
Unlocking Wellness: Probiotics as Key Drivers in Functional Food Innovation and Health Promotion



Abstract
1. Introduction
2. Probiotic Strains, Sources, and Isolation Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Representative Bacteria | Reference |
|---|---|---|
| Lactobacillus | Lactobacillus gasseri, Lactobacillus fermentum, Lactobacillus delbrueckii, Lactobacillus plantarum, Lactobacillus salivarius and Lactobacillus reuteri, etc. | [23] |
| Bifidobacterium | Bifidobacterium bifidum, Bifidobacterium thermophilum, Bifidobacterium adolescentis, Bifidobacterium infantis and Bifidobacterium breve, etc. | [24] |
| Bacillus | Bacillus amyloliquefaciens, Bacillus cereus, Bacillus circulans, Bacillus licheniformis, Bacillus sphaericus, Bacillus subtilis and Bacillus thuringiensis, etc. | [25] |
| Streptococcus | Streptococcus thermophilus and Streptococcus salivarius, etc. | [26] |
| Other | Pediococcus pentosaceus, Lactococcus lactis, Saccharomyces cerevisiae and Saccharomyces boulardii, etc. | [27] |
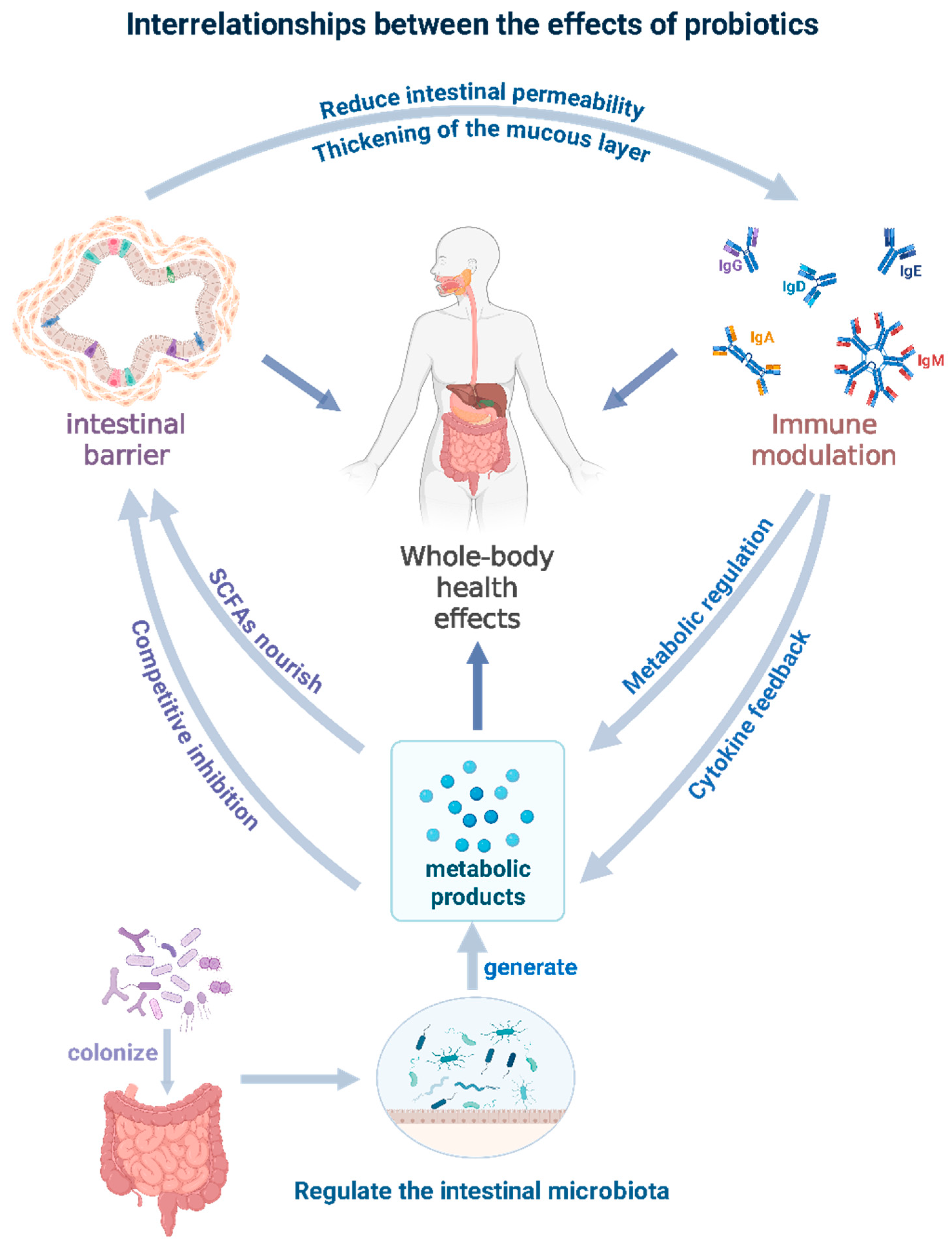
3. The Function of Probiotics
3.1. Regulating the Balance of Intestinal Flora
3.2. Enhancement of Intestinal Barrier Function
3.3. Immunomodulatory Function
3.4. Metabolic Disease Intervention
3.5. The Synergistic Effect of Probiotics and Prebiotics
4. Application of Probiotics in Functional Foods
4.1. Dairy Products
4.1.1. Probiotic Fermented Milk
4.1.2. Probiotic Fermented Yogurt
| Probiotic Fermented Milk Products | Tested Strains | Reference |
|---|---|---|
| High-protein drink, pure probiotic fermented milk | Lactobacillus acidophilus, Bifidobacterium lactis, Lactobacillus paracasei, Streptococcus thermophilus | [95] |
| Compound lactobacillus beverage | Lactobacillus bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus and Bifidobacterium | [96] |
| Probiotic fermented milk with compound fruit and vegetable juice | Lactobacillus bulgaricus, Streptococcus thermophilus | [97] |
| Probiotic fermented goat milk | Acidophilus, Lactobacillus casei, Lactobacillus paracasei 01, Lactobacillus paracasei 431 | [98] |
| Double-protein probiotic yogurt | Lactobacillus bulgaricus, Streptococcus thermophilus, Bifidobacterium | [99] |
| Probiotic yogurt ice cream | Streptococcus thermophilus DMST-H2, Lactobacillus delbrueckii DMLD-H1 | [100] |
| Compound probiotic fermented milk | Lactobacillus paracasei PC-01, Bifidobacterium Lactobacillus Probio-M8 | [101] |
4.2. Plant-Based and Fruit and Vegetable Products
4.2.1. Probiotic Fermentation of Fruit and Vegetable Juice
4.2.2. Probiotic Fermented Soy Products
4.3. Meat Products
4.4. Other Functional Foods
4.4.1. Application of Probiotics in Health Products
4.4.2. Application of Probiotics in Food Additives
5. Technological Innovation of Probiotics in Functional Foods
5.1. Microencapsulation Technology
5.2. Biofilm Delivery Systems
5.3. Multidimensional Environmental Adaptation Techniques
5.4. Genetic Engineering
5.5. Personalized Strain Screening
6. Safety and Evaluation of Probiotics
6.1. Safety Risks of Probiotics
6.2. Safety Evaluation System of Probiotics
7. Final Considerations and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goldin, B.R. Health benefits of probiotics. Br. J. Nutr. 1998, 80, S203–S207. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [CrossRef] [PubMed]
- Onubi, O.J.; Poobalan, A.S.; Dineen, B.; Marais, D.; McNeill, G. Effects of probiotics on child growth: A systematic review. J. Health Popul. Nutr. 2015, 34, 8. [Google Scholar] [CrossRef] [PubMed]
- Essa, M.M.; Bishir, M.; Bhat, A.; Chidambaram, S.B.; Al-Balushi, B.; Hamdan, H.; Govindarajan, N.; Freidland, R.P.; Qoronfleh, M.W. Functional foods and their impact on health. J. Food Sci. Technol. 2023, 60, 820–834. [Google Scholar] [CrossRef]
- Boggia, R.; Zunin, P.; Turrini, F. Functional foods and food supplements. Appl. Sci. 2020, 10, 8538. [Google Scholar] [CrossRef]
- Fentie, E.G.; Lim, K.; Jeong, M.; Shin, J.-H. A comprehensive review of the characterization, host interactions, and stabilization advancements on probiotics: Addressing the challenges in functional food diversification. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13424. [Google Scholar] [CrossRef]
- Chakravarty, K.; Gaur, S.; Kumar, R.; Jha, N.K.; Gupta, P.K. Exploring the multifaceted therapeutic potential of probiotics: A review of current insights and applications. Probiot. Antimicrob. Proteins 2025, 17, 341–363. [Google Scholar] [CrossRef]
- Ray, M.; Manjunath, A.; Halami, P.M. Effect of probiotics as an immune modulator for the management of COVID-19. Arch. Microbiol. 2023, 205, 182. [Google Scholar] [CrossRef]
- Penha Rodrigues Pereira, E.; Silva da Graça, J.; Manfrinato Ferreira, B.; Fasura Balthazar, C.; Xavier-Santos, D.; França Bezerril, F.; Magnani, M.; Sant’Ana, A.S. What are the main obstacles to turning foods healthier through probiotics incorporation? A review of functionalization of foods by probiotics and bioactive metabolites. Food Res. Int. 2024, 176, 113785. [Google Scholar] [CrossRef]
- Juárez-Fernández, M.; Porras, D.; García-Mediavilla, M.V.; Román-Sagüillo, S.; González-Gallego, J.; Nistal, E.; Sánchez-Campos, S. Aging, gut microbiota and metabolic diseases: Management through physical exercise and nutritional interventions. Nutrients 2021, 13, 16. [Google Scholar] [CrossRef]
- Kocsis, T.; Molnár, B.; Németh, D.; Hegyi, P.; Szakács, Z.; Bálint, A.; Garami, A.; Soós, A.; Márta, K.; Solymár, M. Probiotics have beneficial metabolic effects in patients with type 2 diabetes mellitus: A meta-analysis of randomized clinical trials. Sci. Rep. 2020, 10, 11787. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Romero, T.; Flores-Andrade, E.; Bonilla-Zavaleta, E.; González-Arnao, M.T.; Rascón-Díaz, M.P. Probiotic microencapsulation at low temperatures. Cryobiology 2019, 91, 173–174. [Google Scholar] [CrossRef]
- Han, J.; McClements, D.J.; Liu, X.; Liu, F. Oral delivery of probiotics using single-cell encapsulation. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13322. [Google Scholar] [CrossRef] [PubMed]
- Lorena, C.; Imen, N. Taxonomy and systematics of plant probiotic bacteria in the genomic era. AIMS Microbiol. 2017, 3, 383–412. [Google Scholar]
- Papizadeh, M.; Rohani, M.; Nahrevanian, H.; Javadi, A.; Pourshafie, M.R. Probiotic characters of Bifidobacterium and Lactobacillus are a result of the ongoing gene acquisition and genome minimization evolutionary trends. Microb. Pathog. 2017, 111, 118–131. [Google Scholar] [CrossRef]
- Thullner, M.; Regnier, P. Microbial Controls on the biogeochemical dynamics in the subsurface. Rev. Mineral. Geochem. 2019, 85, 265–302. [Google Scholar] [CrossRef]
- Matchado, M.S.; Rühlemann, M.; Reitmeier, S.; Kacprowski, T.; Frost, F.; Haller, D.; Baumbach, J.; List, M. On the limits of 16S rRNA gene-based metagenome prediction and functional profiling. Microb. Genom. 2024, 10, 001203. [Google Scholar] [CrossRef]
- Arikawa, K.; Ide, K.; Kogawa, M.; Saeki, T.; Yoda, T.; Endoh, T.; Matsuhashi, A.; Takeyama, H.; Hosokawa, M. Recovery of strain-resolved genomes from human microbiome through an integration framework of single-cell genomics and metagenomics. Microbiome 2021, 9, 202. [Google Scholar] [CrossRef]
- Cardona, S.; Mostafazadeh, N.; Luan, Q.; Zhou, J.; Peng, Z.; Papautsky, I. Numerical modeling of physical cell trapping in microfluidic chips. Micromachines 2023, 14, 1665. [Google Scholar] [CrossRef]
- Pereira, A.C.; Tenreiro, A.; Cunha, M.V. When FLOW-FISH met FACS: Combining multiparametric, dynamic approaches for microbial single-cell research in the total environment. Sci. Total Environ. 2022, 806, 150682. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Jiang, T.; Yan, Y. Biosensor-assisted titratable CRISPRi high-throughput (BATCH) screening for over-production phenotypes. Metab. Eng. 2023, 75, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Song, H.S.; Kim, Y.B.; Kim, J.Y.; Roh, S.W.; Whon, T.W. Advances in culturomics research on the human gut microbiome: Optimizing medium composition and culture techniques for enhanced microbial discovery. J. Microbiol. Biotechnol. 2024, 34, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Heczko, P.B.; Giemza, M.; Ponikiewska, W.; Strus, M. Importance of Lactobacilli for human health. Microorganisms 2024, 12, 2382. [Google Scholar] [CrossRef] [PubMed]
- Lugli, G.A.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Turroni, F.; Viappiani, A.; Sinderen, D.v.; Ventura, M. Tracking the taxonomy of the genus Bifidobacterium based on a phylogenomic approach. Appl. Environ. Microbiol. 2018, 84, e02249-17. [Google Scholar] [CrossRef]
- Gopikrishna, T.; Suresh Kumar, H.K.; Perumal, K.; Elangovan, E. Impact of Bacillus in fermented soybean foods on human health. Ann. Microbiol. 2021, 71, 1–16. [Google Scholar] [CrossRef]
- Martinovic, A.; Cocuzzi, R.; Arioli, S.; Mora, D. Streptococcus thermophilus: To survive, or not to survive the gastrointestinal tract, that is the question! Nutrients 2020, 12, 2175. [Google Scholar] [CrossRef]
- Qin, H.; Wu, H.; Shen, K.; Liu, Y.; Li, M.; Wang, H.; Qiao, Z.; Mu, Z. Fermented minor grain foods: Classification, functional components, and probiotic potential. Foods 2022, 11, 3155. [Google Scholar] [CrossRef]
- Tang, C.; Lu, Z. Health promoting activities of probiotics. J. Food Biochem. 2019, 43, e12944. [Google Scholar] [CrossRef]
- Knipe, H.; Temperton, B.; Lange, A.; Bass, D.; Tyler, C.R. Probiotics and competitive exclusion of pathogens in shrimp aquaculture. Rev. Aquac. 2021, 13, 324–352. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Nie, X.; Chitrakar, B.; Gao, J.; Sang, Y. Mutual adhesion of Lactobacillus spp. to intestinal cells: A review of perspectives on surface layer proteins and cell surface receptors. Int. J. Biol. Macromol. 2024, 282, 137031. [Google Scholar] [CrossRef]
- Pan, C.; Jiang, X.; Wei, J.; Liu, C.; Zhang, M.; Gao, C.; Chen, R.; Yang, C.; Wang, B.; Yu, M.; et al. Ameba-inspired strategy enhances probiotic efficacy via prebound nutrient supply. Nat. Commun. 2025, 16, 1827. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, S.; Liao, C.; Jia, Y.; Li, J.; Shang, K.; Chen, J.; Cao, P.; Li, W.; Li, Y.; et al. Probiotic properties of chicken-derived highly adherent lactic acid bacteria and inhibition of enteropathogenic bacteria in Caco-2 Cells. Microorganisms 2022, 10, 2515. [Google Scholar] [CrossRef] [PubMed]
- Çelen, T.; Anumudu, C.; Miri, T.; Onyeaka, H.; Fernandez-Trillo, P. Pathogen-responsive delivery of nisin. Food Hydrocoll. 2024, 154, 110076. [Google Scholar] [CrossRef]
- Yang, S.M.; Lee, D.W.; Park, H.J.; Kwak, M.H.; Park, J.M.; Choi, M.-G. Hydrogen peroxide enhances the antibacterial effect of methylene blue-based photodynamic therapy on biofilm-forming bacteria. Photochem. Photobiol. 2019, 95, 833–838. [Google Scholar] [CrossRef]
- Facchin, S.; Bertin, L.; Bonazzi, E.; Lorenzon, G.; De Barba, C.; Barberio, B.; Zingone, F.; Maniero, D.; Scarpa, M.; Ruffolo, C.; et al. Short-chain fatty acids and human health: From metabolic pathways to current therapeutic implications. Life 2024, 14, 559. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef]
- Nayebhashemi, M.; Enayati, S.; Zahmatkesh, M.; Madanchi, H.; Saberi, S.; Mostafavi, E.; Mirbzadeh Ardakani, E.; Azizi, M.; Khalaj, V. Surface display of pancreatic lipase inhibitor peptides by engineered Saccharomyces boulardii: Potential as an anti-obesity probiotic. J. Funct. Foods 2023, 102, 105458. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, Q.; Qi, R.; Wang, J.; Qiu, X.; Liu, Z.; Huang, J. Effects of Lactobacillus plantarum on the intestinal morphology, intestinal barrier function and microbiota composition of suckling piglets. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1908–1918. [Google Scholar] [CrossRef]
- Pridmore, R.D.; Pittet, A.-C.; Praplan, F.; Cavadini, C. Hydrogen peroxide production by Lactobacillus johnsonii NCC 533 and its role in anti-Salmonella activity. FEMS Microbiol. Lett. 2008, 283, 210–215. [Google Scholar] [CrossRef]
- Prado-Rebolledo, O.F.; Delgado-Machuca, J.d.J.; Macedo-Barragan, R.J.; Garcia-Marquez, L.J.; Morales-Barrera, J.E.; Latorre, J.D.; Hernandez-Velasco, X.; Tellez, G. Evaluation of a selected lactic acid bacteria-based probiotic on Salmonella enterica serovar Enteritidis colonization and intestinal permeability in broiler chickens. Avian Pathol. 2017, 46, 90–94. [Google Scholar] [CrossRef]
- Zhou, K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. J. Funct. Foods 2017, 33, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Kakuta, S.; Aihara, Y.; Kamiya, T.; Watanabe, Y.; Osakabe, N.; Hazato, N.; Miyawaki, A.; Yoshikawa, S.; Usami, T.; et al. Visualization of probiotic-mediated Ca2+ signaling in intestinal epithelial cells in vivo. Front. Immunol. 2016, 7, 601. [Google Scholar] [CrossRef] [PubMed]
- Vangay, P.; Ward, T.; Lucas, S.; Beura, L.K.; Sabas, D.; Abramson, M.; Till, L.; Hoops, S.L.; Kashyap, P.; Hunter, R.C.; et al. Industrialized human gut microbiota increases CD8+T cells and mucus thickness in humanized mouse gut. Gut Microbes 2023, 15, 2266627. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Song, J.; Lan, X.; Ma, F.; Jiang, M.; Jiang, C. Calcium-sensitive receptors alters intestinal microbiota metabolites especially SCFAs and ameliorates intestinal barrier damage in neonatal rat endotoxemia. Infect. Drug Resist. 2023, 16, 5707–5717. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, S.; Thaker, H.; Dong, M. Shiga toxins: An update on host factors and biomedical applications. Toxins 2021, 13, 222. [Google Scholar] [CrossRef]
- Fagan, R.P.; Fairweather, N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 2014, 12, 211–222. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Lasaro, M.A.; Salinger, N.; Zhang, J.; Wang, Y.; Zhong, Z.; Goulian, M.; Zhu, J. F1C fimbriae play an important role in biofilm formation and intestinal colonization by the Escherichia coli commensal strain Nissle 1917. Appl. Environ. Microbiol. 2009, 75, 246–251. [Google Scholar] [CrossRef]
- Nzakizwanayo, J.; Kumar, S.; Ogilvie, L.A.; Patel, B.A.; Dedi, C.; Macfarlane, W.M.; Jones, B.V. Disruption of Escherichia coli Nissle 1917 K5 capsule biosynthesis, through loss of distinct kfi genes, modulates interaction with intestinal epithelial cells and impact on cell health. PLoS ONE 2015, 10, e0120430. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Fact. 2020, 19, 23. [Google Scholar] [CrossRef]
- Wiertsema, S.P.; van Bergenhenegouwen, J.; Garssen, J.; Knippels, L.M.J. The interplay between the gut microbiome and the immune system in the context of infectious diseases throughout life and the role of nutrition in optimizing treatment strategies. Nutrients 2021, 13, 886. [Google Scholar] [CrossRef] [PubMed]
- Benjamin-Neelon, S.E. Position of the academy of nutrition and dietetics: Benchmarks for nutrition in child care. J. Acad. Nutr. Diet. 2018, 118, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Perdigon, G.; Alvarez, S.; Rachid, M.; Aguero, G.; Gobbato, N. Immune system stimulation by probiotics. J. Dairy Sci. 1995, 78, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xie, M.; Liu, Y. TLR2 promotes the progression of diabetes mellitus with atherosclerosis via activating NLRP3 inflammasome and MyD88/NF-κB signaling pathway. Sci. Rep. 2025, 15, 16348. [Google Scholar] [CrossRef]
- Shoji, S.; Hanada, K.; Takahashi, M.; Watanabe, K.; Yonemochi, M.; Tomabechi, Y.; Shirouzu, M. The NF-κB regulator IκBβ exhibits different molecular interactivity and phosphorylation status from IκBα in an IKK2-catalysed reaction. FEBS Lett. 2020, 594, 1532–1549. [Google Scholar] [CrossRef]
- Vincenzi, A.; Goettert, M.I.; Volken de Souza, C.F. An evaluation of the effects of probiotics on tumoral necrosis factor (TNF-α) signaling and gene expression. Cytokine Growth Factor Rev. 2021, 57, 27–38. [Google Scholar] [CrossRef]
- Chae, J.M.; Heo, W.; Cho, H.T.; Lee, D.H.; Kim, J.H.; Rhee, M.S.; Park, T.-S.; Kim, Y.K.; Lee, J.H.; Kim, Y.J. Effects of orally-administered Bifidobacterium animalis subsp lactis strain BB12 on dextran sodium sulfate-induced colitis in mice. J. Microbiol. Biotechnol. 2018, 28, 1800–1805. [Google Scholar] [CrossRef]
- Watanabe, N.; Kaminuma, O.; Kitamura, N.; Hiroi, T. Induced treg cells augment the Th17-mediated intestinal inflammatory response in a CTLA4-dependent manner. PLoS ONE 2016, 11, e0150244. [Google Scholar] [CrossRef]
- Tang, C.; Kong, L.; Shan, M.; Lu, Z.; Lu, Y. Protective and ameliorating effects of probiotics against diet-induced obesity: A review. Food Res. Int. 2021, 147, 110490. [Google Scholar] [CrossRef]
- Koutnikova, H.; Genser, B.; Monteiro-Sepulveda, M.; Faurie, J.-M.; Rizkalla, S.; Schrezenmeir, J.; Clément, K. Impact of bacterial probiotics on obesity, diabetes and non-alcoholic fatty liver disease related variables: A systematic review and meta-analysis of randomised controlled trials. BMJ Open 2019, 9, e017995. [Google Scholar] [CrossRef]
- van der Beek, C.M.; Canfora, E.E.; Kip, A.M.; Gorissen, S.H.M.; Olde Damink, S.W.M.; van Eijk, H.M.; Holst, J.J.; Blaak, E.E.; Dejong, C.H.C.; Lenaerts, K. The prebiotic inulin improves substrate metabolism and promotes short-chain fatty acid production in overweight to obese men. Metabolism 2018, 87, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xiao, K.-l.; Chen, L.; Wu, C.; Hu, X.; Zeng, T.; Chen, X.-q.; Li, W.-j.; Deng, X.; Li, H.; et al. Insulin sensitizers improve the GLP-1 secretion and the amount of intestinal L cells on high-fat-diet–induced catch-up growth. Nutrition 2017, 39–40, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-R.; Singhania, R.R.; Patel, A.K.; Tsai, T.-R.; Tsai, M.-L.; Sharma, V.; Dong, C.-D. Novel food isolates with striking α-glucosidase inhibitory activity and probiotic potential for an antidiabetic role. J. Food Sci. Technol. 2024, 61, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Duseja, A.; Acharya, S.K.; Mehta, M.; Chhabra, S.; Shalimar; Rana, S.; Das, A.; Dattagupta, S.; Dhiman, R.K.; Chawla, Y.K. High potency multistrain probiotic improves liver histology in non-alcoholic fatty liver disease (NAFLD): A randomised, double-blind, proof of concept study. BMJ Open Gastroenterol. 2019, 6, e000315. [Google Scholar] [CrossRef]
- Mishima, E.; Abe, T. Role of the microbiota in hypertension and antihypertensive drug metabolism. Hypertens. Res. 2022, 45, 246–253. [Google Scholar] [CrossRef]
- Saleh, M.A.; Norlander, A.E.; Madhur, M.S. Inhibition of interleukin-17a, but not interleukin-17f, signaling lowers blood pressure, and reduces end-organ inflammation in angiotensin ii–induced hypertension. JACC Basic. Transl. Sci. 2016, 1, 606–616. [Google Scholar] [CrossRef]
- Krege, J.H.; Moyer, J.S.; Langenbach, L.L.; Peng, L.; Zhang, S.H.; Maeda, N.; Reddick, R.L.; Smithies, O. Angiotensin-converting enzyme gene and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1245–1250. [Google Scholar] [CrossRef]
- Deehan, E.C.; Al Antwan, S.; Witwer, R.S.; Guerra, P.; John, T.; Monheit, L. Revisiting the concepts of prebiotic and prebiotic effect in light of scientific and regulatory progress—A consensus paper from the global prebiotic association. Adv. Nutr. 2024, 15, 100329. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Tsigalou, C.; Dalamaga, M. Probiotics, prebiotics, synbiotics, postbiotics, and obesity: Current evidence, controversies, and perspectives. Curr. Obes. Rep. 2020, 9, 179–192. [Google Scholar] [CrossRef]
- Sharma, R.; Padwad, Y. Plant-polyphenols based second-generation synbiotics: Emerging concepts, challenges, and opportunities. Nutrition 2020, 77, 110785. [Google Scholar] [CrossRef]
- Chunchai, T.; Thunapong, W.; Yasom, S.; Wanchai, K.; Eaimworawuthikul, S.; Metzler, G.; Lungkaphin, A.; Pongchaidecha, A.; Sirilun, S.; Chaiyasut, C.; et al. Decreased microglial activation through gut-brain axis by prebiotics, probiotics, or synbiotics effectively restored cognitive function in obese-insulin resistant rats. J. Neuroinflamm. 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; O’Riordan, K.J.; Clarke, G.; Cryan, J.F. Feeding gut microbes to nourish the brain: Unravelling the diet–microbiota–gut–brain axis. Nat. Metab. 2024, 6, 1454–1478. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Catapano, A.; Trinchese, G.; Cimmino, F.; Penna, E.; Pizzella, A.; Cristiano, C.; Lama, A.; Crispino, M.; Mollica, M.P. Butyrate improves neuroinflammation and mitochondrial impairment in cerebral cortex and synaptic fraction in an animal model of diet-induced obesity. Antioxidants 2023, 12, 4. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Heidari-soureshjani, R.; Mirhosseini, N.; Kouchaki, E.; Bahmani, F.; Aghadavod, E.; Tajabadi-Ebrahimi, M.; Asemi, Z. Probiotic and selenium co-supplementation, and the effects on clinical, metabolic and genetic status in Alzheimer’s disease: A randomized, double-blind, controlled trial. Clin. Nutr. 2019, 38, 2569–2575. [Google Scholar] [CrossRef]
- Liu, L.; Yang, D.; Liu, H.; Guo, W.; Jiang, Z.; Han, Q.; Liu, Y. Proliferation of probiotics and antioxidant effects of functional oligosaccharides added in fermented dairy product. Int. J. Dairy Technol. 2024, 77, 893–904. [Google Scholar] [CrossRef]
- Azhar, M.A.; Munaim, M.S.A. Design and optimization of a probiotic tablet for gastrointestinal tolerance by a simplex-centroid mixture. Drug Dev. Ind. Pharm. 2020, 47, 1–8. [Google Scholar] [CrossRef]
- Balaghi, Z.; Azima, S.; Motamedifar, M.; Kaviani, M.; Poordast, T.; Zare, N. The effect of lactofem oral probiotic capsule on Lactobacilli colonization and some vaginal health parameters. Gynecol. Obstet. Investig. 2020, 85, 245–251. [Google Scholar] [CrossRef]
- Talwalkar, A.; Miller, C.W.; Kailasapathy, K.; Nguyen, M.H. Effect of packaging materials and dissolved oxygen on the survival of probiotic bacteria in yoghurt. Int. J. Food Sci. Technol. 2010, 39, 605–611. [Google Scholar] [CrossRef]
- Sornsenee, P.; Chimplee, S.; Saengsuwan, P.; Romyasamit, C. Characterization of probiotic properties and development of banana powder enriched with freeze-dried Lacticaseibacillus paracasei probiotics. Heliyon 2022, 8, e11063. [Google Scholar] [CrossRef]
- Zhou, J.S.; Gopal, P.K.; Gill, H.S. Potential probiotic lactic acid bacteria Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019) do not degrade gastric mucin in vitro. Int. J. Food Microbiol. 2001, 63, 81–90. [Google Scholar] [CrossRef]
- Alansari, M.M.; Sahlah, S.A.; Alhumaid, L.; Singh, A.J.R. Probiotic lactobacilli: Can be a remediating supplement for pandemic COVID-19. A review. J. King Saud Univ. Sci. 2020, 33, 101286. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Branco, G.F.; Cruz, A.G.; Fonseca Faria, J.d.A.; Shah, N.P. Probiotic dairy products as functional foods. Compr. Rev. Food Sci. Food Saf. 2010, 9, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Szajnar, K.; Znamirowska, A.; Kuzniar, P. Sensory and textural properties of fermented milk with viability of Lactobacillus rhamnosus and Bifidobacterium animalis ssp. lactis Bb-12 and increased calcium concentration. Int. J. Food Prop. 2020, 23, 582–598. [Google Scholar] [CrossRef]
- Martínez, M.P.; Magnoli, A.P.; González Pereyra, M.L.; Cavaglieri, L. Probiotic bacteria and yeasts adsorb aflatoxin M1 in milk and degrade it to less toxic AFM1-metabolites. Toxicon 2019, 172, 1–7. [Google Scholar] [CrossRef]
- Sanaldi, K.; Coban, A.Y. Detoxification of aflatoxin M1 in different milk types using probiotics. An. Acad. Bras. Cienc. 2023, 95, e20220794. [Google Scholar] [CrossRef]
- Gunenc, A.; Khoury, C.; Legault, C.; Mirrashed, H.; Rijke, J.; Hosseinian, F. Seabuckthorn as a novel prebiotic source improves probiotic viability in yogurt. LWT Food Sci. Technol. 2016, 66, 490–495. [Google Scholar] [CrossRef]
- Mani-Lopez, E.; Palou, E.; Lopez-Malo, A. Probiotic viability and storage stability of yogurts and fermented milks prepared with several mixtures of lactic acid bacteria. J. Dairy Sci. 2014, 97, 2578–2590. [Google Scholar] [CrossRef]
- Laird, E.; Molloy, A.M.; McNulty, H.; Ward, M.; McCarroll, K.; Hoey, L.; Hughes, C.F.; Cunningham, C.; Strain, J.J.; Casey, M.C. Greater yogurt consumption is associated with increased bone mineral density and physical function in older adults. Osteoporos. Int. 2017, 28, 2409–2419. [Google Scholar] [CrossRef]
- Saleem, G.N.; Gu, R.; Qu, H.; Bahar Khaskheli, G.; Rashid Rajput, I.; Qasim, M.; Chen, X. Therapeutic potential of popular fermented dairy products and its benefits on human health. Front. Nutr. 2024, 11, 1328620. [Google Scholar] [CrossRef]
- Gijsbers, L.; Ding, E.L.; Malik, V.S.; de Goede, J.; Geleijnse, J.M.; Soedamah-Muthu, S.S. Consumption of dairy foods and diabetes incidence: A dose-response meta-analysis of observational studies. Am. J. Clin. Nutr. 2016, 103, 1111–1124. [Google Scholar] [CrossRef]
- Farag, M.A.; Saleh, H.A.; El Ahmady, S.; Elmassry, M.M. Dissecting yogurt: The impact of milk types, probiotics, and selected additives on yogurt quality. Food Rev. Int. 2022, 38, 634–650. [Google Scholar] [CrossRef]
- Brodziak, A.; Król, J.; Matwijczuk, A.; Czernecki, T.; Glibowski, P.; Wlazło, Ł.; Litwińczuk, A. Effect of sea buckthorn (Hippophae rhamnoides L.) mousse on properties of probiotic yoghurt. Appl. Sci. 2021, 11, 545. [Google Scholar] [CrossRef]
- Ghafarloo, M.H.; Jouki, M.; Tabari, M. Production and characterization of synbiotic doogh, a yogurt-based Iranian drink by gum arabic, ginger extract and B. bifidum. J. Food Sci. Technol. 2020, 57, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Terpou, A.; Papadaki, A.; Bosnea, L.; Kanellaki, M.; Kopsahelis, N. Novel frozen yogurt production fortified with sea buckthorn berries and probiotics. LWT Food Sci. Technol. 2019, 105, 242–249. [Google Scholar] [CrossRef]
- da Cruz, M.F.; Rocha, R.S.; Silva, R.; Freitas, M.Q.; Pimentel, T.C.; Esmerino, E.A.; Cruz, A.G.; Fidalgo, T.K.d.S.; Maia, L.C. Probiotic fermented milks: Children’s emotional responses using a product-specific emoji list. Food Res. Int. 2021, 143, 110269. [Google Scholar] [CrossRef]
- Li, S.; Tang, S.; He, Q.; Hu, J.; Zheng, J. Changes in proteolysis in fermented milk produced by Streptococcus thermophilus in co-culture with Lactobacillus plantarum or Bifidobacterium animalis subsp. lactis during refrigerated storage. Molecules 2019, 24, 3699. [Google Scholar] [CrossRef]
- Barat, A.; Ozcan, T. Growth of probiotic bacteria and characteristics of fermented milk containing fruit matrices. Int. J. Dairy Technol. 2018, 71, 120–129. [Google Scholar] [CrossRef]
- Mituniewicz-Malek, A.; Zielinska, D.; Ziarno, M. Probiotic monocultures in fermented goat milk beverages—Sensory quality of final product. Int. J. Dairy Technol. 2019, 72, 240–247. [Google Scholar] [CrossRef]
- Soni, R.; Jain, N.K.; Shah, V.; Soni, J.; Suthar, D.; Gohel, P. Development of probiotic yogurt: Effect of strain combination on nutritional, rheological, organoleptic and probiotic properties. J. Food Sci. Technol. 2020, 57, 2038–2050. [Google Scholar] [CrossRef]
- Soukoulis, C.; Lyroni, E.; Tzia, C. Sensory profiling and hedonic judgement of probiotic ice cream as a function of hydrocolloids, yogurt and milk fat content. LWT Food Sci. Technol. 2010, 43, 1351–1358. [Google Scholar] [CrossRef]
- Hao, Y.; Wu, T.; Guo, S.; Kwok, L.-Y.; Zhang, H.; Wang, J. Metabolic dynamics of fermented milk beverages co-fermented with Bifidobacterium animalis subsp. lactis Probio-M8 and Lacticaseibacillus paracasei PC-01 during storage. LWT Food Sci. Technol. 2023, 185, 115196. [Google Scholar] [CrossRef]
- Timmermans, R.A.H.; Nederhoff, A.L.; Groot, M.N.N.; van Boekel, M.A.J.S.; Mastwijk, H.C. Effect of electrical field strength applied by PEF processing and storage temperature on the outgrowth of yeasts and moulds naturally present in a fresh fruit smoothie. Int. J. Food Microbiol. 2016, 230, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Luo, D.; Bao, Y.; Liao, X.; Wu, J. Characterization of diversity and probiotic efficiency of the autochthonous lactic acid bacteria in the fermentation of selected raw fruit and vegetable juices. Front. Microbiol. 2018, 9, 2539. [Google Scholar] [CrossRef] [PubMed]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Szutowska, J.; Rybicka, I.; Pawlak-Lemanska, K.; Gwiazdowska, D. Spontaneously fermented curly kale juice: Microbiological quality, nutritional composition, antioxidant, and antimicrobial properties. J. Food Sci. 2020, 85, 1248–1255. [Google Scholar] [CrossRef]
- Mantzourani, I.; Nikolaou, A.; Kourkoutas, Y.; Alexopoulos, A.; Dasenaki, M.; Mastrotheodoraki, A.; Proestos, C.; Thomaidis, N.; Plessas, S. Chemical profile characterization of fruit and vegetable juices after fermentation with probiotic strains. Foods 2024, 13, 1136. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Yang, J.; Li, X.; Lin, D.; Wang, M. Fermentation broth from fruit and vegetable waste works: Reducing the risk of human bacterial pathogens in soil by inhibiting quorum sensing. Environ. Int. 2024, 188, 108753. [Google Scholar] [CrossRef]
- Rizzi, F.; Juan, B.; Espadaler-Mazo, J.; Capellas, M.; Huedo, P. Lactiplantibacillus plantarum KABP051: Stability in fruit juices and production of bioactive compounds during their fermentation. Foods 2024, 13, 3851. [Google Scholar] [CrossRef]
- Quan, Q.; Liu, W.; Guo, J.; Ye, M.; Zhang, J. Effect of six lactic acid bacteria strains on physicochemical characteristics, antioxidant activities and sensory properties of fermented orange juices. Foods 2022, 11, 1920. [Google Scholar] [CrossRef]
- Golestani, M.; Pourahmad, R. Comparison of three treatments (two fermented treatments and one nonfermented treatment) in production of synbiotic ice cream. J. Food Process. Preserv. 2017, 41, e12839. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mortazavian, A.M. Review article: Technological aspects of prebiotics in probiotic fermented milks. Food Rev. Int. 2011, 27, 192–212. [Google Scholar] [CrossRef]
- Jeminiwa, B.O.; Knight, R.M.; Braden, T.D.; Cruz-espindola, C.; Boothe, D.M.; Akingbemi, B.T. Regulation of the neuroendocrine axis in male rats by soy-based diets is independent of age and due specifically to isoflavone action. Biol. Reprod. 2020, 103, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Kano, M.; Kaga, C. Possibility of breast cancer prevention: Use of soy isoflavones and fermented soy beverage produced using probiotics. Int. J. Mol. Sci. 2015, 16, 10907–10920. [Google Scholar] [CrossRef]
- Lampe, B.J.; English, J.C. Toxicological assessment of nattokinase derived from Bacillus subtilis var. natto. Food Chem. Toxicol. 2016, 88, 87–99. [Google Scholar] [CrossRef]
- Maslarov, D.; Drenska, D.; Maslarova-Gelov, J.; Gelov, I. Understanding the concept of Nattokinase use: A few years after beginning. Biotechnol. Biotechnol. Equip. 2023, 37, 2249552. [Google Scholar] [CrossRef]
- Kim, B.; Byun, B.Y.; Mah, J.-H. Biogenic amine formation and bacterial contribution in Natto products. Food Chem. 2012, 135, 2005–2011. [Google Scholar] [CrossRef]
- Lan, G.; Li, C.; He, L.; Zeng, X.; Zhu, Q. Effects of different strains and fermentation method on nattokinase activity, biogenic amines, and sensory characteristics of natto. J. Food Sci. Technol. 2020, 57, 4414–4423. [Google Scholar] [CrossRef]
- Ma, X.; Liu, Z.; He, Y.; Zhou, Y. Process optimization of co-fermentation natto with Lactobacillus bulgaricus and characteristic analysis. J. Food Sci. Technol. 2024, 62, 716–726. [Google Scholar] [CrossRef]
- Qian, W. Study on the Fermentation of Sausage with Composite Strains. Ph.D. Thesis, Jilin Agricultural University, Jilin, China, 2013. [Google Scholar]
- Gong, Y.; Sun, X.; Zhang, L.; Guo, Y.J.; Li, J.; Yang, Y.; Li, C.; Hu, B.; He, L. Processing technology of low-acidity Sichuan-flavor sausage produced by mixed-strain fermentation. Food Ind. Technol. 2015, 36, 227–232+239. [Google Scholar]
- Rubio, R.; Jofré, A.; Martín, B.; Aymerich, T.; Garriga, M. Characterization of lactic acid bacteria isolated from infant faeces as potential probiotic starter cultures for fermented sausages. Food Microbiol. 2014, 38, 303–311. [Google Scholar] [CrossRef]
- Bis-Souza, C.V.; Barretto Penna, A.L.; da Silva Barretto, A.C. Applicability of potentially probiotic Lactobacillus casei in low-fat Italian type salami with added fructooligosaccharides: In vitro screening and technological evaluation. Meat Sci. 2020, 168. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Corral, S.; Cano-Garcia, L.; Salvador, A.; Belloch, C. Yeast strains as potential aroma enhancers in dry fermented sausages. Int. J. Food Microbiol. 2015, 212, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional foods and nondairy probiotic food development: Trends, concepts, and products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Marinova, V.Y.; Rasheva, I.K.; Kizheva, Y.K.; Dermenzhieva, Y.D.; Hristova, P.K. Microbiological quality of probiotic dietary supplements. Biotechnol. Biotechnol. Equip. 2019, 33, 834–841. [Google Scholar] [CrossRef]
- Saxelin, M.; Lassig, A.; Karjalainen, H.; Tynkkynen, S.; Surakka, A.; Vapaatalo, H.; Jarvenpaa, S.; Korpela, R.; Mutanen, M.; Hatakka, K. Persistence of probiotic strains in the gastrointestinal tract when administered as capsules, yoghurt, or cheese. Int. J. Food Microbiol. 2010, 144, 293–300. [Google Scholar] [CrossRef]
- Xu, C.; Ban, Q.; Wang, W.; Hou, J.; Jiang, Z. Novel nano-encapsulated probiotic agents: Encapsulate materials, delivery, and encapsulation systems. J. Control. Release 2022, 349, 184–205. [Google Scholar] [CrossRef]
- Castro, A.; Aleman, R.S.; Tabora, M.; Kazemzadeh, S.; Pournaki, L.K.; Cedillos, R.; Marcia, J.; Aryana, K. Probiotic characteristics of Streptococcus thermophilus and Lactobacillus bulgaricus as influenced by new food sources. Microorganisms 2023, 11, 2291. [Google Scholar] [CrossRef]
- Zhao, X.; Ge, Y.; Yu, X.; Liu, C.; Li, H.; Wang, X.; Yao, S. Fermentation characteristics of fermented milk with Streptococcus thermophilus CICC 6063 and Lactobacillus helveticus CICC 6064 and volatile compound dynamic profiles during fermentation and storage. Molecules 2024, 29, 1257. [Google Scholar] [CrossRef]
- Vandera, E.; Kakouri, A.; Koukkou, A.-I.; Samelis, J. Major ecological shifts within the dominant nonstarter lactic acid bacteria in mature Greek Graviera cheese as affected by the starter culture type. Int. J. Food Microbiol. 2019, 290, 15–26. [Google Scholar] [CrossRef]
- Sbehat, M.; Mauriello, G.; Altamimi, M. Microencapsulation of probiotics for food functionalization: An update on literature reviews. Microorganisms 2022, 10, 1948. [Google Scholar] [CrossRef]
- Yao, M.; Xie, J.; Du, H.; McClements, D.J.; Xiao, H.; Li, L. Progress in microencapsulation of probiotics: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 857–874. [Google Scholar] [CrossRef] [PubMed]
- Zaeim, D.; Sarabi-Jamab, M.; Ghorani, B.; Kadkhodaee, R.; Tromp, R.H. Electrospray assisted fabrication of hydrogel microcapsules by single- and double-stage procedures for encapsulation of probiotics. Food Bioprocess. Technol. 2017, 102, 250–259. [Google Scholar] [CrossRef]
- Patarroyo, J.L.; Fonseca, E.; Cifuentes, J.; Salcedo, F.; Cruz, J.C.; Reyes, L.H. Gelatin-graphene oxide nanocomposite hydrogels for Kluyveromyces lactis encapsulation: Potential applications in probiotics and bioreactor packings. Biomolecules 2021, 11, 922. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Tian, H.; Yin, X.; Zhou, Y.; Zhu, L. Microencapsulation of 2-phenyl ethanol with methylcellulose/alginate/methylcellulose as the wall material and stability of the microcapsules. Polym. Bull. 2020, 77, 989–1001. [Google Scholar] [CrossRef]
- Al-Hamayda, A.; Abu-Jdayil, B.; Ayyash, M.; Tannous, J. Advances in microencapsulation techniques using Arabic gum: A comprehensive review. Ind. Crops Prod. 2023, 205, 117556. [Google Scholar] [CrossRef]
- Qi, F.; Zhu, J.; Li, M.; Ren, J.; Hu, Y.; Sun, Q. Preparation of W/O/W Lactiplantibacillus plantarum L3 microcapsules using modified low methoxy pectin as wall materials and their application in simulated yoghurt fermentation systems. Int. J. Dairy Technol. 2024, 77, 114–131. [Google Scholar] [CrossRef]
- Díaz Vergara, L.I.; Arata Badano, J.; Aminahuel, C.A.; Vanden Braber, N.L.; Rossi, Y.E.; Pereyra, C.M.; Cavaglieri, L.R.; Montenegro, M.A. Chitosan-glucose derivative as effective wall material for probiotic yeasts microencapsulation. Int. J. Biol. Macromol. 2023, 253, 127167. [Google Scholar] [CrossRef]
- Fauzi, M.A.R.D.; Pudjiastuti, P.; Wibowo, A.C.; Hendradi, E. Preparation, properties and potential of carrageenan-based hard capsules for replacing gelatine: A review. Polymers 2021, 13, 2666. [Google Scholar] [CrossRef]
- Shu, G.; He, Y.; Chen, L.; Song, Y.; Meng, J.; Chen, H. Microencapsulation of Lactobacillus acidophilus by xanthan-chitosan and its stability in yoghurt. Polymers 2017, 9, 733. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Li, X.; Michelon, M.; Leopercio, B.C.; Carvalho, M.S.; Frostad, J.M. Effects of aging on the shelf life and viscoelasticity of gellan gum microcapsules. Food Hydrocoll. 2021, 121, 106982. [Google Scholar] [CrossRef]
- Zhao, D.; Li, Z.; Xia, J.; Kang, Y.; Sun, P.; Xiao, Z.; Niu, Y. Research progress of starch as microencapsulated wall material. Carbohydr. Polym. 2023, 318, 121118. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, P.; Hu, Z.; Chen, Y.; Jin, X.; Deng, R.; Kirk, T.V.; Chen, X.D. Curcumin-loaded microcapsules with soy and whey protein as wall material: In vitro release, and ex vivo absorption based on the rat small intestine. J. Food Eng. 2024, 383, 112254. [Google Scholar] [CrossRef]
- Ye, S.; Fan, Y.; Cui, Y.; Chen, D.; Kong, L.; Liu, Y. Preparation of safflower oil microcapsules with gelatin and a novel exopolysaccharide as wall matrix. J. Food Process Eng. 2017, 40, e12573. [Google Scholar] [CrossRef]
- Boldoo, T.; Chinnasamy, V.; You, N.; Cho, H. Experimental analysis on thermal energy storage performance of micro-encapsulated stearic acid and stearyl alcohol PCM slurries; A comparative study. J. Energy Storage 2023, 73, 109218. [Google Scholar] [CrossRef]
- Yang, S.; Hu, X.; Cang, W.; Ji, S.; Wu, R.; Wu, J. Biofilm-based probiotic delivery system and its application in the food industry. Food Biosci. 2024, 62, 105172. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, J.; Zhang, Y.; Zhao, D.; Li, S.; Dou, H.; Wang, H.; Xia, X. The role of rcpA gene in regulating biofilm formation and virulence in Vibrio parahaemolyticus. Int. J. Food Microbiol. 2024, 418, 110714. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory effects triggered by lactic acid bacteria exopolysaccharides: New insights into molecular interactions with host cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef]
- Olson, J.K.; Navarro, J.B.; Allen, J.M.; McCulloh, C.J.; Mashburn-Warren, L.; Wang, Y.; Varaljay, V.A.; Bailey, M.T.; Goodman, S.D.; Besner, G.E. An enhanced Lactobacillus reuteri biofilm formulation that increases protection against experimental necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G408–G419. [Google Scholar] [CrossRef]
- Crook, N.; Ferreiro, A.; Gasparrini, A.J.; Pesesky, M.W.; Gibson, M.K.; Wang, B.; Sun, X.; Condiotte, Z.; Dobrowolski, S.; Peterson, D.; et al. Adaptive strategies of the candidate probiotic E. coli nissle in the mammalian gut. Cell Host Microbe 2019, 25, 499–512.e498. [Google Scholar] [CrossRef]
- Ma, J.; Xu, C.; Liu, F.; Hou, J.; Shao, H.; Yu, W. Stress adaptation and cross-protection of Lactobacillus plantarum KLDS 1.0628. CyTA-J. Food 2021, 19, 72–80. [Google Scholar] [CrossRef]
- Liu, K.; Fang, H.; Cui, F.; Nyabako, B.A.; Tao, T.; Zan, X.; Chen, H.; Sun, W. ARTP mutation and adaptive laboratory evolution improve probiotic performance of Bacillus coagulans. Appl. Microbiol. Biotechnol. 2020, 104, 6363–6373. [Google Scholar] [CrossRef] [PubMed]
- Bisson, G.; Marino, M.; Poletti, D.; Innocente, N.; Maifreni, M. Turbidimetric definition of growth limits in probiotic Lactobacillus strains from the perspective of an adaptation strategy. J. Dairy Sci. 2021, 104, 12236–12248. [Google Scholar] [CrossRef]
- Yadav, R.; Kumar, V.; Baweja, M.; Shukla, P. Gene editing and genetic engineering approaches for advanced probiotics: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1735–1746. [Google Scholar] [CrossRef]
- Carter, C.S.; Morgan, D.; Verma, A.; Lobaton, G.; Aquino, V.; Sumners, E.; Raizada, M.; Li, Q.; Buford, T.W. Therapeutic delivery of ang(1–7) via genetically modified probiotic: A dosing study. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 75, 1299–1303. [Google Scholar] [CrossRef]
- Fang, T.; Liu, S. Metal-Phenolic Network Directed Coating of Single Probiotic Cell Followed by Photoinitiated Thiol-Ene Click Fortification to Enhance Oral Therapy. Small 2024, 20, 2308146. [Google Scholar] [CrossRef]
- Abou Ayana, I.A.A.; Elgarhy, M.R.; Al-Otibi, F.O.; Omar, M.M.; El-Abbassy, M.Z.; Khalifa, S.A.; Helmy, Y.A.; Saber, W.I.A. Artificial intelligence-powered optimization and milk permeate upcycling for innovative sesame milk with enhanced probiotic viability and sensory appeal. ACS Omega 2024, 9, 25189–25202. [Google Scholar] [CrossRef]
- Celiberto, L.S.; Pinto, R.A.; Rossi, E.A.; Vallance, B.A.; Cavallini, D.C.U. Isolation and characterization of potentially probiotic bacterial strains from mice: Proof of concept for personalized probiotics. Nutrients 2018, 10, 1684. [Google Scholar] [CrossRef]
- Rudyk, M.; Hurmach, Y.; Serhiichuk, T.; Akulenko, I.; Skivka, L.; Berehova, T.; Ostapchenko, L. Multi-probiotic consumption sex-dependently interferes with MSG-induced obesity and concomitant phagocyte pro-inflammatory polarization in rats: Food for thought about personalized nutrition. Heliyon 2023, 9, e13381. [Google Scholar] [CrossRef]
- Boyle, R.J.; Robins-Browne, R.M.; Tang, M.L.K. Probiotic use in clinical practice: What are the risks? Am. J. Clin. Nutr. 2006, 83, 1256–1264. [Google Scholar] [CrossRef]
- Tang, Q.; Hao, Y.; Wang, L.; Lu, C.; Lu, Z. Characterization of a bacterial strain Lactobacillus paracasei LP10266 recovered from an endocarditis patient in Shandong, China. BMC Microbiol. 2021, 21, 183. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.S.; Chen, C.W.; Kuo, Y.W.; Ho, H.H. Lactobacillus spp. reduces ethanol-induced liver oxidative stress and inflammation in a mouse model of alcoholic steatohepatitis. Exp. Ther. Med. 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Antibiotic resistome of Salmonella typhi: Molecular determinants for the emergence of drug resistance. Front. Med. 2021, 15, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.R.; Achkar, J.P.; Brinich, M.A.; Farrell, R.M. Helping patients make informed choices about probiotics: A need for research. Am. J. Gastroenterol. 2009, 104, 809–813. [Google Scholar] [CrossRef]
- Besselink, M.G.H.; Santvoort, H.C.V.; Buskens, E.; Boermeester, M.A.; Gooszen, H.G. Probiotic prophylaxis in predicted severe acute pancreatitis: A randomised, double-blind, placebo-controlled trial. Lancet 2008, 371, 651–659. [Google Scholar] [CrossRef]
- Botes, M.; Reenen, C.A.V.; Dicks, L.M.T. Evaluation of Enterococcus mundtii ST4SA and Lactobacillus plantarum 423 as probiotics by using a gastro-intestinal model with infant milk formulations as substrate. Int. J. Food Microbiol. 2008, 128, 362–370. [Google Scholar] [CrossRef]
- Moreno, M.R.F.; Sarantinopoulos, P.; Tsakalidou, E.; Vuyst, L.D. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef]
- Paudel, R.; Nepal, H.P. Linezolid resistance in vancomycin resistant Enterococci: A worrisome situation. Int. J. Basic. Clin. Pharmacol. 2021, 10, 464–465. [Google Scholar] [CrossRef]
- Nicas, T.I.; Wu, C.Y.; Hobbs, J.N.; Preston, D.A.; Allen, N.E. Characterization of vancomycin resistance in Enterococcus faecium and Enterococcus faecalis. Antimicrob. Agents Chemother. 1989, 33, 1121–1124. [Google Scholar] [CrossRef]
- Graham, K.; Stack, H.; Rea, R. Safety, beneficial and technological properties of enterococci for use in functional food applications—A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef]
- Marco, M.L.; Sanders, M.E.; Ganzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The international scientific association for probiotics and prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- Lim, M.Y.; Cho, Y.; Rho, M. Diverse distribution of resistomes in the human and environmental microbiomes. Curr. Genomics 2018, 19, 701–711. [Google Scholar] [CrossRef]



| Category | Encapsulation Materials | Advantages | Disadvantages | Reference |
|---|---|---|---|---|
| Carbohydrates | Sodium alginate | Mild reaction, forms a gel with Ca2⁺, good film-forming properties, good gelling properties, good thickening properties, and high biocompatibility | Poor porosity and water solubility, sensitive to acidic environments | [135] |
| Gum arabic | High water solubility, viscosity increases significantly with concentration, good oxygen barrier properties | Low mechanical strength, needs to be combined with other materials for reinforcement and protection | [136] | |
| Pectin | Excellent gelling properties, resistant to digestive enzymes in the gastrointestinal tract, and controls the release of probiotics | Weak gel strength, prone to clumping during storage | [137] | |
| Chitosan | High biocompatibility, degradability, bioadhesiveness, and antibacterial properties | Dissolves under acidic conditions, with limited gastric protection; unable to resist external mutation pressure | [138] | |
| Carrageenan | High thermal stability and good film-forming properties | High concentrations may inhibit the growth of probiotics | [139] | |
| Xanthan gum | Wide pH tolerance, often used in combination with other gel materials | High cost, limiting industrial applications | [140] | |
| Gellan gum | Acid and heat resistant, highly stable, and capable of forming high-strength gels at low concentrations | Poor stability in physiological environments, requiring chemical modification for optimization | [141] | |
| Porous starch | Good biocompatibility, high adsorption capacity, low cost | Poor water resistance and mechanical properties, poor processability | [142] | |
| Proteins | Whey protein | High nutritional value, good emulsification, strong solubility | High temperature variability, limiting hot working applications | [143] |
| Gelatin | Biodegradability, excellent film-forming properties, strong oxygen resistance | High purification costs, low concentration gelatin chemical properties unstable | [144] | |
| Lipids | Fatty acids | Biocompatibility, low-temperature processing adaptability | Weak mechanical strength, poor water dispersibility | [145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Zhou, Y.; Xu, H. Unlocking Wellness: Probiotics as Key Drivers in Functional Food Innovation and Health Promotion. Appl. Sci. 2025, 15, 6498. https://doi.org/10.3390/app15126498
Huang Y, Zhou Y, Xu H. Unlocking Wellness: Probiotics as Key Drivers in Functional Food Innovation and Health Promotion. Applied Sciences. 2025; 15(12):6498. https://doi.org/10.3390/app15126498
Chicago/Turabian StyleHuang, Yifei, Yuankun Zhou, and Hengyi Xu. 2025. "Unlocking Wellness: Probiotics as Key Drivers in Functional Food Innovation and Health Promotion" Applied Sciences 15, no. 12: 6498. https://doi.org/10.3390/app15126498
APA StyleHuang, Y., Zhou, Y., & Xu, H. (2025). Unlocking Wellness: Probiotics as Key Drivers in Functional Food Innovation and Health Promotion. Applied Sciences, 15(12), 6498. https://doi.org/10.3390/app15126498