
Effect of Dietary Supplements with ω-3 Fatty Acids, Ascorbic Acid, and Polyphenolic Antioxidant Flavonoid on Gene Expression, Organ Failure, and Mortality in Endotoxemia-Induced Septic Rats


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Dietary Supplement Administration, Endotoxemia Induction, and Parameter Recordings
2.2. Primary Rat Mesenteric Endothelial Cell (RMEC) Isolation and mRNA Expression Determination by RT-qPCR
2.3. In Vivo Permeability Assay and MODS Determination
2.4. Statistical Analyses
3. Results
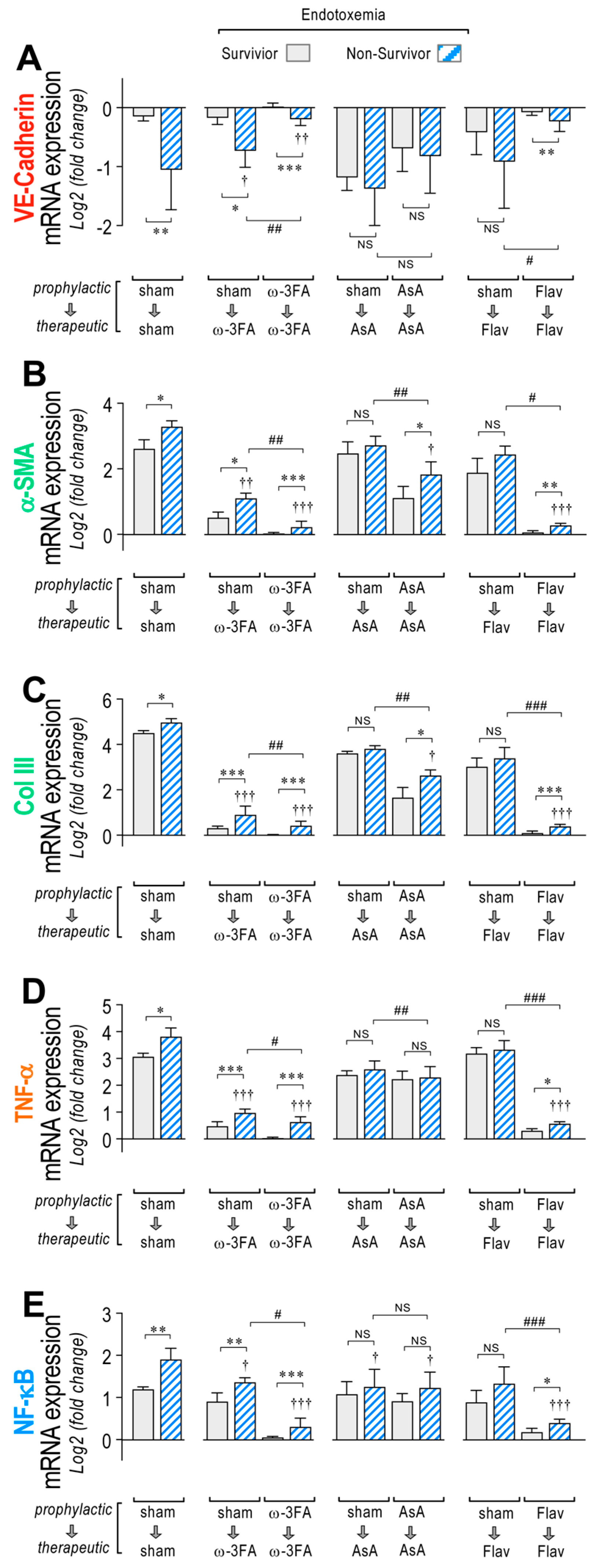
3.1. Impact in EndMT Gene Expression through Dietary Supplementation Based on ω-3 FA, Ascorbic Acid, or Polyphenolic Antioxidant Flavonoids in Endotoxemic Rats
3.2. Effects on Vascular Hyperpermeability through Dietary Supplementation Based on ω-3 FA, Ascorbic Acid, or Polyphenolic Antioxidant Flavonoids in Endotoxemic Rats
3.3. Effects on Endotoxemia-Induced Hypotension through Dietary Supplementation Based on ω-3 FA, Ascorbic Acid, or Polyphenolic Antioxidant Flavonoids in Endotoxemic Rats
3.4. Effects on Endotoxemia-Induced MODS through Dietary Supplementation Based on ω-3 FA, Ascorbic Acid, or Polyphenolic Antioxidant Flavonoids in Endotoxemic Rats
3.5. Impact on Survival and Risk of Death through Dietary Supplementation Based on ω-3 FA, Ascorbic Acid, or Polyphenolic Antioxidant Flavonoids in Endotoxemic Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, Regional, and National Sepsis Incidence and Mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet Lond. Engl. 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.; Rhodes, A.; Alhazzani, W.; Antonelli, M.; Coopersmith, C.M.; French, C.; Machado, F.R.; Mcintyre, L.; Ostermann, M.; Prescott, H.C.; et al. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock 2021. Crit. Care Med. 2021, 49, e1063–e1143. [Google Scholar] [CrossRef] [PubMed]
- Gatica, S.; Eltit, F.; Santibanez, J.F.; Varela, D.; Cabello-Verrugio, C.; Simon, F. Expression Suppression and Activity Inhibition of TRPM7 Regulate Cytokine Production and Multiple Organ Dysfunction Syndrome During Endotoxemia: A New Target for Sepsis. Curr. Mol. Med. 2019, 19, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, M.R. Clinical Studies on Cytokines in Sepsis: Role of Serum Cytokines in the Development of Multiple-Systems Organ Failure. Nephrol. Dial. Transplant. 1994, 9 (Suppl. S4), 94–98. [Google Scholar]
- Remick, D.G. Pathophysiology of Sepsis. Am. J. Pathol. 2007, 170, 1435–1444. [Google Scholar] [CrossRef]
- Riedemann, N.C.; Guo, R.-F.; Ward, P.A. The Enigma of Sepsis. J. Clin. Investig. 2003, 112, 460–467. [Google Scholar] [CrossRef]
- Karima, R.; Matsumoto, S.; Higashi, H.; Matsushima, K. The Molecular Pathogenesis of Endotoxic Shock and Organ Failure. Mol. Med. Today 1999, 5, 123–132. [Google Scholar] [CrossRef]
- Simon, F.; Fernández, R. Early Lipopolysaccharide-Induced Reactive Oxygen Species Production Evokes Necrotic Cell Death in Human Umbilical Vein Endothelial Cells. J. Hypertens. 2009, 27, 1202–1216. [Google Scholar] [CrossRef]
- Sarmiento, D.; Montorfano, I.; Cáceres, M.; Echeverría, C.; Fernández, R.; Cabello-Verrugio, C.; Cerda, O.; Tapia, P.; Simon, F. Endotoxin-Induced Vascular Endothelial Cell Migration Is Dependent on TLR4/NF-ΚB Pathway, NAD(P)H Oxidase Activation, and Transient Receptor Potential Melastatin 7 Calcium Channel Activity. Int. J. Biochem. Cell Biol. 2014, 55, 11–23. [Google Scholar] [CrossRef]
- Dauphinee, S.M.; Karsan, A. Lipopolysaccharide Signaling in Endothelial Cells. Lab. Investig. 2005, 86, 9–22. [Google Scholar] [CrossRef]
- Prado, Y.; Tapia, P.; Eltit, F.; Reyes-Martínez, C.; Feijóo, C.G.; Llancalahuen, F.M.; Riedel, C.A.; Cabello-Verrugio, C.; Stehberg, J.; Simon, F. Sepsis-Induced Coagulopathy Phenotype Induced by Oxidized High-Density Lipoprotein Associated with Increased Mortality in Septic-Shock Patients. Antioxidants 2023, 12, 543. [Google Scholar] [CrossRef]
- Echeverría, C.; Montorfano, I.; Sarmiento, D.; Becerra, A.; Nuñez-Villena, F.; Figueroa, X.F.; Cabello-Verrugio, C.; Elorza, A.A.; Riedel, C.; Simon, F. Lipopolysaccharide Induces a Fibrotic-like Phenotype in Endothelial Cells. J. Cell Mol. Med. 2013, 17, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Vallejos, A.; Olivares, P.; Gatica, S.; Villegas, V.; Echeverria, C.; Cabello-Verrugio, C.; Simon, F. Endotoxemia-Induced Endothelial Fibrosis Inhibition Improves Hypotension, Tachycardia, Multiple Organ Dysfunction Syndrome, Cytokine Response, Oxidative Stress, and Survival. Lab. Investig. 2019, 99, 1173–1192. [Google Scholar] [CrossRef]
- Echeverría, C.; Montorfano, I.; Hermosilla, T.; Armisén, R.; Velásquez, L.A.; Cabello-Verrugio, C.; Varela, D.; Simon, F. Endotoxin Induces Fibrosis in Vascular Endothelial Cells through a Mechanism Dependent on Transient Receptor Protein Melastatin 7 Activity. PLoS ONE 2014, 9, e94146. [Google Scholar] [CrossRef] [PubMed]
- Medici, D.; Shore, E.M.; Lounev, V.Y.; Kaplan, F.S.; Kalluri, R.; Olsen, B.R. Conversion of Vascular Endothelial Cells into Multipotent Stem-like Cells. Nat. Med. 2010, 16, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, E.M.; Potenta, S.; Xie, L.; Zeisberg, M.; Kalluri, R. Discovery of Endothelial to Mesenchymal Transition as a Source for Carcinoma-Associated Fibroblasts. Cancer Res. 2007, 67, 10123–10128. [Google Scholar] [CrossRef]
- Echeverría, C.; Montorfano, I.; Tapia, P.; Riedel, C.; Cabello-Verrugio, C.; Simon, F. Endotoxin-Induced Endothelial Fibrosis Is Dependent on Expression of Transforming Growth Factors Β1 and Β2. Infect. Immun. 2014, 82, 3678–3686. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Montorfano, I.; Becerra, A.; Cerro, R.; Echeverría, C.; Sáez, E.; Morales, M.G.; Fernández, R.; Cabello-Verrugio, C.; Simon, F. Oxidative Stress Mediates the Conversion of Endothelial Cells into Myofibroblasts via a TGF-Β1 and TGF-Β2-Dependent Pathway. Lab. Investig. 2014, 94, 1068–1082. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, C.; Montorfano, I.; Cabello-Verrugio, C.; Armisén, R.; Varela, D.; Simon, F. Suppression of Transient Receptor Potential Melastatin 4 Expression Promotes Conversion of Endothelial Cells into Fibroblasts via Transforming Growth Factor/Activin Receptor-like Kinase 5 Pathway. J. Hypertens. 2015, 33, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Tapia, P.; Gatica, S.; Cortés-Rivera, C.; Otero, C.; Becerra, A.; Riedel, C.A.; Cabello-Verrugio, C.; Kalergis, A.M.; Simon, F. Circulating Endothelial Cells from Septic Shock Patients Convert to Fibroblasts Are Associated with the Resuscitation Fluid Dose and Are Biomarkers for Survival Prediction. Crit. Care Med. 2019, 47, 942–950. [Google Scholar] [CrossRef]
- Rojas, M.; Prado, Y.; Tapia, P.; Carreño, L.J.; Cabello-Verrugio, C.; Simon, F. Oxidized High-Density Lipoprotein Induces Endothelial Fibrosis Promoting Hyperpermeability, Hypotension, and Increased Mortality. Antioxidants 2022, 11, 2469. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, E.M.; Tarnavski, O.; Zeisberg, M.; Dorfman, A.L.; McMullen, J.R.; Gustafsson, E.; Chandraker, A.; Yuan, X.; Pu, W.T.; Roberts, A.B.; et al. Endothelial-to-Mesenchymal Transition Contributes to Cardiac Fibrosis. Nat. Med. 2007, 13, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.; Muñoz-Durango, N.; Riedel, C.A.; Echeverría, C.; Kalergis, A.M.; Cabello-Verrugio, C.; Simon, F. Endothelial-to-Mesenchymal Transition: Cytokine-Mediated Pathways That Determine Endothelial Fibrosis under Inflammatory Conditions. Cytokine Growth Factor Rev. 2017, 33, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Becerra, A.; Rojas, M.; Vallejos, A.; Villegas, V.; Pérez, L.; Cabello-Verrugio, C.; Simon, F. Endothelial Fibrosis Induced by Suppressed STAT3 Expression Mediated by Signaling Involving the TGF-Β1/ALK5/Smad Pathway. Lab. Investig. 2017, 97, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento, D.; Montorfano, I.; Cerda, O.; Cáceres, M.; Becerra, A.; Cabello-Verrugio, C.; Elorza, A.A.; Riedel, C.; Tapia, P.; Velásquez, L.A.; et al. Increases in Reactive Oxygen Species Enhance Vascular Endothelial Cell Migration through a Mechanism Dependent on the Transient Receptor Potential Melastatin 4 Ion Channel. Microvasc. Res. 2015, 98, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.; Vallejos, A.; Echeverria, C.; Varela, D.; Cabello-Verrugio, C.; Simon, F. OxHDL Controls LOX-1 Expression and Plasma Membrane Localization through a Mechanism Dependent on NOX/ROS/NF-ΚB Pathway on Endothelial Cells. Lab. Investig. 2019, 99, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Tapia, P.; Chinchón, E.; Morales, D.; Stehberg, J.; Simon, F. Effectiveness of Short-Term 6-Hour High-Volume Hemofiltration during Refractory Severe Septic Shock. J. Trauma Acute Care Surg. 2012, 72, 1228–1238. [Google Scholar] [CrossRef]
- Barrionuevo, N.; Gatica, S.; Olivares, P.; Cabello-Verrugio, C.; Simon, F. Endothelial Cells Exhibit Two Waves of P-Selectin Surface Aggregation Under Endotoxic and Oxidative Conditions. Protein J. 2019, 38, 667–674. [Google Scholar] [CrossRef]
- Echeverria, C.; Eltit, F.; Santibanez, J.F.; Gatica, S.; Cabello-Verrugio, C.; Simon, F. Endothelial Dysfunction in Pregnancy Metabolic Disorders. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1866, 165414. [Google Scholar] [CrossRef]
- Vallejos, A.; Olivares, P.; Varela, D.; Echeverria, C.; Cabello-Verrugio, C.; Pérez-Leighton, C.; Simon, F. Preventive Leptin Administration Protects Against Sepsis Through Improving Hypotension, Tachycardia, Oxidative Stress Burst, Multiple Organ Dysfunction, and Increasing Survival. Front. Physiol. 2018, 9, 1800. [Google Scholar] [CrossRef]
- Chen, H.; Wang, S.; Zhao, Y.; Luo, Y.; Tong, H.; Su, L. Correlation Analysis of Omega-3 Fatty Acids and Mortality of Sepsis and Sepsis-Induced ARDS in Adults: Data from Previous Randomized Controlled Trials. Nutr. J. 2018, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Körner, A.; Schlegel, M.; Theurer, J.; Frohnmeyer, H.; Adolph, M.; Heijink, M.; Giera, M.; Rosenberger, P.; Mirakaj, V. Resolution of Inflammation and Sepsis Survival Are Improved by Dietary Ω-3 Fatty Acids. Cell Death Differ. 2018, 25, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Xie, Y.-J.; Liu, Z.-M.; Chen, W.-B.; Zhang, R.; Ye, H.-X.; Wang, W.; Liu, X.-Y.; Chen, H.-S. Omega-3 Fatty Acids Impair MiR-1-3p-Dependent Notch3 down-Regulation and Alleviate Sepsis-Induced Intestinal Injury. Mol. Med. 2022, 28, 9. [Google Scholar] [CrossRef] [PubMed]
- Kawade, N.; Tokuda, Y.; Tsujino, S.; Aoyama, H.; Kobayashi, M.; Murai, A.; Horio, F. Dietary Intake of Ascorbic Acid Attenuates Lipopolysaccharide-Induced Sepsis and Septic Inflammation in ODS Rats. J. Nutr. Sci. Vitaminol. 2018, 64, 404–411. [Google Scholar] [CrossRef]
- Marik, P.E. Hydrocortisone, Ascorbic Acid and Thiamine (HAT Therapy) for the Treatment of Sepsis. Focus on Ascorbic Acid. Nutrients 2018, 10, 1762. [Google Scholar] [CrossRef]
- Rungsung, S.; Singh, T.U.; Perumalraja, K.; Mahobiya, A.; Sharma, M.; Lingaraju, M.C.; Parida, S.; Sahoo, M.; Kumar, D. Luteolin Alleviates Vascular Dysfunctions in CLP-Induced Polymicrobial Sepsis in Mice. Pharmacol. Rep. 2022, 74, 1054–1068. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Liu, L.; Wang, Z.; Xie, H.; Feng, Y.; Suo, J.; Wang, M.; Shang, W.; Feng, G. Icariin Improves Sepsis-Induced Mortality and Acute Kidney Injury. Pharmacology 2018, 102, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, H.; Wu, X.; Guo, Y.; Cheng, W.; Qian, F. Fisetin Alleviates Sepsis-Induced Multiple Organ Dysfunction in Mice via Inhibiting P38 MAPK/MK2 Signaling. Acta Pharmacol. Sin. 2020, 41, 1348–1356. [Google Scholar] [CrossRef]
- Tu, T.; Li, B.; Li, X.; Zhang, B.; Xiao, Y.; Li, J.; Qin, F.; Liu, N.; Sun, C.; Liu, Q.; et al. Dietary ω-3 Fatty Acids Reduced Atrial Fibrillation Vulnerability via Attenuating Myocardial Endoplasmic Reticulum Stress and Inflammation in a Canine Model of Atrial Fibrillation. J. Cardiol. 2022, 79, 194–201. [Google Scholar] [CrossRef]
- Singh, J.E. Dietary Sources of Omega-3 Fatty Acids Versus Omega-3 Fatty Acid Supplementation Effects on Cognition and Inflammation. Curr. Nutr. Rep. 2020, 9, 264–277. [Google Scholar] [CrossRef]
- Belin, S.; Kaya, F.; Burtey, S.; Fontes, M. Ascorbic Acid and Gene Expression: Another Example of Regulation of Gene Expression by Small Molecules? Curr. Genom. 2010, 11, 52–57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hitomi, K.; Tsukagoshi, N. Subcellular Biochemistry, Ascorbic Acid: Biochemistry and Biomedical Cell Biology. Subcell. Biochem. 1996, 25, 41–56. [Google Scholar] [PubMed]
- Grano, A.; Tullio, M.C.D. Ascorbic Acid as a Sensor of Oxidative Stress and a Regulator of Gene Expression: The Yin and Yang of Vitamin C. Med. Hypotheses 2007, 69, 953–954. [Google Scholar] [CrossRef] [PubMed]
- Chojkier, M.; Houglum, K.; Solis-Herruzo, J.; Brenner, D.A. Stimulation of Collagen Gene Expression by Ascorbic Acid in Cultured Human Fibroblasts. A Role for Lipid Peroxidation? J. Biol. Chem. 1989, 264, 16957–16962. [Google Scholar] [CrossRef] [PubMed]
- Sawaguchi, S.; Yue, B.Y.; Chang, I.L.; Wong, F.; Higginbotham, E.J. Ascorbic Acid Modulates Collagen Type I Gene Expression by Cells from an Eye Tissue—Trabecular Meshwork. Cell. Mol. Biol. 1992, 38, 587–604. [Google Scholar] [PubMed]
- Martín, M.Á.; Ramos, S. Impact of Dietary Flavanols on Microbiota, Immunity and Inflammation in Metabolic Diseases. Nutrients 2021, 13, 850. [Google Scholar] [CrossRef]
- El-Haleim, E.A.A.; Sallam, N.A. Vitamin D Modulates Hepatic MicroRNAs and Mitigates Tamoxifen-induced Steatohepatitis in Female Rats. Fundam. Clin. Pharm. 2022, 36, 338–349. [Google Scholar] [CrossRef]
- Jenks, D.B.; Ehrlich, Y.; Spolnik, K.; Gregory, R.L.; Yassen, G.H. Residual Antibiofilm Effects of Various Concentrations of Double Antibiotic Paste Used during Regenerative Endodontics after Different Application Times. Arch. Oral Biol. 2016, 70, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Khazri, O.; Mezni, A.; Limam, F.; Aouani, E. Bleomycin-Induced Damage in Rat Lung: Protective Effect of Grape Seed and Skin Extract. Dose Response 2022, 20, 15593258221131648. [Google Scholar] [CrossRef]
- Gatica, S.; Villegas, V.; Vallejos, A.; Olivares, P.; Aballai, V.; Lagos-Meza, F.; Echeverria, C.; Cabello-Verrugio, C.; Varela, D.; Simon, F. TRPM7 Mediates Kidney Injury, Endothelial Hyperpermeability and Mortality during Endotoxemia. Lab. Investig. 2019, 100, 234–249. [Google Scholar] [CrossRef]
- Prado, Y.; Pérez, L.; Eltit, F.; Echeverría, C.; Llancalahuen, F.M.; Tapia, P.; González, P.A.; Kalergis, A.M.; Cabello-Verrugio, C.; Simon, F. Procoagulant Phenotype Induced by Oxidized High-Density Lipoprotein Associates with Acute Kidney Injury and Death. Thromb. Res. 2023, 223, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Semmler, A.; Frisch, C.; Debeir, T.; Ramanathan, M.; Okulla, T.; Klockgether, T.; Heneka, M.T. Long-Term Cognitive Impairment, Neuronal Loss and Reduced Cortical Cholinergic Innervation after Recovery from Sepsis in a Rodent Model. Exp. Neurol. 2007, 204, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Leite, F.B.; Prediger, R.D.; Silva, M.V.; Sousa, J.B.D.; Carneiro, F.P.; Gasbarri, A.; Tomaz, C.; Queiroz, A.J.; Martins, N.T.; Ferreira, V.M. Role of Nicotine on Cognitive and Behavioral Deficits in Sepsis-Surviving Rats. Brain Res. 2013, 1507, 74–82. [Google Scholar] [CrossRef]
- Besseling, P.J.; Pieters, T.T.; Nguyen, I.T.N.; Bree, P.M.D.; Willekes, N.; Dijk, A.H.; Bovée, D.M.; Hoorn, E.J.; Rookmaaker, M.B.; Gerritsen, K.G.; et al. A Plasma Creatinine- and Urea-Based Equation to Estimate Glomerular Filtration Rate in Rats. Am. J. Physiol. Renal Physiol. 2021, 320, F518–F524. [Google Scholar] [CrossRef] [PubMed]
- Roche, H.M. Dietary Lipids and Gene Expression. Biochem. Soc. Trans. 2004, 32, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Clarke, S.D.; Heird, W.C. Polyunsaturated Fatty Acids and Gene Expression. Curr. Opin. Clin. Nutr. 2004, 7, 151–156. [Google Scholar] [CrossRef]
- Wahle, K.W.J.; Rotondo, D.; Heys, S.D. Polyunsaturated Fatty Acids and Gene Expression in Mammalian Systems. Proc. Nutr. Soc. 2003, 62, 349–360. [Google Scholar] [CrossRef]
- Corral-Jara, K.F.; Cantini, L.; Poupin, N.; Ye, T.; Rigaudière, J.P.; Vincent, S.D.S.; Pinel, A.; Morio, B.; Capel, F. An Integrated Analysis of MiRNA and Gene Expression Changes in Response to an Obesogenic Diet to Explore the Impact of Transgenerational Supplementation with Omega 3 Fatty Acids. Nutrients 2020, 12, 3864. [Google Scholar] [CrossRef]
- Poulsen, H.E.; Jensen, B.R.; Weimann, A.; Jensen, S.A.; Sørensen, M.; Loft, S. Antioxidants, DNA Damage and Gene Expression. Free Radical Res. 2000, 33, S33–S39. [Google Scholar]
- Palmer, H.J.; Paulson, K.E. Reactive Oxygen Species and Antioxidants in Signal Transduction and Gene Expression. Nutr. Rev. 2009, 55, 353–361. [Google Scholar] [CrossRef]
- Feng, N.-H.; Chu, S.-J.; Wang, D.; Hsu, K.; Lin, C.-H.; Lin, H.-I. Effects of Various Antioxidants on Endotoxin-Induced Lung Injury and Gene Expression: MRNA Expressions of MnSOD, Interleukin-1beta and INOS. Chin. J. Physiol. 2004, 47, 111–120. [Google Scholar] [PubMed]
- Abdelgawad, L.M.; Abdelaziz, A.M.; Sabry, D.; Abdelgwad, M. Influence of Photobiomodulation and Vitamin D on Osteoblastic Differentiation of Human Periodontal Ligament Stem Cells and Bone-like Tissue Formation through Enzymatic Activity and Gene Expression. Biomol. Concepts 2020, 11, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Molano, A.; Meydani, S.N. Vitamin E, Signalosomes and Gene Expression in T Cells. Mol. Asp. Med. 2012, 33, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Adolfsson, O.; Lee, C.; Prolla, T.A.; Ordovas, J.; Meydani, S.N. Vitamin E and Gene Expression in Immune Cells. Ann. N. Y. Acad. Sci. 2004, 1031, 96–101. [Google Scholar] [CrossRef]
- Kao, P.-F.; Lee, W.-S.; Liu, J.-C.; Chan, P.; Tsai, J.-C.; Hsu, Y.-H.; Chang, W.-Y.; Cheng, T.-H.; Liao, S.-S. Downregulation of Superoxide Dismutase Activity and Gene Expression in Cultured Rat Brain Astrocytes after Incubation with Vitamin C. Pharmacology 2003, 69, 1–6. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Parra, D.; Milagro, F.I.; Goyenechea, E.; Larrarte, E.; Margareto, J.; Martnez, J.A. Differential Expression of Oxidative Stress and Inflammation Related Genes in Peripheral Blood Mononuclear Cells in Response to a Low-Calorie Diet: A Nutrigenomics Study. Omics J. Integr. Biol. 2008, 12, 251–261. [Google Scholar] [CrossRef]
- Ulven, S.M.; Holven, K.B. Metabolomic and Gene Expression Analysis to Study the Effects of Dietary Saturated and Polyunsaturated Fats. Curr. Opin. Lipidol. 2020, 31, 15–19. [Google Scholar] [CrossRef]
- Islam, M.S.; Castellucci, C.; Fiorini, R.; Greco, S.; Gagliardi, R.; Zannotti, A.; Giannubilo, S.R.; Ciavattini, A.; Frega, N.G.; Pacetti, D.; et al. Omega-3 Fatty Acids Modulate the Lipid Profile, Membrane Architecture, and Gene Expression of Leiomyoma Cells. J. Cell. Physiol. 2018, 233, 7143–7156. [Google Scholar] [CrossRef]
- Kemse, N.; Kale, A.; Chavan-Gautam, P.; Joshi, S. Increased Intake of Vitamin B 12, Folate, and Omega-3 Fatty Acids to Improve Cognitive Performance in Offspring Born to Rats with Induced Hypertension during Pregnancy. Food Funct. 2018, 9, 3872–3883. [Google Scholar] [CrossRef]
- Kemse, N.; Sundrani, D.; Kale, A.; Joshi, S. Maternal Micronutrients, Omega-3 Fatty Acids and Gene Expression of Angiogenic and Inflammatory Markers in Pregnancy Induced Hypertension Rats. Arch. Med. Res. 2017, 48, 414–422. [Google Scholar] [CrossRef]
- Makris, G.C.; Geroulakos, G.; Makris, M.C.; Mikhailidis, D.P.; Falagas, M.E. The Pleiotropic Effects of Statins and Omega-3 Fatty Acids against Sepsis: A New Perspective. Expert Opin. Investig. Drugs 2010, 19, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Svahn, S.L.; Ulleryd, M.A.; Grahnemo, L.; Ståhlman, M.; Borén, J.; Nilsson, S.; Jansson, J.-O.; Johansson, M.E. Dietary Omega-3 Fatty Acids Increase Survival and Decrease Bacterial Load in Mice Subjected to Staphylococcus Aureus-Induced Sepsis. Infect. Immun. 2016, 84, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Wolbrink, D.R.J.; Grundsell, J.R.; Witteman, B.; van de Poll, M.; van Santvoort, H.C.; Issa, E.; Dennison, A.; van Goor, H.; Besselink, M.G.; Bouwense, S.A.W.; et al. Are Omega-3 Fatty Acids Safe and Effective in Acute Pancreatitis or Sepsis? A Systematic Review and Meta-Analysis. Clin. Nutr. 2020, 39, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- Barton, R.G.; Wells, C.L.; Carlson, A.; Singh, R.; Sullivan, J.J.; Cerra, F.B. Dietary Omega-3 Fatty Acids Decrease Mortality and Kupffer Cell Prostaglandin E2 Production in a Rat Model of Chronic Sepsis. J. Trauma Inj. Infect. Crit. Care 1991, 31, 768–774. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Lee, S.-M. Effect of Ascorbic Acid on Hepatic Vasoregulatory Gene Expression during Polymicrobial Sepsis. Life Sci. 2004, 75, 2015–2026. [Google Scholar] [CrossRef]
- Fowler, A.A.; Syed, A.A.; Knowlson, S.; Sculthorpe, R.; Farthing, D.; DeWilde, C.; Farthing, C.A.; Larus, T.L.; Martin, E.; Brophy, D.F.; et al. Phase I Safety Trial of Intravenous Ascorbic Acid in Patients with Severe Sepsis. J. Transl. Med. 2014, 12, 32. [Google Scholar] [CrossRef]
- Schmidt, T.; Kahn, R.; Kahn, F. Ascorbic Acid Attenuates Activation and Cytokine Production in Sepsis-like Monocytes. J. Leukoc. Biol. 2022, 112, 491–498. [Google Scholar] [CrossRef]
- Fisher, B.J.; Kraskauskas, D.; Martin, E.J.; Farkas, D.; Wegelin, J.A.; Brophy, D.; Ward, K.R.; Voelkel, N.F.; Fowler, A.A.; Natarajan, R. Mechanisms of Attenuation of Abdominal Sepsis Induced Acute Lung Injury by Ascorbic Acid. Am. J. Physiol.-Lung Cell. 2012, 303, L20–L32. [Google Scholar] [CrossRef]
- Kim, J.; Stolarski, A.; Zhang, Q.; Wee, K.; Remick, D. Hydrocortisone, ascorbic acid, and thiamine therapy decrease renal oxidative stress and acute kidney injury in murine sepsis. Shock 2022, 58, 426–433. [Google Scholar] [CrossRef]
- Ragoonanan, D.; Tran, N.; Modi, V.; Nickelsen, P.M. Unanswered Questions on the Use of Hydrocortisone, Ascorbic Acid, and Thiamine Therapy in Sepsis and Septic Shock. Am. J. Health Syst. Pharm. 2022, 79, 1626–1633. [Google Scholar] [CrossRef]
- Wang, K.; Yin, L.; Song, Y.; Zhang, M.; Lu, Y.; Wang, S. The Use of Hydrocortisone, Ascorbic Acid and Thiamine in Patients with Sepsis and Septic Shock—A Systematic Review. J. Pharm. Pract. 2022, 35, 89719002210971. [Google Scholar] [CrossRef] [PubMed]
- Assouline, B.; Faivre, A.; Verissimo, T.; Sangla, F.; Berchtold, L.; Giraud, R.; Bendjelid, K.; Sgardello, S.; Elia, N.; Pugin, J.; et al. Thiamine, Ascorbic Acid, and Hydrocortisone as a Metabolic Resuscitation Cocktail in Sepsis: A Meta-Analysis of Randomized Controlled Trials with Trial Sequential Analysis. Crit. Care Med. 2021, 49, 2112–2120. [Google Scholar] [CrossRef] [PubMed]
- Vetterli, L.; Brun, T.; Giovannoni, L.; Bosco, D.; Maechler, P. Resveratrol Potentiates Glucose-Stimulated Insulin Secretion in INS-1E β-Cells and Human Islets through a SIRT1-Dependent Mechanism. J. Biol. Chem. 2011, 286, 6049–6060. [Google Scholar] [CrossRef] [PubMed]
- Scazzocchio, B.; Varì, R.; Filesi, C.; D’Archivio, M.; Santangelo, C.; Giovannini, C.; Iacovelli, A.; Silecchia, G.; Volti, G.L.; Galvano, F.; et al. Cyanidin-3-O-β-Glucoside and Protocatechuic Acid Exert Insulin-Like Effects by Upregulating PPARγ Activity in Human Omental Adipocytes. Diabetes 2011, 60, 2234–2244. [Google Scholar] [CrossRef]
- Esser, D.; Geleijnse, J.M.; Matualatupauw, J.C.; Dower, J.I.; Kromhout, D.; Hollman, P.C.H.; Afman, L.A. Pure Flavonoid Epicatechin and Whole Genome Gene Expression Profiles in Circulating Immune Cells in Adults with Elevated Blood Pressure: A Randomised Double-Blind, Placebo-Controlled, Crossover Trial. PLoS ONE 2018, 13, e0194229. [Google Scholar] [CrossRef] [PubMed]
- Uzunallı, G.; Bora-Tatar, G.; Dayangaç-Erden, D.; Erdem-Yurter, H. Effects of Flavonoid Quercetin on Survival of Motor Neuron Gene Expression. Cell Biol. Int. 2015, 39, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Krishnan, M.; Kim, M.; Yoon, Y.K.; Kim, Y. Rhamnetin, a Natural Flavonoid, Ameliorates Organ Damage in a Mouse Model of Carbapenem-Resistant Acinetobacter Baumannii-Induced Sepsis. Int. J. Mol. Sci. 2022, 23, 12895. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Rong, Y.; Wang, Y.; Guo, Y.; Shan, L.; Yu, X.; Li, L.; Si, J.; Li, X.; Ma, K. A Systematic Study of the Mechanism of Acacetin Against Sepsis Based on Network Pharmacology and Experimental Validation. Front. Pharmacol. 2021, 12, 683645. [Google Scholar] [CrossRef]
- Rabha, D.J.; Singh, T.U.; Rungsung, S.; Kumar, T.; Parida, S.; Lingaraju, M.C.; Paul, A.; Sahoo, M.; Kumar, D. Kaempferol Attenuates Acute Lung Injury in Caecal Ligation and Puncture Model of Sepsis in Mice. Exp. Lung Res. 2018, 44, 63–78. [Google Scholar] [CrossRef]
- Bösch, F.; Altendorf-Hofmann, A.; Koliogiannis, V.; Ilhan, H.; Jacob, S.; Pretzsch, E.; Nölting, S.; Werner, J.; Klauschen, F.; Auernhammer, C.J.; et al. Immuno-Histochemical Correlation of Fibrosis-Related Markers with the Desmoplastic Reaction of the Mesentery in Small Intestine Neuroendocrine Neoplasms. J. Cancer Res. Clin. 2022, 148, 1–9. [Google Scholar] [CrossRef]
- Chaves, F.J.; Cruz, I.; Lopes, C.; Morais, M.D. Polyposis Coli Associated with Fibrosis of Mesentery, Mesocolon and Retroperitoneal Tissues: A Rare Variant of Gardner’s Syndrome. Am. J. Gastroenterol. 1976, 65, 163–167. [Google Scholar] [PubMed]
- Meng, J.; Mao, Y.; Zhou, J.; Chen, Z.; Huang, S.; Wang, Y.; Huang, L.; Zhang, R.; Shen, X.; Lv, W.; et al. Mesenteric Abnormalities Play an Important Role in Grading Intestinal Fibrosis in Patients with Crohn’s Disease: A Computed Tomography and Clinical Marker-Based Nomogram. Ther. Adv. Gastroenterol. 2022, 15, 17562848221122504. [Google Scholar] [CrossRef] [PubMed]
- Vacek, L.; Bravený, P. Effect of Angiotensin II on Blood Pressure and on Microvascular Beds in Mesentery, Skin, and Skeletal Muscle of the Rat. Microvasc. Res. 1978, 16, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Nagy, J.A.; Manseau, E.J.; Dvorak, H.F. Vascular Permeability Factor/Vascular Endothelial Growth Factor-Mediated Signaling in Mouse Mesentery Vascular Endothelium. Cancer Res. 1998, 58, 1278–1284. [Google Scholar]
- Aird, W.C. Endothelium as an Organ System. Crit. Care Med. 2004, 32, S271–S279. [Google Scholar] [CrossRef]
- Lee, W.L.; Slutsky, A.S. Sepsis and Endothelial Permeability. N. Engl. J. Med. 2010, 363, 689–691. [Google Scholar] [CrossRef]
- Siddall, E.; Khatri, M.; Radhakrishnan, J. Capillary Leak Syndrome: Etiologies, Pathophysiology, and Management. Kidney Int. 2017, 92, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Spicer, A.; Calfee, C.S. Fixing the Leak: Targeting the Vascular Endothelium in Sepsis. Crit. Care 2012, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Wang, D.; Kong, L.; Li, M.; Feng, Z.; Shuai, B.; Wang, L.; Wei, Y.; Li, H.; Wu, S.; et al. Intermedin Protects against Sepsis by Concurrently Re-Establishing the Endothelial Barrier and Alleviating Inflammatory Responses. Nat. Commun. 2018, 9, 2644. [Google Scholar] [CrossRef]
- Liu, J.-Y.; Dickter, J.K. Nosocomial Infections A History of Hospital-Acquired Infections. Gastrointest. Endosc. Clin. N. Am. 2020, 30, 637–652. [Google Scholar] [CrossRef]
- Tan, X.Y.D.; Wiseman, T.; Betihavas, V. Risk Factors for Nosocomial Infections and/or Sepsis in Adult Burns Patients: An Integrative Review. Intensive Crit. Care Nurs. 2022, 73, 103292. [Google Scholar] [CrossRef] [PubMed]
- Preiser, J.-C.; Arabi, Y.M.; Berger, M.M.; Casaer, M.; McClave, S.; Montejo-González, J.C.; Peake, S.; Blaser, A.R.; Berghe, G.V.D.; van Zanten, A.; et al. A Guide to Enteral Nutrition in Intensive Care Units: 10 Expert Tips for the Daily Practice. Crit. Care 2021, 25, 424. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.; Hoffman, L. Enteral Nutrition in the Mechanically Ventilated Patient. Nutr. Clin. Pract. 2019, 34, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.; Gokorsch, S.; Fegbeutel, C.; Hattar, K.; Rosseau, S.; Walmrath, D.; Seeger, W.; Grimminger, F. Parenteral Nutrition with Fish Oil Modulates Cytokine Response in Patients with Sepsis. Am. J. Resp. Crit. Care 2003, 167, 1321–1328. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Yang, E.; Zhang, N.; Cao, S.; Zhou, Y. Fish Oil–Supplemented Parenteral Nutrition Could Alleviate Acute Lung Injury, Modulate Immunity, and Reduce Inflammation in Rats with Abdominal Sepsis. Nutr. Res. 2015, 35, 784–791. [Google Scholar] [CrossRef]
- Sukhotnik, I.; Shany, A.; Bashenko, Y.; Hayari, L.; Chemodanov, E.; Mogilner, J.; Coran, A.G.; Shaoul, R. Parenteral but Not Enteral Omega-3 Fatty Acids (Omegaven) Modulate Intestinal Regrowth After Massive Small Bowel Resection in Rats. JPEN J. Parenter. Enter. Nutr. 2010, 34, 503–512. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prado, Y.; Echeverría, C.; Feijóo, C.G.; Riedel, C.A.; Cabello-Verrugio, C.; Santibanez, J.F.; Simon, F. Effect of Dietary Supplements with ω-3 Fatty Acids, Ascorbic Acid, and Polyphenolic Antioxidant Flavonoid on Gene Expression, Organ Failure, and Mortality in Endotoxemia-Induced Septic Rats. Antioxidants 2023, 12, 659. https://doi.org/10.3390/antiox12030659
Prado Y, Echeverría C, Feijóo CG, Riedel CA, Cabello-Verrugio C, Santibanez JF, Simon F. Effect of Dietary Supplements with ω-3 Fatty Acids, Ascorbic Acid, and Polyphenolic Antioxidant Flavonoid on Gene Expression, Organ Failure, and Mortality in Endotoxemia-Induced Septic Rats. Antioxidants. 2023; 12(3):659. https://doi.org/10.3390/antiox12030659
Chicago/Turabian StylePrado, Yolanda, Cesar Echeverría, Carmen G. Feijóo, Claudia A. Riedel, Claudio Cabello-Verrugio, Juan F. Santibanez, and Felipe Simon. 2023. "Effect of Dietary Supplements with ω-3 Fatty Acids, Ascorbic Acid, and Polyphenolic Antioxidant Flavonoid on Gene Expression, Organ Failure, and Mortality in Endotoxemia-Induced Septic Rats" Antioxidants 12, no. 3: 659. https://doi.org/10.3390/antiox12030659
APA StylePrado, Y., Echeverría, C., Feijóo, C. G., Riedel, C. A., Cabello-Verrugio, C., Santibanez, J. F., & Simon, F. (2023). Effect of Dietary Supplements with ω-3 Fatty Acids, Ascorbic Acid, and Polyphenolic Antioxidant Flavonoid on Gene Expression, Organ Failure, and Mortality in Endotoxemia-Induced Septic Rats. Antioxidants, 12(3), 659. https://doi.org/10.3390/antiox12030659