Abstract
Thanks to their various promising properties, bioactive peptides extracted from microalgae have recently attracted great attention from scientists. These compounds have been mostly obtained by enzymatic hydrolysis. In the present study, proteins from the marine microalga Rhodomonas sp. were hydrolysated into peptides using alcalase and pepsin. After obtaining the peptides, we characterized them and the crude protein by Fourier-transform infrared spectroscopy. Then, the biological activities of all protein preparations were evaluated. Antioxidant activity was investigated using DPPH (2,2-Diphenyl-1-picrylhydrazyl) radical-scavenging and ferric reducing power assays. The obtained results showed that crude protein presents higher antioxidant activity (74% at 1 mg/mL) compared to peptides obtained after hydrolysis by pepsin (56.5% at 1 mg/mL) and alcalase (47.5% at 1 mg/mL). The anti-inflammatory activity was also evaluated by inhibition of the denaturation of albumin assay. An interesting anti-inflammatory activity was obtained using protein extract. It was improved after enzymatic hydrolysis using pepsin to reach 89% of inhibition at 500 µg/mL. The obtained data showed that the marine microalga Rhodomonas sp. could be a potential source of valuable proteins and peptides for the pharmaceutical and cosmetic fields.
1. Introduction
The overloading of nutrients in aquatic ecosystems provides ideal conditions for the excessive growth of marine organisms, which comprise approximately half of the total global biodiversity [1]. In parallel, oxidative stress and reactive oxygen species (ROS) have been increased in these kinds of environments, which could cause severe cellular damage [2]. Therefore, in order to defend against oxidative bursts, microalgae and other oceanic organisms have developed multiple defensive mechanisms, mainly the accumulation of valuable biomolecules, which could stimulate antioxidant activity [3]. Thus, the interest in extracting new antioxidant agents from aquatic species has remarkably increased. Aquatic ecosystems include a great diversity of marine species, 245,500 of which have been identified thanks to advances in molecular biology. All of these organisms constitute blue biotechnology and are classified into different major categories, namely, vertebrates, invertebrates, bacteria, fungi, and algae. Algae constitute the most abundant resource in the aquatic environment. They are a very diverse group, approximately 50,000 species of which have been isolated and identified [4]. They are chlorophyllous organisms, classified into macroalgae and microalgae. Macroalgae represent a polyphyletic group of multicellular algae inhabiting the shallow littoral zone with sufficient light to accomplish photosynthesis [5]. Microalgae can be prokaryotic or eukaryotic. Prokaryotic microalgae are represented by cyanobacteria, which possess a wide variety of ecological characteristics that have allowed them to colonize most aquatic ecosystems, while eukaryotic microalgae are classified into Chlorophyceae, Rhodophyceae, and Phaeophyceae. Their chloroplasts contain several flattened thylakoids that contain chlorophylls and other pigments responsible for their pigmentation. Microalgae cultivation is an effective process used for CO2 sequestration and carbon footprint reduction [6]. Thus, microalgae biomass production is a promising strategy to combine the reduction of anthropogenic emissions with the production of biomass useful in various fields [7,8,9]. Microalgae biomass has been considered as a new source of metabolites [3,10,11,12]. They are highly explored due to their richness in valuable molecules, mainly proteins, polysaccharides, fatty acids, and pigments. In particular, many microalgae strains, such as Arthrospira sp., Aphanizomenon sp., Dunaliella sp. and Chlorella sp., are characterized by their high protein content, exceeding 50%, and are actually used as human food supplements [8]. In fact, they have protein and amino acid profiles with high dietary value.
Anti-inflammatory activity has been also reported for multiple compounds derived from different microalgae species, such as those belonging to Chlorella, Dunaliella, and Phaeodactylum genera [13,14]. Such products could prevent various occurrences caused by inflammations, such as the increase in protein denaturation, increase in vascular permeability, and biological membrane alteration [15]. In this frame, the isolation of bioactive peptides is a novel approach attracting scientists’ attention. Nowadays, several therapeutic peptides, obtained by the proteolytic degradation of some proteins present in bacteria, plants, and animals, have been reported as potential therapeutic agents [16]. Those peptides are characterized by specific biological activities coupled with low toxicity.
Antioxidant and anti-inflammatory capacities are among the most popular biological activities of peptides [17,18]. Both animal and plant proteins are known to contain potential bioactive sequences; however, most studies to date have involved milk proteins [19]. The most common and easiest method by which produce bioactive peptides is enzymatic hydrolysis [16]. It generates peptides with specific molecular weights and a specific amino acid composition. In addition, it allows the control of the hydrolysis process as well as the hydrolysis degree. Enzymatic hydrolysis could create particular structural motifs in the generated peptides, conferring unique structural properties that influence their specific mode of action [20]. Bioactive peptides are generally 3 to 20 amino acid residues in length and have molecular weights of less than 6000 Da [21].
Microalgae, especially those produced at an industrial scale, are considered a promising bioactive peptides source with highly beneficial features. In this frame, peptides with antioxidant potential have been extracted from various Chlorophycae species, mainly Chlorella vulgaris, Navicula incerta, Chlorella ellipsoidea, and Dunaliella salina [22,23,24,25,26]. Anti-inflammatory and antihypertensive peptides have been also isolated from cyanobacteria species belonging to the Spirulina genus [18].
In general, the average protein content of the microalgae biomass is around 25–30% of the total biomass. The protein content and amino acid composition of microalgae biomass are affected by many biotic and abiotic factors and by the genomic variation of species [12,27]. However, the biological activities of proteins can be determined by their amino acid composition, their size, as well as their 3D conformation. It is important to note that proteins and peptides from cryptophytes are not as well studied and exploited as those from other phyla. Rhodomonas genus is a cryptophyte microalga that is still poorly studied in terms of its biodiversity, distribution, and food web interactions despite its widespread occurrence in coastal ecosystems. Indeed, it is widely used in aquaculture feed due to its high nutritional value. In fact, Rhodomonas species are able to produce a large variety of metabolites, including proteins, carbohydrates, carotenoids, chlorophylls a and c, phycoerythrin, as well as fatty acids [28]. Regarding its biochemical composition, The Rhodomonas genus is considered a suitable microalga for a variety of applications in biomedical, biotechnological, and nutraceutical domains due to its high production of commercially valuable compounds such as amino acids, polar lipids, and unsaturated eicosapentaenoic and docosahexaenoic fatty acids [29]. In addition, it represents a source of natural proteins that could compete favorably with the conventional protein sources. In addition, the Rhodomonas genus constitutes a promising candidate for the production of phycoerythrin, an interesting fluorescent protein, used as a natural dye in food and cosmetics as well as a fluorescent marker in biomedical research. This phycobiliprotein is a potential therapeutic agent that has been shown to have multiple benefits in human health thanks to its antioxidant, anticancer, and anti-inflammatory properties [30]. This adds value for the Rhodomonas genus to be considered a potential strain for several promising bioactivities. These potentialities of Rhodomonas proteins could be improved after the hydrolysis and generation of peptides. Hence, it will be interesting to isolate peptides from Rhodomonas species and then study their biological activities, something that has not been previously investigated.
In the present study, proteins and peptides hydrolysates were prepared from a strain of the Rhodomonas genus [30] and were then further utilized to investigate their antioxidant and anti-inflammatory activities.
2. Materials and Methods
2.1. Microalgal Strain, Growth Conditions, and Protein Accumulation Monitoring
A local marine microalgae strain was previously isolated from the Tunisian coast Sidi Mansour and identified as belonging to the Rhodomonas genus [31]. It was maintained in culture in F/2 medium [31] at 23 ± 2 °C under continuous lighting at an intensity irradiance of 80 μmol photons/m2/s. Cultures were inoculated at an initial cell density of 104 cells/mL and were performed in triplicate. Microalgae cell concentrations were determined by cell counts every 48 h using a light microscope (Motic microscope AE 2000, Spain) and Malassez counting chamber (Supe Rior, U, 200 mm) [32]. The concentration of accumulated protein in Rhodomonas sp. cells was estimated every 48 h by Lowry assay [3] as previously reported by Derbel et al. [30], using bovine serum albumin (BSA) as standard.
2.2. Biochemical Characterization of Rhodomonas sp. Biomass
Rhodomonas sp. biomass was recovered during the stationary phase and then characterized. Total protein content in Rhodomonas sp. biomass was determined by the Kjeldahl method [33] using a 500 mg dry microalgae sample. Ash content determination was performed gravimetrically after incineration at 550 °C in an oven (SANTE, Model: STM-3-12, 220V). Carbohydrates, lipids, and chlorophyll content were determined as previously detailed by Ben Mohamed et al. [34].
2.3. Preparation of Rhodomonas sp. Proteins Extract
Crude protein from Rhodomonas sp. was obtained after the microalga cell lysis step. Eight cell lysis methods were tested:
Method I: 30 mg of alumina powder was added to 30 mg of microalgal biomass and suspended in 3 mL of distilled water. Grinding of the sample was performed using a cold mortar (PZRT 60 mm Manual Ceramics Grinder) in ice for 15 min.
Method II: 30 mg of microalgal biomass was suspended in 3 mL of distilled water and vortexed for two minutes. The sample was frozen at −20 °C and defrosted until completely thawed. This procedure was repeated five times to allow cell lysis.
Method III: 30 mg of microalgal biomass was suspended in 3 mL of distilled water and vortexed for two minutes. Sonication was then performed for 5 min, using an ultrasonic bath (Ultrasons, J.P. Selecta, Barcelona, Spain), filled with cold distilled water.
Method IV: similar to Method III, differing only by the sonication time: 10 min (cycle: 5 min sonication, 5 min rest time) instead of 5 min.
Method V: this treatment is similar to Method II, differing only by the sonication time: 15 min instead of 10 min.
Method VI: 30 mg of microalgal biomass was suspended in 3 mL of distilled water and vortexed for two minutes. The sample was then subjected to probe sonication at 20 kHz for 5 min on ice (cycle: 20 s pulse time, 20 s rest time) using a probe sonicator (Sonics, 130 W).
Method VII: similar to Method VI, differing only by the sonication time: 10 min instead of 5 min.
Method VIII: similar to Method VI, differing only by the sonication time: 15 min instead of 5 min.
After lysis steps, cells were centrifuged at 6000× g for 30 min at 4 °C and supernatants were recovered. Proteins’ concentrations in the obtained crude extracts were determined through Lowry method [3] using bovine serum albumin (BSA) as standard.
Proteins were precipitated as described by Montone et al. [35]. Briefly, ice-cold acetone was added to each recovered supernatant (2:1). The mixture was incubated at −20 °C during 24 h and then centrifuged at 6000× g for 30 min at 4 °C. The obtained pellet was collected and freeze-dried.
2.4. Cell density and Proteins Production Assessments
Cell growth and proteins’ accumulation rates of the studied strain were monitored for 22 consecutive days. The microalgal growth was determined by counting using a light microscope (Motic microscope AE 2000, Madrid, Spain) and Malassez counting chamber (Supe Rior, U, 200 mm). Proteins’ concentrations were determined through Lowry method [3] using bovine serum albumin (BSA) as standard.
2.5. Preparation of Rhodomonas sp. Peptides Hydrolysates
The proteins from Rhodomonas sp. at 30 mg/mL were hydrolyzed separately by alcalase and pepsin. The hydrolysis of proteins by alcalase (Sigma-Aldrich) was performed using an enzyme:proteins ratio (w/w) of 1:70, at 50 °C, pH 9, for 4 h. After hydrolysis, the reaction was stopped by decreasing the pH of the reactional mixture to 2 with HCl (0.5 M). The obtained peptides were cooled at room temperature, then centrifuged at 6000× g for 30 min. The supernatant was recovered and its pH was adjusted to 7 using NaOH (0.5 M) before being freeze-dried. For hydrolysis of proteins with pepsin (Biobasic), an enzyme:proteins ratio (w/w) of 1:25 was used. The enzymatic reaction was performed at 37 °C, pH 3, for 4 h [36]. After hydrolysis, the reaction was stopped by increasing the pH to 8 with NaOH (0.5 M), then the mixtures were centrifuged at 6000× g for 30 min. Supernatants were collected and then freeze-dried.
The degree of hydrolysis (DH) was determined as previously described by Soto-Sierra et al. [37] Equation (1).
where,
DH (%) = h/htot
htot = number of peptide bonds in the substrate per g of protein (∼8);
h = number of hydrolyzed peptide bonds per g of protein calculated as follows:
where,
h = (V ∗ N)/(mα NH2)
V = volume of titrant (NaOH (mL));
N = normality of the titrant (NaOH);
m = mass of protein (g);
α = mean degree of dissociation calculated as follows:
α = 10 (pH.pK)/(1 + 10(pH.pK))
2.6. FTIR Spectroscopy
Fourier infrared spectra were determined for Rhodomonas sp. crude protein as well as peptide hydrolysates using a VERTEX 70 FT-IR instrument equipped with an ATR A225 module (palladium–diamond). The IR spectra (50 scans) were obtained at room temperature in the spectral range 4000–400 cm−1, using OPUS 7.2 software (Bruker Optics GmbH, Billerica, MA, USA). Spectra were analyzed using the same software, and results were exported for plotting using Origin 8.5.1 software SR2 (OriginLab, Northampton, MA, USA).
2.7. Antioxidant Activity Evaluation
2.7.1. DPPH Radical-Scavenging Activity
The DPPH (2,2-diphenyl-1-picrylhydrazyl) radical-scavenging ability of the protein extract and peptide hydrolysates was estimated following the method described by Kirby and Schmidt [38]. Five hundred microliters of each sample was mixed with 375 μL of ethanol (99%) and 125 μL of DPPH (Sigma-Aldrich, D132) solution prepared in ethanol (0.02% (w/v)). Afterward, the mixtures were incubated for 30 min at room temperature in darkness. Then, A517 was measured. Five concentrations (0.1, 0.3, 0.5, 0.8, 1 mg/mL) were tested for each sample in triplicates. Ascorbic acid was used as the positive control. The DPPH (2,2-Diphenyl-1-picrylhydrazyl)-scavenging activity was calculated using the following Equation (2):
where Acontrol, Ablank, and Asample were the absorbance of the negative control (without sample), the blank (without DPPH solution), and the samples, respectively.
DPPH-radical-scavenging activity (%) = (Acontrol + Ablank − Asample)/Acontrol × 100
2.7.2. Ferric-Reducing Power Assay
The ferric-reducing capacities of Rhodomonas sp. proteins and peptides were evaluated by the method reported by Yıldırım et al. [39]. Briefly, 0.5 mL of each sample was mixed with 1.25 mL of distilled water and 1.25 mL of potassium ferricyanide (1% (m/v)). The mixture was incubated for 30 min at 50 °C and then treated with 1.25 mL of trichloroacetic acid (10% (w/v)). After centrifuging for 10 min at 10,000× g, 1.25 mL of the obtained supernatant was recovered, which was then mixed with 1.25 mL of distilled water and 0.25 mL of ferric chloride solution (0.1% (w/v)). After incubation for 10 min in darkness, A700 was measured. Five different concentrations (0.1, 0.3, 0.5, 0.8, 1 mg/mL) were tested for each sample in triplicates. Ascorbic acid (Sigma-Aldrich, Darmstadt, Germany) was used as positive control. All experiments were performed in triplicate.
2.8. In vitro Anti-Inflammatory Activity Evaluation
The ability of Rhodomonas sp. proteins and peptides to inhibit albumin denaturation was evaluated using the method described by Chakou et al. [40] with slight modification. Five different concentrations (0.1, 0.2, 0.3, 0.4, 0.5 mg/mL) were prepared for each proteins and peptides sample in Tris buffer (0.05 M, pH 6.6). Five hundred microliters of each sample was mixed with 1% (w/v) of aqueous solution of bovine albumin fraction V (Sigma-Aldrich, Saint-Louis, MI, USA). The reaction mixture was first incubated at 37 °C for 20 min, then heated at 70 °C for 5 min. After cooling at room temperature, the A600 of the mixture was determined. Diclofenac (Sodic Diclophenac) was used as the positive control. All assays were set up in triplicates.
The albumin denaturation inhibition percentage was calculated according to Equation (3)
% inhibition = 100 ∗ (Acontrol − Asample)/Acontrol
3. Results and Discussion
3.1. Biochemical Characterization of Rhodomonas sp. Biomass
The composition of Rhodomonas sp. biomass, collected at the early stationary phase, was determined. The obtained results showed that Rhodomonas sp. accumulated 34.5 mg/100 mg DW of proteins, 14.18 mg/100 mg DW of carbohydrate, 13 mg/100 mg DW of lipids, 15.73 mg/100 mg DW of ash, and 0.46 mg/100 mg DW of Chlorophyll (Table 1). These results revealed that organic constituents were the most abundant components with a proportion greater than 75 mg/100 mg DW [41]. As previously reported, proteins were the major organic components. In fact, several studies have shown the richness of Rhodomonas species in proteins. R. lens and R. salina species were shown to contain a large amount of proteins, ranging from 30% to 60% [42,43,44,45]. The carbohydrate content was relatively low compared with data reported by Fernández-Reiriz et al. [46] showing a carbohydrate content of 28.9% for Rhodomonas sp. However, the obtained carbohydrate content was comparable to that reported for R. lens, i.e., 11% [44]. Furthermore, the majority of microalgae, including Rhodomonas genus, accumulate lipids, in particular, triacylglycerols. Under conditions of macronutrient deficiency, such as nitrogen starvation, the cell stores the carbon flux obtained from photosynthesis as lipids [47]. In addition, a high ash level was found in Rhodomonas sp. microalgae biomasses [46]. It is worth noting that microalgae biomass composition is extremely dependent on the culture media used as well as environmental factors [48,49].

Table 1.
Biochemical composition (% of dry weight) of Rhodomonas sp. biomass.
3.2. Cell Density and Protein Production Assessments
In order to determine the culture phase coinciding with maximum protein production using Rhodomonas sp. strain, cell growth and protein production were monitored (Figure 1). The obtained data showed a classical sigmoidal growth curve. A lag phase occurred during the first 72 h. Then, a typically exponential phase was reached and lasted 9 days, followed by the stationary phase between the 11th and 22nd days.
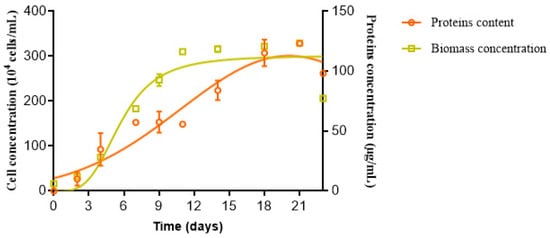
Figure 1.
Assessment of Rhodomonas sp. cell density and protein production in accordance with growth phases.
In parallel to cell number monitoring, protein production was taken into consideration. It was observed that the protein production was in parallel to the biomass production, reaching its maximum after 21 days of culture. Several studies reported that protein production was correlated with nutrient availability. In fact, the maximum protein accumulation was achieved during the logarithmic phase, and it then decreased according to nutrient concentrations [50]. However, Sui et al., [51] proved that the highest essential amino acid values were obtained at the stationary phase for the Dunaliella salina species.
3.3. Total Intracellular Protein Extraction Using Different Lysis Methods
Different cell lysis methods were compared to find the most efficient protocol suitable for intracellular protein recovery from Rhodomonas sp. The results revealed that biomass grinding into alumina powder seems to be the most efficient method for Rhodomonas sp. cell lysis compared to physical disruption by sonication and freeze-thawing (Figure 2). In concordance, Meijer and Wijffels [52] found that sonic disruption was not efficient for protein extraction from Rhodomonas species, with an efficiency of 4%, but was still reliable for Chlorella species, with an efficiency of 103%. Various methods have been employed for microalgae cellular membrane disintegration or disruption, including mechanical, physical, or nonmechanical techniques [53]. The success of the lysis method depends on microalgae cell wall composition [53,54].

Figure 2.
Comparison of different cell lysis methods for the recovery of intracellular proteins from Rhodomonas sp.
3.4. Peptides Hydrolysed by Alcalase and Pepsin
Enzymatic hydrolysis is a technique for specifically fragmenting a raw material in order to extract peptides of interest in the presence of proteases. During enzymatic hydrolysis, proteases will break the peptide bonds between two adjacent amino acids in the primary sequence of a protein, thus generating at least two peptides. The enzymatic hydrolysis of a protein extract makes it possible to obtain peptide fractions potentially endowed with biological activities. Alcalase and pepsin were used separately to release peptides from Rhodomonas sp. protein extract. The use of specific or even nonspecific proteases is the most common way to produce bioactive peptides, as it takes less time to reach the degree of hydrolysis, and it is possible to control the hydrolysis process in order to achieve peptides with specific molecular weights and a specific amino acid composition [55]. Alcalase is considered the most effective enzyme to produce a high percentage of protein recovery [55]. In addition, it has been reported that pepsin hydrolysate exerts stronger biological activities, which may be due to the random cleavages of hydrophobic amino acids by pepsin [56].
The peptides obtained after enzymatic hydrolysis revealed a DH of 34.58% and 43.24% using alcalase and pepsin, respectively. These results indicated that Rhodomonas sp. protein extract was suitable as substrate for both used proteases.
3.5. Proteins and Peptides Characterization
Fourier-transform infrared (FTIR) spectroscopy is a method widely used for the characterization of proteins and peptides [57]. It has been reported that proteins and peptides generate nine distinct infrared absorption bands (i.e., amide A, amide B, and amide I–VII) [58,59]. The most predominant bands are recorded at approximately 1650 cm−1 and at 1550 cm−1, corresponding to the amide I and amide II bands.
Figure 3 represents FTIR spectra of proteins and peptides samples. The characteristic bands of amide I, amide II, and amide III, as well as the C–C and C=O stretching vibration are detectable in the obtained FTIR spectra.
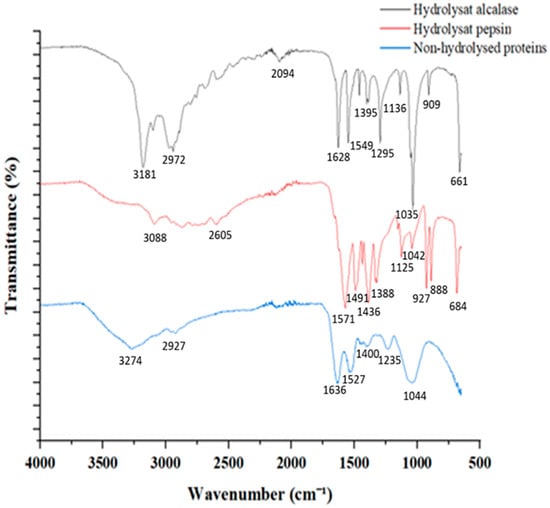
Figure 3.
FTIR spectra of Rhodomonas sp. crude protein and peptides produced after alcalase and pepsin hydrolysis.
The main peaks associated with crude protein preparation were observed at 3274 cm−1, 2927 cm−1, 1636 cm−1, 1527 cm−1, and 1044 cm−1, corresponding to N–H stretch, O–H stretch, C=O stretch at amide I; N–H deformation/C–N stretch at amide II, and C–O stretch, respectively (Table 2). After the digestion of proteases, the spectra of the produced peptides revealed the appearance and shift of several peaks compared to the nonhydrolyzed control (Figure 3). In addition, peaks related to the amide bands, which does not change location, showed a decreasing amplitude in the peptides’ spectra. The shifted and/or appeared peaks in the peptides spectra were recorded in various regions, mainly at 1500–1400 cm−1, associated with C-terminal groups. Protein digestion using proteases generates COO− groups.
Table 2.
Summary of wave numbers and corresponding functional groups.
Recently, FTIR spectroscopy has been considered an efficient approach for the structural characterization of proteins and peptides [57]. It could provide information on peptides and proteins size as well as on the secondary structure of protein [60]. It is also a useful technique for the investigation of protein-related parameters, mainly the effects of solvent, temperature, and pH on proteins [62,63]. Kristoffersen et al. [57] proved the potential of using FTIR for monitoring protein sizes during enzymatic digestion.
3.6. Antioxidant Activity
Oxidative bursts can be avoided by the action of antioxidants through different mechanisms, including the formation of chelate complexes with pro-oxidant metals, free radical scavenging, quenching of singlet oxygen and photosensitizers, and the deactivation of peroxydinitrite and lipoxygenases [64]. Synthetic antioxidant drugs available on the market have certain side effects, which are not conducive to long-term use. Hence, interests are gradually focusing on natural antioxidants, which are safer than chemically synthesized antioxidants, and some of these antioxidants may be more efficient than synthetic drugs. Therefore, the present study aims to investigate the antioxidant potential of proteins and peptides derived from the cryptophyte microalga Rhodomonas sp. through DPPH radical-scavenging and ferric reducing power assays.
DPPH radical-scavenging activities were found to increase proportionately to the concentration in all tested samples (Figure 4a). The crude protein used at 1 mg/mL showed an interesting antioxidant activity against DPPH, reaching 74%. This antioxidant activity was much higher than that previously reported for proteins extracted from Schizochytrium limacinum and Porphyra columbina [65,66]. In concordance, the crude protein showed dose-dependent reducing capacities, with a maximum reached at 1 mg/mL concentration (Figure 4b). The important antioxidant activity obtained for crude protein may be due to the presence of amino acids involved in free radical deactivation. Antioxidant peptide molecules can achieve quenching of reactive oxygen/nitrogen species through two pathways, namely, hydrogen atom transfer (HAT) and single electron transfer (SET) reactions [67]. Particularly, Met has been considered an important free radical scavenger in biological systems [68]. This is due to the Met radical, which is very labile to oxidation and may potentially scavenge free radicals before they attack other amino acid residues critical to protein structure or function. To obtain additional structural information on the antioxidant peptides derived from Rhodomonas sp. protein hydrolysate, the active peptides need to be purified and their amino acid sequences should be investigated.

Figure 4.
Antioxidant activities of proteins and peptide hydrolysates using (a) DPPH radical-scavenging assay and ferric reducing power assay (b).
The important antioxidant activity obtained for crude protein indicates that Rhodomonas sp. could be a good candidate as a natural antioxidant source for use in the food and cosmetic fields.
Moderate antioxidant activities were found for peptide hydrolysates obtained by enzymatic digestion using alcalase and pepsin, separately. This finding confirms the suggestion of Alzahrani et al. [69] that enzymatic hydrolysis could not increase antioxidant activity if the specific peptide having antioxidant activity was not liberated or the active peptide was inactivated by digestion. They suggested also that the inactivation of peptides could be induced by the presence of exoproteases in the commercial protease preparations, through the splitting of some amino acids from the active N-terminal or C-terminal of the peptides. In contrast, studies performed on microalgae Porphyra columbina [66] and Palmaria palmeadas [70] found that antioxidant activities were enhanced in peptides. Hence, the impact of protein hydrolysis on antioxidant activity seems to be dependent on the microalgal species, the degree of hydrolysis, and the type of enzyme used [71]. In this frame, several studies have reported antioxidant activity enhancement after enzymatic hydrolysis. In fact, the DPPH radical-scavenging capacity as well as ferric reducing power, obtained using proteins extracted from Schizochytrium limacinum, were shown to improve significantly after hydrolysis using pepsin, trypsin, or α-chymotrypsin digestion [65]. Cian et al. [66] found that the antiradical activity of protein extract of Porphyra columbina was also enhanced after enzymatic hydrolysis, from 35% to 54% at 4 mg/mL proteins concentration.
3.7. Anti-Inflammatory Activity
Inflammation is a complex biological process that involves many signaling biomolecules at the intracellular level as well as coordinated communication between different immune cells. It appears when healthy tissues are exposed to biological, physical, or chemical stimuli. Protein denaturation is a well-documented cause of inflammation; thus, it is considered as a marker for inflammatory disease [72]. The stimulation and activation of immune cells, such as macrophages and neutrophils, induce the generation of proinflammatory cytokines such as tumor necrosis factor (TNF-α), interleukin 1β (IL-1β), and mitogen-activated protein kinase (MAPK). TNF-α can be involved in several crucial events for the initiation of inflammation [72]. Interleukin 1β may stimulate signal transduction by activation of the nuclear factor-κB (NF-κB) and the MAPK. NF-κB, when activated, may, in turn, induce further expression of multiple cytokines, including TNF-α and IL-1β, and promote the inflammatory process.
Microalgal anti-inflammatory compounds may act through one or more mechanisms involved in inflammation, including the release of chemicals such as leukotrienes, prostaglandins, bradykinin, histamine, platelet-activating factor, and interleukin-1 from tissues and migrating cells [14].
The denaturation of proteins corresponds to proteins losing their secondary, tertiary, and quaternary structures, resulting in loss of biological function [73]. This process could be induced by the application of external stress, such as heat, or by exposure to chemicals, such as organic solvents [15]. Hence, to investigate the anti-inflammatory activity, the ability of protein extract and peptide hydrolysates to inhibit the heat-induced denaturation of albumin was evaluated.
As seen in Figure 5, crude protein exhibited an inhibiting effect of about 67% at 0.5 mg/mL. The anti-inflammatory activity was found to be improved after enzymatic hydrolysis by the acidic protease: pepsin. In fact, at 0.5 mg/mL concentration, pepsin hydrolysate revealed a much higher capacity to inhibit albumin denaturation (89%). The in vitro anti-inflammatory activity of pepsin hydrolysate was similar to that of the standard drug diclofenac used at the same concentration. In addition, the anti-inflammatory activity of pepsin hydrolysate was much higher than that of the aqueous extracts of the medicinal plants Cannabis sativa (<70%) and Ficus racemose (<80%) [74].
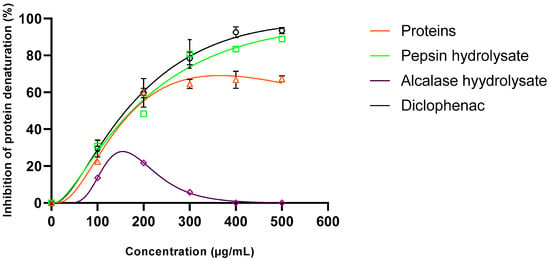
Figure 5.
Inhibition of albumin denaturation by crude protein extract and peptides at different concentrations.
In contrast, the anti-inflammatory capacity was lost after hydrolysis by alcalase. Changing the protease modifies the resulting peptide’s length, sequence, and consequently, biological activities.
4. Conclusions
Crude protein and peptides from Rhodomonas sp. were prepared and tested for their antioxidant and anti-inflammatory activities. Crude protein may potentially serve as a natural antioxidant source in the cosmetic and pharmaceutical fields. This antioxidant potential was not increased after enzymatic hydrolysis by alcalase and pepsin. Concerning the anti-inflammatory activity, it was found to be improved by enzymatic hydrolysis using pepsin. Despite these encouraging results, further in vivo tests are required.
Author Contributions
Conceptualization, J.E., I.F., P.M. and S.A.; methodology, H.D., W.M. and J.E.; formal analysis, H.D. and J.E. supervision, J.E., I.F., P.M. and S.A.; investigation, writing—original draft, H.D. and J.E.; Writing—review and editing, J.E., I.F., P.M. and S.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This work was supported by grants from the Tunisian Ministry of Higher Education and Scientific Research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kar, B.; Kumar, R.S.; Karmakar, I.; Dola, N.; Bala, A.; Mazumder, U.K.; Hadar, P.K. Antioxidant and in vitro anti-inflammatory activities of Mimusops elengi leaves. Asian Pac. J. Trop. Biomed. 2012, 2, 976–980. [Google Scholar] [CrossRef]
- Tierney, M.S.; Croft, A.K.; Hayes, M. A review of antihypertensive and antioxidant activities in macroalgae. Bot. Mar. 2010, 53, 387–408. [Google Scholar] [CrossRef]
- Gargouch, N.; Karkouch, I.; Elleuch, J.; Elkahoua, S.; Michaud, P.; Abdelkafi, S.; Laroche, C.; Fendri, I. Enhanced B-phycoerythrin production by the red microalga Porphyridium marinum: A powerful agent in industrial applications. Int. J. Biol. Macromol. 2018, 120, 2106–2114. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Nelson, D.R.; Mystikou, A.; Daakour, S.; Salehi-Ashtiani, K. Advances in microalgal research and engineering development. Curr. Opin. Biotechnol. 2019, 59, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Miladi, R.; Manghisi, A.; Armeli Mincante, S.; Genovese, G.; Abdelkafi, S.; Morabito, M. A DNA barcoding survey of Ulva (Chlorophyta) in Tunisia and Italy reveals the presence of the overlooked alien Ulva ohnoi. Cryptogam. Algol. 2018, 39, 85–107. [Google Scholar] [CrossRef]
- Uma, V.S.; Usmani, Z.; Sharma, M.; Diwan, D.; Sharma, M.; Guo, M.; Tuohy, M.G.; Makatsoris, C.; Zhao, X.; Thakur, V.K.; et al. Valorisation of algal biomass to value-added metabolites: Emerging trends and opportunities. Phytochem. Rev. 2022. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Hadjkacem, F.; Hentati, F.; Drira, R.; Pierre, G.; Gardarin, C.; Delattre, C.; El Alaoui-Talibi, Z.; Cherkaoui, E.M.; et al. Influence of the sulfate content of the exopolysaccharides from Porphyridium sordidum on their elicitor activities on date palm vitroplants. Plant Physiol. Biochem 2022, 186, 99–106. [Google Scholar] [CrossRef]
- Elleuch, J.; Hadj Kacem, F.; Ben Amor, F.; Hadrich, B.; Michaud, P.; Fendri, I.; Abdelkafi, S. Extracellular neutral protease from Arthrospira platensis: Production, optimization and partial characterization. Int. J. Biol. Macromol. 2021, 167, 1491–1498. [Google Scholar] [CrossRef]
- Hadjkacem, F.; Pierre, G.; Christophe, G.; Elleuch, J.; Fendri, I.; Boual, Z.; Ould El Hadj, M.D.; El Alaoui-Talibi, Z.; El Modafar, C.; Dubessay, P.; et al. Bioconversion of the brown Tunisian seaweed Halopteris scoparia: Application to energy. Energies 2022, 15, 4342. [Google Scholar] [CrossRef]
- Elleuch, F.; Ben Hlima, H.; Barkallah, M.; Baril, P.; Abdelkafi, S.; Pichon, C.; Fendri, I. Carotenoids overproduction in Dunaliella sp.: Transcriptional changes and new insights through lycopene cyclase regulation. Appl. Sci. 2019, 9, 5389. [Google Scholar] [CrossRef]
- Ben Amor, F.; Barkallah, M.; Eleuch, F.; Karkouch, N.; Dammak, M.; Baréa, B.; Villeneuve, P.; Abdelkafi, S.; Fendri, I. Cyanobacteria as sources of marine bioactive compounds: Molecular specific detection based on Δ9 desaturase gene. Int. J. Biol. Macromol. 2017, 105, 1440–1445. [Google Scholar] [CrossRef]
- Tounsi, L.; Hentati, F.; Ben Hlima, H.; Barkallah, M.; Smaoui, S.; Fendri, I.; Philippe, M.; Abdelkafi, S. Microalgae as feedstock for bioactive polysaccharides. Int. J. Biol. Macromol. 2022, 221, 1238–1250. [Google Scholar] [CrossRef]
- Guzmán, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Tabarzad, M.; Atabaki, V.; Hosseinabadi, T. Anti-inflammatory activity of bioactive compounds from microalgae and cyanobacteria by focusing on the mechanisms of action. Mol. Biol. Rep. 2020, 47, 6193–6205. [Google Scholar] [CrossRef]
- Leelaprakash, G.; Dass, S.M. In vitro anti-inflammatory activity of methanol extract of Enicostemma axillare. Int. J. Drug Dev. Res. 2011, 3, 189–196. [Google Scholar]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive peptides: Synthesis, sources, applications, and proposed mechanisms of action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Guzmán, F.; Wong, G.; Román, T.; Cárdenas, C.; Alvárez, C.; Schmitt, P.; Albericio, F.; Rojas, V. Identification of antimicrobial peptides from the microalgae Tetraselmis suecica (Kylin) Butcher and bactericidal activity improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef]
- Vo, T.S.; Ryu, B.; Kim, S.K. Purification of novel anti-inflammatory peptides from enzymatic hydrolysate of the edible microalgal Spirulina maxima. J. Funct. Foods 2013, 5, 1336–1346. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food. Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef]
- Anh, T.L.Q.; Hoa, N.T.Q.; Nguyen, P.D.T.; Thanh, H.V.; Nguyen, P.B.; Anh, L.T.H.; Dao, D.T.A. Soybean Protein Extraction by Alcalase and Flavourzyme, Combining Thermal Pretreatment for Enteral Feeding Product. Catalysts 2020, 10, 829. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Arguelles, E.; Laurena, A.C.; Martinez-Goss, M.R.; Monsalud, R.G. Antibacterial activity, total phenolic content and antioxidant capacity of a green microalga Desmodesmus sp.(U-AU2) from Los Baños, Laguna (Philippines). J. Nat. Stud. 2017, 16, 1–13. [Google Scholar]
- Kang, K.H.; Qian, Z.J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.K. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in HepG2/CYP2E1 cells induced by ethanol. Phytother Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 13, 3419–3425. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Xia, E.; Zhai, L.; Huang, Z.; Liang, H.; Yang, H.; Song, G.; Li, W.; Tang, H. Optimization and identification of antioxidant peptide from underutilized Dunaliella salina protein: Extraction, in vitro gastrointestinal digestion, and fractionation. Biomed Res. Int. 2019, 2019, 6424651. [Google Scholar] [CrossRef]
- Garofalo, C.; Norici, A.; Mollo, L.; Osimani, A.; Aquilanti, L. Fermentation of microalgal biomass for innovative food production. Microorganisms 2022, 10, 2069. [Google Scholar] [CrossRef]
- Oostlander, P.C.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Optimization of Rhodomonas sp. under continuous cultivation for industrial applications in aquaculture. Algal Res. 2020, 47, 101889. [Google Scholar] [CrossRef]
- Latsos, C.; van Houcke, J.; Blommaert, L.; Verbeeke, G.P.; Kromkamp, J.; Timmermans, K.R. Effect of light quality and quantity on productivity and phycoerythrin concentration in the cryptophyte Rhodomonas sp. J. Appl. Phycol. 2021, 33, 729–741. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Pancha, I.; Jain, D.; Mishra, S. Effect of preservatives for food grade C-Phycoerythrin, isolated from marine cyanobacteria Pseudanabaena sp. Int. J. Biol. Macromol. 2010, 47, 597–602. [Google Scholar] [CrossRef]
- Derbel, H.; Elleuch, J.; Tounsi, L.; Nicolo, M.S.; Rizzo, M.G.; Michaud, P.; Fendri, I.; Abdelkafi, S. Improvement of biomass and phycoerythrin production by a strain of Rhodomonas sp. isolated from the Tunisian coast of Sidi Mansour. Biomolecules 2022, 12, 885. [Google Scholar] [CrossRef]
- Elleuch, J.; Ben Amor, F.; Chaaben, Z.; Frikha, F.; Michaud, P.; Fendri, I.; Abdelkafi, S. Zinc biosorption by Dunaliella sp. AL-1: Mechanism and effects on cell metabolism. Sci. Total Environ. 2021, 773, 145024. [Google Scholar] [CrossRef]
- Ben Hlima, H.; Bohli, T.; Kraiem, M.; Ouederni, A.; Mellouli, L.; Michaud, P.; Abdelkafi, S.; Smaoui, S. Combined effect of Spirulina platensis and Punica granatum peel extacts: Phytochemical content and antiphytophatogenic activity. App. Sci. 2019, 9, 5475. [Google Scholar] [CrossRef]
- Ben Mohamed, J.; Elleuch, J.; Drira, M.; Esteban, M.Á.; Michaud, P.; Abdelkafi, S.; Fendri, I. Characterization and Biotechnological Potential of Two Native Marine Microalgae Isolated from the Tunisian Coast. Appl. Sci. 2021, 11, 5295. [Google Scholar] [CrossRef]
- Montone, C.M.; Capriotti, A.L.; Cavaliere, C.; La Barbera, G.; Piovesana, S.; Zenezini Chiozzi, R.; Laganà, A. Peptidomic strategy for purification and identification of potential ACE-inhibitory and antioxidant peptides in Tetradesmus obliquus microalgae. Anal. Bioanal. Chem. 2018, 410, 3573–3586. [Google Scholar] [CrossRef]
- Qi, W.; Su, R.X.; He, Z.M.; Zhang, Y.B.; Jin, F.M. Pepsin-induced changes in the size and molecular weight distribution of bovine casein during enzymatic hydrolysis. J. Dairy Sci. 2007, 90, 5004–5011. [Google Scholar] [CrossRef]
- Soto-Sierra, L.; Wilken, L.R.; Mallawarachchi, S.; Nikolov, Z.L. Process development of enzymatically-generated algal protein hydrolysates for specialty food applications. Algal Res. 2021, 55, 102248. [Google Scholar] [CrossRef]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrières, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the Tunisian brown seaweed Cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef]
- Yıldırım, A.; Mavi, A.; Kara, A.A. Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef]
- Chakou, F.Z.; Boual, Z.; Hadj, M.D.O.E.; Belkhalfa, H.; Bachari, K.; El Alaoui-Talibi, Z.; El Modafar, C.; Hadjkacem, F.; Fendri, I.; Abdelkafi, S.; et al. Pharmacological investigations in traditional utilization of Alhagi maurorum Medik. in saharan Algeria: In vitro study of anti-inflammatory and antihyperglycemic activities of water-soluble polysaccharides extracted from the seeds. Plants 2021, 10, 2658. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Whitney, C.G.; MacPherso, M.J.; Bhatti, S.; Banskota, A.H.; Stefanova, R.; McGinn, P.J. Biochemical characterization of microalgal biomass from freshwater species isolated in Alberta, Canada for animal feed applications. Algal Res. 2015, 11, 435–447. [Google Scholar] [CrossRef]
- Latsos, C.; Van Houcke, J.; Timmermans, K.R. The effect of nitrogen starvation on biomass yield and biochemical constituents of Rhodomonas sp. Front. Mar. Sci. 2020, 7, 563333. [Google Scholar] [CrossRef]
- Renaud, S.; Thinh, L.; Lambrinidis, G.; Parry, D. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Seixas, P.; Coutinho, P.; Ferreira, M.; Otero, A. Nutritional value of the cryptophyte Rhodomonas lens for Artemia sp. J. Exp. Mar. Biol. 2009, 381, 1–9. [Google Scholar] [CrossRef]
- Dunstan, G.A.; Brown, M.R.; Volkman, J.K. Cryptophyceae and rhodophyceae; chemotaxonomy, phylogeny, and application. Phytochemistry 2005, 66, 2557–2570. [Google Scholar] [CrossRef]
- Fernández-Reiriz, M.J.; Perez-Camacho, A.; Ferreiro, M.J.; Blanco, J.; Planas, M.; Campos, M.J.; Labarta, U. Biomass production and variation in the biochemical profile (total protein, carbohydrates, RNA, lipids and fatty acids) of seven species of marine microalgae. Aquaculture 1989, 83, 17–37. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Sánchez-Bayo, A.; Morales, V.; Rodríguez, R.; Vicente, G.; Bautista, L.F. Cultivation of microalgae and cyanobacteria: Effect of operating conditions on growth and biomass Composition. Molecules 2020, 25, 2834. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of exopolysaccharides production by Porphyridium sordidum and their potential to induce defense responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. [Google Scholar] [CrossRef]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Sui, Y.; Muys, M.; Van de Waal, D.B.; D’Adamo, S.; Vermeir, P.; Fernandes, T.V.; Vlaeminck, S.E. Enhancement of co-production of nutritional protein and carotenoids in Dunaliella salina using a two-phase cultivation assisted by nitrogen level and light intensity. Bioresour. Technol. 2019, 287, 121398. [Google Scholar] [CrossRef] [PubMed]
- Meijer, E.A.; Wijffels, R.H. Development of a fast, reproducible and effective method for the extraction and quantification of proteins of micro-algae. Biotechnol. Technol. 1998, 12, 353–358. [Google Scholar] [CrossRef]
- Corrêa, P.S.; Morais Júnior, W.G.; Martins, A.A.; Caetano, N.S.; Mata, T.M. Microalgae biomolecules: Extraction, separation and purification methods. Processes 2021, 9, 10. [Google Scholar] [CrossRef]
- Allard, B.; Templier, J. Comparison of neutral lipid profile of various trilaminar outer cell wall (TLS)-containing microalgae with emphasis on algaenan occurrence. Phytochemistry 2000, 54, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef]
- Xu, S.; Shen, Y.; Li, Y. Antioxidant activities of sorghum kafirin alcalase hydrolysates and membrane/gel filtrated fractions. Antioxidants 2019, 8, 131. [Google Scholar] [CrossRef]
- Kristofersen, K.A.; Amerongen, A.; Böcker, U.; Lindberg, D.; Wubshet, S.G.; de Vogel-van den Bosch, H.; Horn, S.J.; KristianAfseth, N. Fourier-transform infrared spectroscopy for monitoring proteolytic reactions using dryflms treated with trifuoroacetic acid. Sci. Rep. 2020, 10, 7844. [Google Scholar] [CrossRef]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta 2007, 1767, 1073–1101. [Google Scholar] [CrossRef]
- Kafle, B.; Böcker, U.; Wubshet, S.G.; Dankel, K.; Måge, I.; O’Farrell, M.; Afseth, N.K. Fourier-transform infrared spectroscopy for characterization of liquid protein solutions: A comparison of two sampling techniques. Vib. Spectrosc. 2023, 124, 103490. [Google Scholar] [CrossRef]
- León-López, A.; Fuentes-Jiménez, L.; Hernández-Fuentes, A.D.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Hydrolysed collagen from sheepskins as a source of functional peptides with antioxidant activity. Int. J. Mol. Sci. 2019, 20, 3931. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Spalding, K.E.; Ashton, K.M.; Board, R.; Butler, H.J.; Dawson, T.P.; Harris, D.A.; Hughes, C.S.; Jenkins, C.A.; Jenkinson, M.D.; et al. Fourier-transform infrared spectroscopy of biofluids: A practical approach. Transl. Biophotonics 2020, 3, e202000025. [Google Scholar] [CrossRef]
- Andersen, P.V.; Veiseth-Kent, E.; Wold, J.P. Analyzing pH-induced changes in a myofibril model system with vibrational and fluorescence spectroscopy. Meat Sci. 2017, 125, 1–9. [Google Scholar] [CrossRef]
- Polumbryk, M.; Ivanov, S.; Polumbryk, O. Antioxidants in food systems. Mechanism of action. Ukr. J. Food Sci. 2013, 1, 15–40. [Google Scholar]
- Moaveni, S.; Salami, M.; Khodadadi, M.; McDougall, M.; Emam-Djomeh, Z. Investigation of S. limacinum microalgae digestibility and production of antioxidant bioactive peptides. LWT 2022, 154, 112468. [Google Scholar] [CrossRef]
- Cian, R.E.; Martínez-Augustin, O.; Drago, S.R. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Int. Food Res. J. 2012, 49, 364–372. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gomez, B.; Barba, F.J.; Mora, L.; Perez-Santaescolastica, C.; Toldra, F. Bioactive peptides as natural antioxidants in food products—A review. Trends. Food. Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Alzahrani, M.A.J.; Perera, C.O.; Hemar, Y. Production of bioactive proteins and peptides from the diatom Nitzschia laevis and comparison of their in vitro antioxidant activities with those from Spirulina platensis and Chlorella vulgaris. Int. J. Food Sci. Technol. 2018, 53, 676–682. [Google Scholar] [CrossRef]
- Park, E.Y.; Imazu, H.; Matsumura, Y.; Nakamura, Y.; Sato, K. Effects of peptide fractions with different isoelectric points from wheat gluten hydrolysates on lipid oxidation in pork meat patties. J. Agric. Food Chem. 2012, 60, 7483–7488. [Google Scholar] [CrossRef]
- Sbroggio, M.F.; Montilha, M.S.; Figueiredo, V.R.G.; Georgetti, S.R.; Kurozawa, L.E. Influence of the degree of hydrolysis and type of enzyme on antioxidant activity of okara protein hydrolysates. J. Food Sci. Technol. 2016, 36, 375–381. [Google Scholar] [CrossRef]
- Okeke, E.S.; Enechi, O.C.; Nkwoemeka, N.E. Membrane stabilization, albumin denaturation, protease inhibition, and antioxidant activity as possible mechanisms for the anti-inflammatory effects of flavonoid-rich extract of Peltophorum pterocarpum (DC) K. Heyne (FREPP) Stem Bark. In Proceeding of the First International Electronic Conference on Antioxidants in Health and Disease, Virtual, 1–15 December 2020. [Google Scholar]
- Ruiz-Ruiz, J.C.; Matus-Basto, A.J.; Acereto-Escoffié, P.; Segura-Campos, M.R. Antioxidant and anti-inflammatory activities of phenolic compounds isolated from Melipona beecheii honey. Food Agric. Immunol. 2017, 28, 1424–1437. [Google Scholar] [CrossRef]
- Rastogi, S.; Iqbal, M.S.; Ohri, D. In vitro study of anti-inflammatory and antioxidant activity of some medicinal plants and their interrelationship. Asian J. Pharm. Clin. Res. 2018, 11, 2455–3891. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).