Antioxidative 1,4-Dihydropyridine Derivatives Modulate Oxidative Stress and Growth of Human Osteoblast-Like Cells In Vitro

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Compounds
2.2. Total Antioxidative Capacity (TAC) Assay
2.3. Total Oxidative Capacity (TOC) Assay
2.4. Cell Culture and Treatments
2.5. Cellular Viability (Tetrazolium Reduction Assay)
2.6. Cell Proliferation (BrdU Assay)
2.7. Measurement of intracellular ROS production
2.8. Determination of Glutathione (GSH) Levels
2.9. Statistical Analysis
3. Results
3.1. Total Antioxidative Capacity
3.2. Total Oxidative Capacity
3.3. Short-Term (1-h) Treatment of HOS Cells with DHPs and Hydrogen Peroxide
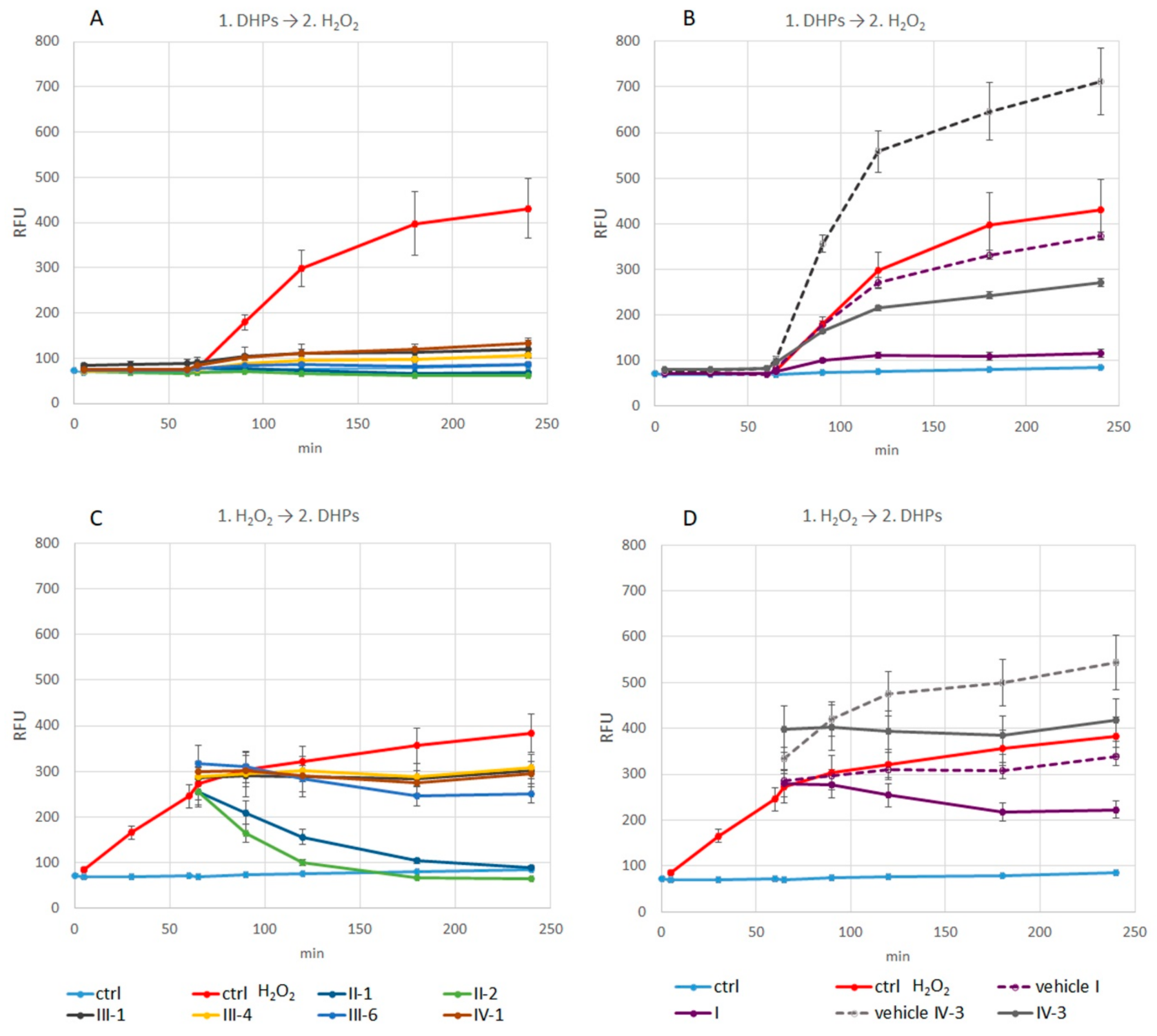
3.3.1. Measurement of Intracellular ROS Production
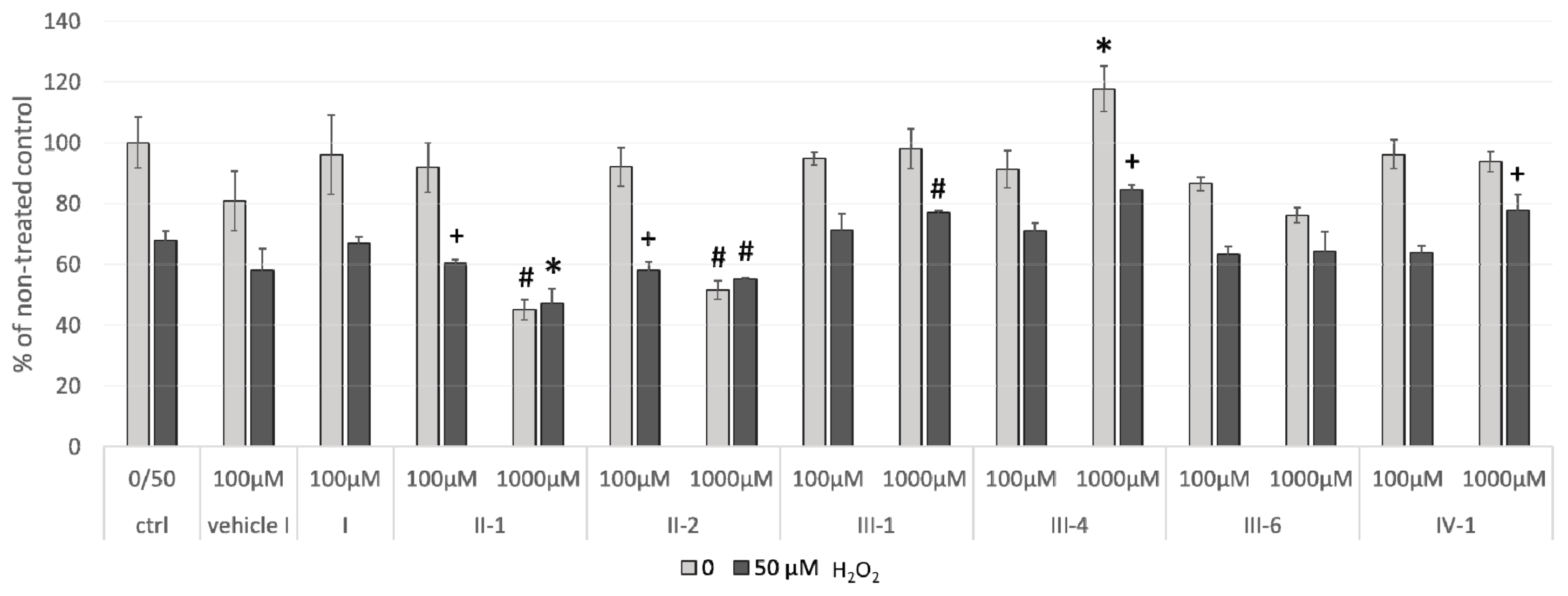
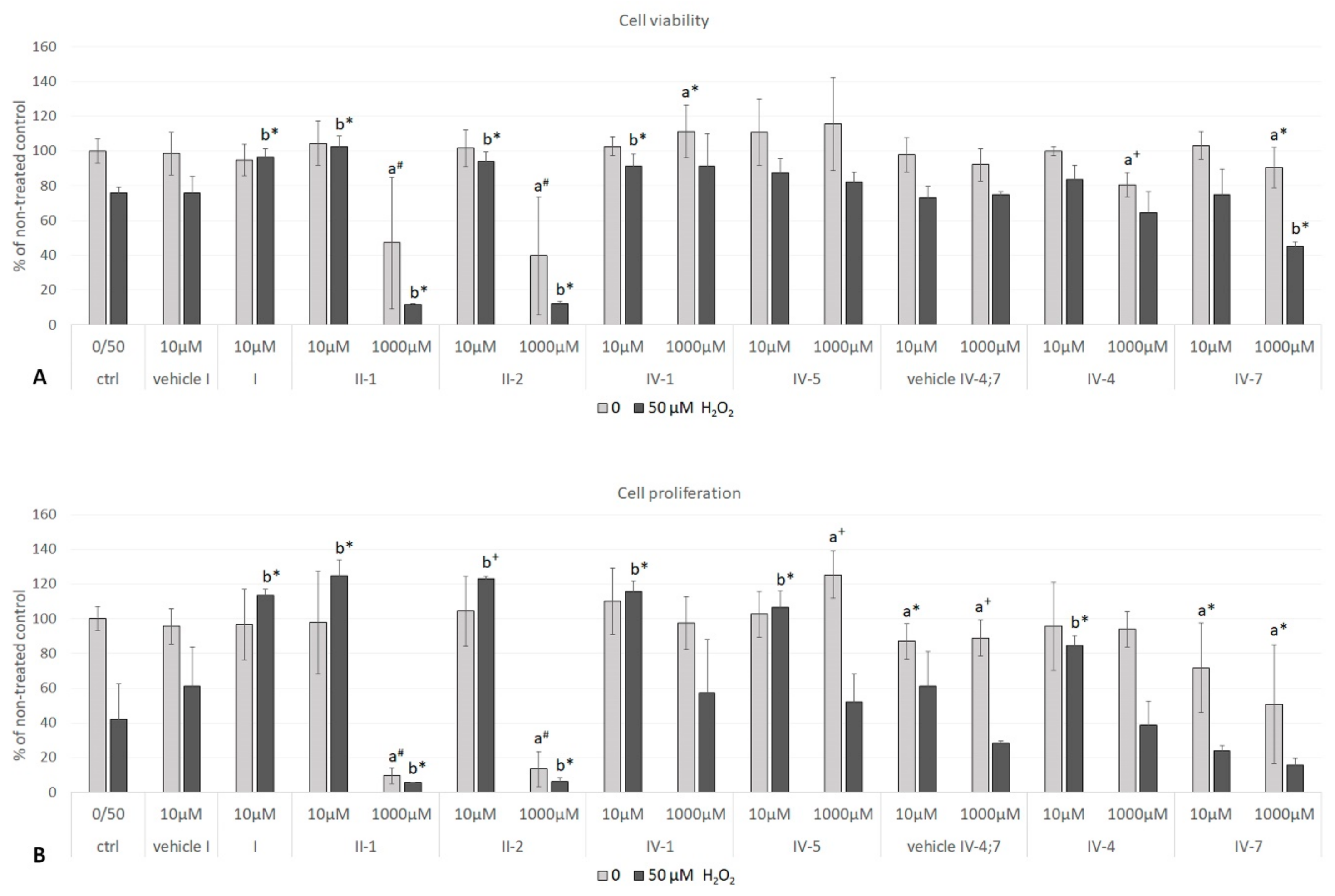
3.3.2. Cellular Viability
3.4. The Effects of 24-h Pre-Treatment of Cells with DHPs before Exposure to Hydrogen Peroxide or Tert-Butyl Hydroperoxide
3.4.1. Hydrogen Peroxide as a Stressor
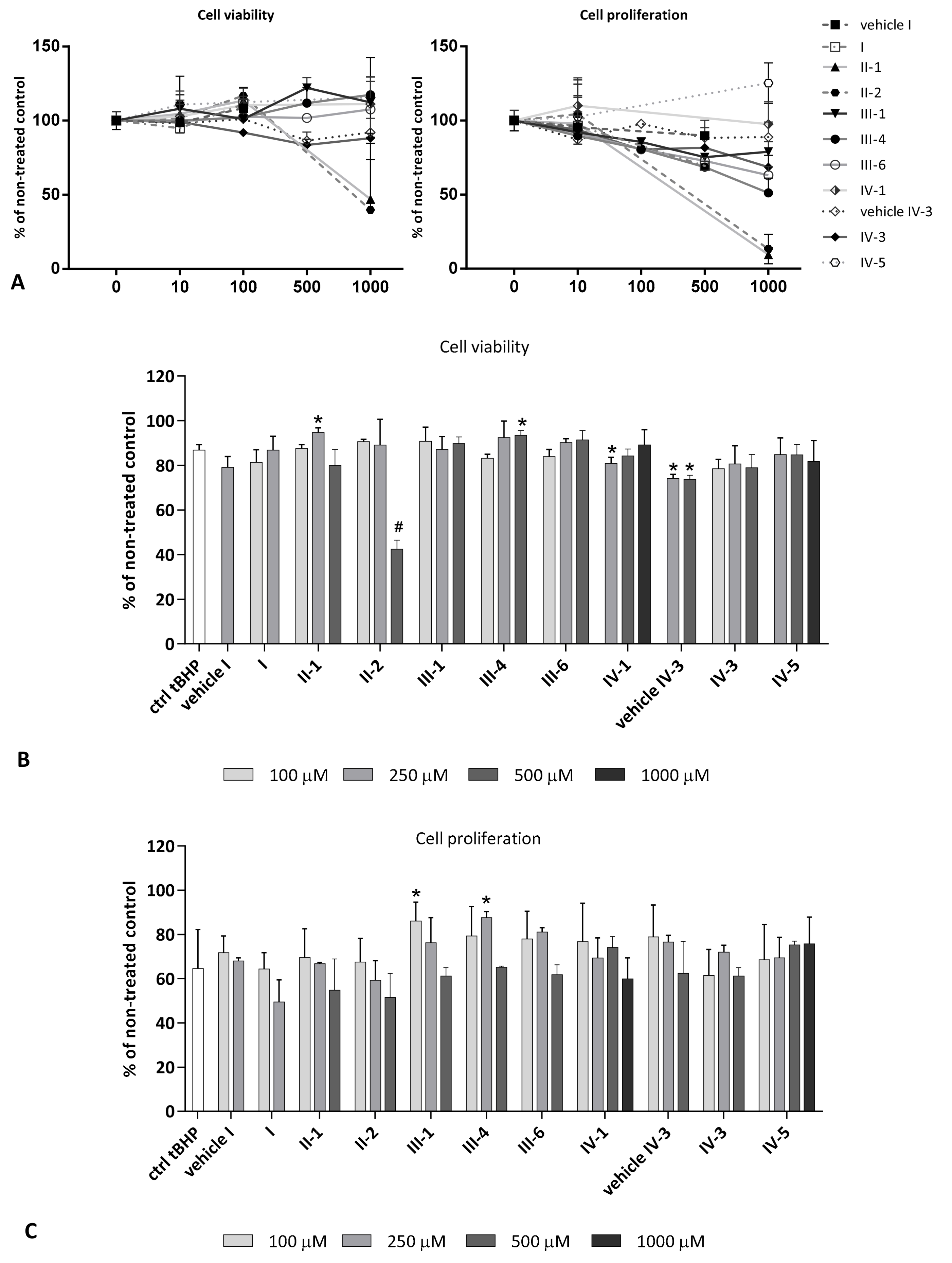
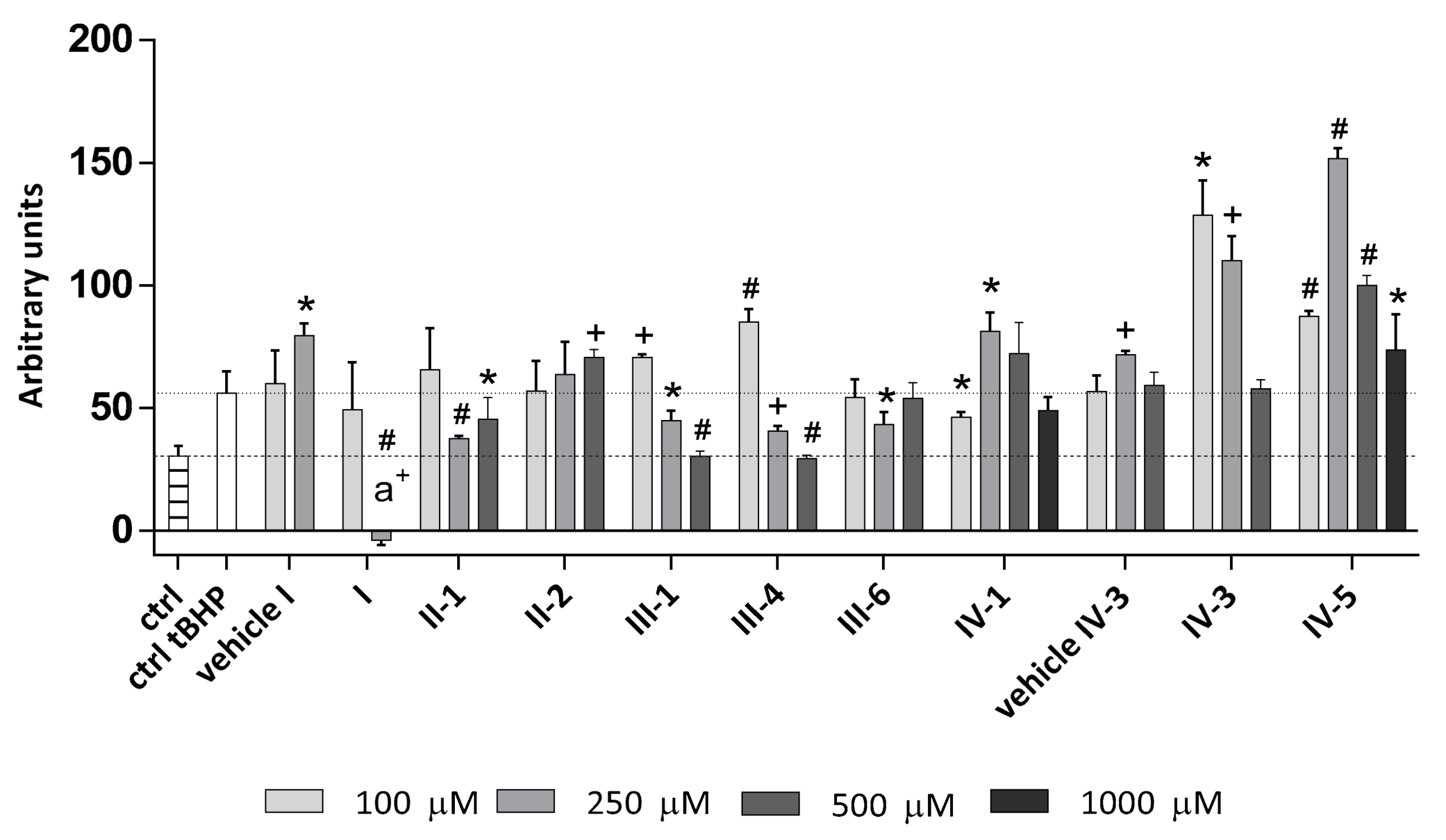
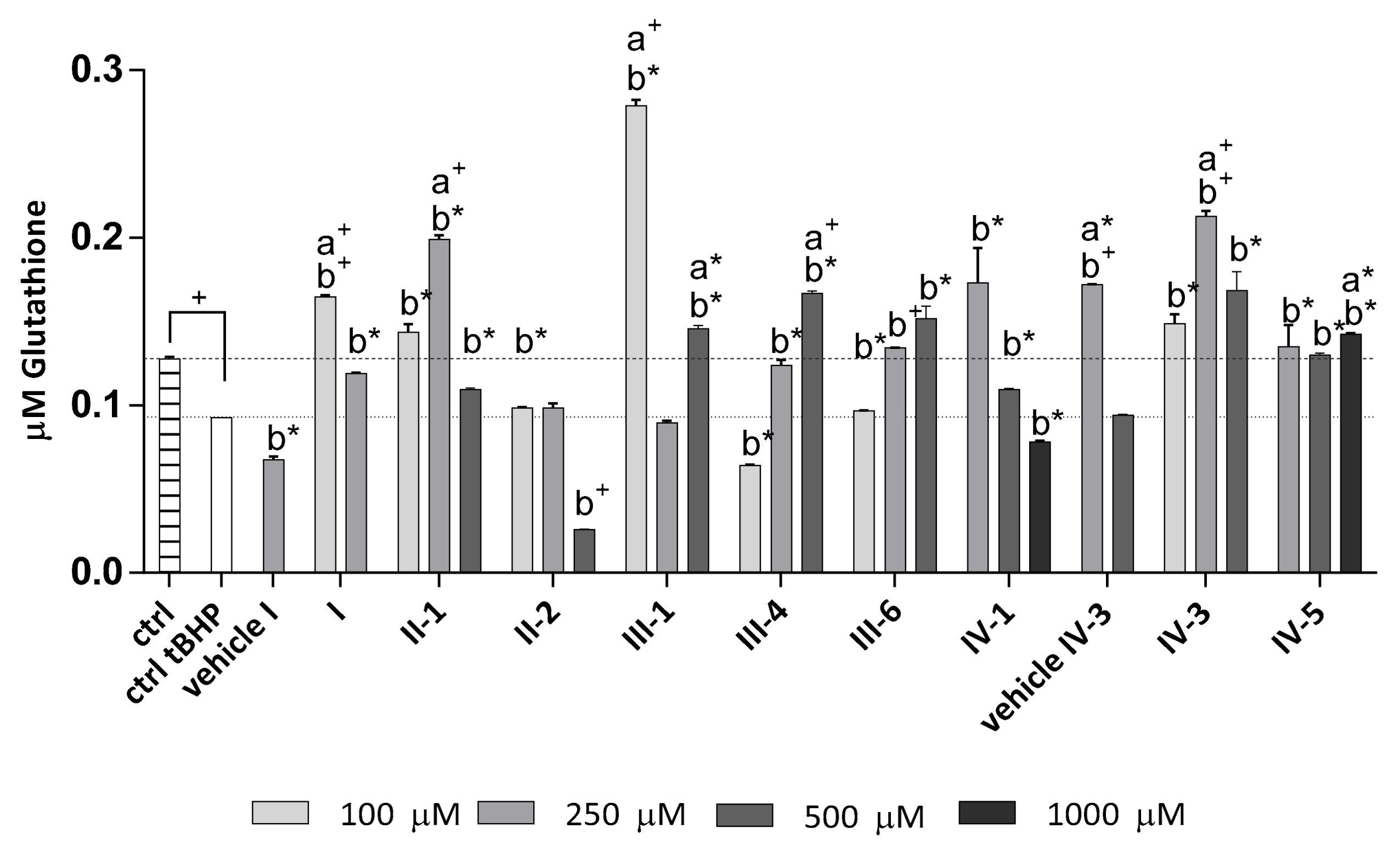
3.4.2. Tert-Butyl Hydroperoxide(tBHP) as a Stressor
Cell Viability and Proliferation
Intracellular ROS Production
Determination of Total Glutathione Level in the Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DHP(s) | 1,4-dihydropyridine(s) |
| OS | oxidative stress |
| AO | Antioxidant |
| AOA | antioxidative activity |
| TAC | total antioxidative capacity |
| TOC | total oxidative capacity |
| ROS | reactive oxygen species |
| HRP | horse radish peroxidase |
| TMB | 3,3′,5,5′-tetramethylbenzydine |
| DCFH-DA | 2′,7′-dichlorodihydrofluorescein diacetate |
| tBHP | tert-butyl hydroperoxide |
| BrdU | 5-bromo-2′-deoxyuridine |
| HOS | human osteosarcoma. |
References
- Sies, H. Oxidative stress: Introductory remarks. In Oxidative Stress; Sies, H., Ed.; Academic Press: London, UK, 1985; pp. 1–7. [Google Scholar]
- Milkovic, L.; Siems, W.; Siems, R.; Zarkovic, N. Oxidative stress and antioxidants in carcinogenesis and integrative therapy of cancer. Curr. Pharm. Des. 2014, 20, 6529–6542. [Google Scholar] [CrossRef] [PubMed]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. Corrigendum to “European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS)”. Redox Biol. 2018, 14, 694–696. [Google Scholar] [CrossRef] [PubMed]
- Long, M.J.C.; Poganik, J.R.; Ghosh, S.; Aye, Y. Subcellular redox targeting: Bridging in vitro and in vivo chemical biology. ACS Chem. Biol. 2017, 12, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Žarković, N.; Lončarić, I.; Čipak, A.; Jurić, G.; Wonisch, W.; Borović, S.; Waeg, G.; Vuković, T.; Žarković, K. [Pathophysiological characteristics of secondary messengers of free radicals and oxidative stress]. Patofiziološke značajke sekundarnih glasnika slobodnih radikala i oksidativni stress. In Oksidativni Stres i Djelotvornost Antioksidansa (Second Messengers of Free Radicals; Oxidative Stres); Bradamante, V., Lacković, Z., Eds.; Medicinska Naklada: Zagreb, Croatia, 2001; pp. 13–32. ISBN 953-176-127-2. [Google Scholar]
- Swarnalatha, G.; Prasanthi, G.; Sirisha, N.; Madhusudhana Chetty, C. 1,4-Dihydropyridines: A multtifunctional molecule—A review. Int. J. ChemTech Res. 2011, 3, 75–89. [Google Scholar]
- Velena, A.; Zarkovic, N.; Gall Troselj, K.; Bisenieks, E.; Krauze, A.; Poikans, J.; Duburs, G. 1,4-Dihydropyridine derivatives: Dihydronicotinamide analogues—Model compounds targeting oxidative stress. Oxid. Med. Cell. Longev. 2016, 2016, 1892412. [Google Scholar] [CrossRef] [PubMed]
- Mercola, M.; Cashman, J.; Lanier, M.; Willems, E.; Schade, D. Compounds for Stem Cell Differentiation. U.S. Patent OO9233926B2, 12 January 2016. [Google Scholar]
- Velẽna, A.; Zilbers, J.; Duburs, G. Derivatives of 1,4-dihydropyridines as modulators of ascorbate-induced lipid peroxidation and high-amplitude swelling of mitochondria, caused by ascorbate, sodium linoleate and sodium pyrophosphate. Cell Biochem. Funct. 1999, 17, 237–252. [Google Scholar] [CrossRef]
- Ryabokon, N.I.; Goncharova, R.I.; Duburs, G.; Rzeszowska-Wolny, J. A 1,4-dihydropyridine derivative reduces DNA damage and stimulates DNA repair in human cells in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 587, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Uldrikis, Y.R.; Zidermane, A.A.; Biseniex, E.A.; Preisa, I.E.; Dubur, G.Y.; Tirzit, G.D. Esters of 2,6-dimethyl-1,4-dihydropyridine-3,5-dicarboxylic Acid and Method of Obtaining. Thereof. Patent WO 8000345 A1, 8 August 1978. [Google Scholar]
- Buraka, E.; Chen, C.Y.-C.; Gavare, M.; Grube, M.; Makarenkova, G.; Nikolajeva, V.; Bisenieks, I.; Brūvere, I.; Bisenieks, E.; Duburs, G.; et al. DNA-binding studies of AV-153, an antimutagenic and DNA repair-stimulating derivative of 1,4-dihydropiridine. Chem. Biol. Interact. 2014, 220, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Ošiņa, K.; Rostoka, E.; Sokolovska, J.; Paramonova, N.; Bisenieks, E.; Duburs, G.; Sjakste, N.; Sjakste, T. 1,4-Dihydropyridine derivatives without Ca2+-antagonist activity up-regulate Psma6 mRNA expression in kidneys of intact and diabetic rats. Cell Biochem. Funct. 2016, 34, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Ošiņa, K.; Leonova, E.; Isajevs, S.; Baumane, L.; Rostoka, E.; Sjakste, T.; Bisenieks, E.; Duburs, G.; Vīgante, B.; Sjakste, N. Modifications of expression of genes and proteins involved in DNA repair and nitric oxide metabolism by carbatonides [disodium-2,6-dimethyl-1,4-dihydropyridine-3,5-bis(carbonyloxyacetate) derivatives] in intact and diabetic rats. Arh. Hig. Rada Toksikol. 2017, 68, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Rostoka, E.; Sokolovska, J.; Sjakste, N.; Sjakste, T.; Ošiņa, K. 1,4-Dihidropiridīna atvasinājums DNS pārrāvumu novēršanai cukura diabēta apstākļos. Patent of the Republic of Latvia, Nr. P-15-89 LV15181B, 20 August 2015. [Google Scholar]
- Marie, P.J.; Kassem, M. Osteoblasts in osteoporosis: Past, emerging, and future anabolic targets. Eur. J. Endocrinol. 2011, 165, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.G.; Rao, A.V. Oxidative stress and antioxidants in the risk of osteoporosis—Role of the antioxidants lycopene and and polyphenols. Top Osteoporos. 2013, 117–161. [Google Scholar] [CrossRef]
- Marinucci, L.; Balloni, S.; Fettucciari, K.; Bodo, M.; Talesa, V.N.; Antognelli, C. Nicotine induces apoptosis in human osteoblasts via a novel mechanism driven by H2O2 and entailing Glyoxalase 1-dependent MG-H1 accumulation leading to TG2-mediated NF-κB desensitization: Implication for smokers-related osteoporosis. Free Radic. Biol. Med. 2018, 117, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Shimodaka, K.; Okura, F.; Shimizu, Y.; Saito, I.; Yanaihara, T. Osteoblast cells (MG-63 and HOS) have aromatase and 5α-reductase activities. Biochem. Mol. Biol. Int. 1996, 39, 109–116. [Google Scholar]
- Rose, F.R.; Cyster, L.A.; Grant, D.M.; Scotchford, C.A.; Howdle, S.M.; Shakesheff, K.M. In vitro assessment of cell penetration into porous hydroxyapatite scaffolds with a central aligned channel. Biomaterials 2004, 25, 5507–5514. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Lee, S.R.; Park, H.J.; Byun, H.E.; Sohn, E.H.; Kim, B.O.; Rhee, D.K.; Pyo, S. Kobophenol A enhances proliferation of human osteoblast-like cells with activation of the p38 pathway. Int. Immunopharmacol. 2013, 17, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Norcross, B.E.; Clement, G.; Weinstein, M. The Hantzsch pyridine synthesis: A factorial design experiment for the introductory organic laboratory. J. Chem. Educ. 1969, 46, 694–695. [Google Scholar] [CrossRef]
- Uldrikis, J.; Preisa, I.; Duburs, G.; Zidermane, A.; Bisenieks, E.; Tirzitis, G. 2,6-Dimethyl-1,4-dihydropyridine-3,5-dicarboxylic Acid Esters and Method for Preparing Same. WO Patent 8000345 A1 19800306, 6 March 1980. [Google Scholar]
- Pubulis, K.; Stonans, I.; Jonane-Osa, I.; Jansone, I.; Bisenieks, E.; Kalvins, I.; Vigante, B.; Bruvere, I.; Uldrikis, J.; Zuka, L.; et al. Antiviral Efficacy of Disodium 2,6-dimethyl-1,4-dihydropyridine-3,5-bis(carbonyloxyacetate) and Its Derivatives. European Patent Application EP 2 578 218 A1, 6 October 2011. [Google Scholar]
- Bisenieks, E.; Duburs, G.; Stonans, I.; Jaschenko, E.; Domracheva, I.; Poikans, J.; Bruvere, I.; Kalvins, I.; Shestakova, I.; Uldrikis, J.; et al. Pharmaceutical Combination of 5-fluorouracil and Derivative of 1,4-dihydropyridine and Its Use in the Treatment of Cancer. EP Patent 2228365 A1, 15 September 2010. [Google Scholar]
- Dubur, G.Y.; Uldrikis, Y.R. Preparation of 3,5-diethoxycarbonyl-2,6-dimethyl-1,4-dihydro-isonicotinic acid and 3,5-diacetyl-2,6-dimethyl-1,4-dihydroisoni-cotinic acid and their salts. Chem. Heterocycl. Compd. 1972, 5, 762–763. [Google Scholar] [CrossRef]
- Bisenieks, E.A.; Dubur, G.Y.; Uldrikis, Y.R.; Veveris, M.M.; Kimenis, A.A.; Ivanov, E.V. 2-(2,6-Dimethyl-3,5-diethoxycarbonyl-1,4-dihydropyridine-4-carboxamide Glutaric Acid Its Disodium Salt and Method of Their Preparation. DE Patent 3337521 A1, 26 April 1984. [Google Scholar]
- Stonans, I.; Jansone, I.; Jonane-Osa, I.; Bisenieks, E.; Duburs, G.; Kalvins, I.; Vigante, B.; Uldrikis, J.; Bruvere, I.; Zuka, L.; et al. Derivatives of 1,4-dihydropyridine Possessing Antiviral Efficacy. U.S. Patent 20130131126A1, 23 May 2013. [Google Scholar]
- Bruvere, I.; Bisenieks, E.; Poikans, J.; Uldrikis, J.; Plotniece, A.; Pajuste, K.; Rucins, M.; Vigante, B.; Kalme, Z.; Gosteva, M.; et al. Dihydropyridine derivatives as cell growth modulators in vitro. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Borovic, S.; Tirzitis, G.; Tirzite, D.; Cipak, A.; Khoschsorur, G.A.; Waeg, G.; Tatzber, F.; Scukanec-Spoljar, M.; Zarkovic, N. Bioactive 1,4-dihydroisonicotinic acid derivatives prevent oxidative damage of liver cells. Eur. J. Pharmacol. 2006, 537, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Tatzber, F.; Griebenow, S.; Wonisch, W.; Winkler, R. Dual method for the determination of peroxidase activity and total peroxides-iodide leads to a significant increase of peroxidase activity in human sera. Anal. Biochem. 2003, 316, 147–153. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rodrigues, C.; Mósca, A.F.; Martins, A.P.; Nobre, T.; Prista, C.; Antunes, F.; Gasparovic, A.C.; Soveral, G. Rat aquaporin-5 is pH-gated induced by phosphorylation and is implicated in oxidative stress. Int. J. Mol. Sci. 2016, 17, 2090. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Shahzad, D.; Larik, F.A.; Channar, P.A.; Mahfooz, H.; Abbas, Q.; Hassan, M.; Raza, H.; Seo, S.-Y.; Shabir, G. Synthesis of 4-aryl-2,6-dimethyl-3,5-bis-N-(aryl)-carbamoyl-1,4-dihydropyridines as novel skin protecting and anti-aging agents. Bangladesh J. Pharmacol. 2017, 12, 25. [Google Scholar] [CrossRef]
- Hu, R.; Li, S.-L.; Bai, H.-T.; Wang, Y.-X.; Liu, L.-B.; Lv, F.-T.; Wang, S. Regulation of oxidative stress inside living cells through polythiophene derivatives. Chin. Chem. Lett. 2016, 27, 545–549. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Li, Y.; Wang, F.; Yang, F.; Xu, W. Synthesis and radioprotective activity of mitochondria targeted dihydropyridines in vitro. Int. J. Mol. Sci. 2017, 18, 2233. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, E.V.; Ponomarjova, T.V.; Merkusev, G.N.; Dubur, G.J.; Bisenieks, E.A.; Dauvarte, A.Z.; Pilscik, E.M. A new skin radioprotective agent Diethon (experimental study). Radiobiol. Radiother. (Berl.) 1990, 31, 69–78. [Google Scholar] [PubMed]
- Hong, S.E.; Urahashi, S.; Kamata, R. Skin radioprotector (Diethone) modifying dermal response of radiation on rats. J. Korean Soc. Ther. Radiol. 1989, 7, 15–22. [Google Scholar]
- Pandupuspitasari, N.S.; Khan, F.A.; Sameeullah, M.; Rehman, Z.U.; Sudjatmogo; Muktiani, A.; Yi, J. Effects of diludine on the production, oxidative status, and biochemical parameters in transition cows. J. Environ. Agric. Sci. 2016, 6, 3–9. [Google Scholar]
- Fernandes, M.A.S.; Santos, M.S.; Vicente, J.A.F.; Moreno, A.J.M.; Velena, A.; Duburs, G.; Oliveira, C.R. Effects of 1,4-dihydropyridine derivatives (cerebrocrast, gammapyrone, glutapyrone, and diethone) on mitochondrial bioenergetics and oxidative stress: A comparative study. Mitochondrion 2003, 3, 47–59. [Google Scholar] [CrossRef]
- Filipan-Litvić, M.; Litvić, M.; Vinković, V. An efficient, metal-free, room temperature aromatization of Hantzsch-1,4-dihydropyridines with urea-hydrogen peroxide adduct, catalyzed by molecular iodine. Tetrahedron 2008, 64, 5649–5656. [Google Scholar] [CrossRef]
- Tirzit, G.D.; Kazush, E.Y.; Dubur, G.Y. Influence of 1,4-dihydropyridine derivatives on the generation of hydroxyl radicals. Chem. Heterocycl. Compd. 1992, 28, 435–437. [Google Scholar] [CrossRef]
- Josephy, P.D.; Eling, T.; Mason, R.P. The horseradish peroxidase-catalyzed oxidation of 3,5,3’,5’-tetramethylbenzidine. Free radical and charge-transfer complex intermediates. J. Biol. Chem. 1982, 257, 3669–3675. [Google Scholar] [PubMed]
- Duburs, G.Y.; Kumerova, A.O.; Uldrikis, Y.R. Enzymic oxidation of hydrogenated pyridines with peroxidase-hydrogen peroxide system. Latv. PSR ZA Vestis 1970, 73, 73–77. [Google Scholar]
- Afanasyeva, M.S.; Taraban, M.B.; Polyakov, N.E.; Purtov, P.A.; Leshina, T.V.; Grissom, C.B. Elementary steps of enzymatic oxidation of nifedipine catalyzed by horseradish peroxidase. J. Phys. Chem. B 2006, 110, 21232–21237. [Google Scholar] [CrossRef] [PubMed]
- Huyser, E.S.; Harmony, J.A.K.; McMillian, F.L. Peroxide oxidations of dihydropyridine derivatives. J. Am. Chem. Soc. 1972, 94, 3176–3180. [Google Scholar] [CrossRef]
- Lovaković, T.; Poljak-Blazi, M.; Duburs, G.; Cipak, A.; Cindrić, M.; Vigante, B.; Bisenieks, E.; Jaganjac, M.; Mrakovcić, L.; Dedić, A.; et al. Growth modulation of human cells in vitro by mild oxidative stress and 1,4-dihydropyridine derivative antioxidants. Coll. Antropol. 2011, 35, 137–141. [Google Scholar] [PubMed]
- Wilson, C. Bone: Oxidative stress and osteoporosis. Nat. Rev. Endocrinol. 2014, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Alía, M.; Ramos, S.; Mateos, R.; Bravo, L.; Goya, L. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxicol. 2005, 19, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Feng, H.; Sun, W.; Liu, K.; Lu, J.-J.; Chen, X. Tert-butyl hydroperoxide (t-BHP) induced apoptosis and necroptosis in endothelial cells: Roles of NOX4 and mitochondrion. Redox Biol. 2017, 11, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.C.; Teng, C.C.; Shen, C.H.; Huang, W.S.; Lu, C.C.; Kuo, H.C.; Tung, S.Y. Protective effect of black garlic extracts on tert-butyl hydroperoxide-induced injury in hepatocytes via a c-Jun N-terminal kinase-dependent mechanism. Exp. Ther. Med. 2018, 15, 2468–2474. [Google Scholar] [CrossRef] [PubMed]
- Langston, J.W.; Li, W.; Harrison, L.; Aw, T.Y. Activation of promoter activity of the catalytic subunit of γ-glutamylcysteine ligase (GCL) in brain endothelial cells by insulin requires antioxidant response element 4 and altered glycemic status: Implication for GCL expression and GSH synthesis. Free Radic. Biol. Med. 2011, 51, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Aktay, G.; Emre, M.H.; Polat, A. Influence of dihydropyridine calcium antagonist nitrendipine on benzo(a)pyrene-induced oxidative stress. Arch. Pharm. Res. 2011, 34, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Zarkovic, N.; Saso, L. Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol. 2017, 12, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Lisk, C.; McCord, J.; Bose, S.; Sullivan, T.; Loomis, Z.; Nozik-Grayck, E.; Schroeder, T.; Hamilton, K.; Irwin, D.C. Nrf2 activation: A potential strategy for the prevention of acute mountain sickness. Free Radic. Biol. Med. 2013, 63, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N. 4-hydroxynonenal as a bioactive marker of pathophysiological processes. Mol. Aspects Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Zarkovic, K.; Jakovcevic, A.; Zarkovic, N. Contribution of the HNE-immunohistochemistry to modern pathological concepts of major human diseases. Free Radic. Biol. Med. 2017, 111, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Gasparovic, A.C.; Milkovic, L.; Sunjic, S.B.; Zarkovic, N. Cancer growth regulation by 4-hydroxynonenal. Free Radic. Biol. Med. 2017, 111, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Hoppe, A.; Detsch, R.; Boccaccini, A.R.; Zarkovic, N. Effects of Cu-doped 45S5 bioactive glass on the lipid peroxidation-associated growth of human osteoblast-like cells in vitro. J. Biomed. Mater. Res. A 2014, 102, 3556–3561. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Cipak Gasparovic, A.; Zarkovic, N. Overview on major lipid peroxidation bioactive factor 4-hydroxynonenal as pluripotent growth-regulating factor. Free Radic. Res. 2015, 49, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Mouthuy, P.-A.; Snelling, S.J.B.; Dakin, S.G.; Milković, L.; Gašparović, A.Č.; Carr, A.J.; Žarković, N. Biocompatibility of implantable materials: An oxidative stress viewpoint. Biomaterials 2016, 109, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Herbert, V. Prooxidant effects of antioxidant vitamins. Introduction. J. Nutr. 1996, 126, 1197S–1200S. [Google Scholar] [CrossRef] [PubMed]
- Tafazoli, S.; Wright, J.S.; O’Brien, P.J. Prooxidant and antioxidant activity of vitamin E analogues and troglitazone. Chem. Res. Toxicol. 2005, 18, 1567–1574. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C. Pro-Oxidants or Antioxidants. (Pro-oxidants and Antioxidants. What Characterizes an Antioxidant). In Proceedings of the Sunrise Free Radical School Free Radical Biology (SFRBM), San Francisco, CA, USA, 19–21 November 2009; University of Otago: Christchurch, New Zealand, 2009; pp. 1–33. Available online: https://pdfs.semanticscholar.org/presentation/83c9/974cfea4d3171558be4814673d22bbb7f857.pdf (accessed on 16 July 2018).
- Shimizu, H.; Nakagami, H.; Yasumasa, N.; Mariana, O.K.; Kyutoku, M.; Koriyama, H.; Nakagami, F.; Shimamura, M.; Rakugi, H.; Morishita, R. Cilnidipine, but not amlodipine, ameliorates osteoporosis in ovariectomized hypertensive rats through inhibition of the N-type calcium channel. Hypertens. Res. 2012, 35, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, K.; Liu, Y.; Maekawa, T.; Ishikawa, E.; Motosugi, Y.; Ando, H.; Tsuruoka, S.; Fujimura, A. Protective effect of amlodipine against osteoporosis in stroke-prone spontaneously hypertensive rats. Eur. J. Pharmacol. 2010, 635, 227–230. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Compounds | Trivial Name | R3 | R4 | R5 | Solubility | Reference |
|---|---|---|---|---|---|---|
| I | Diethone | Ethanol with 10% DMSO | [23] | |||
| II-1 | Carbatone | ONa | H | OCH2COONa | Water | [24] |
| II-2 | J-9-133-2 | ONa | H | OCH3 | Water | [25] |
| II-3 | J-9-46 | OCH2CH3 | H | OCH2CH3 | Ethanol | [25] |
| II-4 | Metcarbatone | ONa | CH3 | OCH2COONa | Water | [26] |
| II-5 | Etcarbatone | ONa | CH2CH3 | OCH2COONa | Water | [26] |
| II-6 | J-9-117 (Styrylcarbatone) | ONa | CH=CHPh | OCH2COONa | Water | *MM |
| III-1 | Thiocarbatone | ONa | H | SCH2COONa | Water | MM |
| III-2 | TK-2 | OCH3 | H | SCH2COOCH3 | Ethanol with 10% DMSO | MM |
| III-3 | E-170-4 (TK-1) | OCH2CH3 | H | SCH2COOCH2CH3 | Ethanol with 10% DMSO | MM |
| III-4 | E2-134-1 | ONa | H | OCH2CH3 | Water | MM |
| III-5 | E2-135 | OCH2CH3 | H | OCH2CH3 | Ethanol | MM |
| III-6 | E2-136-2 | ONa | H | OCH2COONa | Water | MM |
| III-7 | E2-131 | ONa | CH2CH3 | OCH2CH3 | Water | MM |
| III-8 | E2-130-3 | OCH2CH3 | CH2CH3 | OCH2CH3 | Ethanol | MM |
| III-9 | E2-120 (ETK-2) | OCH3 | CH2CH3 | SCH2COOCH3 | Ethanol | MM |
| III-10 | E2-113 (ETK-1) | OCH2CH3 | CH2CH3 | SCH2COOCH2CH3 | Ethanol | MM |
| III-11 | E-163-1 | ONa | 3-Py | OCH2CH3 | Water | MM |
| III-12 | E-163-K | OCH2CH3 | 3-Py | OCH2CH3 | Ethanol | MM |
| IV-1 | AV-153 | OCH2CH3 | ONa | OCH2CH3 | Water | [27] |
| IV-2 | EE-126 | OCH3 | ONa | OCH3 | Water | MM |
| IV-3 | E3-46 | OCH2CH3 | OH | OCH2COOCH2CH3 | Ethanol | MM |
| IV-4 | V-6-55-1 | OCH2COOCH2CH3 | OH | OCH2COOCH2CH3 | Ethanol | MM |
| IV-5 | AV-154-Na | CH3 | ONa | CH3 | Water | [27] |
| IV-6 | J-11-61B | CH3 | OH | OCH2CH3 | Ethanol | MM |
| IV-7 | J-11-71-2 | OCH2CH3 | OH | Ethanol | MM | |
| IV-8 | Glutapyrone | OCH2CH3 | NHCH(CH2)2COONa|COONa | OCH2CH3 | Water | [28] |
| V | J-12-25 | Ethanol | MM |
| Compounds | Trivial Name | Concentrations Tested | ||
|---|---|---|---|---|
| 10 µM | 100 µM | 1000 µM | ||
| Equivalent to mM Uric Acid | ||||
| I | Diethone | 1.463 ± 1.016 | 1.465 ± 0.085 | >10 |
| II-1 | Carbatone | 0.438 ± 0.177 | 1.534 ± 0.064 | 9.692 ± 0.036 |
| II-2 | J-9-133-2 | 0.585 ± 0.074 | 3.756 ± 0.312 | >10 |
| II-3 | J-9-46 | 0 | 0.148 ± 0.088 | 2.284 ± 0.504 |
| II-4 | Metcarbatone | 0 | 0.526 ± 0.033 | 0.852 ± 0.013 |
| II-5 | Etcarbatone | 0.027 ± 0.038 | 0.057 ± 0.019 | 0.206 ± 0.085 |
| II-6 | J-9-117 (Styrylcarbatone) | 0.680 ± 0.051 | 0.421 ± 0.596 | 1.073 ± 0.04 |
| III-1 | Thiocarbatone | 1.279 ± 0.046 | 2.735 ± 0.363 | >10 |
| III-2 | TK-2 | 0 | 0 | 0 |
| III-3 | E-170-4 (TK-1) | 0 | 0 | 0 |
| III-4 | E2-134-1 | 0.770 ± 0.075 | 0.541 ± 0.304 | >10 |
| III-5 | E2-135 | 0.027 ± 0.302 | 0.564 ± 0.422 | 1.968 ± 0.294 |
| III-6 | E2-136-2 | 0.498 ± 0.115 | 1.082 ± 0.175 | >10 |
| III-7 | E2-131 | 0.778 ± 0.085 | 0 | 0 |
| III-8 | E2-130-3 | 0.355 ± 0.234 | 0.285 ± 0.398 | 0.980 ± 0.129 |
| III-9 | E2-120 (ETK-2) | 0 | 0.042 ± 0.007 | 0 |
| III-10 | E2-113 (ETK-1) | 0.032 ± 0.195 | 0.077 ± 0.219 | 0 |
| III-11 | E-163-1 | 0 | 0 | 0 |
| III-12 | E-163-K | 0 | 0 | 0 |
| IV-1 | AV-153 | 0.153 ± 0.216 | 2.005 ± 0.053 | 9.828 ± 0.081 |
| IV-2 | EE-126 | 0.856 ± 0.614 | 0.304 ± 0.078 | 0.292 ± 0.083 |
| IV-3 | E3-46 | 0.075 ± 0.163 | 1.542 ± 0.120 | >10 |
| IV-4 | V-6-55-1 | 0.636 ± 0.220 | 4.013 ± 0.228 | >10 |
| IV-5 | AV-154-Na | 0.481 ± 0.028 | 2.774 ± 0.199 | >10 |
| IV-6 | J-11-61B | 0 | 0.948 ± 0.056 | 8.222 ± 0 |
| IV-7 | J-11-71-2 | 0 | 3.160 ± 0.094 | >10 |
| IV-8 | Glutapyrone | 0.190 ± 0.015 | 0.127 ± 0.096 | 0 |
| V | J-12-25 | 0.016 ± 0.193 | 0.050 ± 0.040 | 0.012 ± 0.044 |
| Compounds | Trivial Name | Concentrations Tested | ||
|---|---|---|---|---|
| 10 µM | 100 µM | 1000 µM | ||
| Equivalent to µM H2O2 | ||||
| I | Diethone | 1.346 ± 1.904 | 0 | 0 |
| II-1 | Carbatone | 2.692 ± 0.000 | 18.846 ± 1.088 | 1.538 ± 0.544 |
| II-2 | J-9-133-2 | 0.250 ± 0.374 | 0 | 0 |
| II-3 | J-9-46 | 0 | 0 | 0 |
| II-4 | Metcarbatone | 0 | 0 | 0 |
| II-5 | Etcarbatone | 0 | 0 | 0 |
| II-6 | J-9-117 (Styrylcarbatone) | 0.577 ± 0.816 | 0 | 0.192 ± 0.272 |
| III-1 | Thiocarbatone | 0 | 0 | 0 |
| III-2 | TK-2 | 0 | 0 | 7.558 ± 1.791 |
| III-3 | E-170-4 (TK-1) | 0 | 3.375±1.768 | 38.529 ± 1.663 |
| III-4 | E2-134-1 | 0 | 0 | 0 |
| III-5 | E2-135 | 0 | 0 | 0 |
| III-6 | E2-136-2 | 0 | 0 | 0 |
| III-7 | E2-131 | 0 | 5.411 ± 1.663 | 6.588 ± 3.328 |
| III-8 | E2-130-3 | 0 | 0 | 0 |
| III-9 | E2-120 (ETK-2) | 0 | 0 | 13.235 ± 5.407 |
| III-10 | E2-113 (ETK-1) | 0 | 0 | 0.147±0.208 |
| III-11 | E-163-1 | 0 | 4.5 ± 1.179 | 13.059 ± 1.664 |
| III-12 | E-163-K | 0 | 0 | 0 |
| IV-1 | AV-153 | 6.923 ± 0.544 | 41.154 ± 1.088 | 5.00 ± 4.351 |
| IV-2 | EE-126 | 3.625 ± 0.884 | 3.938 ± 0.442 | 3.938 ± 0.442 |
| IV-3 | E3-46 | 0 | 7.243 ± 0.114 | 0 |
| IV-4 | V-6-55-1 | 0.481 ± 0.680 | 0 | 0 |
| IV-5 | AV-154-Na | 3.938 ± 0.442 | 3.625 ± 0.884 | 0 |
| IV-6 | J-11-61B | 0 | 0 | 0 |
| IV-7 | J-11-71-2 | 0.938 ± 0.000 | 1.875 ± 0.000 | 0 |
| IV-8 | Glutapyrone | 0 | 0 | 0 |
| V | J-12-25 | 0 | 0 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milkovic, L.; Vukovic, T.; Zarkovic, N.; Tatzber, F.; Bisenieks, E.; Kalme, Z.; Bruvere, I.; Ogle, Z.; Poikans, J.; Velena, A.; et al. Antioxidative 1,4-Dihydropyridine Derivatives Modulate Oxidative Stress and Growth of Human Osteoblast-Like Cells In Vitro. Antioxidants 2018, 7, 123. https://doi.org/10.3390/antiox7090123
Milkovic L, Vukovic T, Zarkovic N, Tatzber F, Bisenieks E, Kalme Z, Bruvere I, Ogle Z, Poikans J, Velena A, et al. Antioxidative 1,4-Dihydropyridine Derivatives Modulate Oxidative Stress and Growth of Human Osteoblast-Like Cells In Vitro. Antioxidants. 2018; 7(9):123. https://doi.org/10.3390/antiox7090123
Chicago/Turabian StyleMilkovic, Lidija, Tea Vukovic, Neven Zarkovic, Franz Tatzber, Egils Bisenieks, Zenta Kalme, Imanta Bruvere, Zaiga Ogle, Janis Poikans, Astrida Velena, and et al. 2018. "Antioxidative 1,4-Dihydropyridine Derivatives Modulate Oxidative Stress and Growth of Human Osteoblast-Like Cells In Vitro" Antioxidants 7, no. 9: 123. https://doi.org/10.3390/antiox7090123
APA StyleMilkovic, L., Vukovic, T., Zarkovic, N., Tatzber, F., Bisenieks, E., Kalme, Z., Bruvere, I., Ogle, Z., Poikans, J., Velena, A., & Duburs, G. (2018). Antioxidative 1,4-Dihydropyridine Derivatives Modulate Oxidative Stress and Growth of Human Osteoblast-Like Cells In Vitro. Antioxidants, 7(9), 123. https://doi.org/10.3390/antiox7090123