrossRef]
- Siddhuraju, P.; Becker, K. Antioxidant properties of various solvent extracts of total phenolic constituents from three different agroclimatic origins of drumstick tree (Moringa oleifera Lam.) leaves. J. Agric. Food Chem. 2003, 9, 2144–2155. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 76, 70–76. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kuban, V. Determination of total content of phenolic compounds and their antioxidant activity in vegetables: Evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 8, 607–616. [Google Scholar] [CrossRef]
- Apak, R.; Guçlu, K.; Ozyurek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and Vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC Method. J. Agric. Food Chem. 2004, 29, 7970–7981. [Google Scholar] [CrossRef]
- Gasic, K.; Hernandez, A.; Korban, S.S. RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction. Plant Mol. Biol. Rep. 2004, 22, 437–438. [Google Scholar] [CrossRef]
- Portu, J.; Santamaría, P.; Alfaro, L.I.; López, R.; Cerdán, G.T. Methyl jasmonate foliar application to Tempranillo vineyard improved grape and wine phenolic content. J. Agric. Food Chem. 2015, 63, 2328–2337. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Tassoni, A.; Franceschetti, M.; Righetti, L.; Naldrett, M.J.; Bagni, N. Chitosan treatment induces changes of protein expression profile and stilbene distribution in Vitis vinifera cell suspensions. Proteomics 2009, 9, 610–624. [Google Scholar] [CrossRef]
- Duxbury, M.; Hotter, G.; Reglinski, T.; Sharpe, N. Effect of chitosan and 5-chlorosalicylic acid on total phenolic content of grapes and wine. Am. J. Enol. Viticult. 2004, 55, 191–194. [Google Scholar]
- Reglinski, T.; Elmer, G.; Taylor, J.T.; Wood, N.; Hoyte, S.M. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef]
- Rabêlo, V.M.; Magalhães, P.C.; Bressanin, L.A.; Carvalho, D.T.; Reis, C.O.; Karam, D.; Doriguetto, A.C.; Santos, M.H.; Filho, P.P.S.S.; Souza, T.C. The foliar application of a mixture of semisynthetic chitosan derivatives induces tolerance to water deficit in maize, improving the antioxidant system and increasing photosynthesis and grain yield. Sci. Rep. 2019, 9, 8164. [Google Scholar] [CrossRef]
- García, Y.R.; Cascales, I.R.; Ortín, A.B.B.; Muñoz, R.G.; Cutillas, A.M.; Plaza, E.G. Increasing bioactive phenolic compounds in grapes: Response of six monastrell grape clones to benzothiadiazole and methyl jasmonate treatments. Am. J. Enol. Vitic. 2013, 64, 459–465. [Google Scholar] [CrossRef]
- Espada, M.A.C.; Wood, K.V.; Bordelon, B.; Watkins, B.A. Anthocyanin quantification and radical scavenging capacity of Concord, Norton, and Marechal Foch grapes and wines. J. Agric. Food Chem. 2004, 52, 6779–6786. [Google Scholar] [CrossRef]
- Yang, J.; Martinson, T.E.; Liu, R.H. Phytochemical profiles and antioxidant activities of wine grape. Food Chem. 2009, 116, 332–339. [Google Scholar] [CrossRef]
- Kapłan, M.; Najda, A.; Klimek, K.; Borowy, A. Effect of gibberellic acid (GA3) inflorescence application on content of bioactive compounds and antioxidant potential of grape (Vitis L.) ‘Einset Seedless’ berries. S. Afr. J. Enol. Vitic. 2019, 40, 1–10. [Google Scholar] [CrossRef]
- Khan, W.M.; Prithiviraj, B.; Smith, D.L. Effect of foliar application of chitin and chitosan oligosaccharides on photosynthesis of maize and soybean. Photosynthetica 2002, 40, 621–624. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, X.; Yang, J.; Yin, H.; Wang, W.; Lu, H.; Du, Y. Nitric oxide production and its functional link with OIPK in tobacco defense response elicited by chitooligosaccharide. Plant Cell Rep. 2011, 30, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Dousseau, S.; Rodrigues, A.C.; Sousa Lira, J.M.; Ribeiro Junior, P.M.; Pacheco, F.V.; de Alvarenga, A.A.; Vilela Resende, M.L.; Filha Ferreira de Paula, A.C.C. Exogenous chitosan application on antioxidant systems of Jaborandi. Cienc. Rural 2016, 46, 191–197. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Siadat, S.A.; Bakhshandeh, A.; Pirbalouti, A.G.; Hashemi, M. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis Celak. Crop J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Avramova, V.; AbdElgawad, H.; Vasileva, I.; Petrova, A.S.; Holek, A.; Mariën, J.; Asard, H.; Beemster, G.T. High antioxidant activity facilitates maintenance of cell division in leaves of drought tolerant maize hybrids. Front. Plant Sci. 2017, 8, 84. [Google Scholar] [CrossRef]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Factors influencing antioxidant compounds in rice. Cri. Rev. Food Sci. Nutr. 2017, 57, 893–922. [Google Scholar] [CrossRef]
- Tepperman, J.M.; Dunsmuir, P. Transformed plants with elevated levels of chloroplastic SOD are not more resistant to superoxide toxicity. Plant Mol. Biol. 1990, 14, 501–511. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Yu, J.; Cheng, Y.; Ruan, M.; Wang, R.; Ye, Q.; Zhou, G.; Li, Z.; Yao, Z.; Yang, Y.; et al. The SOD gene family in tomato: Identification, phylogenetic relationships, and expression patterns. Front. Plant Sci. 2016, 7, 1279. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Thiele, D.J. Molecular mechanisms of copper uptake and distribution. Curr. Opin. Plant Biol. 2002, 6, 171–180. [Google Scholar] [CrossRef]
- Hu, X.; Hao, C.; Cheng, Z.M.; Zhong, Y. Genome-wide identification, characterization, and expression analysis of the grapevine superoxide dismutase (SOD) family. Int. J. Genomics 2019, 7350414. [Google Scholar] [CrossRef]
- Leng, X.; Jia, H.; Sun, X.; Shangguan, L.; Mu, Q.; Wang, B.; Fang, J. Comparative transcriptome analysis of grapevine in response to copper stress. Sci. Rep. 2015, 17, 17749. [Google Scholar] [CrossRef]
- Chamnanmanoontham, N.; Pongprayoon, W.; Pichayangkura, R.; Roytrakul, S.; Chadchawan, S. Chitosan enhances rice seedling growth via gene expression network between nucleus and chloroplast. Plant Growth Regul. 2015, 75, 101–114. [Google Scholar] [CrossRef]
- Choudhary, R.C.; Kumarswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci. Rep. 2017, 7, 9754. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Anatomy of a nonhost disease resistance response of pea to Fusarium solani: PR gene elicitation via DNase, chitosanand chromatin alterations. Front. Plant Sci. 2015, 6, 373. [Google Scholar] [CrossRef]
- Zeng, K.; Deng, Y.; Ming, J.; Deng, L. Induction of disease resistance and ROS metabolism in navel oranges by chitosan. Sci. Hortic. 2010, 126, 223–228. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Rudolf, J.R.; Resurreccion, A.V.A. Elicitation of Resveratrol in Peanut Kernels by Application of Abiotic Stresses. J. Agric. Food Chem. 2005, 53, 10186–10192. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Combined elicitation of chitosan and ultraviolet C enhanced stilbene production and expression of chitinase and β-1, 3-glucanase in Vitis vinifera cell suspension cultures. Plant Cell Tissue Organ Cult. 2016, 124, 105–117. [Google Scholar] [CrossRef]
- Brasili, E.; Miccheli, A.; Marini, F.; Praticò, G.; Sciubba, F.; Di Cocco, M.E.; Cechinel, V.F.; Tocci, N.; Valletta, A.; Pasqua, G. Metabolic Profile and Root Development of Hypericum perforatum L. In vitro Roots under Stress Conditions Due to Chitosan Treatment and Culture Time. Front. Plant Sci. 2016, 7, 507. [Google Scholar] [CrossRef]
- Badiali, C.; De Angelis, G.; Simonetti, G.; Elisa, B.; Tobaruela, E.C.; Purgatto, E.; Yin, H.; Valletta, A.; Pasqua, G. Chitosan oligosaccharides affect xanthone and VOC biosynthesis in Hypericum perforatum root cultures and enhance the antifungal activity of root extracts. Plant Cell Rep. 2018, 37, 1471–1484. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Qing-Yan, G.; Xin, W.; Qi-Ping, Q.; Zi-Ying, W.; Jing, L.; Fu, Y.-J. Chitosan elicitation of Isatis tinctoria L. hairy root cultures for enhancing flavonoid productivity and gene expression and related antioxidant activity. Ind. Crop. Prod. 2018, 124, 28–35. [Google Scholar] [CrossRef]
- Valletta, A.; De Angelis, G.; Badiali, C.; Brasili, E.; Miccheli, A.; Di Cocco, M.E.; Pasqua, G. Acetic acid acts as an elicitor exerting a chitosan-like effect on xanthone biosynthesis in Hypericum perforatum L. root cultures. Plant Cell Rep. 2016, 35, 1009–1020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| Fe-SOD | Superoxide dismutase | F 5′ CCTTTGTGAACCTAGGCGAACC 3′ R 5′ TGGCCGGGTTAGCTTGAACTC 3′ |
| Cu/Zn-SOD | Superoxide dismutase | F 5′ AGATTGGCATGTGGTGTTGTTG 3′ R 5′ ACTCCCACATTACCCAACAACA 3′ |
| CAT | Catalase | F 5′ GGTGTTCACACCTTCACTCT 3′ R 5′ GAGATCCTGAGTAGCATGACTG 3′ |
| APX | Ascorbate peroxidase | F 5′ ATCTGGTGGTCATACTCTGG 3′ R 5′ TCTAGGAGAGCCTTGTCTGA 3′ |
| GR | Glutathione reductase | F 5′ AATACTAGGGGGAGGGTACA 3′ R 5′ GTTGTCCTTGGATGCAGA 3′ |
| Grx | Glutaredoxin | F 5′ CTGCTCTCACAGCTAAAAGC 3′ R 5′ CATCACAGAGTCACATCCAC 3′ |
| Rboh | Respiratory burst oxidase | F 5′ CTGATTCTCAGTAGGAACTGGT 3′ R 5′ GAGGTTTGGTCATGTATAGTGC 3′ |
| AO | Amine oxidase | F 5′ GTGAATCCGAACAAGAGAAC 3′ R 5′ AGACTTATTGTAGGGTGTGACC 3′ |
| POD | Peroxidase | F 5′ CCTGATCCAACACTAGATGC 3′ R 5′ GTCTGACTGAAGAAGTCCAGAG 3′ |
| PPO | Polyphenol oxidase | F 5′ CTAGAACTCCAGGTTCATGC 3′ R 5′ GCTTCTCGTCGTAGAGTGAT 3′ |
| Variety/Tissue | Harvesting Time/Treatment | |||||
|---|---|---|---|---|---|---|
| During Veraison | After Veraison | At Complete Maturation | ||||
| Control | Chitosan | Control | Chitosan | Control | Chitosan | |
| ‘Touriga Franca’ | ||||||
| berry seeds | 109.69 ± 5.04 b | 119.27 ± 6.58 a | 89.95 ± 5.13 c | 101.92 ± 2.47 b | 94.67 ± 2.26 c | 93.65 ± 5.10 c |
| berry skins | 105.54 ± 2.73 c | 136.69 ± 5.40 a | 110.61 ± 4.67 c | 135.86 ± 5.37 a | 112.70 ± 4.23 c | 122.80 ± 1.92 ab |
| cluster stems | NA | NA | NA | NA | 88.90 ± 5.55 b | 114.44 ± 5.78 a |
| leaves | 70.28 ± 6.27 c | 87.27 ± 1.70 a | 63.11 ± 0.56 d | 83.18 ± 0.86 a | 55.65 ± 1.09 e | 78.71 ± 1.48 b |
| shoots | NA | NA | NA | NA | 37.23 ± 2.36 a | 34.09 ± 0.64 a |
| ‘Tinto Cão‘ | ||||||
| berry seeds | 192.64 ± 5.34 b | 199.19 ± 2.64 a | 154.15 ± 1.86 d | 182.14 ± 5.54 c | 146.32 ± 4.25 e | 144.05 ± 1.23 e |
| berry skins | 80.50 ± 3.61 f | 99.03 ± 2.14 e | 112.24 ± 5.07 d | 135.13 ± 0.60 b | 129.95 ± 4.17 c | 163.36 ± 9.57 a |
| cluster stems | NA | NA | NA | NA | 158.61 ± 5.28 b | 208.97 ± 8.37 a |
| leaves | 223.28 ± 10.10 b | 246.79 ± 1.92 a | 191.77 ± 3.86 d | 195.77 ± 3.86 d | 204.76 ± 8.89 c | 201.16 ± 3.16 cd |
| shoots | NA | NA | NA | NA | 49.59 ± 2.44 a | 52.65 ± 1.51 a |
| Variety/Tissue | Harvesting time/Treatment | |||||
|---|---|---|---|---|---|---|
| During Veraison | After Veraison | At Complete Maturation | ||||
| Control | Chitosan | Control | Chitosan | Control | Chitosan | |
| ‘Touriga Franca’ | ||||||
| berry seeds | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| berry skins | 16.52 ± 0.44 e | 22.35 ± 0.91 cd | 20.86 ± 0.87 d | 24.49 ± 0.34 ab | 26.02 ± 1.02 a | 23.28 ± 0.55 bc |
| cluster stems | NA | NA | NA | NA | 8.26 ± 0.02 b | 17.66 ± 0.01 a |
| leaves | 0.39 ± 0.02 b | 0.50 ± 0.01 a | 0.32 ± 0.02 c | 0.44 ± 0.01 ab | 0.39 ± 0.01 b | 0.53 ± 0.01 a |
| shoots | NA | NA | NA | NA | 1.55 ± 0.11 a | 1.12 ± 0.02 a |
| ‘Tinto Cão‘ | ||||||
| berry seeds | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| berry skins | 5.32 ± 0.19 f | 8.30 ± 0.40 e | 21.16 ± 1.32 d | 23.39 ± 0.25 c | 28.74 ± 0.97 b | 35.16 ± 1.16 a |
| cluster stems | NA | NA | NA | NA | 2.25 ± 0.01 b | 2.68 ± 0.02 a |
| leaves | 0.76 ± 0.06 b | 0.93 ± 0.04 a | 0.67 ± 0.01 c | 0.77 ± 0.03 b | 0.78 ± 0.03 b | 0.77 ± 0.03 b |
| shoots | NA | NA | NA | NA | 0.14 ± 0.02 a | 0.15 ± 0.01 a |
| Variety/Tissue | Harvesting time/Treatment | |||||
|---|---|---|---|---|---|---|
| During Veraison | After Veraison | At Complete Maturation | ||||
| Control | Chitosan | Control | Chitosan | Control | Chitosan | |
| ‘Touriga Franca’ | ||||||
| berry seeds | 33.19±4.57 e | 141.11 ± 5.95 a | 48.23 ± 5.28 d | 124.93 ± 14.32 b | 95.35 ± 5.32 c | 125.98 ± 4.54 b |
| berry skins | 74.24 ± 3.68 b | 84.78 ± 4.78 a | 59.72 ± 3.78 c | 82.09 ± 6.41 a | 47.41 ± 3.02 d | 57.11 ± 0.76 c |
| cluster stems | NA | NA | NA | NA | 31.63 ± 3.11 a | 20.52 ± 2.28 b |
| leaves | 41.75 ± 5.42 a | 36.90 ± 1.11 b | 35.97 ± 3.45 b | 32.47 ± 2.33 c | 17.06 ± 1.15 d | 32.47 ± 3.37 c |
| shoots | NA | NA | NA | NA | 17.26 ± 1.62 b | 37.02 ± 0.66 a |
| ‘Tinto Cão‘ | ||||||
| berry seeds | 129.04 ± 5.34 b | 140.95 ± 2.64 a | 108.50 ± 1.86 c | 131.63 ± 5.54 b | 106.81 ± 1.23 c | 113.64 ± 4.25 c |
| berry skins | 43.69 ± 3.61 b | 47.89 ± 2.14 a | 26.47 ± 5.07 d | 40.40 ± 0.60 b | 32.03 ± 4.17 c | 47.55 ± 9.57 a |
| cluster stems | NA | NA | NA | NA | 102.66 ± 5.28 b | 140.65 ± 8.37 a |
| leaves | 64.78 ± 10.10 c | 88.19 ± 1.92 a | 62.67 ± 3.86 c | 77.53 ± 3.86 b | 89.43 ± 8.89 a | 78.72 ± 3.16 b |
| shoots | NA | NA | NA | NA | 30.75 ± 2.44 b | 37.97 ± 1.51 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues. Antioxidants 2019, 8, 525. https://doi.org/10.3390/antiox8110525
Singh RK, Soares B, Goufo P, Castro I, Cosme F, Pinto-Sintra AL, Inês A, Oliveira AA, Falco V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues. Antioxidants. 2019; 8(11):525. https://doi.org/10.3390/antiox8110525
Chicago/Turabian StyleSingh, Rupesh K., Bruno Soares, Piebiep Goufo, Isaura Castro, Fernanda Cosme, Ana L. Pinto-Sintra, António Inês, Ana A. Oliveira, and Virgílio Falco. 2019. "Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues" Antioxidants 8, no. 11: 525. https://doi.org/10.3390/antiox8110525
APA StyleSingh, R. K., Soares, B., Goufo, P., Castro, I., Cosme, F., Pinto-Sintra, A. L., Inês, A., Oliveira, A. A., & Falco, V. (2019). Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues. Antioxidants, 8(11), 525. https://doi.org/10.3390/antiox8110525