Abstract
Vitellogenesis is a pivotal reproductive process of the yolk formation in crustaceans. Vitellogenin (VTG) is the precursor of main yolk proteins and synthesized by endogenous estrogens. The intertidal mud crab (Macrophthalmus japonicus) inhabits sediment and is a good indicator for assessing polluted benthic environments. The purpose of this study was to identify potential responses of M. japonicus VTG under environmental stresses caused by chemical pollutants, such as 1, 10, and 30 µg L−1 concentrations in di(2-ethylhexyl) phthalate (DEHP), bisphenol A (BPA) and irgarol. We characterized the M. japonicus VTG gene and analyzed the transcriptional expression of VTG mRNA in M. japonicus exposed to various chemicals and exposure periods. A phylogenetic analysis revealed that the M. japonicus VTG clustered closely with Eriocheir sinensis (Chinese mitten crab) VTG, in contrast with another clade that included the VTG ortholog of other crabs. The basal level of VTG expression was the highest in the hepatopancreas and ovaries, and tissues. VTG expression significantly increased in the ovaries and hepatopancreas after 24 h exposure to DEHP. Increased responses of VTG transcripts were found in M. japonicus exposed to DEHP and BPA for 96 h; however, VTG expression decreased in both tissues after irgarol exposure. After an exposure of 7 d, VTG expression significantly increased in the ovaries and hepatopancreas for all concentrations of all chemicals. These results suggest that the crustacean embryogenesis and endocrine processes are impaired by the environmental chemical pollutants DEHP, BPA, and irgarol.
1. Introduction
Vitellogenesis is a hormonally regulated process for synthesizing yolk proteins and an important step in the reproductive development of crustaceans [1]. Vitellogenin (VTG), a precursor molecule of vitellin, is stored in crustacean hepatopancreas tissue and transported by the hemolymph to the ovaries. The sites of VTG synthesis are in the hepatopancreas and ovary tissues in decapod crustaceans [2,3,4,5]. VTG genes have shown specific levels of expressions that vary by tissue, sex, and development stage [6]. VTG inductions were observed in aquatic invertebrates by exposure to estrogenic compounds [7,8]. After endosulfan exposure, VTG gene expression was downregulated in Pandalus shrimp (Pandalopsis japonica) [9]. The regulation of VTG gene may be affected by several pollutants [7,8,9].
Environmental chemical pollution is globally one of the most critical ecological problems, owing to the high risks it poses to ecosystems and human life [10]. Bisphenol A (BPA), a common endocrine disrupting chemical (EDC), is a carbon-based synthetic compound and a xenoestrogen that interferes with estrogen receptor signaling [11]. BPA concentrations observed in U.S. river estuaries were approximately 4 μg L−1 to 21 μg L−1 [12,13]. Exposure to BPA induced transcription responses of the heat shock protein 90 (HSP90) gene in the marine crab Charybdis japonica [14]. Di-(2-ethylhexyl) phthalate (DEHP), a suspected EDC, is widely used as plasticizer and may induce reproductive and developmental toxicities in aquatic environments [15]. DEHP toxicity induced changes in the VTG levels in African sharptooth catfish (Clarias gariepinus) and the Chinese rare minnow (Gobiocypris rarus) [16,17]. DEHP levels were 0.47 to 12 μg L−1 in the China river estuary [18]. Irgarol, which is used as an algicide, has been found in coastal and estuarine environments because it is commonly used in antifouling systems [19,20]. Irgarol exposure induced downregulation of HSP90 expression in hard coral (Acropora tenuis) [21]. Worldwide, the environmental concentrations of irgarol in antifouling systems range from 1 ng L−1 to 1 mg L−1 [22]. Irgarol’s long half-life poses an ecological risk in estuarine ecosystems [20]. Although these pollutants are ubiquitously distributed in aquatic environments, there is not enough information about the effects of embryogenesis in crustaceans.
Crabs make up one of the largest crustacean families. The accumulation and synthesis of VTG is vital for oocyte and embryo development in crabs [2,6,23]. A burrowing mud crab, Macrophthalmus japonicus (Ocypodoidea), inhabits estuarine intertidal mud flats throughout the Indo-Pacific regions of Japan and Korea [24,25,26,27]. M. japonicus is also a major bioturbator of tidal flats and plays a crucial role in purifying sediment [28]. The movement of M. japonicus may affect the distribution of macro-infauna because of the accompanying disturbance in the physical and chemical properties of the sediment [25]. Previous studies have suggested that M. japonicus may be a useful indicator for modeling the exposure effects of chemical toxicity in the sediment environment [26,27,29,30].
In this study, we assessed the effects of crustacean embryogenesis and endocrine process by environmental chemical pollutants such as BPA, DEHP, and irgarol using the intertidal mud crab. To accomplish this, we characterized the M. japonicus VTG gene and analyzed phylogenetic relationships. The transcriptional responses of VTG and the tissue distribution of VTG mRNA expression were investigated from M. japonicus ovaries and hepatopancreases, plus other tissues, following exposure to either BPA, DEHP, or irgarol.
2. Materials and Methods
2.1. Organisms
M. japonicus was prepared from marine product markets in Yeosu (Jeonnam, Korea). On average, crabs ranged from 2.5 to 3.5 cm in carapace height, 2.7 to 4.3 cm in carapace width and 4 to 11 g in body weight. The crabs were placed in glass containers (45.7 × 35.6 × 30.5 cm) with natural seawater and aeration. All samples acclimatized for 1 d at 18 ± 1 °C in temperature, 25% salinity, and a 12 h light-dark schedule. Non-damaged crabs were selected. Experiments were performed based on the guidelines of the Chonnam National University Institutional Animal Care and Use Committee.
2.2. Exposure Experiments
Irgarol (2-(tert-butylamino)-4-(cyclo-propylamino)-6-(methyl-thio)-s-triazine) and BPA (99.9% pure) were prepared from Sigma-Aldrich (St. Louis, MO, USA). DEHP solutions were obtained from a solid compound (99%, Junsei Chemical Co. Ltd., Japan). The 10 mg L−1 stock solutions of BPA, DEHP, and irgarol were prepared by dissolving the chemicals in 99% acetone at room temperature. For working solutions of 1, 10, and 30 µg L−1 for each chemical; the stock solution was diluted with seawater. In solvent controls, a solvent concentration was <0.5% acetone.
For each chemical exposure, the crabs (n = 120) were randomly divided into four experimental groups (1, 10, and 30 µg L−1 treatment solutions, as well as solvent controls). Ten crabs were exposed with one of three doses of BPA, DEHP, or irgarol. Treatment times were 24 h, 96 h, and 7 d. Three individuals were subjected to tissue extractions for each time interval in each chemical treatment condition and control crab. The seawater was changed every day by adding the equivalent concentration of each chemical during the experiment. No food was provided during the experimental period. All experiments were run in three replicates with independent samples. Following exposure, tissue samples were immediately extracted from the crabs.
2.3. Characterization of the M. Japonicus Vitellogenin (VTG) Genes and Phylogenetic Analysis
VTG gene sequences were obtained using the 454 GS FLX transcriptomic database of the M. japonicus body [31]. The identified VTG cDNA sequences were compared with the VTG sequences in crustacean species available on the national center for biotechnology information (NCBI) database, using basic local alignment search tool (BLAST) searching (http://www.ncbi.nlm.nih.gov). The deduced amino acid sequences were obtained using the ExPASy translation program and aligned using the Clustal W2 tool.
To analyze the M. japonicus VTG gene in a phylogenetic tree, 14 crustacean VTG sequences were downloaded from NCBI for comparison of their similarities with the deduced amino acid sequences of the M. japonicus VTG. The GeneDoc Program (ver. 2.6.001) was used to display the multiple aligned sequences. A construction of the phylogenetic tree was made by neighbor-joining analyses of the Mega X program (version 10.04). The bootstrap value was calculated by 1000 replicates.
2.4. RNA Isolation and mRNA Expression Analysis
Total RNA from M. japonicus was obtained using the RNAiso Plus reagent (Takara, Dalian, China) following the manufacturer’s protocol. Recombinant DNase I (RNase free) (Takara, Kusatsu, Japan) treatment was used to eliminate contamination of genomic DNA. Integrity and quantity of the extracted RNA were checked using 0.8% agarose gel for electrophoresis and a Nano-Drop 1000 (Thermo Fisher Scientific, Foster City, CA, USA). cDNA synthesis was performed with 3 µg of total RNA using the SuperScript™III RT kit (Invitrogen, Foster City, CA, USA) following the manufacturer’s protocol. After synthesis, the diluted cDNA (50-fold) was stored at −80 °C.
To evaluate VTG gene responses in multiple tissues, the total RNA was extracted from M. japonicus in a seawater control. Quantitative real-time PCR (polymerase chain reaction, RT-PCR) was performed using the ExicyclerTM96 (Bioneer, Daejeon, Korea) with the master mix (Bioneer, Daejeon, Korea). The glyceraldehyde-3-phosphate dehydrogenase (Mj GAPDH) gene of M. japonicus was used as an internal reference [16]. The primer sequences were: Mj_VTG forward 5′-CTTGGGCTCTCCAGTTC TTG-3′; Mj_VTG reverse 5′-CCACGTATGCCTCTTTTGGT-3′; Mj_GAPDH forward 5′-TGCTGATGCACCCATGTTTG-3′; Mj_GAPDH reverse 5′-AGGCCCTGGACAATCTCAA AG-3′. The size of the PCR product was 164 bp for the VTG gene and 147 bp for the GAPDH gene. The reaction mixture (20 µL) contained 6 µL of diluted cDNA, 10 µL of 2x SYBR green dye (Bioneer, Daejeon, Korea), 0.5 µL of forward and reverse primers (10 µM), and 3.0 µL of RNase-free water. The following RT-PCR cycle was used for amplification of the VTG gene: 94 °C for 40 s, followed by 38 cycles of 94 °C for 15 s, 53 °C for 30 s and 72 °C for 40 s. The ExicyclerTM96 real time system program (version 3.54.8) was used for the verification of the RT-PCR baseline. The calculation for relative transcript levels was used by the 2−Δ Δct method [32] and normalized with GAPDH.
2.5. Statistical Analysis
Statistical analysis was conducted using statistical package for the social sciences (SPSS) 12.0 KO (SPSS Inc., Chicago, IL, USA). Statistical analysis for tissue distribution was performed using Dunnett’s multiple range test of One-Way Analysis of Variance (ANOVA). An independent sample t-test was used to compare significant differences in the transcriptional expression of VTG between the ovaries and hepatopancreas under different chemical concentrations. A two-way ANOVA was conducted to assess the effects of dose and exposure to each chemical on VTG expression. Differences were statistically significant at p < 0.05 (*) and p < 0.01 (**). Data are presented as the mean ± SD.
3. Results
3.1. Characterization of the M. Japonicus VTG Gene
The partial cDNA of the M. japonicus VTG gene was obtained from the GS-FLX transcriptome database of the M. japonicus crab [31]. The M. japonicus VTG DNA was 3700 bp long, included an open reading frame (ORF) of 1202 amino acids, and included a lipoprotein amino acid terminal region of VTG (Figure 1). The M. japonicus VTG nucleotide sequence was 87% homologous with that of Eriocheir sinensis (KC699915). The predicted amino acid sequences of the VTG were 84%, 72%, 65%, and 63% homologous with those of E. sinensis (AGM75775), Longpotamon honanense (freshwater crab; AKI23633), Scylla paramamosain (green mud crab; ACO36035), and Portunus trituberculatus (swimming crab; AAX94762), respectively. Thus, the VTG gene of M. japonicus does not show a high homology with that of the other crabs (Figure 1). A phylogenetic analysis revealed two clades of VTG of the crabs (Figure 2). One clade was composed of VTGs from M. japonicus, E. sinensis and L. honanense. Another clade had homologous VTGs from S. paramamosain, P. trituberculatus, Charybdis feriata (Indo-Pacific crab; AAU93694), and Callinectes sapidus (blue crab; ABC41925). In addition, the black tiger shrimp (Penaeus monodon) (ABB89953) VTG formed another clade with the VTG of lobster (Homarus americanus; ABO09863), multiple prawn species, and other related shrimp species (Figure 2).

Figure 1.
Vitellogenin (VTG) gene of the intertidal mud crab, M. japonicus. Multiple sequence alignments of the deduced M. japonicus VTG gene sequences with the homologous sequences of other crabs.
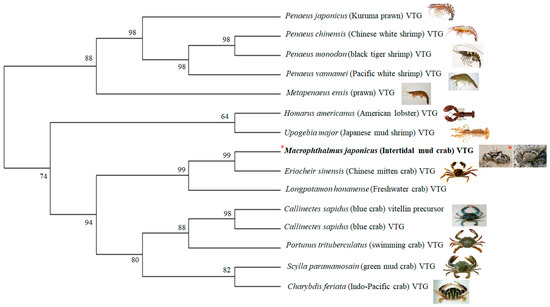
Figure 2.
Phylogenetic tree of M. japonicus VTG gene constructed by neighbor-joining analysis (bootstrap value 1000). The numbers at the nodes are the percentage bootstrap values. GenBank accession numbers for VTGs are: M. japonicus VTG (Acc **), Eriocheir sinensis (KC699915), Longpotamon honanense (AKI23633), Callinectes sapidus vitellin precursor (AEI59132), C. sapidus VTG (ABC41925), Portunus trituberculatus (AAX94762), Scylla paramamosain (ACO36035), Charybdis feriata (AAU93694), Homarus americanus (ABO09863), Upogebia major (BAF91417), Penaeus japonicus (BAD98732), Penaeus chinensis (ABC86571), Penaeus monodon (ABB89953), Penaeus vannamei (AAP76571), and Metapenaeus ensis (AAN40700). ** Accession #s to be deposited in GenBank.
3.2. Basal Levels of M. Japonicus VTG Expression in Multiple Tissues
The basal expression levels of VTG were observed in six tissues (gill, hepatopancreas, muscle, ovaries, heart, and stomach) (Figure 3). The fold change of each VTG mRNA expression in each tissue was based on the VTG expression level in gill tissue (when the set value of the gill = 1). The M. japonicus VTG gene expression was expressed differentially in all investigated tissues. The high level of M. japonicus VTG expression was evident in the ovaries and hepatopancreas, whereas relatively low levels were observed in the gill, muscle, stomach, and heart. The M. japonicus VTG mRNA expression varied significantly between the crab gill and ovaries, or hepatopancreas (p < 0.05).
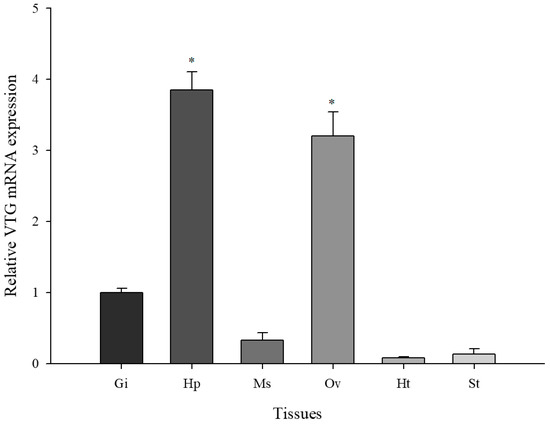
Figure 3.
Basal expression of M. japonicus VTG transcripts among different tissues (Gi, gill; Hp, hepatopancreas; Ms, muscle; Ov, ovaries; Ht, heart; St, stomach). Each tissue was collected from 10 mud crabs. The experiments were performed in triplicate. The data are represented as the mean ± SD. The relative expression levels of each gene in each tissue were compared with the respective gene expression levels in gills (relative to set value of gill = 1). Significant differences at * p < 0.05 are indicated with an asterisk.
3.3. VTG Gene Expressions in M. Japonicus Ovaries and Hepatopancreas after DEHP, BPA, or Irgarol Exposures
After an exposure of 24 h, VTG mRNA expression generally increased in M. japonicus hepatopancreas exposed to DEHP, BPA or irgarol (Figure 4A). In particular, exposure to 1 and 30 µg L−1 DEHP significantly induced VTG gene expression (p < 0.05) relative to the control (when the value of the control = 1). In the ovaries, transcriptional levels of VTG increased slightly after BPA exposure (Figure 4B). In contrast, irgarol slightly decreased VTG expression at a relatively low concentration of 1 µg L−1, although VTG response was somewhat induced at a relatively high irgarol concentration of 30 µg L−1. After exposure to different concentrations of DEHP, VTG gene expression was significantly upregulated at 10 (2.8-fold) and 30 µg L−1 (3.3-fold) of DEHP in a concentration-dependent manner (p < 0.05).
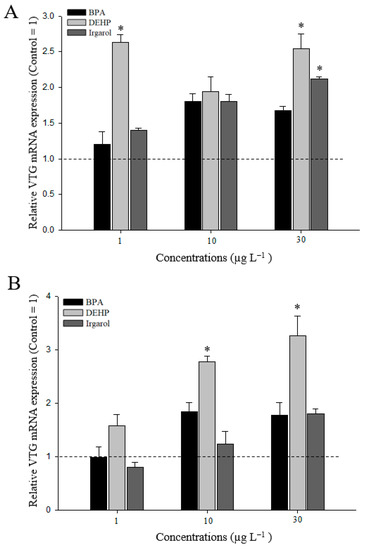
Figure 4.
Transcriptional expression of VTG gene in M. japonicus hepatopancreas (A) and ovaries (B) exposed to each dose of bisphenol A (BPA), di(2-ethylhexyl) phthalate (DEHP), and irgarol for 24 h. The values were normalized against glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Values of each bar represent the mean ± SD. A statistically significant difference is presented by an asterisk at * p < 0.05 as compare to the control (relative control value of VTG = 1).
The VTG gene response increased in hepatopancreas exposed to DEHP and BPA for 96 h, whereas a decrease in VTG expression resulted from irgarol exposure (Figure 5A). The up-regulation of VTG gene expression was significantly different at a relatively high concentration of BPA (30 µg L−1) (p < 0.05). In addition, DEHP exposure significantly induced the up-regulation of the VTG expression in the hepatopancreas. The highest VTG expression was observed at 30 µg L−1 (4.9-fold) DEHP (p < 0.01). In contrast, after 96 h of irgarol exposure, VTG decreased in a dose-dependent manner. In the M. japonicus ovaries, BPA and DEHP exposure significantly upregulated transcriptional levels of VTG following exposure to all concentrations (p < 0.05) (Figure 5B). Similar to the VTG gene response in the hepatopancreas, the VTG expression in the M. japonicus ovaries downregulated after irgarol exposure for 96 h. The highest level of VTG expression was found at 30 µg L−1 (5.1-fold) of DEHP (p < 0.01).
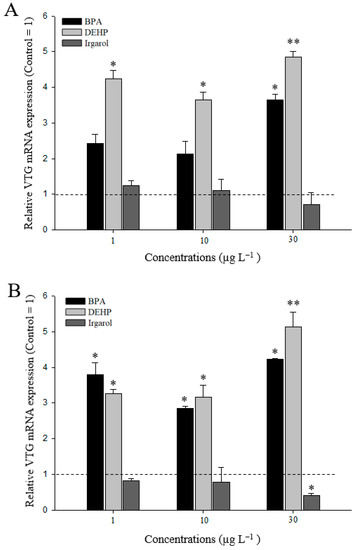
Figure 5.
Expression of VTG mRNA in M. japonicus hepatopancreas (A) and ovaries (B) after exposure to each dose of BPA, DEHP, and irgarol for 96 h. The values were normalized against GAPDH. Values of each bar represent the mean ± SD. A statistically significant difference is presented by an asterisk at * p < 0.05 and ** p < 0.01 as compare to the control (relative control value of VTG = 1).
At 7 d of exposure, there were different responses of the VTG gene between the hepatopancreas and ovary tissues (Figure 6). In the hepatopancreas, significant up-regulation of the VTG gene was observed at relatively low concentrations of 1 µg L−1 of BPA (4.3 fold) and DEHP (5.1 fold) (Figure 6A). The responses of VTG gene expression decreased in a concentration-dependent manner. After irgarol exposure, VTG mRNA expression slightly increased in the M. japonicus hepatopancreas, compared to the control level, for 7 d. In terms of VTG response in the ovaries, VTG expression was upregulated at all concentrations of BPA, DEHP, or irgarol exposure (Figure 6B). BPA exposure significantly increased VTG gene expression at 1 (4.3-fold), 10 (4.0-fold), and 30 µg L−1 (5.1-fold) of BPA (p < 0.05). DEHP treatment also significantly induced the VTG mRNA level at 1 (4.2 fold), 10 (5.3 fold) and 30 µg L−1 (7.2 fold) of DEHP (p < 0.01). The trend of VTG in response to DEHP exposure increased in a dose-dependent manner. In addition, significant expression of VTG was found at a relatively high concentration of 30 µg L−1 (3.8 fold) of irgarol, although VTG gene expression generally increased in M. japonicus ovaries exposed to irgarol for 7 d. Furthermore, there are no significant difference in weight changes between the control and treated groups after 7 d exposures to each chemical (Supplementary Figure S1).
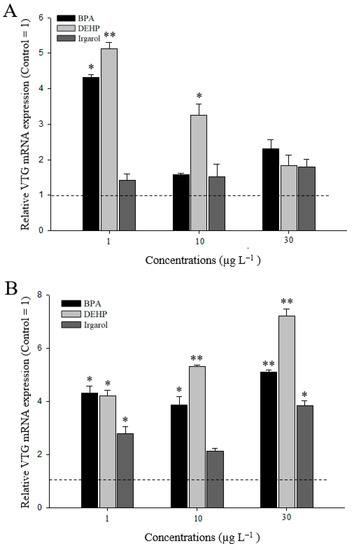
Figure 6.
Expression of VTG mRNA in M. japonicus hepatopancreas (A) and ovaries (B) after exposure to each dose of BPA, DEHP, and irgarol for 7 d. The values were normalized against GAPDH. Values of each bar represent the mean ± SD. A statistically significant difference was presented by an asterisk at * p < 0.05 and ** p < 0.01 as compare to the control (relative control value of VTG = 1).
With different exposure times, VTG gene expression for 24 or 96 h did not differ significantly between the hepatopancreas and ovary tissues (p > 0.05). VTG expression levels for 7 d were significantly different between the hepatopancreas and gill (p < 0.001). An increase of the VTG gene expression was observed to be time-dependent in M. japonicus hepatopancreas exposed to the relatively low concentrations of 1 µg L−1 BPA and DEHP (Supplementary Figure S2). In ovary tissue, the up-regulations of VTG gene were found to be time-dependent at all concentrations of BPA and DEHP (Supplementary Figure S3).
4. Discussion
To understand VTG gene responses following exposure to environmental chemical pollutants, the cDNA sequence of M. japonicus VTG was identified and analyzed using phylogenetic analysis. The three functional domains of the crustacean VTG gene are the lipoprotein domain at the N-terminus (LPD_N), the DUF1943 domain, and the von Willebrand factors type-D domain (VWD) [2,23,33]. In this study, we characterized the M. japonicus VTG with 3700 bp ORF, which encodes a protein of 1202 amino acids. The deduced protein region of the M. japonicus VTG was predicted as the LPD_N of VTG, which is a conserved region found in various lipid transport proteins [34]. In multiple alignments of the amino acid sequences, M. japonicus VTG showed high homology with the protein from E. sinensis. The phylogenetic analysis revealed two clusters representing shrimps or crabs among the crustaceans. In these clusters, three clades were represented by crabs, including M. japonicus, other crabs, and H. americanus.
In this study, it was observed that M. japonicus mainly synthesizes VTG in the hepatopancreas and ovaries. In crustaceans, the hepatopancreas is a critical storage system of nutrients and energy in the forms of carbohydrates, proteins, and lipids, which are essential in regulating an energy balance during the reproductive cycle [35]. The hepatopancreas was also reported to be the main source of VTG activity in decapod species, although VTG expression was also found in the ovaries [5]. The primary expression site of the VTG gene in crustaceans has been a controversial topic. VTG production was only observed in the hepatopancreas of Macrobrachium rosenbergii (freshwater prawn), Cherax quadricarinatus (red claw crayfish), Pandalus hypsinotus (coonstriped shrimp) and P. trituberculatus [2,36,37,38]. In contrast, the VTG gene was expressed in both the hepatopancreas and ovaries of Upogebia major (mud shrimp) and marine crabs such as S. paramamosain, Carcinus maenas (green crab), C. sapidus, C. japonica and E. sinensis [9,23,39,40,41]. We identified that the major sites for VTG synthesis in M. japonicus are the hepatopancreas and ovaries.
The endocrine disruptor BPA is presently considered a ubiquitous emerging pollutant in aquatic environments. In this study, exposures to BPA triggered up-regulation of M. japonicus VTG expression in the hepatopancreas and ovary tissues for different exposure periods. BPA exposure increased VTG levels in various fish, such as fathead minnows, goldfish, and bluegills [42]. BPA toxicity at lower concentrations (<15 µg L−1) showed induced ovarian expression of a steroidogenic gene involving VTG in G. rarus [43]. In addition, different modulations of VTG were reported in two VTG genes in the gilthead bream Sparus aurata and G. rarus [43,44]. These results indicate that induction of VTG following BPA exposure is a major response to the stressful effects of endocrine-disruption.
The endocrine effects of DEHP, a distinctive EDC, are noticeable in marine ecosystems where high concentrations of DEHP have been found in river estuaries and coastal areas [45]. In our study, DEHP exposure generally increased M. japonicus VTG expression in the tested tissues. The up-regulation of VTG gene expression in M. japonicus was more sensitive to DEHP exposure than to exposure to the other chemicals tested. In addition, transcriptional responses of the VTG gene in the hepatopancreas were found to be significant under relatively low concentrations of DEHP for the different exposure times, whereas VTG was expressed in the ovaries under all concentrations of DEHP tested. Adeogun reported that DEHP exposure induced time- and concentration-dependent increases of VTG levels in C. gariepinus [17]. Long exposure to low doses of DEHP reduced oocyte maturation [46] and increased the levels of hepatic VTG transcripts in G. rarus [16]. In addition, VTG levels in marine medaka livers increased after DEHP exposure [47]. These results strongly emphasize the suitability of using the VTG gene as a biomarker to assess the environmental presence of DEHP.
Since the use of irgarol as an algaecide has increased, irgarol has been detected frequently in coastal environments [48]. However, there is not enough information about the potential effects of irgarol exposure on the endocrine system of crustaceans. In our study, the responses of VTG gene expression following exposure to irgarol were different. After a 24 h exposure to irgarol, the M. japonicus VTG gene was expressed in a dose-dependent manner. The up-regulation of VTG relative to the control decreased in the hepatopancreas and ovaries when exposed to irgarol for 96 h. VTG gene expression recovered after 7 d of irgarol exposure. These results are the first report of the endocrine effects of irgarol exposure in crustaceans. A recent study reported that irgarol exposure induced changes in thyroid endpoints in the inland silverside fish (Menidia beryllina) under salinity and temperature changes [28]. In A. tenuis, exposure to irgarol for 7 d resulted in body color changes and reduced transcriptional expression of the HSP90 gene [29]. In a previous study, irgarol exposure induced expression changes of digestion related genes and chitinase genes [17]. Thus, VTG gene expression was affected by environmental chemical pollutants such as BPA, DEHP, and irgarol. After EDC exposures, the expression pattern of the M. japonicus VTG gene was correlated with short-term EDC exposure in the hepatopancreas (p = 0.791) and long-term EDC exposure in ovary tissue (p = 0.927).
In conclusion, our results demonstrate that M. japonicus is sensitive to several chemical pollutants, as measured by changes of the VTG gene expression in the hepatopancreas and ovaries. The results suggest that the molecular balance of VTG expression levels of intertidal mud crabs appear to be affected by EDCs such as BPA, DEHP, and irgarol at even relatively low concentrations of 1 µg L−1. The expression levels of the VTG gene are important factors to be considered for evaluating xenoestrogenic responses. The change of the VTG responses are related with disturbed processes of reproductive physiology such as egg yolk generation in M. japonicus, because of the up-regulation of VTG expression in the ovaries after EDC exposures. The crab hepatopancreas is an organ for VTG synthesis and storage of energy accumulation, which can support lipid transportation for oogenesis [49]. EDC exposure also impaired the molecular balance of the VTG level through the disturbance of energy metabolisms in M. japonicus hepatopancreas. Up or down-regulation of VTG gene may result in sensitive defense activities of M. japonicus crabs to potential EDCs pollutants, given the major function of VTG in stabilizing the endocrine process and triggering lipid uptake for energy storage during crustacean embryogenesis. Our findings suggest that M. japonicus VTG may be a useful key to estimate the endocrine effects of antifoulants such as irgarol, BPA, and DEHP. In further studies, we will need to demonstrate the effects of one or more combined exposures of these EDCs in VTG protein synthesis.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3417/9/7/1401/s1, Figure S1: The relative weight (g) in M. japonicus crabs exposed to each dose of BPA, DEHP, and irgarol for 7 d. Figure S2: Transcriptional expression of VTG gene in M. japonicus hepatopancreas exposed to each dose of BPA, DEHP, and irgarol for each exposure period. Figure S3: Transcriptional expression of VTG gene in M. japonicus ovary exposed to each dose of BPA, DEHP, and irgarol for each exposure period.
Author Contributions
Conceptualization: K.P. and I.-S.K.; Methodology: K.P and I.-S.K.; Formal Analysis: K.P., H.J. and D.-K.K.; Investigation: K.P., D.-K.K. and H.J.; Resources: K.P. and I.-S.K.; Writing—Original Draft Preparation: K.P.; Writing—Review and Editing: H.J., D.-K.K. and I.-S.K.; Supervision: I.-S.K.; Project Administration: I.-S.K.; Funding Acquisition: I.-S.K.
Funding
This study was supported by the National Research Foundation of Korea, Korea, which is funded by the Korean Government (NRF-2018-R1A6A1A-03024314).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tsukimura, B.; Waddy, S.L.; Vogel, J.M.; Linder, C.J.; Bauer, D.K.; Borst, D.W. Characterization and quantification of yolk proteins in the lobster, Homarus americanus. J. Exp. Zool. 2002, 292, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xu, H.T.; Dai, Z.M.; Yang, W.J. Molecular characterization and expression analysis of vitellogenin in the marine crab Portunus trituberculatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 142, 456–464. [Google Scholar] [CrossRef]
- Okumura, T.; Yamano, K.; Sakiyama, K. Vitellogenin gene expression and hemolymph vitellogenin during vitellogenesis, final maturation, and oviposition in female kuruma prawn, Marsupenaeus japonicus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, T. Mechanisms and control of vitellogenesis in crustaceans. Fish. Sci. 2011, 77, 1–21. [Google Scholar] [CrossRef]
- Jia, X.; Chen, Y.; Zou, Z.; Lin, P.; Wang, Y.; Zhang, Z. Characterization and expression profile of Vitellogenin gene from Scylla paramamosain. Gene 2013, 520, 119–130. [Google Scholar] [CrossRef]
- Thongda, W.; Chung, J.S.; Tsutsui, N.; Zmora, N.; Katenta, A. Seasonal variations in reproductive activity of the blue crab, Callinectes sapidus: Vitellogenin expression and levels of vitellogenin in the hemolymph during ovarian development. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 179, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Matozzo, V.; Gagne, F.; Marin, M.G.; Ricciardi, F.; Blaise, C. Vitellogenin as a biomarker of exposure to estrogenic compounds in aquatic invertebrates: A review. Environ. Int. 2008, 34, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kwak, T.S.; Kwak, I.S. Vitellogenin Gene Characterization and Expression of Asian Paddle Crabs (Charybdis japonica) following Endocrine Disrupting Chemicals. Ocean Sci. J. 2014, 49, 127–135. [Google Scholar] [CrossRef]
- Jeon, J.M.; Lee, S.O.; Kim, K.S.; Baek, H.J.; Kim, S.; Kim, I.K.; Mykles, D.L.; Kim, H.W. Characterization of two vitellogenin cDNAs from a Pandalus shrimp (Pandalopsis japonica): Expression in hepatopancreas is down-regulated by endosulfan exposure. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 157, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Neris, J.B.; Olivares, D.M.M.; Velasco, F.G.; Luzardo, F.H.M.; Correia, L.O.; González, L.N. HHRISK: A code for assessment of human health risk due to environmental chemical pollution. Ecotoxicol. Environ. Saf. 2018, 170, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Shafei, A.; Ramzy, M.M.; Hegazy, A.I.; Husseny, A.K.; El-Hadary, U.G.; Taha, M.M.; Mosa, A.A. The molecular mechanisms of action of the endocrine disrupting chemical bisphenol A in the development of cancer. Gene 2018, 647, 235–243. [Google Scholar] [CrossRef]
- Kolpin, D.W.; Furlong, E.T.; Meyer, M.T.E.; Michael, T.; Zaugg, S.D.; Barber, L.B.; Buxton, H.T. Pharmaceuticals, hormones, and other organic wastewater contaminants in U.S.streams, 1999–2000: A national reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Xiang-Li, L.I.; Liang, R.J. Bisphenol A pollution of surface water and its environmental factors. J. Ecol. Rural Environ. 2009, 25, 94–97. [Google Scholar]
- Park, K.; Kwak, I.S. Characterize and Gene Expression of Heat Shock Protein 90 in Marine Crab Charybdis japonica following Bisphenol A and 4-Nonylphenol Exposures. Environ. Health Toxicol. 2014, 29, e2014002. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Gu, M.B.; Sang, B.I.; Kwack, S.J.; Kim, K.B.; Lee, B.M. Risk reduction of adverse effects due to di-(2-ethylhexyl) phthalate (DEHP) by utilizing microbial degradation. J. Toxicol. Environ. Health. A 2009, 72, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; Zhang, L.; Ma, Y.; Han, J.; Yang, L.; Zhou, B. Endocrine disruption by di-(2-ethylhexyl)-phthalate in Chinese rare minnow (Gobiocypris rarus). Environ. Toxicol. Chem. 2013, 32, 1846–1854. [Google Scholar] [CrossRef]
- Adeogun, A.O.; Ibor, O.R.; Imiuwa, M.E.; Omogbemi, E.D.; Chukwuka, A.V.; Omiwole, R.A.; Arukwe, A. Endocrine disruptor responses in African sharptooth catfish (Clarias gariepinus) exposed to di-(2-ethylhexyl)-phthalate. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 213, 7–18. [Google Scholar] [CrossRef]
- Li, X.; Yin, P.; Zhao, L. Phthalate esters in water and surface sediments of the Pearl River Estuary: Distribution, ecological, and human health risks. Environ. Sci. Pollut. Res. Int. 2016, 23, 19341–19349. [Google Scholar] [CrossRef]
- Sapozhnikova, Y.; Wirth, E.; Schiff, K.; Fulton, M. Antifouling biocides in water and sediments from California marinas. Mar. Pollut. Bull. 2013, 69, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.B.; Diamante, G.; Giroux, M.; Xu, E.G.; Abessa, D.M.S.; Schlenk, D. Changes in thyroid status of Menidia beryllina exposed to the antifouling booster irgarol: Impacts of temperature and salinity. Chemosphere 2018, 209, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, H.; Minamide, S.; Takeuchi, I. Identification and characterization of heat shock protein 90 (HSP90) in the hard coral Acropora tenuis in response to Irgarol 1051. Mar. Pollut. Bull. 2018, 133, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Konstantinou, I.K.; Albanis, T.A. Worldwide occurrence and effects of antifouling paint booster biocides in the aquatic environment: A review. Environ. Int. 2004, 30, 235–248. [Google Scholar] [CrossRef]
- Li, L.; Li, X.J.; Wu, Y.M.; Yang, L.; Li, W.; Wang, Q. Vitellogenin regulates antimicrobial responses in Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2017, 69, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, J.; Nishida, M.; Wada, K. Genetic and behavioral diversity in the Macrophthalmus japonicus species complex (Crustacea: Brachyura: Ocypodidae). Mar. Biol. 2002, 140, 1–8. [Google Scholar]
- Tanaka, Y.; Aoki, S.; Okamoto, K. Effects of the bioturbating crab Macrophthalmus japonicus on abiotic and biotic tidal mudflat characteristics in the Tama River, Tokyo Bay, Japan. Plankton. Benthos. Res. 2017, 12, 34–43. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Kim, W.S.; Park, K.; Kim, J.; Lee, M.O.; Kwak, I.S. Chitinase gene responses and tissue sensitivity in an intertidal mud crab (Macrophthalmus japonicus) following low or high salinity stress. Cell Stress Chaperones 2015, 20, 517–526. [Google Scholar] [CrossRef]
- Park, K.; Kwak, T.S.; Kim, W.S.; Kwak, I.S. Changes in exoskeleton surface roughness and expression of chitinase genes in mud crab Macrophthalmus japonicus following heavy metal differences of estuary. Mar. Pollut. Bull. 2019, 138, 11–18. [Google Scholar] [CrossRef]
- Park, K.; Nikapitiya, C.; Kwak, T.S.; Kwak, I.S. Antioxidative-related Genes Expression Following Perfluorooctane Sulfonate (PFOS) Exposure in the Intertidal Mud Crab, Macrophthalmus japonicus. Ocean Sci. J. 2015, 50, 547–556. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Kim, W.S.; Park, K.; Kwak, I.S. Identification of potential markers and sensitive tissues for low or high salinity stress in an intertidal mud crab (Macrophthalmus japonicus). Fish Shellfish Immunol. 2014, 41, 407–416. [Google Scholar] [CrossRef]
- Park, K.; Nikapitiya, C.; Kim, W.S.; Kwak, T.S.; Kwak, I.S. Changes of exoskeleton surface roughness and expression of crucial participation genes for chitin formation and digestion in the mud crab (Macrophthalmus japonicus) following the antifouling biocide irgarol. Ecotoxicol. Environ. Saf. 2016, 132, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Nikapitiya, C.; Kwak, I.S. Identification and expression of proteolysis response genes for Macrophthalmus japonicus exposure to irgarol toxicity. Ann. Limnol. Int. J. Limnol. 2016, 52, 65–74. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2_ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tiu, S.H.; Hui, H.L.; Tsukimura, B.; Tobe, S.S.; He, J.G.; Chan, S.M. Cloning and expression study of the lobster (Homarus americanus) vitellogenin: Conservation in gene structure among decapods. Gen. Comp. Endocrinol. 2009, 160, 36–46. [Google Scholar] [CrossRef]
- Anderson, T.A.; Levitt, D.G.; Banaszak, L.J. The structural basis of lipid interactions in lipovitellin, a soluble lipoprotein. Structure 1998, 6, 895–909. [Google Scholar] [CrossRef]
- Yang, J.; Sun, H.; Qian, Y.; Yang, J. Impairments of cadmium on vitellogenin accumulation in the hepatopancreas of freshwater crab Sinopotamon henanense. Environ. Sci. Pollut. Res. Int. 2017, 24, 18160–18167. [Google Scholar] [CrossRef]
- Okuno, A.; Yang, W.J.; Jayasankar, V.; Saido-Sakanaka, H.; Huong, D.T.; Jasmani, S.; Atmomarsono, M.; Subramoniam, T.; Tsutsui, N.; Ohira, T.; et al. Deduced primary structure of vitellogenin in the giant freshwater prawn, Macrobrachium rosenbergii, and yolk processing during ovarian maturation. J. Exp. Zool. 2002, 292, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Abdu, U.; Davis, C.; Khalaila, I.; Sagi, A. The vitellogenin cDNA of Cherax quadricarinatus encodes a lipoprotein with calcium binding ability, and its expression is induced following the removal of the androgenic gland in a sexually plastic system. Gen. Comp. Endocrinol. 2002, 127, 263–272. [Google Scholar] [CrossRef]
- Tsutsui, N.; Saido-Sakanaka, H.; Yang, W.J.; Jayasankar, V.; Jasmani, S.; Okuno, A.; Ohira, T.; Okumura, T.; Aida, K.; Wilder, M.N. Molecular characterization of a cDNA encoding vitellogenin in the coonstriped shrimp, Pandalus hypsinotus and site of vitellogenin mRNA expression. J. Exp. Zool. A Ecol. Genet. Physiol. 2004, 301, 802–814. [Google Scholar] [CrossRef]
- Zmora, N.; Trant, J.; Chan, S.M.; Chung, J.S. Vitellogenin and its messenger RNA during ovarian development in the female blue crab, Callinectes sapidus: Gene expression, synthesis, transport, and cleavage. Biol. Reprod. 2007, 77, 138–146. [Google Scholar] [CrossRef]
- Kang, B.J.; Nanri, T.; Lee, J.M.; Saito, H.; Han, C.H.; Hatakeyama, M.; Saigusa, M. Vitellogenesis in both sexes of gonochoristic mud shrimp, Upogebia major (Crustacea): Analyses of vitellogenin gene expression and vitellogenin processing. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 589–598. [Google Scholar] [CrossRef]
- Ding, X.; Nagaraju, G.P.C.; Novotney, D.; Lovett, D.L.; Borst, D.W. Yolk protein expression in the green crab, Carcinus maenas. Aquaculture 2010, 298, 325–331. [Google Scholar] [CrossRef]
- Allner, B.; Hennies, M.; Lerche, C.F.; Schmidt, T.; Schneider, K.; Willner, M.; Stahlschmidt-Allner, P. Kinetic determination of vitellogenin induction in the epidermis of cyprinid and perciform fishes: Evaluation of sensitive enzyme-linked immunosorbent assays. Environ. Toxicol. Chem. 2016, 35, 2916–2930. [Google Scholar] [CrossRef]
- Maradonna, F.; Nozzi, V.; Dalla Valle, L.; Traversi, I.; Gioacchini, G.; Benato, F.; Colletti, E.; Gallo, P.; Di Marco Pisciottano, I.; Mita, D.G.; et al. A developmental hepatotoxicity study of dietary bisphenol A in Sparus aurata juveniles. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 166, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, J.; Xu, P.; Yuan, C.; Qin, F.; Liu, S.; Zheng, Y.; Yang, Y.; Wang, Z. Low-dose bisphenol A disrupts gonad development and steroidogenic genes expression in adult female rare minnow Gobiocypris rarus. Chemosphere 2014, 112, 435–442. [Google Scholar] [CrossRef]
- Heindler, F.M.; Alajmi, F.; Huerlimann, R.; Zeng, C.; Newman, S.J.; Vamvounis, G.; van Herwerden, L. Toxic effects of polyethylene terephthalate microparticles and Di(2-ethylhexyl)phthalate on the calanoid copepod, Parvocalanus crassirostris. Ecotoxicol. Environ. Saf. 2017, 141, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yang, Y.; Gao, Y.; Wang, X.; Zhou, B. The impact of long term exposure to phthalic acid esters on reproduction in Chinese rare minnow (Gobiocypris rarus). Environ. Pollut. 2015, 203, 130–136. [Google Scholar] [CrossRef]
- Ye, T.; Kang, M.; Huang, Q.; Fang, C.; Chen, Y.; Shen, H.; Dong, S. Exposure to DEHP and MEHP from hatching to adulthood causes reproductive dysfunction and endocrine disruption in marine medaka (Oryzias melastigma). Aquat. Toxicol. 2014, 146, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Bao, V.W.W.; Leung, K.M.Y.; Lui, G.C.S.; Lam, M.H.W. Acute and chronic toxicities of Irgarol alone and in combination with copper to the marine copepod Tigriopus japonicus. Chemosphere 2013, 90, 1140–1148. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Hou, X.; Yue, W.; Huang, S.; Wang, C. Tissue expression profiles unveil the gene interaction of hepatopancreas, eyestalk, and ovary in the orecocious female Chinese mitten crab, Eriocheir sinensis. BMC Genet. 2019, 20, 12. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).