Musical Training and Brain Volume in Older Adults

 ,
,
Abstract
1. Introduction
2. Method
2.1. Participants
2.2. Music Questionnaire
2.3. Active Engagement
- Twenty items;
- Active musical engagement behaviors (e.g., “I keep track of new music that I come across”, “I often read or search the internet for things related to music”);
- Deliberate allocation of time and money on musical activities (e.g., “I don’t spend much of my disposable income on music”, “I listen attentively to music for _ hours per day”).
2.4. Perceptual Abilities
- Fifteen items;
- Self-assessment of a cognitive musical ability, most of them related to musical listening skills;
- Music listening skills (e.g., “I can compare and discuss differences between two performances or versions of a musical piece”, “I can tell when people sing or play out of tune”).
2.5. Musical Training
- Eleven items;
- Extent of musical training and practice (e.g., “I engaged in regular daily practice of a musical instrument including voice for __ years”, “At the peak of my interest I practiced on my primary instrument including voice for __ hours per day”);
- Degree of self-assessed musicianship (“I would not consider myself a musician”, “I have never been complimented for my talents as a musical performer”).
2.6. Singing Abilities
- Seven items
- Skills and activities related to singing (e.g., “After hearing a new song two or three times I can usually sing it by myself”, “I am not able to sing in harmony when somebody is singing a familiar tune”).
2.7. Emotions
- Nine items;
- Mainly active behaviors related to emotional responses to music (e.g., “I am able to talk about the emotions that a piece of music evokes in me”, “I sometimes choose music that can trigger shivers down my spine”).
2.8. Brain Structure
2.9. Statistical Analysis
3. Results
3.1. Music, Age, Sex, and Education
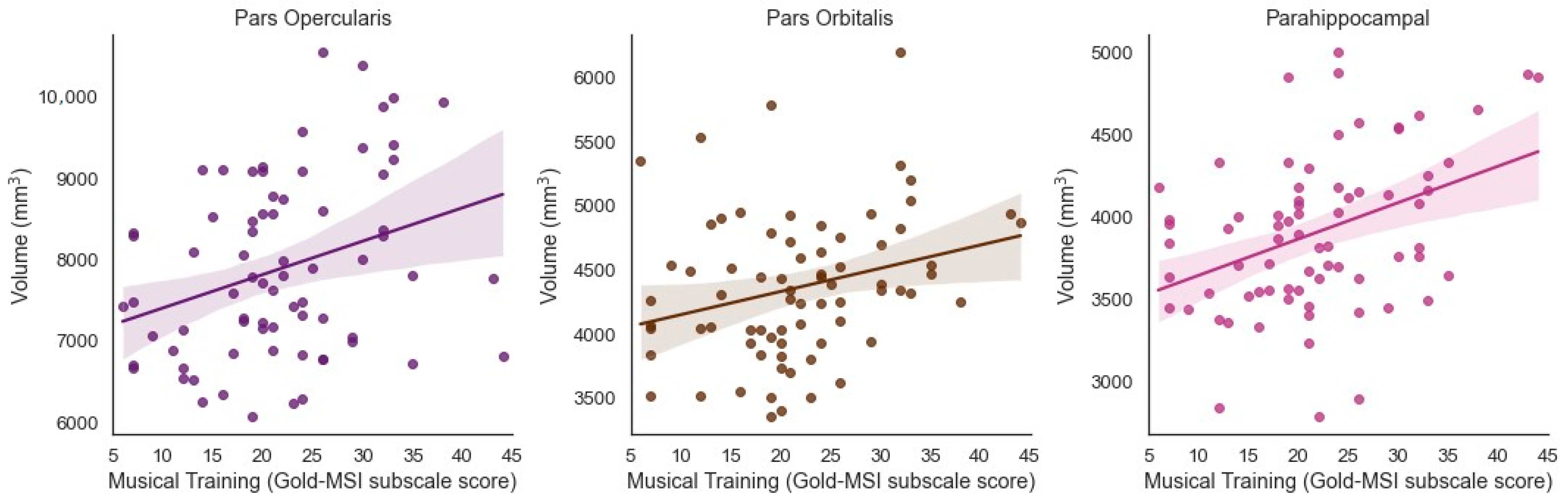
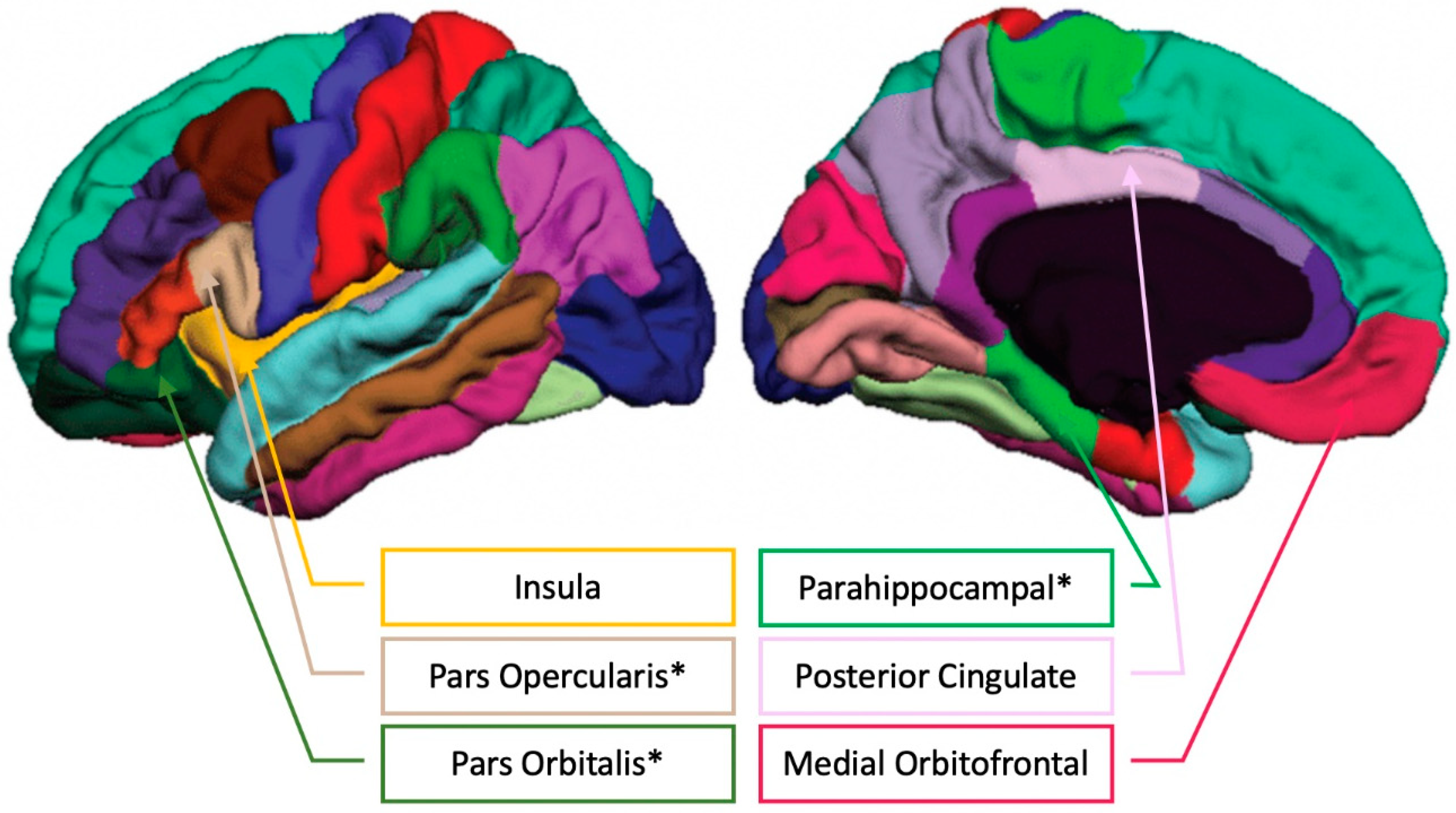
3.2. Musical Training and Bilateral Brain Volume
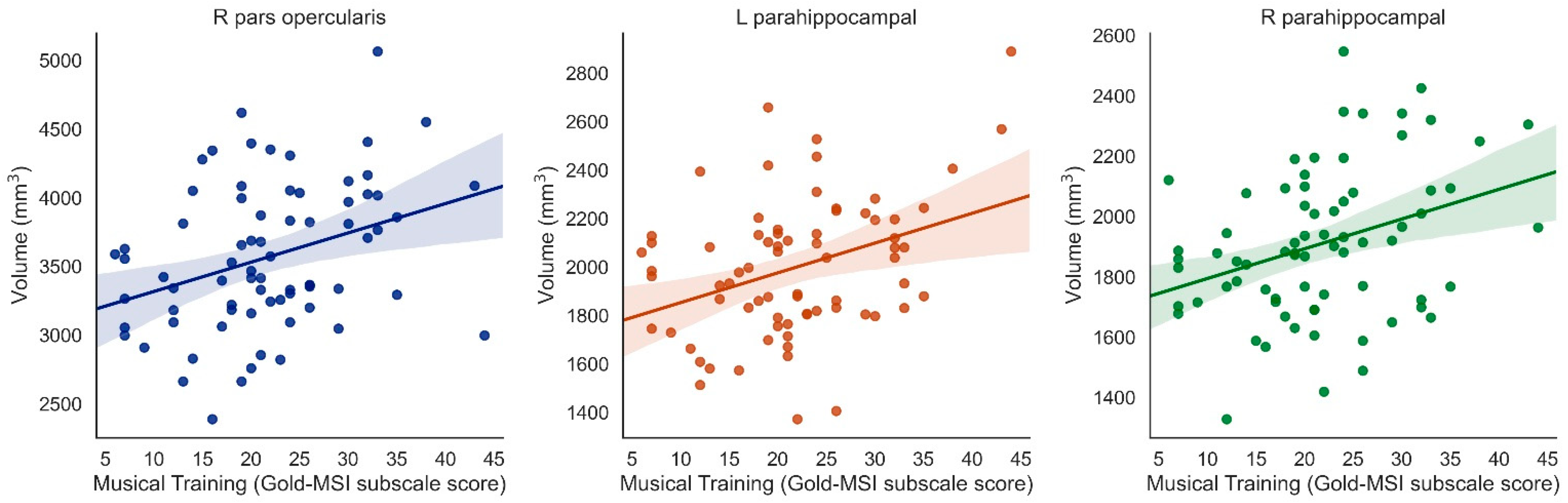
3.3. Laterality of Results
3.4. Exploratory Analyses (Uncorrected, p < 0.05)
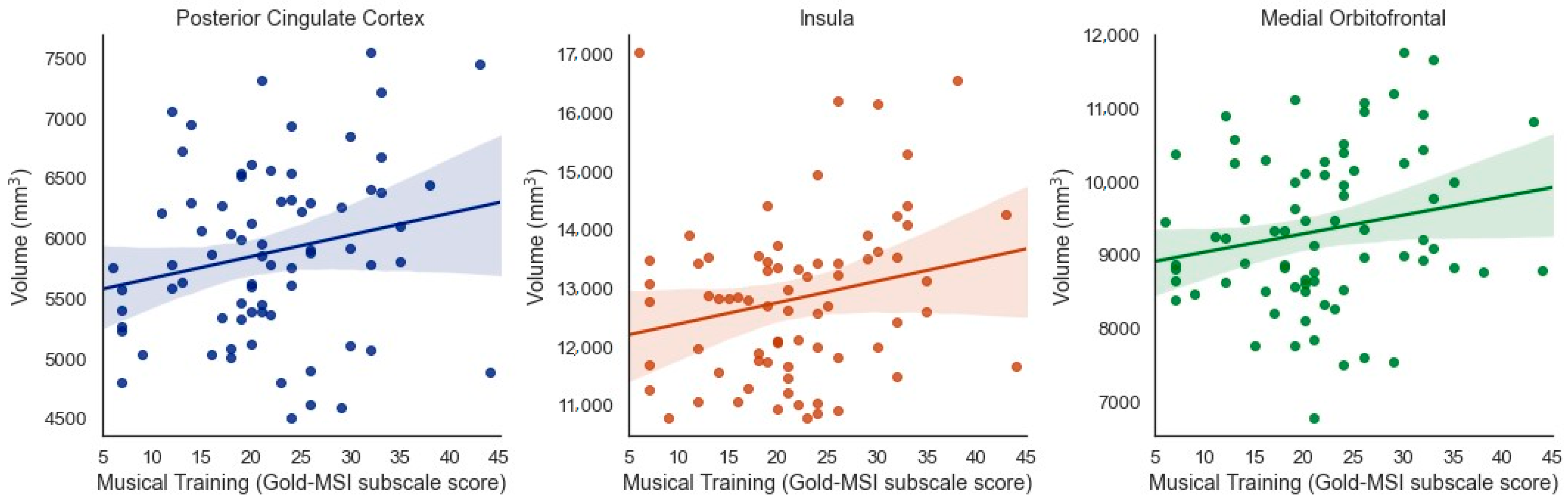
3.5. Specificity of Results
4. Discussion
4.1. Music and Language
4.2. Music, Memory, and Executive Function
4.3. Music, Emotion, and Reward
4.4. Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borella, E.; Carretti, B.; De Beni, R. Working memory and inhibition across the adult life-span. Acta Psychol. 2008, 128, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Grady, C.L. The cognitive neuroscience of ageing. Nat. Rev. Neurosci. 2012, 13, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Hedman, A.M.; Van Haren, N.E.; Schnack, H.G.; Kahn, R.S.; Pol, H.E.H. Human brain changes across the life span: A review of 56 longitudinal magnetic resonance imaging studies. Hum. Brain Mapp. 2012, 33, 1987–2002. [Google Scholar] [CrossRef] [PubMed]
- Reuter-Lorenz, P.A.; Park, D.C. Human Neuroscience and the Aging Mind: A New Look at Old Problems. J. Gerontol. Ser. B 2010, 65, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T.A. Influence of age on practice effects in longitudinal neurocognitive change. Neuropsychology 2010, 24, 563–572. [Google Scholar] [CrossRef]
- Allen, J.S.; Bruss, J.; Brown, C.K.; Damasio, H. Normal neuroanatomical variation due to age: The major lobes and a parcellation of the temporal region. Neurobiol. Aging 2005, 26, 1245–1260. [Google Scholar] [CrossRef]
- Resnick, S.M.; Pham, D.L.; Kraut, M.A.; Zonderman, A.B.; Davatzikos, C. Longitudinal Magnetic Resonance Imaging Studies of Older Adults: A Shrinking Brain. J. Neurosci. 2003, 23, 3295–3301. [Google Scholar] [CrossRef]
- Storsve, A.B.; Fjel, A.M.; Tamnes, C.K.; Westlye, L.T.; Overbye, K.; Aasland, H.W. Differential longitudinal changes in cortical thickness, surface area and volume across the adult life span: Regions of accelerating and decelerating change. J. Neurosci. 2014, 34, 8488–8498. [Google Scholar] [CrossRef]
- Walhovd, K.B.; Westlye, L.T.; Amlien, I.; Espeseth, T.; Reinvang, I.; Raz, N.; Agartz, I.; Salat, D.H.; Greve, D.N.; Fischl, B.; et al. Consistent neuroanatomical age-related volume differences across multiple samples. Neurobiol. Aging 2011, 32, 916–932. [Google Scholar] [CrossRef]
- Wilson, R.S.; Beckett, L.A.; Barnes, L.L.; Schneider, J.A.; Bach, J.; Evans, D.A.; Bennett, D.A. Individual differences in rates of change in cognitive abilities of older persons. Psychol. Aging 2002, 17, 179–193. [Google Scholar] [CrossRef]
- Kolb, B.; Gibb, R.; Gorny, G. Experience-dependent changes in dendritic arbor and spine density in neocortex vary qualitatively with age and sex. Neurobiol. Learn. Mem. 2003, 79, 1–10. [Google Scholar] [CrossRef]
- Erickson, K.I.; Kramer, A.F. Aerobic exercise effects on cognitive and neural plasticity in older adults. Br. J. Sports Med. 2008, 43, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.F.; Colcombe, S. Fitness Effects on the Cognitive Function of Older Adults: A Meta-Analytic Study—Revisited. Perspect. Psychol. Sci. 2018, 13, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.J.; Blumenthal, J.A.; Hoffman, B.M.; Cooper, H.M.; Strauman, T.A.; Welsh-Bohmer, K.A.; Browndyke, J.N.; Sherwood, A. Aerobic Exercise and Neurocognitive Performance: A Meta-Analytic Review of Randomized Controlled Trials. Psychosom. Med. 2010, 72, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, C.; Hu, X.; Liu, X.; Song, W. The Effects of Mind-Body Exercise on Cognitive Performance in Elderly: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2018, 15, 2791. [Google Scholar] [CrossRef]
- Hänggi, J.; Koeneke, S.; Bezzola, L.; Jäncke, L. Structural neuroplasticity in the sensorimotor network of professional female ballet dancers. Hum. Brain Mapp. 2009, 31, 1196–1206. [Google Scholar] [CrossRef]
- Román-Caballero, R.; Arnedo, M.; Triviño, M.; Lupiáñez, J. Musical practice as an enhancer of cognitive function in healthy aging—A systematic review and meta-analysis. PLoS ONE 2018, 13, e0207957. [Google Scholar] [CrossRef]
- Kattenstroth, J.-C.; Kalisch, T.; Eholt, S.; Tegenthoff, M.; Dinse, H.R. Six months of dance intervention enhances postural, sensorimotor, and cognitive performance in elderly without affecting cardio-respiratory functions. Front. Aging Neurosci. 2013, 5, 5. [Google Scholar] [CrossRef]
- Duan, X.; He, S.; Liao, W.; Liang, D.; Qiu, L.; Wei, L.; Li, Y.; Liu, C.; Gong, Q.; Chen, H. Reduced caudate volume and enhanced striatal-DMN integration in chess experts. Neuroimage 2012, 60, 1280–1286. [Google Scholar] [CrossRef]
- Verghese, J.; Lipton, R.B.; Katz, M.J.; Hall, C.B.; Derby, C.A.; Kuslansky, G.; Ambrose, A.F.; Sliwinski, M.; Buschke, H. Leisure Activities and the Risk of Dementia in the Elderly. N. Engl. J. Med. 2003, 348, 2508–2516. [Google Scholar] [CrossRef]
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of Alzheimer’s disease. Alzheimers Dement. 2007, 3, 186–191. [Google Scholar] [CrossRef]
- Zatorre, R.J.; Chen, J.L.; Penhune, V.B. When the brain plays music: Auditory–motor interactions in music perception and production. Nat. Rev. Neurosci. 2007, 8, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Herholz, S.C.; Zatorre, R.J. Musical Training as a Framework for Brain Plasticity: Behavior, Function, and Structure. Neuron 2012, 76, 486–502. [Google Scholar] [CrossRef] [PubMed]
- Bugos, J.A.; Perlstein, W.M.; McCrae, C.S.; Brophy, T.S.; Bedenbaugh, P.H. Individualized Piano Instruction enhances executive functioning and working memory in older adults. Aging Ment. Health 2007, 11, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Seinfeld, S.; Figueroa, H.; Ortiz-Gil, J.; Sanchez-Vives, M.V. Effects of music learning and piano practice on cognitive function, mood and quality of life in older adults. Front. Psychol. 2013, 4, 810. [Google Scholar] [CrossRef]
- Hanna-Pladdy, B.; Mackay, A. The relation between instrumental musical activity and cognitive aging. Neuropsychology 2011, 25, 378–386. [Google Scholar] [CrossRef]
- Hanna-Pladdy, B.; Gajewski, B.J. Recent and Past Musical Activity Predicts Cognitive Aging Variability: Direct Comparison with General Lifestyle Activities. Front. Hum. Neurosci. 2012, 6, 198. [Google Scholar] [CrossRef]
- Moussard, A.; Bermudez, P.; Alain, C.; Tays, W.; Moreno, S. Life-long music practice and executive control in older adults: An event-related potential study. Brain Res. 2016, 1642, 146–153. [Google Scholar] [CrossRef]
- Efauvel, B.; Egroussard, M.; Emutlu, J.; Arenaza-Urquijo, E.M.; Eeustache, F.; Desgranges, B.; Platel, H. Musical practice and cognitive aging: Two cross-sectional studies point to phonemic fluency as a potential candidate for a use-dependent adaptation. Front. Aging Neurosci. 2014, 6, 227. [Google Scholar] [CrossRef]
- Gaser, C.; Schlaug, G. Brain Structures Differ between Musicians and Non-Musicians. J. Neurosci. 2003, 23, 9240–9245. [Google Scholar] [CrossRef]
- James, C.E.; Oechslin, M.S.; Van De Ville, D.; Hauert, C.-A.; Descloux, C.; Lazeyras, F. Musical training intensity yields opposite effects on grey matter density in cognitive versus sensorimotor networks. Brain Struct. Funct. 2014, 219, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.; Fujii, S.; Li, H.; Schlaug, G. The cerebellum’s contribution to beat interval discrimination. NeuroImage 2017, 163, 177–182. [Google Scholar] [CrossRef]
- Nigmatullina, Y.; Hellyer, P.J.; Nachev, P.; Sharp, D.J.; Seemungal, B.M. The Neuroanatomical Correlates of Training-Related Perceptuo-Reflex Uncoupling in Dancers. Cereb. Cortex 2015, 25, 554–562. [Google Scholar] [CrossRef]
- Müllensiefen, D.; Gingras, B.; Musil, J.; Stewart, L. The Musicality of Non-Musicians: An Index for Assessing Musical Sophistication in the General Population. PLoS ONE 2014, 9, e89642. [Google Scholar] [CrossRef]
- Vaquero, L.; Hartmann, K.; Ripollés, P.; Rojo, N.; Sierpowska, J.; François, C.; Camara, E.; Van Vugt, F.T.; Mohammadi, B.; Samii, A.; et al. Structural neuroplasticity in expert pianists depends on the age of musical training onset. NeuroImage 2016, 126, 106–119. [Google Scholar] [CrossRef]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Ségonne, F.; Dale, A.; Busa, E.; Glessner, M.; Salat, D.; Hahn, H.; Fischl, B. A hybrid approach to the skull stripping problem in MRI. NeuroImage 2004, 22, 1060–1075. [Google Scholar] [CrossRef] [PubMed]
- Gronenschild, E.H.B.M.; Habets, P.; Jacobs, H.I.L.; Mengelers, R.; Rozendaal, N.; Van Os, J.; Marcelis, M. The Effects of FreeSurfer Version, Workstation Type, and Macintosh Operating System Version on Anatomical Volume and Cortical Thickness Measurements. PLoS ONE 2012, 7, e38234. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.M.; Fischl, B.; Sereno, M.I. Cortical surface-based analysis: I. Segmentation and surface reconstruction. Neuroimage 1999, 9, 179–194. [Google Scholar] [CrossRef]
- Dale, A.M.; Sereno, M.I. Improved localization of cortical activity by combining EEG and MEG with MRI cortical surface reconstruction: A linear approach. J. Cogn. Neurosci. 1993, 5, 162–176. [Google Scholar] [CrossRef]
- Fischl, B.; Dale, A.M. Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proc. Natl. Acad. Sci. USA 2000, 97, 11050–11055. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Tourville, J.A. 101 Labeled Brain Images and a Consistent Human Cortical Labeling Protocol. Front. Behav. Neurosci. 2012, 6, 171. [Google Scholar] [CrossRef] [PubMed]
- Degé, F.; Kerkovius, K. The effects of drumming on working memory in older adults. Ann. N. Y. Acad. Sci. 2018, 1423, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, A.N.; Diaz, A.F.; Rogalsky, C. The relationship between the neural computations for speech and music perception is context-dependent: An activation likelihood estimate study. Front. Psychol. 2015, 6, 1138. [Google Scholar] [CrossRef] [PubMed]
- Levitin, D.J.; Menon, V. Musical structure is processed in “language” areas of the brain: A possible role for Brodmann Area 47 in temporal coherence. NeuroImage 2003, 20, 2142–2152. [Google Scholar] [CrossRef] [PubMed]
- Bugos, J.A. The benefits of music instruction on processing speed, verbal fluency, and cognitive control in aging. Music Educ. Res. Int. 2010, 4, 1–8. [Google Scholar]
- Parbery-Clark, A.; Skoe, E.; Lam, C.; Kraus, N. Musician Enhancement for Speech-In-Noise. Ear Hear. 2009, 30, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Strait, D.L.; Anderson, S.; Hittner, E.; Kraus, N. Musical Experience and the Aging Auditory System: Implications for Cognitive Abilities and Hearing Speech in Noise. PLoS ONE 2011, 6, e18082. [Google Scholar] [CrossRef] [PubMed]
- Strait, D.L.; Kraus, N. Can You Hear Me Now? Musical Training Shapes Functional Brain Networks for Selective Auditory Attention and Hearing Speech in Noise. Front. Psychol. 2011, 2, 113. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.M.; Ceci, S.J. When and where do we apply what we learn? A taxonomy for far transfer. Psychol. Bull. 2002, 128, 612–637. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Levitin, D. The rewards of music listening: Response and physiological connectivity of the mesolimbic system. NeuroImage 2005, 28, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Sammler, D.; Koelsch, S.; Friederici, A.D. Are left fronto-temporal brain areas a prerequisite for normal music-syntactic processing? Cortex 2011, 47, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Maess, B.; Koelsch, S.; Gunter, T.C.; Friederici, A.D. Musical syntax is processed in Broca’s area: An MEG study. Nat. Neurosci. 2001, 4, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Patel, A. Why would Musical Training Benefit the Neural Encoding of Speech? The OPERA Hypothesis. Front. Psychol. 2011, 2, 142. [Google Scholar] [CrossRef] [PubMed]
- Parbery-Clark, A.; Anderson, S.; Hittner, E.; Kraus, N. Musical experience offsets age-related delays in neural timing. Neurobiol. Aging 2012, 33, 1483.e1–1483.e4. [Google Scholar] [CrossRef]
- White-Schwoch, T.; Carr, K.W.; Anderson, S.; Strait, D.L.; Kraus, N. Older Adults Benefit from Music Training Early in Life: Biological Evidence for Long-Term Training-Driven Plasticity. J. Neurosci. 2013, 33, 17667–17674. [Google Scholar] [CrossRef]
- Abdul-Kareem, I.A.; Stancak, A.; Parkes, L.M.; Sluming, V. Increased gray matter volume of left pars opercularis in male orchestral musicians correlate positively with years of musical performance. J. Magn. Reson. Imaging 2010, 33, 24–32. [Google Scholar] [CrossRef]
- Slumingab, V.; Barricka, T.; Howard, M.; Cezayirli, E.; Mayese, A.; Robertsa, N. Voxel-Based Morphometry Reveals Increased Gray Matter Density in Broca’s Area in Male Symphony Orchestra Musicians. NeuroImage 2002, 17, 1613–1622. [Google Scholar] [CrossRef]
- Sluming, V.; Brooks, J.; Howard, M.; Downes, J.J.; Roberts, N. Broca’s Area Supports Enhanced Visuospatial Cognition in Orchestral Musicians. J. Neurosci. 2007, 27, 3799–3806. [Google Scholar] [CrossRef]
- Bermudez, P.; Lerch, J.P.; Evans, A.C.; Zatorre, R.J. Neuroanatomical Correlates of Musicianship as Revealed by Cortical Thickness and Voxel-Based Morphometry. Cereb. Cortex 2008, 19, 1583–1596. [Google Scholar] [CrossRef]
- Mansens, D.; Deeg, D.J.H.; Comijs, H.C. The association between singing and/or playing a musical instrument and cognitive functions in older adults. Aging Ment. Health 2017, 22, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Williamon, A.; Egner, T. Memory structures for encoding and retrieving a piece of music: An ERP investigation. Cogn. Brain Res. 2004, 22, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Williamson, V.J.; Baddeley, A.D.; Hitch, G.J. Musicians’ and nonmusicians’ short-term memory for verbal and musical sequences: Comparing phonological similarity and pitch proximity. Mem. Cogn. 2010, 38, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, M.S.; Van De Ville, D.; Lazeyras, F.; Hauert, C.-A.; James, C.E. Degree of Musical Expertise Modulates Higher Order Brain Functioning. Cereb. Cortex 2012, 23, 2213–2224. [Google Scholar] [CrossRef] [PubMed]
- George, E.M.; Coch, D. Music training and working memory: An ERP study. Neuropsychologia 2011, 49, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.M.; Schultz, A.P.; Huijbers, W.; Van Dijk, K.R.; Hedden, T.; A Sperling, R. The parahippocampal gyrus links the default-mode cortical network with the medial temporal lobe memory system. Hum. Brain Mapp. 2014, 35, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Hampson, M.; Driesen, N.R.; Skudlarski, P.; Gore, J.C.; Constable, R.T. Brain Connectivity Related to Working Memory Performance. J. Neurosci. 2006, 26, 13338–13343. [Google Scholar] [CrossRef] [PubMed]
- Andrews-Hanna, J.R.; Smallwood, J.; Spreng, R.N. The default network and self-generated thought: Component processes, dynamic control, and clinical relevance. Ann. N. Y. Acad. Sci. 2014, 1316, 29–52. [Google Scholar] [CrossRef] [PubMed]
- Belden, A.; Zeng, T.; Przysinda, E.; Anteraper, S.A.; Whitfield-Gabrieli, S.; Loui, P. Improvising at rest: Differentiating jazz and classical music training with resting state functional connectivity. NeuroImage 2020, 207, 116384. [Google Scholar] [CrossRef] [PubMed]
- Grassi, M.; Meneghetti, C.; Toffalini, E.; Borella, E. Auditory and cognitive performance in elderly musicians and nonmusicians. PLoS ONE 2017, 12, e0187881. [Google Scholar] [CrossRef]
- Bialystok, E.; DePape, A.-M. Musical expertise, bilingualism, and executive functioning. J. Exp. Psychol. Hum. Percept. Perform. 2009, 35, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Loureiro, M.A.; Carthery, M.T. Long-term musical training may improve different forms of visual attention ability. Brain Cogn. 2013, 82, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kirino, E. Reorganization of the thalamocortical network in musicians. Brain Res. 2017, 1664, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Blood, A.J.; Zatorre, R.J.; Bermudez, P.; Evans, A.C. Emotional responses to pleasant and unpleasant music correlate with activity in paralimbic brain regions. Nat. Neurosci. 1999, 2, 382–387. [Google Scholar] [CrossRef]
- Gosselin, N.; Samson, S.; Adolphs, R.; Noulhiane, M.; Roy, M.; Hasboun, D.; Baulac, M.; Peretz, I. Emotional responses to unpleasant music correlates with damage to the parahippocampal cortex. Brain 2006, 129, 2585–2592. [Google Scholar] [CrossRef]
- Chanda, M.L.; Levitin, D.J. The neurochemistry of music. Trends Cogn. Sci. 2013, 17, 179–193. [Google Scholar] [CrossRef]
- Panksepp, J. The Emotional Sources of “Chills” Induced by Music. Music Percept. 1995, 13, 171–207. [Google Scholar] [CrossRef]
- Grewe, O.; Nagel, F.; Kopiez, R.; Altenmüller, E. How Does Music Arouse “Chills”? Investigating Strong Emotions, Combining Psychological, Physiological, and Psychoacoustical Methods. Ann. N. Y. Acad. Sci. 2005, 1060, 446–449. [Google Scholar] [CrossRef]
- Altenmüller, E.; Esiggel, S.; Mohammadi, B.; Samii, M.; Münte, T.F. Play it again sam: Brain correlates of emotional music recognition. Front. Psychol. 2014, 5, 114. [Google Scholar] [CrossRef]
- Steinbeis, N.; Koelsch, S.; Sloboda, J.A. The Role of Harmonic Expectancy Violations in Musical Emotions: Evidence from Subjective, Physiological, and Neural Responses. J. Cogn. Neurosci. 2006, 18, 1380–1393. [Google Scholar] [CrossRef]
- Blood, A.J.; Zatorre, R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.; McGill, J. Music, the food of neuroscience? Nat. Cell Biol. 2005, 434, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Salimpoor, V.N.; Benovoy, M.; Larcher, K.; Dagher, A.; Zatorre, R.J. Anatomically distinct dopamine release during anticipation and experience of peak emotion to music. Nat. Neurosci. 2011, 14, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Dosenbach, N.U.F.; Visscher, K.M.; Palmer, E.D.; Miezin, F.M.; Wenger, K.K.; Kang, H.C.; Burgund, E.D.; Grimes, A.L.; Schlaggar, B.L.; Petersen, S.E. A Core System for the Implementation of Task Sets. Neuron 2006, 50, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, M.F.S.; E Walton, M.; Kennerley, S.W.; Bannerman, D.M. Action sets and decisions in the medial frontal cortex. Trends Cogn. Sci. 2004, 8, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Anat. Embryol. 2010, 214, 655–667. [Google Scholar] [CrossRef]
- Uddin, L.Q. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Chand, G.B.; Wu, J.; Hajjar, I.; Qiu, D. Interactions of the Salience Network and Its Subsystems with the Default-Mode and the Central-Executive Networks in Normal Aging and Mild Cognitive Impairment. Brain Connect. 2017, 7, 401–412. [Google Scholar] [CrossRef]
- Wang, D.; Belden, A.; Hanser, S.B.; Geddes, M.R.; Loui, P. Resting-State Connectivity of Auditory and Reward Systems in Alzheimer’s Disease and Mild Cognitive Impairment. Front. Hum. Neurosci. 2020, 14, 280. [Google Scholar] [CrossRef]
- Martínez-Molina, N.; Mas-Herrero, E.; Rodríguez-Fornells, A.; Zatorre, R.J.; Marco-Pallarés, J. White Matter Microstructure Reflects Individual Differences in Music Reward Sensitivity. J. Neurosci. 2019, 39, 5018–5027. [Google Scholar] [CrossRef] [PubMed]
- Sachs, M.E.; Ellis, R.J.; Schlaug, G.; Loui, P. Brain connectivity reflects human aesthetic responses to music. Soc. Cogn. Affect. Neurosci. 2016, 11, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Zamorano, A.M.; Cifre, I.; Montoya, P.; Riquelme, I.; Kleber, B. Insula-based networks in professional musicians: Evidence for increased functional connectivity during resting state fMRI. Hum. Brain Mapp. 2017, 38, 4834–4849. [Google Scholar] [CrossRef] [PubMed]
- Loui, P.; Patterson, S.; Sachs, M.E.; Leung, Y.; Zeng, T.; Przysinda, E. White Matter Correlates of Musical Anhedonia: Implications for Evolution of Music. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Stern, Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef]
- Tierney, A.T.; Bergeson-Dana, T.R.; Pisoni, D.B. Effects of early musical experience on auditory sequence memory. Empir. Music. Rev. 2008, 3, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Hanser, S.B. Music therapy in cardiac health care: Current issues in research. Cardiol. Rev. 2014, 22, 37–42. [Google Scholar] [CrossRef]
- Särkämö, T.; Tervaniemi, M.; Laitinen, L.S.; Numminen, A.; Kurki, M.; Johnson, J.K.; Rantanen, P. Cognitive, Emotional, and Social Benefits of Regular Musical Activities in Early Dementia: Randomized Controlled Study. Gerontologist 2013, 54, 634–650. [Google Scholar] [CrossRef]
- Green, C.S.; Bavelier, D.; Kramer, A.F.; Vinogradov, S.; Ansorge, U.; Ball, K.K.; Bingel, U.; Chein, J.M.; Colzato, L.S.; Edwards, J.D.; et al. Improving Methodological Standards in Behavioral Interventions for Cognitive Enhancement. J. Cogn. Enhanc. 2019, 3, 2–29. [Google Scholar] [CrossRef]
- Park, D.C.; Bischof, G.N. The aging mind: Neuroplasticity in response to cognitive training. Dialog. Clin. Neurosci. 2013, 15, 109–119. [Google Scholar]
- Music on Our Minds: The Rich Potential of Music to Promote Brain Health and Mental Well-Being. 2020. Available online: https://www.aarp.org/content/dam/aarp/health/brain_health/2020/06/gcbh-music-report-english.doi.10.26419-2Fpia.00103.001.pdf (accessed on 25 December 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean (SD) (Present Study) | Percentile Relative to Norms * |
|---|---|---|
| Active Engagement | 31.33 (9.3) | 17% |
| Perceptual Abilities | 45.48 (8.4) | 48% |
| Musical Training | 21.74 (8.5) | 47% |
| Singing Abilities | 26.75 (8.4) | 46% |
| Emotions | 30.49 (5.6) | 44% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaddock-Heyman, L.; Loui, P.; Weng, T.B.; Weisshappel, R.; McAuley, E.; Kramer, A.F. Musical Training and Brain Volume in Older Adults. Brain Sci. 2021, 11, 50. https://doi.org/10.3390/brainsci11010050
Chaddock-Heyman L, Loui P, Weng TB, Weisshappel R, McAuley E, Kramer AF. Musical Training and Brain Volume in Older Adults. Brain Sciences. 2021; 11(1):50. https://doi.org/10.3390/brainsci11010050
Chicago/Turabian StyleChaddock-Heyman, Laura, Psyche Loui, Timothy B. Weng, Robert Weisshappel, Edward McAuley, and Arthur F. Kramer. 2021. "Musical Training and Brain Volume in Older Adults" Brain Sciences 11, no. 1: 50. https://doi.org/10.3390/brainsci11010050
APA StyleChaddock-Heyman, L., Loui, P., Weng, T. B., Weisshappel, R., McAuley, E., & Kramer, A. F. (2021). Musical Training and Brain Volume in Older Adults. Brain Sciences, 11(1), 50. https://doi.org/10.3390/brainsci11010050