Abstract
The production of postbiotics for food applications has been growing in recent years owing to their biological potential and superior technological performance over probiotics. Their production involves the use of synthetic culture media, and in this work, we propose using sugarcane straw as a source of sugar and biological components and a sustainable alternative. Thus, this by-product was used as a substrate to produce a postbiotic extract using Saccharomyces cerevisiae as a fermentative microorganism. Sugarcane straw underwent a precedent saccharification step to release the fermentable sugars. The final extracts were characterized for their total content of sugars, phenolic compounds, organic acids, and their respective chromatographic profiles. Seventeen different polyphenols were identified with the predominance of three classes, the hydroxybenzoic acids, hydroxycinnamic acids, and flavonoids, where ferulic acid, 3,4-dihydroxybenzaldehyde, and 2,5-dihydroxybenzoic acid were most prevalent. The current work evaluated the potential use of this postbiotic extract for food applications, its antioxidant activity, gut microbiota modulatory effect, and intestinal anti-inflammatory potential. The resultant extracts showed considerable antioxidant activity and the ability to lower the pro-inflammatory mediators (i.e., interleukin 6, 8, and tumor necrosis factor-alpha) in Caco-2 cells. During the fecal fermentability assay, no modulatory effect was observed on the main beneficial bacteria, such as Lactobacillus and Bifidobacterium. Nevertheless, a significant increase in short-chain fatty acids, namely, acetate, propionate, butyrate, and valerate was observed. Moreover, the extract also demonstrated capacity to inhibit the proliferation of putrefactive bacteria such as Enterobacteriaceae. Finally, sustainable postbiotic extracts produced by S. cerevisiae fermentation using sugarcane straw as a substrate exhibited relevant biological properties with potential use as food and nutraceutical ingredients.
1. Introduction
The development of food products and ingredients with the potential to modulate gut microbiota has been growing in recent years, owing to the overall impact on health. These products can be prebiotics, probiotics, symbiotics, and more recently, postbiotics [1,2]. Definitions of these concepts have been published by the International Scientific Association of Probiotics and Prebiotics (ISAPP). Prebiotics are compounds that are specifically exploited by microorganisms giving a health advantage, while probiotics are living bacteria that, when consumed in the right amounts, help the host’s health. A mixture of live microorganisms and substrate(s) that host microorganisms use in a specific way to promote the host’s health is referred to as a symbiotic connection. Postbiotic extract is a “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” [3,4]. Postbiotics involves not only dead microorganisms, fractions, and lysates that may include cell wall fractions and exopolysaccharides, but it also involves metabolites such as enzymes, short-chain fatty acids (SCFA), peptides, peptidoglycans, teichoic acids, cell surface proteins, polysaccharides, and organic acids that are produced by living microorganisms that can be excreted and released after cellular lysis [5]. Due to the high heterogeneity of substances classified as postbiotics, they might have a high diversity of mechanisms of action. Because of their dose safety, long shelf-life, and their action as signaling molecules, they exert pharmacological properties such as anti-inflammatory, immunomodulatory, anti-obesogenic, antihypertensive, hypocholesterolemic, anti-proliferative, and antioxidant activities [6].
The production of postbiotics has been facilitated by the use of culture media containing synthetic compounds and saccharose, and the most widely used microorganisms are derived from lactic acid bacteria, specifically from the Lactobacillus genera and/or yeasts, particularly Saccharomyces cerevisiae [7]. Finding alternative substrate sources that are plentiful, affordable, and renewable is necessary, though, e.g., the agricultural and industrial by-products, which represent a significant opportunity to produce postbiotics rich in other compounds with enhanced bioactivities at reduced costs [7]. Some works have already reported using by-products as substrates for postbiotic production, such as cheese whey, milk permeate, and lignocellulosic biomass [8,9]. Recently, it was demonstrated that the juice from Parmentiera aculeata, a plant that is available in Mexico, increased synthesis of postbiotics like succinic acid, formic, acetic, propionic, and lactic acids as well as development of Lactobacillus plantarum [10].
Lignocellulosic biomass has grown in importance within the context of biorefinery, which aims to effectively use biomass to produce bioproducts with high-added value with sustainability. The most affordable and copious renewable carbon source currently accessible is lignocellulosic biomass [11]. Despite the potential that postbiotics have in the food industry, they may present some issues related to their production costs, hence, they are less competitive than their counterparts made from plants or chemical compounds. Using agro-industrial residues, such as sugarcane straw, presents an alternative to achieve an economical and sustainable process. Sugarcane (Saccharum officinarum L.) is a crop that produces a higher quantity of by-products discarded after harvesting, including straw residues [12]. According to estimates, 140 kg of dry straw are produced for every ton of sugarcane processed [13]. Straw may have two final destinations: it can be used for land cover or burned to supply energy to industries. The primary constituents of sugarcane straw are cellulose (34–43%), hemicelluloses (23–38%, primarily xylans), and lignin (11–25%), with extractives such polyphenols and inorganic compounds making up the remainder [14]. The lignocellulosic material undergoes hydrolysis to release monosaccharides that microbes can utilize for alcohol production [15]. Several studies have focused on sugarcane straw as a potential source of xylooligosaccharides (XOS), which are released by the action of enzymes, being well known for their prebiotic potential and considered as emerging prebiotics [16,17,18,19]. Saccharomyces cerevisiae and sugarcane straw hydrolysates have also been used for ethanol production from lignocellulosic biomass through fermentation [20]. This work has explored and demonstrated the potential of those extracts may have as a source of postbiotics that could modulate the gut microbiota while exerting other beneficial effects.
Therefore, the present work aims to demonstrate the use of sugarcane straw as a substrate to produce postbiotics by S. cerevisiae fermentation. The beneficial effect of the extracts on gut metabolic activity modulation, while helping to reduce intestinal inflammation by decreasing pro-inflammatory mediators and inhibiting potential pathogenic Enterobacteriaceae, was characterized, focusing on their potential use as food and nutraceutical ingredients.
2. Materials and Methods
2.1. Reagents
Appendix A contains a description of all the chemicals and reagents utilized in this investigation.
2.2. Production of Postbiotic Extract
2.2.1. Sugarcane Straw Saccharification Conditions
Raízen (São Paulo, Brazil) contributed the sugarcane straw. Samples were transported to the CBQF-UCP laboratory (Porto, Portugal) and immediately dried in a furnace (Nabertherm, Lilienthal, Germany) at 40 °C and crushed (knife mill SM100, Retsch, Haan, Germany) to a particle size ˂ 4 mm. Subsequently, it was sieved on a 900 µm sieve (Retsch GmbH, Haan, Germany), with an amplitude of 100 for 10 min.
Straw powder was dispersed in a supplemented buffer solution in a ratio of 1:20 (v:w). Supplement solution was composed of 10 g/L peptone, 5 g/L yeast extract, 2 g/L tri-ammonium citrate (C6H14N2O7), 2 g/L potassium phosphate dibasic (K2HPO4), 5 g/L sodium acetate (NaO2CCH3), 0.1 g/L magnesium sulfate heptahydrate (MgSO4·7H2O), and 0.05 g/L manganese (II) sulfate tetrahydrate (MNSO4·4H2O). This supplement was added to the sodium citrate buffer solution, containing 4.31 g/L of citric acid (C6H8O7) and 8.68 g/L sodium citrate (NA3C6H5O7), and then the pH was adjusted to 5.0 with NaOH (1 M) and autoclaved at 121 °C for 10 min.
The saccharification process was performed for 72 h with 36.2 FPU (filter paper units) of Celluclast® per gram cellulose at 50 °C, 150 rpm [21]. A saccharification control was performed with a sample under the same conditions without enzyme addition. All the experimental conditions were performed in duplicate. Along the process, samples were collected after 0, 24, 48, and 72 h, and the total sugars decay was determined along the saccharification process.
At the end of saccharification, the biomass was separated by centrifugation (15 min at 5000 rpm) and then filtered (0.22 µm, Sartorius stedim, Gottingen, Germany) under sterile conditions.
2.2.2. Fermentation Conditions and Extract Production
Overnight, inoculum of Saccharomyces cerevisiae DSM 70449 (DMSZ-German Collection of Microorganisms and Cell Cultures, Braunschweig, Germany) was prepared in yeast malt broth and incubated at 30 °C. After saccharification, the extract was used for fermentation with an inoculum of S. cerevisiae with a final cellular concentration at 106 CFU/mL determined by optical density measurement at 600 nm (UV-1900 UV-VIS spectrophotometer, Shimadzu, Kyoto, Japan) [22] and incubated at 30 °C, for 72 h (in duplicate).
At 0, 24, 48, and 72 h, samples were taken for evaluation of cellular concentration, pH, and total sugar content measurements. At the end, sonication was applied for mechanical cell lysis (CY-500, Optic Ivymen System, Comecta; Barcelona, Spain) for 10 min with pulses of 15 s on and 45 s off, at the temperature of 20 °C and an amplitude of 70%. After that, samples were centrifuged (30 min at 4480× g), and the supernatant was stored at −80 °C before freeze-drying (gamma 2–16 LSCplus, Martin Christ, Osterode am Harz, Germany). The obtained powder was further characterized. The entire process flow is described in Figure 1.
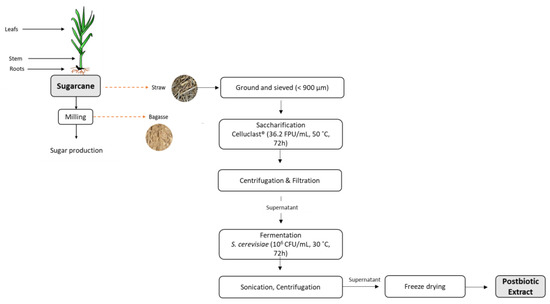
Figure 1.
Scheme of sugarcane straw postbiotic extract production.
2.3. Cellular Concentration Measurements
Viable counts were determined in samples of sugarcane straw fermented with S. cereviseae at 0, 24, 48, and 72 h by employing the spread technique to plate the samples on potato dextrose agar (PDA), serial decimal dilutions in peptone water, and a 24-h incubation period at 30 °C. All assays were performed in duplicated.
2.4. Postbiotic Extract Chemical Characterization
2.4.1. Total Sugars
Total sugars were evaluated by the phenol-sulphuric acid method [23]. Briefly, to 80 μL of each sample, 150 µL of the 5% phenol solution and 1 mL of sulphuric acid (95%, v/v) were combined and added. Samples were incubated in an oven at 100 °C for 10 min. After cooling, the absorbance was measured in a spectrophotometer (Shimadzu, Kyoto, Japan) at 490 nm. All assays were performed in duplicate. Glucose was used for the calibration curve (5–0.3125 mg/mL).
2.4.2. Quantification of the Total Phenolic Content
The Folin–Ciocalteu assay was performed according to the method described by [24]. The analysis samples were previously prepared at 25 mg/mL in deionized water. In a test tube, 50 µL of sample was mixed with 50 µL of Folin–Ciocalteu’s reagent at 1 N, 1000 µL of sodium carbonate at 7.5% (w/v), and 1400 µL of deionized water. The mixture was incubated for 1 hour in the dark at room temperature, after which the absorbance at a wavelength of 750 nm was measured. Gallic acid (0.493 to 0.062 mg/mL) calibration curve was used to calculate the total phenolic content, with results expressed as mg of gallic acid equivalents (GA) per g of dry weight (dw) sample.
2.4.3. Organic Acids, Sugars, and Short-Chain Fatty Acids Analysis by HPLC-RID
The dry extract was previously dispersed in water at a concentration of 25 mg/mL and filtered with 0.45 µm filter (Minisart; Sartorius stedim, Gottingen, Germany). Chromatographic analysis was performed with high-performance liquid chromatography, Agilent 1200 HPLC system with a refractive index (RID) detector (1260 Infinity II, Agilent Technologies; Santa Clara, CA, USA), and diode array detector (DAD) operating at 50 °C. Separation was performed in the Aminex HPX87Pb column (BioRad, Hercules, CA, USA) with a mobile phase of 0.005 M of sulphuric acid at a flow rate of 0.6 mL/min. Following centrifugation, the supernatants were filtered (0.22 µm) and immediately analyzed by HPLC in duplicate [25]. Using their matching calibration curves, malic acid, lactate, acetate, butyrate, ethanol, xylose, mannose, galactose, glucose, arabinose, and fructose were identified and measured.
2.4.4. Individual Phenolic Compounds Identification by LC-ESI-QqTOF-HRMS
The extracts were dispersed in ultra-pure water at 50 mg/mL and filtered with 0.45 µm filter (Minisart; Sartorius stedim, Gottingen, Germany) before analysis by LC-ESI-UHR-QqTOF-MS [26]. A UHPLC UltiMate 3000 Dionex (Thermo Scientific) equipped with an ultrahigh resolution was used to carry out the separation. Mass spectrometer with 50,000 full-sensitivity resolution (FSR) using Qq-time-of-flight (UHR-QqTOF) (Impact II, Bruker Daltonics, Bremen, Germany) was employed. Utilizing an Acclaim RSLC 120 C18 column (100 mm × 2.1 mm, 2.2 m), metabolites were separated (Dionex). Acetonitrile with 0.1% formic acid and 0.1% aqueous formic acid were the mobile phases (solvent B). The following gradient conditions were used during the separation process, which lasted 24.5 min, at a flow rate of 0.25 mL/min: 0 min, 0% B; 10 min, 21.0% B; 14 min, 27% B; 18.30 min, 58%; 20.0 min, 100%; 24.0 min, 100%; 24.10 min, 0%; and 26.0 min, 0%. The injection had a 5 L volume.
Mass spectrometers with ultrahigh-resolution quadrupole–quadrupole time-of-flight (UHR–QqTOF) are employed. In an auto MS scan mode, spectra with a m/z range of 20 to 1000 are obtained, and parameters for MS analysis are determined using these data. The selected parameters included the end plate offset voltage of 500 V, the capillary voltage of 3.0 KV, the drying gas temperature of 200 °C, the drying gas flow of 8.0 L/min, the nebulizing gas pressure of 2 Bar, the collision radio frequency (RF) of 250 to 1000 Vpp, the transfer time of 25 to 70 s, the collision cell energy of 5 eV, and the pre-pulse storage of 6 s. Sodium formate clusters were utilized for post-acquisition internal mass calibration, and it was injected using a syringe pump at the beginning of each chromatographic analysis. Accurate mass and isotope rate estimates with the designated mSigma proved the compound’s elemental composition (Bruker Daltonics). The mSigma results provided assurance that the lowest chemical nature for mass was presented. A compound’s precise mass [M − H]− was used to identify it. Quantification (μg/mL) was performed by external calibration, and for the quantification of hydroxybenzoic acids, the standards 2,5-dihydroxybenzoic acid, 4-hydroxybenzaldehyde, 4-hydroxybenzoic acid, 3,4-dihydroxybenzalhedyde, and protocatehuic acid were used, respectively, for each compound. For hydroxycinnamic acids, p-coumaric acid and the ferulic acid were also used for feruloylquinic acids quantification. The flavonoids standards vitexin, luteolin, and tricin were used for the quantification of isovitexin 2″-O-arabinoside, luteolin-8-C-glucoside isomers, and tricin derivatives, respectively.
2.5. Postbiotic Extract Biological Characterization
2.5.1. Impact of the Postbiotic Extract on Gut Microbiota
Fermentability assay to assess the impact of the extract on gut microbiota was performed using tube-independent batch culture fermentations, which were carried out using fresh human fecal inoculum from fecal samples of healthy donors [27]. The composition of basal nutrient medium was made according to [28]. In this experiment, each condition tested had 3 fermentation sampling tubes (i.e., 0, 24, and 48 h) and was performed in triplicate. The tested conditions were: (1) sugarcane straw with S. cerevisiae; (2) fructo-oligosaccharides (as a positive prebiotic control); and (3) inoculum control (media with only fecal inoculum). The substrates were tested at 1% (w/v) (i.e., 10 g/L). The batch fermentation was conducted under anaerobic conditions at 37 °C for 48 h, in which fermentation sampling tubes were collected from their respective time point for bacterial enumeration by plating, pH measurements, and short-chain fatty acids (SCFA) and lactate analysis performed by HPLC, as previously described [27].
2.5.2. Radical Scavenging Activity
ABTS Radical Cation
The 2.2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS+) radical cation test was used to calculate the total antioxidant capacity. Potassium persulfate (2.45 mM) and ABTS (7 mmol/L) were combined in a 1:1 (v/v) ratio to create the ABTS+ solution. On a 96-well microplate, the reaction was carried out in five minutes at 30 °C with diluted ABTS (OD of 0.700 ± 0.020 at 734 nm) and the samples (15 μL of each sample in duplicated reacted with 200 μL of ABTS). After that, OD was measured in a microplate reader (Synergy H1 microplate reader, Biotek, Winooski, Vermont, EUA) at 734 nm [29]. For the calibration curve, Trolox was used (0.0075–0.075 mg/mL). A serial of sample dilutions were performed (0.39–12.5 mg/mL) in methanol to evaluate the IC50 (mg/g dw).
DPPH Radical Cation
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical test was used to calculate the DPPH. The working solution was created up to a final OD of 0.600 ± 0.100 at 515 nm after the DPPH solution (600 μM) was made in methanol. After mixing the sample (25 μL) and DPPH (175 μL) in duplicate, the OD was measured at 515 nm in a microplate reader after 30 min at 25 °C (Synergy H1 microplate reader, Biotek, Winooski, Vermont, EUA) [29]. For the calibration curve, Trolox was used (0.0075–0.075 mg/mL). A serial of sample dilutions was performed (0.14–12.5 mg/mL) in methanol to evaluate the IC50 (mg/g dw).
Oxygen Radical Absorption Capacity (ORAC)
The antioxidant capacity was evaluated using the Oxygen Radical Absorbance Capacity (ORAC) assay [30]. In summary, the reaction took place at 40 °C in 75 mM phosphate buffer (pH 7.4), and the final test mixture (200 μL) contained the following compounds: Trolox (10–80 μM) or sample (0.08–1.667 mg/mL), fluorescein (70 nM), AAPH (14 mM). For 137 min (104 cycles), the fluorescence was captured using a 485 nm excitation and 580 nm emission wavelength (Synergy H1; BioTek, Winooski, VT, USA). We utilized 96-well black polystyrene microplates from Nunc in Denmark. Daily, AAPH and Trolox solutions were prepared, and fluorescein stock solution (1.17 mM) in 75 mM phosphate buffer was diluted (pH 7.4). At least three separate runs were carried out for each sample, and all reaction mixes were prepared in duplicate. The results were expressed as µM of Trolox equivalent per g dry weight.
2.5.3. Cytotoxicity Evaluation
Three different cell lines were used in this work Caco-2 cells. All cells were culture in a humidified atmosphere at 37 °C with 5 CO2 as monolayers. They were cultured in DMEM with 4.5 g/L, L-glutamine without pyruvate, 10% FBS, 1% Pen-Strep and 1% of NEAA. Hep G2 cells were cultured in the same media but without NEAA. Caco-2 cells were used between passages 23 and 36.
As previously mentioned, cytotoxicity was assessed in accordance with the International Organization for Standardization (ISO, 2009) standard [22,31].
On a 96-well microplate, cells were planted at a density of 1 × 104 cells per well (Nunclon Delta, Thermo Scientific, Waltham, MA, USA). After 24 hours, the culture medium was carefully removed and replaced with culture medium that included extract (1.56 and 50 mg/mL). The DMSO at 10% (v/v) was used as a death control and plain culture media was used as growth control. Using a micro-plate reader, the fluorescence (excitation: 560 nm; emission: 590 nm) was quantified (Synergy H1 microplate reader, Biotek, Winooski, Vermont, EUA) after 24 h incubation followed by Presto Blue addition. All assays were performed in quadruplicate.
2.5.4. Immunomodulatory Effect
For this assay, Caco-2 cells were seeded at 2.5 × 105 cells/well and incubated for 24 h at 37 °C [32]. After that, the extract was added (0.3125 mg/mL) after culture media replacement, and the plate was re-incubated again for 24 h. The IL-1β (Invitrogen, Waltham, MA, USA) was used as inflammation control, and basal activity control was made with plain media. The supernatants were stored at −80 °C for further analysis.
According to the manufacturer’s instructions, the Human IL-6 Elisa Kit High Sensitivity (Abcam, Cambridge, UK), the Legend Max Human Elisa Kit IL-8, and the Legend Max Human Elisa Kit TNF-α (BioLegend, San Diego, CA, USA) were used for the enzyme-linked immunosorbent assay (ELISA) to detect Interleukins 6 (IL-6), 8 (IL-8), and Tumor Necrosis Factor alpha (TNF-α). Findings were given in interleukin (IL) pg/mL of sample. Relative Percentage of Production, which compares the interleukin quantity in the basal (non-stimulated) control to the results from any proteomic-based test, is a measure of the test’s effectiveness. The interleukin content of the basal control was 100%.
2.6. Statistical Analysis
The normality distribution was assessed using the Shapiro-Wilk test with a significance threshold of p < 0.05. The Tukey’s post hoc test was employed for the statistically significant variables and the ANOVA was used to test for significant differences (homogeneity of variance was adopted at the p < 0.05 significance level). Version 14.0.0.15 of STATISTICA was the program used.
3. Results and Discussion
3.1. Saccharification and Fermentation Process
The sugarcane straw went through a saccharification process with a commercial cellulase to breakdown cellulose and hemicellulose to release monosaccharides that will be the sugar source for the yeast. The saccharification occurred for 72 h, and the total sugar content increased from 1.72 ± 0.10 to 3.46 ± 1.06 mg/mL. Saccharification is the conversion of cellulose biomass to useful substances such as fermentable sugars through the enzymatic hydrolysis of the polysaccharide. Cellulase application during saccharification operates mainly in cellulose and hemicellulose degradation, releasing more water soluble carbohydrates [14].
This final sugar rich mixture was applied for fermentation with S. cerevisiae, and the yeast growth and total sugars content along the 72 h are described in Figure 2. It was possible to observe an increase in yeast growth of ca. 5 log cycles accompanied by a consumption of ca. 2 mg/mL of the total sugars.
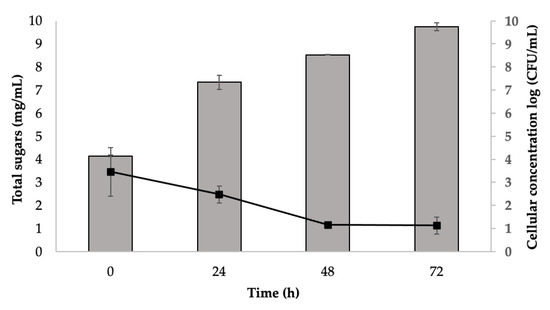
Figure 2.
Total sugar content (■) (mg/mL, average ± SD) and S. cerevisiae growth (■) (Log cycles, average ± SD) measured in cell broth along 72 h fermentation process of sugarcane straw that went through saccharification.
3.2. Postbiotic Extract General Composition
The final extract, obtained after freeze-drying the supernatant of sonicated cells, resulting from sequential sugarcane straw saccharification and fermentation with S. cerevisiae, was assessed in terms of total phenolic and total protein content and compared with samples that crossed the same process without the yeast fermentation step (control) (Table 1).

Table 1.
Levels of the major biochemical constituents (mg/g DW; means ± SD) in sugarcane straw powder extract before and after S. cerevisiae fermentation.
In control samples, the total phenolic content was 12.20 mg gallic acid equivalents/g dry weight extract, and after yeast fermentation, it increased (p < 0.05) to 16.15 mg GA/g dw. Fermentation of soybean meal with S. cerevisiae demonstrated an increase in total phenolic content [33]. In this work, the straw extract obtained after saccharification may have bounded polyphenols into proteins or polysaccharides from the cell walls, and thus, yeast had the capacity to hydrolyze flavonoid glycosides into aglycones [34,35] through the action of β-glucosidases releasing additional total phenolics compounds that will contribute to a higher antioxidative effect [36].
The protein content in extracts after yeast fermentation is due to the release of extracellular protein throughout the fermentation process and/or that which is initially present in the extract used as a substrate [37]. However, in this work, the total protein content of the extract remained similar (p > 0.05) in samples with and without yeast fermentation (32–33 g/100 g). Depending on the biomass used for fermentation, different behaviors were reported in the literature for protein contents after yeast fermentation; one example of that was in okara, a soybean by-product, which showed an increase from 32.8 to 40.5 g/100 g in protein content after S. cerevisiae fermentation [38].
3.3. Organic Acids and Sugars Profiles
The analysis of the main sugars and organic acids was carried out by high-performance liquid chromatography with a refractive index detection (HPLC-RID) (Table 1).
The only monosaccharide detected was glucose with an amount of 41.5 mg/g dw, which disappeared after S. cerevisiae fermentation. The same happened with the disaccharide cellobiose, with amounts of 12.5 mg/g dw detected in the non-fermented extract. The presence of these sugars results from their accumulation at the end of saccharification process, and then the yeast uses the glucose as carbon source, resulting in a decrease in its content [39]. As opposed to that, S. cerevisiae is not capable of hydrolyzing cellobiose [40]. The decrease in cellobiose content observed in this study after yeast fermentation may be related to the fact that the extract was sonicated after fermentation, a which process may have promoted the hydrolysis of oligomers released in saccharification leading to a breakdown of disaccharides [41].
The acetic acid was the main organic acid detected in all samples, and increases after yeast fermentation were observed from 80.7 to 194.17 mg/g of dry extract. Regarding the other organic acids, lactic acid and propionate were only detected in extracts obtained after yeast fermentation. The lactic acid reached amounts of 9.83 mg/g of dry extract, respectively. It was shown that the wild-type strains of Saccharomyces cerevisiae only create trace amounts of D-lactate, most likely because of the action of the methylglyoxal bypass. Saccharomyces cerevisiae does not produce significant amounts of organic acids spontaneously [42,43].
Due to the overexpression of aldehyde dehydrogenase-coding genes, the increase in acetic acid level seen following yeast fermentation may be connected to the yeast’s reaction to osmotic stress [44].
3.4. Phenolic Compounds Profile
The polyphenolic profile of the sugarcane straw extracts obtained after a sequential enzymatic saccharification and fermentation with S. cerevisiae was compared with samples without yeast fermentation to verify the impact of this process on polyphenols content. A total of 17 compounds were identified in any resulting extract (Table 2), but it was only possible to quantify 14 (Table 3).
Table 2.
LC-ESI-UHR-QqTOF-MS data of phenolic compounds identified in straw extract after saccharification.
Table 3.
LC-ESI-UHR-QqTOF-MS data of phenolic compounds quantified in straw extract after saccharification and with and without S. cerevisiae fermentation (means ± SD, µg/g dry extract).
The phenolic profile obtained was identical to other sugarcane materials such as sugarcane leaves genotypes [45], molasses extract [46], and bagasse [47]. Although the category of compounds was the same throughout all studies, the diversity of compounds discovered, and their concentration were somewhat distinct from those found in this study. Most of the reports in the literature described phenolic extractions using organic solvents such as ethanol [47], but in this work, the solvent used was water, and the extract went through a sterilization process before the saccharification process. The exposure of the extract to high temperature and low pressure (121 °C, 10 min 1.1 atm) applied for sterilization may has been responsible for the lower content of polyphenols detected in this extract [48]. The aqueous environment was not favorable for flavonoids in the extract, since sugarcane straw is characterized by the presence of several compounds such as luteolin and apigenin with moieties of glucose or arabinoside and O-glycosylated or the presence of tricin and luteolin aglycone [49,50].
Within hydroxybenzoic acids, six distinct compounds were found, and the two most representative were 2,5-dihydroxybenzoic acid isomer 2 (24.56 ± 4.35 μg/g dw) and 4-hydroxybenzaldehyde (17.71 ± 0.17 μg/g dw), with the ions m/z 153 [M − H]− (C7H5O4) and m/z 121 [M − H]− (C7H5O2), respectively. Around 27% of the extract’s measured components fall into this category. High levels have been linked to systemic infections. Hydroxybenzoic acid derivatives are in charge for plants’ defense response against pathogens [51]. In addition to serving plant purposes, they also possess intriguing biological characteristics that make them appealing to the food business. 2,5-Dihydroxybenzoic acid has shown interesting bioactive properties such as antidiabetic potential [52].
After yeast fermentation, the content of all the compounds within this class decreased except for 3,4-dihydroxybenzaldehyde, which increased from 7.17 to 24.82 μg/g dw. 3,4-dihydroxybenzaldehyde is known to be covalently linked to cell wall polysaccharides, and during yeast fermentation, there may occur a release of these compounds from polysaccharides that are more accessible to quantify. Similar results were reported for sorghum fermented with a lactic bacteria [53].
Hydroxycinnamic acids were the predominant class representing 71% of whole-extract composition. The presence of quinic acid esterified with ferulic acid units was detected in four compounds. Additionally, free ferulic and p-coumaric acids were detected. Ferulic acid promotes the development of lignin in mature cane, making it a molecular marker that changes with genotypes, cell types, and tissues within the same plant [54]. Immunomodulatory effect by ferulic and p-coumaric acids has been reported due to its antioxidant capacity that exerts a cytoprotective effect [55].
Ferulic acid (m/z 193 [M − H]− (C10H9O4)) was present in higher quantities with 110.36 ± 0.01 μg/g dw. After yeast fermentation, the only hydroxycinnamic acid that decreased was ferulic acid, by ca. 39%. The remaining compounds did not present a significant variation (Table 3).
Saccharomyces cerevisiae growth may be inhibited by some phenolic compounds, and to overcome this issue, certain phenolic compounds can be catabolized and metabolically processed in situ by yeast. According to published reports, yeast can convert ferulic acid into some intermediates, such as the isomer dihydroferulic acid, in the first two hours of cultivation. This yeast can also produce benzeneacetic acid, 2′,5′-dihydroxyacetophenone, 5-allyl-1-methoxy-2,3-dihydroxybenzene, 4-hydroxyphenylethanol, 4-vinylguaiacol, and phenylethyl alcohol [56]. Decarboxylases, the most well-known of which is phenyl acrylic acid decarboxylase, were demonstrated to promote the conversion of ferulic acid in S. cerevisiae [57].
Another significant class of phenolic chemicals that are prominently present in sugarcane are flavonoids. Among these, luteolin with glucose moieties and tricin with glucose, glucuronide, or neospheroside moieties were found. The luteolin and isovitexin derivatives were detected, but they were lower than the quantification limit. Tricin-7-O-glucoside (m/z 491 [M − H]− (C25H31O10)), tricin-7-O-rhamnosyl-glucuronide (m/z 651 [M − H]− (C29H31O17)) presented the higher content among all the flavones identified with 1.57 ± 0.02 and 1.27 ± 0.02 μg/g dw, respectively. With only 2% of all detected phenolic compounds belonging to this class, the extract had the lowest representation of these compounds. Many studies claim that due to tricin’s pharmacological efficiency, bioactive characteristics, and low toxicity, it has demonstrated its clinical utility over time [58]. Due to the qualities it possesses, such as anticancer capabilities [59,60], antiobesity [61], antidiabetic [62], antioxidant [63,64], anti-inflammatory [65,66], and antiviral [67] tricin is regarded as a promising nutraceutical. The fermentation process led to a decrease in tricin-7-O-glucoside from 1.57 to 0.75 μg/g dw, and tricin-7-O-rhamnosyl-glucuronide presented an increase of 8%.
3.5. Antioxidant Activity
The antioxidant capacity of the extracts was determined by ABTS and DPPH through the IC50 (mg/mL) and ORAC results were expressed in μmol Trolox equivalents/g dry weight (Table 4).
Table 4.
Antioxidant capacity of sugarcane straw extract after cellulose saccharification with and without (control) S. cerevisiae fermentation.
According ABTS, DPPH, and ORAC methods, the capacity of the extract to neutralize free radicals increased after S. cerevisiae fermentation of sugarcane straw extract, since a significant decrease (p < 0.05) in IC50 was observed for ABTS and DPPH and an increase in (p < 0.05) Trolox equivalents for ORAC (Table 4), whose reaction mechanism may be through hydrogen atom transfer.
Different antioxidant activities have been reported for different S. cerevisiae strains. A study between PY S. cerevisiae var. boulardii and S. cerevisiae BY4742 showed that var. boulardii had 6–10-fold higher scavenging activity (DPPH), and this was due to higher content of total phenolics and flavonoids [68]. Another work described an increase in the antioxidant capacity of apple juice fermented sequentially with S. cerevisiae, dealcoholization, and L. plantarum [69]. It was also reported that autolysates of baker’s yeast, Pichia fermentans BY5 and Issatchenkia orientalis BY10, showed antioxidant activity in scavenging of DPPH radical, nitric oxide, hydroxyl radical, and metal ion chelating [70,71,72].
The bioactive peptides in yeast hydrolysates and other antioxidant substances such glutathione, Maillard reaction products, sulfur-containing amino acids, and polysaccharides are likely what give them their antioxidative properties [73].
Besides the contribution of yeast autolysates to the antioxidant capacity, it was also reported in the literature that S. cerevisiae had the capacity to increase the bioaccessibility of phenolic content, thus increasing the radical scavenging properties [33]. The fermentation of 4 cereals (wheat, buckwheat, wheat germ, barley, and rye) by L. rhamnosus and S. cerevisiae showed an increase in the antioxidant activity [74]. Additionally, an increased antioxidant capacity was observed in a Lactobacillus paracasei postbiotic extract tested in liver of male albino rats [75]. Antioxidant activity in butter samples increased after the addition of ferulago angulata extract when combined with postbiotic compounds produced by Lactiplantibacillus plantarum subsp. plantarum [76].
As previously described in the present work, the total phenolic content increase observed in straw extract obtained after saccharification and yeast fermentation may be contributing to antioxidant activity increase.
3.6. Impact of Postbiotics on Gut Microbiota Modulation
The effect of the extract in gut microbiota, an in vitro fermentation model was performed using feces from five donors. Fermentation sampling was performed at times 0, 24, and 48 h, to quantify the pH, the total and specific bacteria, such as Enterobacteriaceae, lactic acid bacteria, and Bifidobacterium, and finally the short-chain fatty acids (SCFA). Obtained data were compared with two controls, one positive prebiotic designated as FOS and one negative only with inoculum control (IC). The use of FOS was for control of the impact on gut modulation in terms of microorganisms composition, in the evaluation of the main beneficial bacteria such as Lactobacillus and Bifidobacterium.
The common pH of the human colon varies between 6.0 and 8.0 [77], and in the negative control (IC), no significant differences (p > 0.05) were observed after 48 h. On the other hand, FOS presented a high decrease from 7.15 to 3.64, and in the straw extract, pH decreased from 7.24 to 6.72 (Figure 3). The pH decrease, although slight, may create a favorable environment for the reproduction of some beneficial bacteria while inhibiting harmful microorganisms.
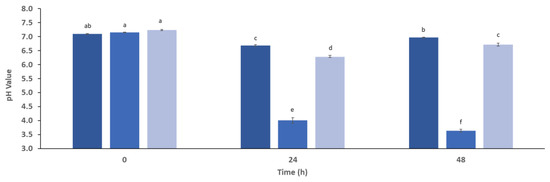
Figure 3.
The changes in pH value of simulated gut microbiota fermentation (five donors) along 48 h in the presence of inoculum control that represents media with only fecal inoculum (IC, ■), fructooligosaccharides (FOS, ■), and straw extract after fermentation with S. cerevisiae (■). a,b,c,d,e,f Different letters mean significant differences (p < 0.05) between samples.
The fermented straw extract as well as FOS used as the positive control and no extract addition as negative control were tested and were incubated for 48 h with fecal bacteria. Figure 4 depicts modifications in bacterial populations following this therapy.
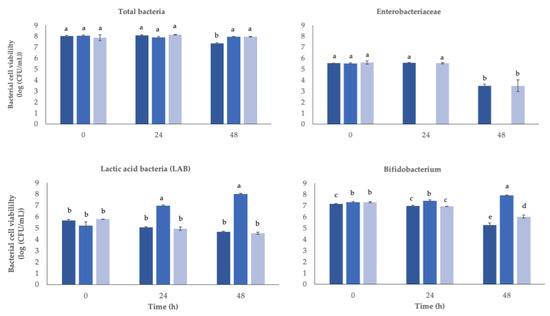
Figure 4.
Changes in bacterial viable cell counts (log CFU/mL) in batch cultures after 48 h at 37 °C of inoculation of different substrates. Inoculum control (IC, ■), fructo-oligosaccharides (FOS, ■), and straw extract after fermentation with S. cerevisiae (■). a,b,c,d,e Different letters mean statically significant (p < 0.05) differences between samples.
After 24 or 48 hours of fermentation, no significant changes (p > 0.05) in the total number of bacteria were found, presenting a value of 8 log (CFU/mL) over the 48 h.
In lactic acid bacteria (LAB), no significant differences (p > 0.05) were seen along fermentation time in the inoculums with straw extract or in the negative control. On the other side, the positive control (FOS) increased from 5.2 to 8.0 log (CFU/mL) after 48 h. Bifidobacterium, where no extract was added, presented a decrease by the end of 48 h, while in the positive control, the presence of FOS stimulated this group of bacteria to increase from 7.3 to 7.9 log (CFU/mL), and in the presence of straw extract, the group decreased (p < 0.05) from 7.3 to 6.0 log (CFU/mL).
The group of Enterobacteriaceae cells were not detected in positive controls after 24 and 48 h fermentation, while in the negative control and in the presence of straw extract, the group decreased significantly only after 48 h fermentation, from 5.5 to 3.5 log (CFU/mL).
The inoculum with FOS was the only one that promoted the growth of LAB and bifidobacteria, and this was due to its prebiotic capacity. One to three molecules of fructose bonded directly to the fructose residue of sucrose with β-1,2 linkage form the non-digestible oligosaccharide known as FOS. Short-chain fatty acids including acetic, propionic, and n-butyric acids, carbon dioxide, hydrogen, methane, amino acids, and vitamins are produced during the fermentation of FOS by the gut bacteria. The increase in SCFA decreases the pH, which is favorable for Bifidobacterium and Lactobacillus growth, while it suppresses the pathogenic bacteria [78], also seen in this work (Figure 4).
Prebiotics primarily target strains of Lactobacillus and Bifidobacterium, among others, which can combat chronic diseases because intestinal fermentation generates SCFAs. They are produced as a byproduct of the intestinal microbiota’s metabolism, primarily from dietary fiber that is not digestible [79]. Figure 5 shows the values obtained for SCFA in the cultures.
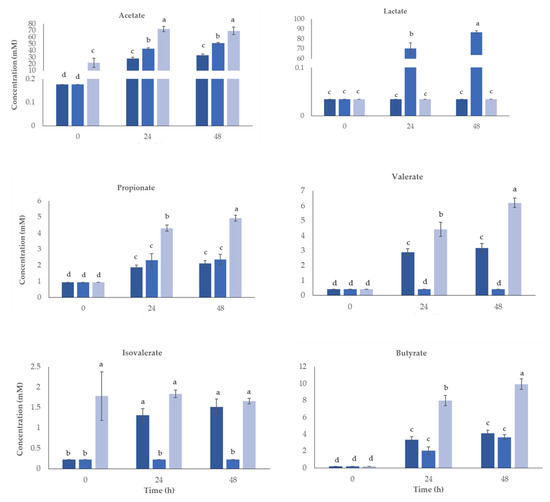
Figure 5.
Concentration (mM) of the short-chain fatty acids (SCFA) and lactate produced for 48 h of human colonic fermentation using straw extract obtained after fermentation with S. cerevisiae. Inoculum control (IC, ■), fructo-oligosaccharides (FOS, ■), and straw extract after fermentation with S. cerevisiae (■). a,b,c,d Significant variations (p < 0.05) between samples for each acid are denoted by different letters.
Without the interference of a nutritional source, the IC gives background data on the intrinsic and established gut microbiota metabolite production. As expected, the IC has the lowest SCFA production because there is no additional substrate to feed the intestinal bacteria. After FOS administration, an increase in SCFA production was observed. Thus, the administration of prebiotics induces the synthesis of SCFA, which has been reported to exert positive effects against the inflammatory behavior of the intestinal barrier. According to these findings, prebiotics not only aid in the growth and development of particular colonic bacteria but also encourage the synthesis of bacterial metabolites that may have positive health effects [80].
The acetate was the main SCFA quantified in all samples after 48 h fermentation (p < 0.05) when compared with negative and positive controls. The last two increased along fermentation time but to a lower extent. Fermentation for 24 h was enough for the maximum content of acetate, showing an increase from 21.6 to 69.3 mM. According to reports, this material is the most common substance in the large intestine and the main consequence of bifidobacterial fermentation in a human fecal environment [81,82]. The abundance of acetate is crucial for reducing inflammation and blocking pathogen invasion such as Enterohaemorrhagic Escherichia coli [83,84].
Lactate increased significantly in the presence of FOS, but no differences were observed in negative control or in the presence of straw extract.
Butyrate increased along fermentation for all conditions tested, but the increase observed was higher when fermentation occurred in the presence of straw extract, presenting 9.94 mM by comparison with FOS, which only had 3.6 mM after 48 h. Several in vitro investigations have shown that butyrate can cause apoptosis in colonic tumor cell lines, and it is thought that butyrate is a beneficial metabolite of gut bacterial activity [85]. Butyrate formation occurs in certain Firmicutes bacteria, through two mechanisms, one via butyrate kinase and another by butyric acetate CoA transferase [86]. Butyric acid has beneficial effects on the intestinal mucosa and influences cell development, making it crucial for human health. It has anti-inflammatory qualities and lowers the likelihood of developing colon cancer [81,87]. Colitis treatment options have also included butyrate irrigation (enema). As a result, substances that stimulate the formation of butyrate at intestinal epithelial cells are of interest to the food industry supplements.
Regarding propionate concentrations, the most significant increase was in the presence of straw extract, increasing from 0.95 to 4.93 mM, while the FOS inoculum had a lower increase (0.95–2.37 mM). Due to its anti-lipogenic, cholesterol-lowering, anti-inflammatory, and anti-carcinogenic properties, propionate may have positive impacts on health [88]. Propionic acid content may be related to the high numbers of Bacteroides present or to the presence of specific compounds in the extracts [89]. Its production was associated with side chains found in xilooligosaccharides [90]. Another reason may be the presence of oligosaccharides derived from hemicellulose, which is a rich source of xylan, which is composed by xylooligosaccharides (XOS) [91]. According to some reports, XOS promotes the growth of Lactobacillus at the expense of the growth of dangerous bacteria, leading to an increase in short-chain fatty acids including butyrate, acetate, and propionate [18,92].
Richness of postbiotic extract in lactate and acetate can positively influence downstream butyrate and propionate content at the level of the gut when converted by butyrate and propionate producers [93]. Microbes such as Anaerobutyricum and Anaerostipes species can convert lactate to butyrate via the CoA-transferase pathway. Moreover, there are two ways that lactate can be converted to propionate: either through the acrylate pathway, which is used by microorganisms such as Coprococcus catus and Megasphaera elsdenii, or by the succinate pathway, which is used by microbes such as Veillonella species [94,95,96].
Valerate did not increase in the positive control; however, straw extract induced an increase (p < 0.05) from 0.4 to 6.2 mM after 48 h. In the negative control, also visible was an increase in valerate content but to a lower extent, increasing to 3.2 mM. The isovalerate only revealed differences in the negative control, which increased from 0.2 to 1.5 mM.
The treatment of sugarcane straw during saccharification with cellulase can release xylooligosaccharides (XOS) but also cello-oligosaccharides (COS), which are oligomers of β-1,4-linked D-glucose units that have proved prebiotic effect by stimulating in vitro growth of several Lactobacillus and Bifidobacterium strains [97,98]. In this work, no significant growth was observed for Bifidobacterium or lactic acid bacteria (LAB), probably because the amount of oligomers released was not strong enough to stimulate bacteria growth but was enough to increment their production of SCFA. It is described in the literature that XOS or COS production from lignocellulosic material requires other enzymes such as xylanases or higher temperatures and alkaline pH to increase the material breakdown [16].
Besides the potential increase in oligomers in the extract medium that may exert prebiotic effect, the observed decrease in Bifidobacterium growth after 48 h and no changes observed in the growth of LAB may be related to the presence of some polyphenols. It was reported that depending on the phenolic compounds, they may stimulate the Bifidobacterium growth, as was the case with coumaric acid, caffeic acid, and vanillic acid [99], or may inhibit it, as was the case with some flavonoid aglycones [100].
Studies on both humans and animals (rats, pigs, and chickens) revealed an increase in the growth of Lactobacillus and a decrease in Entorabacteriaceae, Clostridium, and Bacteroides, after stimulus with monomeric flavan-3-ols from blackcurrant extract [101] or grape seed extract [102]. In this work, also visible was a decrease in Entorabacteriaceae group after 48 h contact with fermented straw extract, indicating that the polyphenols content was inhibiting their growth.
These findings allow us to state that sugarcane straw extract fermented with S. cerevisiae has a potential prebiotic effect, as besides not stimulating the growth of LAB or Bifidobacterium, a significant increase in SCFA content was seen, namely, acetate, propionate, butyrate, and valerate, while it inhibited putrefactive bacteria such as Entorabacteriaceae.
3.7. Influence of the Extract on Cells Viability
The sublethal concentration was defined through the cytotoxicity assay before conducting the Caco-2 inflammatory assay. The Caco-2 cells were used in this investigation since they are pertinent to the examination of the effects of food components on intestinal epithelial tissue. [103].
The extract showed toxicity for concentrations above 3.125 mg/mL (metabolic inhibition above 30%) after 24 h treatment (Figure 6). Thus, this concentration was considered safe and was used in subsequent assays.
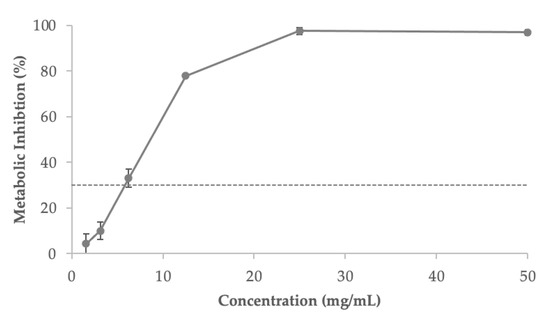
Figure 6.
The metabolic inhibitory action of the fermented sugarcane straw extract on the intestinal Caco-2 cell line, as a base for estimating its cytotoxicity profile. The dotted line shows the 30% cytotoxicity threshold. Values expressed are the mean ± standard deviation.
3.8. Anti-Inflammatory Effect
Interleukin 6 (IL-6) and tumor necrosis factor alpha (TNF-α) are overexpressed in the intestine as a result of the inflammatory bowel disease, which is linked to immune system dysregulation [104]. The regulation of those biomarkers through diet could aid in disease prevention and be used as a co-adjuvant for disease treatment [105].
The possible inflammatory protective impact of sugarcane extract was evaluated using a pre-incubation of the Caco-2 cells media with extracts and a co-incubation (extract + IL-1 pro-inflammatory cytokine). The levels of three cytokines, IL-6, IL-8, and TNF-, were also assessed (Figure 7). The extract (3.125 mg/mL) was employed because it was the greatest dosage deemed physiologically safe for Caco-2 cells. IL-1 was used to provide the inflammatory stimulus. This cytokine is known to have a role in the beginning and amplification of inflammatory activity, which causes Caco-2 cells to produce biomarkers and inflammation mediators. [106].
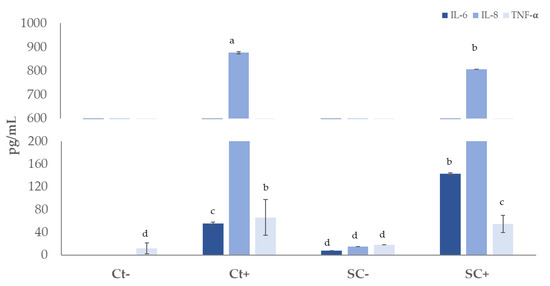
Figure 7.
Immunomodulatory effect on Caco-2 cell line after exposition to fermented sugarcane straw extract (0.625 mg/mL) with (SC+) and without S. cerevisiae (SC−), using IL-1β as a pro-inflammatory stimulus. The concentration of interleukin-8 (IL-8), interleukin-6 (IL-6), and tumor necrosis factor (TNF-) (mean ± SD) were measured. a,b,c,d Different letters indicate significant differences (p < 0.05).
In basal conditions, the presence of interleukins was not detected, and an amount of 11.4 pg/mL of TNF-α was detected. The extract without stimulus was tested to verify if it has a pro-inflammatory effect, and cytokines quantities between 7 and 18 pg/mL were detected. After exposure to stimulus and with 3.125 mg/mL extract, a significant (p < 0.05) decrease in IL-8 was detected from 876 to 806 pg/mL, compared to the positive control (challenge with IL-1β). IL-8 is a cytokine that promotes inflammation and is implicated in chemotaxis. Natural substances having the ability to block the synthesis of IL-8 are appealing because the expression of IL-8 by cancer cells enhances tumor angiogenesis, increases cancer cell proliferation and survival, and facilitates tumor escape from immune cells. Additionally, cancer patients’ poor prognoses are linked to IL-8 expression by cancer cells [107].
The TNF-α reduced from 65.7 to 54.4 pg/mL, corresponding to a decrease of 17%, while IL-6 increased from 55.0 to 142.6 pg/mL when compared with IL-1β stimulus.
TNF-α is produced by macrophages and is responsible to control the host immune response [108]. Meanwhile, IL-6 stimulates the differentiation or proliferation of several non-immune cells while also inducing an immunological response through the formation of effector T cells [109]. In this work, the fermented extract induced the production of IL-6 similarly to a work where a yeast-fermented Korean liquor by-product was tested which was mainly composed by polysaccharide fractions [110]. Monosaccharides showed immune-enhancing effects, and the activation of macrophages was dependent on the polysaccharides molecular weight [111].
Despite the fact that the exact processes by which polyphenols cause their anti-inflammatory effects are not entirely understood, it has been hypothesized that they do so by altering the synthesis of eicosanoids, blocking immune cells that have been activated, or by inhibiting nitric oxide synthase and xyclooxygenase-2 through its inhibitory effects on nuclear factor NF-κβ [112,113].
It was stated that ferulic acid has a strong anti-inflammatory effect, and exerts protection action in the intestinal epithelial barrier dysfunction when stimulated with by LPS [114]. Some studies have discussed the potential of sugarcane polyphenols as anti-inflammatory agents. When Caco-2 cells were treated with IL-1β, a sugarcane stem aqueous extract (50 mol gallic acid equivalent/L) caused a 1.5-fold reduction in IL-8 release [115].
A recent work has described the anti-inflammatory effect of postbiotic extracts produced from four strains, namely, Lactobacillus salivarius, Lactobacillus acidophilus, Lactobacillus plantarum, and Bifidobacterium longum subsp. infantis [116]. Postbiotic extracts from Lactobacillus delbrueckii CIDCA 133 strain reduced the inflammatory markers in a mouse model [117].
4. Conclusions
This work highlights the potential use of a by-product derived from the sugarcane industry, sugarcane straw, to produce postbiotics, through sequential saccharification with cellulase and S. cerevisiae fermentation, with potential application as a food ingredient.
The most prevalent phenolic components in the extract were hydroxycinnamic acids, which were also present in smaller amounts in hydroxybenzoic acids and flavonoids. The extract exhibited capacity to neutralize free radicals while exerting a prebiotic effect, through the increase in short-chain fatty acids, namely, acetate, propionate, and butyrate. Another property identified was the capacity to control pro-inflammatory mediators (IL-8 and TNF-α) in Caco-2 cells, while exerting immune enhancing effect by increasing IL-6. Additionally, the extracts produced in this work, and by this two-step biological process, were shown to modulate gut microbiota especially in terms of production of important SCFA, and to inhibit the growth of potential pathogenic Enterobacteriaceae. However, further studies should be conducted to guarantee efficient and safe usage in humans, as well as other product development strategies, if considering the use of the extract in the improvement of ingredients for functional applications.
As a result, it is possible to see fermented sugarcane straw as a viable source of organic bioactive substances that can be produced within the context of a sustainable and circular economy and potentially used as food ingredients.
Author Contributions
Conceptualization, A.R.M., C.O. and A.L.S.O.; methodology, A.L.S.O., M.S., M.J.C., N.M.d.C., E.M.C. and M.D.; validation, A.R.M. and C.O.; formal analysis, A.L.S.O. and M.S.; investigation, A.L.S.O., M.S., M.J.C., N.M.d.C., E.M.C., S.S. and M.D.; resources, M.P. and A.R.M.; writing—original draft preparation, A.L.S.O. and M.S.; writing—review and editing, A.R.M. and C.O.; supervision, A.R.M.; project administration, M.P. and A.R.M.; funding acquisition, M.P. and A.R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Amyris Bio Products Portugal Unipessoal Lda and Escola Superior de Biotecnologia—Universidade Católica Portuguesa through the Alchemy project, Capturing high value from industrial fermentation bioproducts. This research was funded by Fundo Europeu de Desenvolvimento Regional (FEDER) with a grant number POCI-01−0247-FEDER-027578.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This study did not report any data.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Acetonitrile, 2.2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid), DMSO, formic acid, Folin–Ciocalteu reagent, gallic acid, glucose, yeast extract, magnesium sulfate heptahydrate, peptone, phenol, potassium persulfate, sodium citrate, sulphuric acid, sodium carbonate, tricin, luteolin, protocatechuic acid, vanillic acid, p-coumaric acid, caffeic acid, ferulic acid, chlorogenic acid, 2,5-dihydroxybenzoic acid, 4-hydroxybenzaldehyde, 4-hydroxybenzoic acid, 3,4-dihydroxybenzalhedyde, syringic acid monosodium phosphate, fluorescein disodium salt, Trolox, malic, lactic, acetic, butyric and ethanol, xylose, mannose, galactose, glucose, arabinose fructose, tween 80, vitamin K (Sigma-Aldrich; Sintra, Portugal).
Tri-ammonium citrate (VWR International; Pennsylvania, PA, USA). Potassium phosphate dibasic (Honeywell; Charlotte, NC, USA). Sodium acetate, manganese (II) sulfate tetrahydrate (Merk KGaA; Darmstadt, Germany). Citric acid (AppliChem GmbH). Celluclast (Novozyme, Denmark). Yeast malt broth (YM; Biokar Diagnostics; Allonne, France).
Sodium chloride (AppliChem GmbH), dipotassium hydrogen phosphate, potassium dihydrogen phosphate, magnesium sulfate heptahydrate (Merk KGaA; Darmstadt, Germany), calcium chloride hexahydrate, sodium hydrogen carbonate (Sigma-Aldrich; St. Louis, MO, USA), L-cysteine HCL, bile salts, resazurin solution, fructooligosaccharides (FOS) (Thermo Fisher Scietific; Waltham, MA, USA).
Vitexin, diosmetin, isoschaftoside, orientin, viexin-2-O-rhamnoside (Extrasynthése, France). The 2,2-diphenyl-1-picrylhydrazyl (DPPH˙) was acquired to Alfa Aesar, Thermo Fisher Scientific; Waltham, MA, USA, the AAPH solutions from Acros Organics, Thermo Fisher Scientific; Branchburg, NJ, USA, and Presto Blue (Thermofisher, Waltham, MA, USA).
Caco-2 cells (ATCC HTB-37) were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA).
References
- Sabater, C.; Calvete-Torre, I.; Villamiel, M.; Moreno, F.J.; Margolles, A.; Ruiz, L. Vegetable waste and by-products to feed a healthy gut microbiota: Current evidence, machine learning and computational tools to design novel microbiome-targeted foods. Trends Food Sci. Technol. 2021, 118, 399–417. [Google Scholar] [CrossRef]
- Chaudhari, A.; Dwivedi, M.K. The concept of probiotics, prebiotics, postbiotics, synbiotics, nutribiotics, and pharmabiotics. In Probiotics in the Prevention and Management of Human Diseases; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–11. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A step beyond pre-and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar]
- Duarte, M.; Oliveira, A.L.; Oliveira, C.; Pintado, M.; Amaro, A.; Madureira, A.R. Current postbiotics in the cosmetic market—An update and development opportunities. Appl. Microbiol. Biotechnol. 2022, 106, 5879–5891. [Google Scholar] [CrossRef]
- Amiri, S.; Rezazadeh-Bari, M.; Alizadeh-Khaledabad, M.; Rezaei-Mokarram, R.; Sowti-Khiabani, M. Fermentation optimization for co-production of postbiotics by Bifidobacterium lactis BB12 in cheese whey. Waste Biomass Valorization 2021, 12, 5869–5884. [Google Scholar] [CrossRef]
- Lee, F.H.; Wan, S.Y.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdul Rahim, R.; Idrus, Z. Comparative study of extracellular proteolytic, cellulolytic, and hemicellulolytic enzyme activities and biotransformation of palm kernel cake biomass by lactic acid bacteria isolated from Malaysian foods. Int. J. Mol. Sci. 2019, 20, 4979. [Google Scholar] [CrossRef]
- Lara-Cervantes, T.d.J.; Carrillo-Inungaray, M.L.; Balderas-Hernández, V.E.; Aguilar-Zárate, P.; Veana, F. Valorization of Parmentiera aculeata juice in growth of probiotics in submerged culture and their postbiotic production: A first approach to healthy foods. Arch. Microbiol. 2022, 204, 679. [Google Scholar] [CrossRef]
- Outeiriño, D.; Costa-Trigo, I.; Pinheiro de Souza Oliveira, R.; Pérez Guerra, N.; Salgado, J.M.; Domínguez, J.M. Biorefinery of Brewery Spent Grain by Solid-State Fermentation and Ionic Liquids. Foods 2022, 11, 3711. [Google Scholar] [CrossRef]
- Ali, S.E.; Yuan, Q.; Wang, S.; Farag, M.A. More than sweet: A phytochemical and pharmacological review of sugarcane (Saccharum officinarum L.). Food Biosci. 2021, 44, 101431. [Google Scholar] [CrossRef]
- Saad, M.B.W.; Oliveira, L.R.M.; Cândido, R.G.; Quintana, G.; Rocha, G.J.M.; Gonçalves, A.R. Preliminary studies on fungal treatment of sugarcane straw for organosolv pulping. Enzyme Microb. Technol. 2008, 43, 220–225. [Google Scholar] [CrossRef]
- dos Santos Rocha, M.S.R.; Pratto, B.; de Sousa, R.; Almeida, R.M.R.G.; da Cruz, A.J.G. A kinetic model for hydrothermal pretreatment of sugarcane straw. Bioresour. Technol. 2017, 228, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Szymańska, K.; Kordala, N.; Dąbrowska, A.; Bednarski, W.; Juszczuk, A. Evaluation of Mucor indicus and Saccharomyces cerevisiae capability to ferment hydrolysates of rape straw and Miscanthus giganteus as affected by the pretreatment method. Bioresour. Technol. 2016, 212, 262–270. [Google Scholar] [CrossRef]
- Ávila, P.F.; Martins, M.; de Almeida Costa, F.A.; Goldbeck, R. Xylooligosaccharides production by commercial enzyme mixture from agricultural wastes and their prebiotic and antioxidant potential. Bioact. Carbohydr. Diet. Fibre 2020, 24, 100234. [Google Scholar] [CrossRef]
- Martins, M.; Silva, K.C.G.; Ávila, P.F.; Sato, A.C.K.; Goldbeck, R. Xylo-oligosaccharide microparticles with synbiotic potential obtained from enzymatic hydrolysis of sugarcane straw. Food Res. Int. 2021, 140, 109827. [Google Scholar] [CrossRef]
- Ávila, P.F.; Martins, M.; Goldbeck, R. Enzymatic Production of Xylooligosaccharides from Alkali-Solubilized Arabinoxylan from Sugarcane Straw and Coffee Husk. BioEnergy Res. 2021, 14, 739–751. [Google Scholar] [CrossRef]
- Scott, K.P.; Grimaldi, R.; Cunningham, M.; Sarbini, S.R.; Wijeyesekera, A.; Tang, M.L.K.; Lee, J.; Yau, Y.F.; Ansell, J.; Theis, S. Developments in understanding and applying prebiotics in research and practice—An ISAPP conference paper. J. Appl. Microbiol. 2020, 128, 934–949. [Google Scholar] [CrossRef]
- da Silva, A.S.; Inoue, H.; Endo, T.; Yano, S.; Bon, E.P.S. Milling pretreatment of sugarcane bagasse and straw for enzymatic hydrolysis and ethanol fermentation. Bioresour. Technol. 2010, 101, 7402–7409. [Google Scholar] [CrossRef]
- Ping, L.; Brosse, N.; Sannigrahi, P.; Ragauskas, A. Evaluation of grape stalks as a bioresource. Ind. Crops Prod. 2011, 33, 200–204. [Google Scholar] [CrossRef]
- Costa, E.M.; Pereira, C.F.; Ribeiro, A.A.; Casanova, F.; Freixo, R.; Pintado, M.; Ramos, O.L. Characterization and Evaluation of Commercial Carboxymethyl Cellulose Potential as an Active Ingredient for Cosmetics. Appl. Sci. 2022, 12, 6560. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- de Carvalho, N.M.; Oliveira, D.L.; Costa, C.M.; Pintado, M.; Madureira, A.R. Can Supplemented Skim Milk (SKM) Boost Your Gut Health? Fermentation 2022, 8, 126. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Barros, A.S.; Silva Ferreira, A.C.; Silva, A.M.S. Influence of the temperature and oxygen exposure in red Port wine: A kinetic approach. Food Res. Int. 2015, 75, 337–347. [Google Scholar] [CrossRef]
- de Carvalho, N.M.; Oliveira, D.L.; Saleh, M.A.D.; Pintado, M.; Madureira, A.R. Preservation of Human Gut Microbiota Inoculums for In Vitro Fermentations Studies. Fermentation 2021, 7, 14. [Google Scholar] [CrossRef]
- de Carvalho, N.M.; Walton, G.E.; Poveda, C.G.; Silva, S.N.; Amorim, M.; Madureira, A.R.; Pintado, M.E.; Gibson, G.R.; Jauregi, P. Study of in vitro digestion of Tenebrio molitor flour for evaluation of its impact on the human gut microbiota. J. Funct. Foods 2019, 59, 101–109. [Google Scholar] [CrossRef]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Volatile composition, phenolic content, and in vitro antioxidant activity of red wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar] [CrossRef]
- Contreras, M.D.M.; Hernández-Ledesma, B.; Amigo, L.; Martín-Álvarez, P.J.; Recio, I. Production of antioxidant hydrolyzates from a whey protein concentrate with thermolysin: Optimization by response surface methodology. LWT-Food Sci. Technol. 2011, 44, 9–15. [Google Scholar] [CrossRef]
- ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2022.
- Machado, M.; Costa, E.M.; Silva, S.; Rodriguez-Alcalá, L.M.; Gomes, A.M.; Pintado, M. Pomegranate Oil’s Potential as an Anti-Obesity Ingredient. Molecules 2022, 27, 4958. [Google Scholar] [CrossRef]
- Chi, C.-H.; Cho, S.-J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of insoluble-bound phenolics to antioxidant properties of wheat. J. Agric. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef]
- Daroit, D.J.; Silveira, S.T.; Hertz, P.F.; Brandelli, A. Production of extracellular β-glucosidase by Monascus purpureus on different growth substrates. Process Biochem. 2007, 42, 904–908. [Google Scholar] [CrossRef]
- Wang, L.; Wei, W.; Tian, X.; Shi, K.; Wu, Z. Improving bioactivities of polyphenol extracts from Psidium guajava L. leaves through co-fermentation of Monascus anka GIM 3.592 and Saccharomyces cerevisiae GIM 2.139. Ind. Crops Prod. 2016, 94, 206–215. [Google Scholar] [CrossRef]
- Sreeramulu, G.; Zhu, Y.; Knol, W. Kombucha fermentation and its antimicrobial activity. J. Agric. Food Chem. 2000, 48, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Queiroz Santos, V.A.; Nascimento, C.G.; Schmidt, C.A.P.; Mantovani, D.; Dekker, R.F.H.; da Cunha, M.A.A. Solid-state fermentation of soybean okara: Isoflavones biotransformation, antioxidant activity and enhancement of nutritional quality. LWT 2018, 92, 509–515. [Google Scholar] [CrossRef]
- Parisutham, V.; Chandran, S.-P.; Mukhopadhyay, A.; Lee, S.K.; Keasling, J.D. Intracellular cellobiose metabolism and its applications in lignocellulose-based biorefineries. Bioresour. Technol. 2017, 239, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Duvnjak, Z.; Turcott, G.; Duan, Z.D. Production and consumption of sorbitol and fructose by Saccharomyces cerevisiae ATCC 36859. J. Chem. Technol. Biotechnol. 1991, 52, 527–537. [Google Scholar] [CrossRef]
- Liu, D.; Tang, W.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Monosaccharide composition analysis of polysaccharides from natural sources: Hydrolysis condition and detection method development. Food Hydrocoll. 2021, 116, 106641. [Google Scholar] [CrossRef]
- Martins, A.M.T.B.S.; Cordeiro, C.A.A.; Freire, A.M.J.P. In situ analysis of methylglyoxal metabolism in Saccharomyces cerevisiae. FEBS Lett. 2001, 499, 41–44. [Google Scholar] [CrossRef]
- Abbott, D.A.; Zelle, R.M.; Pronk, J.T.; van Maris, A.J.A. Metabolic engineering of Saccharomyces cerevisiae for production of carboxylic acids: Current status and challenges. FEMS Yeast Res. 2009, 9, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Kalliopi, R.; Paola, D.; Simone, G.; Fabrizio, T.; Rosanna, T.; Sandra, T.; Giovanna, S.; Luca, R.; Luca, C. Candida zemplinina Can Reduce Acetic Acid Produced by Saccharomyces cerevisiae in Sweet Wine Fermentations. Appl. Environ. Microbiol. 2012, 78, 1987–1994. [Google Scholar] [CrossRef]
- Coutinho, I.D.; Baker, J.M.; Ward, J.L.; Beale, M.H.; Creste, S.; Cavalheiro, A.J. Metabolite Profiling of Sugarcane Genotypes and Identification of Flavonoid Glycosides and Phenolic Acids. J. Agric. Food Chem. 2016, 64, 4198–4206. [Google Scholar] [CrossRef] [PubMed]
- Deseo, M.A.; Elkins, A.; Rochfort, S.; Kitchen, B. Antioxidant activity and polyphenol composition of sugarcane molasses extract. Food Chem. 2020, 314, 126180. [Google Scholar] [CrossRef]
- Zheng, R.; Su, S.; Zhou, H.; Yan, H.; Ye, J.; Zhao, Z.; You, L.; Fu, X. Antioxidant/antihyperglycemic activity of phenolics from sugarcane (Saccharum officinarum L.) bagasse and identification by UHPLC-HR-TOFMS. Ind. Crops Prod. 2017, 101, 104–114. [Google Scholar] [CrossRef]
- Sanchiz, A.; Pedrosa, M.M.; Guillamón, E.; Arribas, C.; Cabellos, B.; Linacero, R.; Cuadrado, C. Influence of boiling and autoclave processing on the phenolic content, antioxidant activity and functional properties of pistachio, cashew and chestnut flours. LWT 2019, 105, 250–256. [Google Scholar] [CrossRef]
- Goulas, V.; Papoti, V.T.; Exarchou, V.; Tsimidou, M.Z.; Gerothanassis, I.P. Contribution of Flavonoids to the Overall Radical Scavenging Activity of Olive (Olea europaea L.) Leaf Polar Extracts. J. Agric. Food Chem. 2010, 58, 3303–3308. [Google Scholar] [CrossRef]
- Greeff, J.; Joubert, J.; Malan, S.F.; van Dyk, S. Antioxidant properties of 4-quinolones and structurally related flavones. Bioorg. Med. Chem. 2012, 20, 809–818. [Google Scholar] [CrossRef]
- Campos, L.; Granell, P.; Tárraga, S.; López-Gresa, P.; Conejero, V.; Bellés, J.M.; Rodrigo, I.; Lisón, P. Salicylic acid and gentisic acid induce RNA silencing-related genes and plant resistance to RNA pathogens. Plant Physiol. Biochem. 2014, 77, 35–43. [Google Scholar] [CrossRef]
- Mechchate, H.; Es-safi, I.; Mohamed Al kamaly, O.; Bousta, D. Insight into Gentisic Acid Antidiabetic Potential Using In Vitro and In Silico Approaches. Molecules 2021, 26, 1932. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Elahi, F.; Daliri, E.B.-M.; Han, S.-I.; Oh, D.-H. Impact of thermal treatment and fermentation by lactic acid bacteria on sorghum metabolite changes, their antioxidant and antidiabetic activities. Food Biosci. 2022, 45, 101502. [Google Scholar] [CrossRef]
- Kiyota, E.; Mazzafera, P.; Sawaya, A.C.H.F. Analysis of Soluble Lignin in Sugarcane by Ultrahigh Performance Liquid Chromatography–Tandem Mass Spectrometry with a Do-It-Yourself Oligomer Database. Anal. Chem. 2012, 84, 7015–7020. [Google Scholar] [CrossRef]
- Kilani-Jaziri, S.; Mokdad-Bzeouich, I.; Krifa, M.; Nasr, N.; Ghedira, K.; Chekir-Ghedira, L. Immunomodulatory and cellular anti-oxidant activities of caffeic, ferulic, and p-coumaric phenolic acids: A structure–activity relationship study. Drug Chem. Toxicol. 2017, 40, 416–424. [Google Scholar] [CrossRef]
- Adeboye, P.T.; Bettiga, M.; Aldaeus, F.; Larsson, P.T.; Olsson, L. Catabolism of coniferyl aldehyde, ferulic acid and p-coumaric acid by Saccharomyces cerevisiae yields less toxic products. Microb. Cell Fact. 2015, 14, 149. [Google Scholar] [CrossRef] [PubMed]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-vinyl and 4-ethyl derivatives from hydroxycinnamic acids: Occurrence of volatile phenolic flavour compounds in beer and distribution of Pad1-activity among brewing yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Yazawa, K.; Kurokawa, M.; Obuchi, M.; Li, Y.; Yamada, R.; Sadanari, H.; Matsubara, K.; Watanabe, K.; Koketsu, M.; Tuchida, Y.; et al. Anti-Influenza Virus Activity of Tricin, 4′,5,7-trihydroxy-3′,5′-dimethoxyflavone. Antivir. Chem. Chemother. 2011, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.G.; Gao, S.; Lee, J.K.; Chan, Y.-Y.; Wong, E.C.; Zheng, T.; Li, X.-X.; Shaw, P.-C.; Simmonds, M.S.J.; Lau, C.B. A Natural Flavone Tricin from Grains Can Alleviate Tumor Growth and Lung Metastasis in Colorectal Tumor Mice. Molecules 2020, 25, 3730. [Google Scholar] [CrossRef]
- Oyama, T.; Yasui, Y.; Sugie, S.; Koketsu, M.; Watanabe, K.; Tanaka, T. Dietary tricin suppresses inflammation-related colon carcinogenesis in male Crj: CD-1 mice. Cancer Prev. Res. 2009, 2, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Imm, J.Y. Antiobesity Effect of Tricin, a Methylated Cereal Flavone, in High-Fat-Diet-Induced Obese Mice. J. Agric. Food Chem. 2018, 66, 9989–9994. [Google Scholar] [CrossRef]
- Ji, J.; Yang, X.; Flavel, M.; Shields, Z.P.-I.; Kitchen, B. Antioxidant and Anti-Diabetic Functions of a Polyphenol-Rich Sugarcane Extract. J. Am. Coll. Nutr. 2019, 38, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Lauberte, L.; Telysheva, G.; Cravotto, G.; Andersone, A.; Janceva, S.; Dizhbite, T.; Arshanitsa, A.; Jurkjane, V.; Vevere, L.; Grillo, G.; et al. Lignin–Derived antioxidants as value-added products obtained under cavitation treatments of the wheat straw processing for sugar production. J. Clean. Prod. 2021, 303, 126369. [Google Scholar] [CrossRef]
- Shang, Y.-F.; Cha, K.H.; Lee, E.H.; Pan, C.-H.; Um, B.-H. Optimization, bio accessibility of tricin and anti-oxidative activity of extract from black bamboo leaves. Free Radic. Antioxid. 2016, 6, 64–71. [Google Scholar] [CrossRef]
- Li, X.X.; Chen, S.G.; Yue, G.G.L.; Kwok, H.F.; Lee, J.K.M.; Zheng, T.; Shaw, P.C.; Simmonds, M.S.J.; Lau, C.B.S. Natural flavone tricin exerted anti-inflammatory activity in macrophage via NF-κB pathway and ameliorated acute colitis in mice. Phytomedicine 2021, 90, 153625. [Google Scholar] [CrossRef] [PubMed]
- Shalini, V.; Jayalekshmi, A.; Helen, A. Mechanism of anti-inflammatory effect of tricin, a flavonoid isolated from Njavara rice bran in LPS induced hPBMCs and carrageenan induced rats. Mol. Immunol. 2015, 66, 229–239. [Google Scholar] [CrossRef]
- Solyanik, G.I.; Zulphigarov, O.S.; Prokhorova, I.V.; Pyaskovskaya, O.N.; Kolesnik, D.L.; Atamanyuk, V.P. A Comparative Study on Pharmacokinetics of Tricin, a Flavone from Gramineous Plants with Antiviral Activity. J. Biosci. Med. 2021, 9, 76–91. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Datta, S.; Timson, D.J.; Annapure, U.S. Antioxidant properties and global metabolite screening of the probiotic yeast Saccharomyces cerevisiae var. boulardii. J. Sci. Food Agric. 2017, 97, 3039–3049. [Google Scholar] [CrossRef]
- Li, H.; Huang, J.; Wang, Y.; Wang, X.; Ren, Y.; Yue, T.; Wang, Z.; Gao, Z. Study on the nutritional characteristics and antioxidant activity of dealcoholized sequentially fermented apple juice with Saccharomyces cerevisiae and Lactobacillus plantarum fermentation. Food Chem. 2021, 363, 130351. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, Y.; Chen, L.; Zhao, C.; Maubois, J.; Jiang, T.; Li, H.; He, S. Antioxidant activity of two yeasts and their attenuation effect on 4-nitroquinoline 1-oxide induced in vitro lipid peroxidation. Int. J. Food Sci. Technol. 2010, 45, 555–561. [Google Scholar] [CrossRef]
- Hassan, H.M.M. Antioxidant and immunostimulating activities of yeast (Saccharomyces cerevisiae) autolysates. World Appl. Sci. J. 2011, 15, 1110–1119. [Google Scholar]
- Aditi, S.; Sarbjit, S.K.; Om, P.S. Screening of indigenous yeast isolates obtained from traditional fermented foods of Western Himalayas for probiotic attributes. J. Yeast Fungal Res. 2011, 2, 117–126. [Google Scholar]
- Takalloo, Z.; Nikkhah, M.; Nemati, R.; Jalilian, N.; Sajedi, R.H. Autolysis, plasmolysis and enzymatic hydrolysis of baker’s yeast (Saccharomyces cerevisiae): A comparative study. World J. Microbiol. Biotechnol. 2020, 36, 68. [Google Scholar] [CrossRef]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Osman, A.; El-Gazzar, N.; Almanaa, T.N.; El-Hadary, A.; Sitohy, M. Lipolytic postbiotic from Lactobacillus paracasei manages metabolic syndrome in albino wistar rats. Molecules 2021, 26, 472. [Google Scholar] [CrossRef] [PubMed]
- Anvar, S.A.; Rahimyan, D.; Golestan, L.; Shojaee, A.; Pourahmad, R. Butter fortified with spray-dried encapsulated Ferulago angulata extract nanoemulsion and postbiotic metabolite of Lactiplantibacillus plantarum subsp. plantarum improves its physicochemical, microbiological and sensory properties. Int. J. Dairy Technol. 2023. [Google Scholar] [CrossRef]
- Watson, E.-J.; Giles, J.; Scherer, B.L.; Blatchford, P. Human faecal collection methods demonstrate a bias in microbiome composition by cell wall structure. Sci. Rep. 2019, 9, 16831. [Google Scholar] [CrossRef]
- Oku, T.; Nakamura, S. Fructooligosaccharide: Metabolism through gut microbiota and prebiotic effect. Food Nutr. J. 2017, 2, 1–10. [Google Scholar] [CrossRef]
- Tornero-Martínez, A.; Cruz-Ortiz, R.; Jaramillo-Flores, M.E.; Osorio-Díaz, P.; Ávila-Reyes, S.V.; Alvarado-Jasso, G.M.; Mora-Escobedo, R. In vitro fermentation of polysaccharides from Aloe vera and the evaluation of antioxidant activity and production of short chain fatty acids. Molecules 2019, 24, 3605. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health promoting microbial metabolites produced by gut microbiota after prebiotics metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef]
- Wong, J.M.W.; De Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Hague, A.; Elder, D.J.E.; Hicks, D.J.; Paraskeva, C. Apoptosis in colorectal tumour cells: Induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int. J. Cancer 1995, 60, 400–406. [Google Scholar] [CrossRef] [PubMed]
- McRorie Jr, J.W.; McKeown, N.M. Understanding the physics of functional fibers in the gastrointestinal tract: An evidence-based approach to resolving enduring misconceptions about insoluble and soluble fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Hoyles, L.; Flint, H.J.; Dumas, M.-E. Colonic bacterial metabolites and human health. Curr. Opin. Microbiol. 2013, 16, 246–254. [Google Scholar] [CrossRef]
- Fehlbaum, S.; Prudence, K.; Kieboom, J.; Heerikhuisen, M.; Van den Broek, T.; Schuren, F.H.J.; Steinert, R.E.; Raederstorff, D. In vitro fermentation of selected prebiotics and their effects on the composition and activity of the adult gut microbiota. Int. J. Mol. Sci. 2018, 19, 3097. [Google Scholar] [CrossRef]
- Rodrigues, D.; Walton, G.; Sousa, S.; Rocha-Santos, T.A.P.; Duarte, A.C.; Freitas, A.C.; Gomes, A.M.P. In vitro fermentation and prebiotic potential of selected extracts from seaweeds and mushrooms. LWT 2016, 73, 131–139. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; Van de Wiele, T.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef]
- Vazquez, M.J.; Alonso, J.L.; Domınguez, H.; Parajo, J.C. Xylooligosaccharides: Manufacture and applications. Trends Food Sci. Technol. 2000, 11, 387–393. [Google Scholar] [CrossRef]
- Scott, E.; De Paepe, K.; Van de Wiele, T. Postbiotics and Their Health Modulatory Biomolecules. Biomolecules 2022, 12, 1640. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Duncan, S.H.; Sheridan, P.O.; Walker, A.W.; Flint, H.J. Microbial lactate utilisation and the stability of the gut microbiome. Gut Microbiome 2022, 3, e3. [Google Scholar] [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Karnaouri, A.; Matsakas, L.; Krikigianni, E.; Rova, U.; Christakopoulos, P. Valorization of waste forest biomass toward the production of cello-oligosaccharides with potential prebiotic activity by utilizing customized enzyme cocktails. Biotechnol. Biofuels 2019, 12, 285. [Google Scholar] [CrossRef]
- Ávila, P.F.; Silva, M.F.; Martins, M.; Goldbeck, R. Cello-oligosaccharides production from lignocellulosic biomass and their emerging prebiotic applications. World J. Microbiol. Biotechnol. 2021, 37, 73. [Google Scholar] [CrossRef] [PubMed]
- Gwiazdowska, D.; Juś, K.; Jasnowska-Małecka, J.; Kluczyńska, K. The impact of polyphenols on Bifidobacterium growth. Acta Biochim. Pol. 2015, 62, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Molan, A.-L.; Liu, Z.; Kruger, M. The ability of blackcurrant extracts to positively modulate key markers of gastrointestinal function in rats. World J. Microbiol. Biotechnol. 2010, 26, 1735–1743. [Google Scholar] [CrossRef]
- Viveros, A.; Chamorro, S.; Pizarro, M.; Arija, I.; Centeno, C.; Brenes, A. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 2011, 90, 566–578. [Google Scholar] [CrossRef]
- Nunes, A.R.; Gonçalves, A.C.; Alves, G.; Falcão, A.; Garcia-Viguera, C.; Moreno, D.A.; Silva, L.R. Valorisation of Prunus avium L. By-Products: Phenolic Composition and Effect on Caco-2 Cells Viability. Foods 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G.; Ng, S.C. Understanding and preventing the global increase of inflammatory bowel disease. Gastroenterology 2017, 152, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Edwards, A.; Martín-Pérez, L.; Jiménez-Aspee, F.; Theoduloz, C.; Schmeda-Hirschmann, G.; Larrosa, M. Anti-inflammatory effect of polyphenols from Chilean currants (Ribes magellanicum and R. punctatum) after in vitro gastrointestinal digestion on Caco-2 cells: Anti-inflammatory activity of in vitro digested Chilean currants. J. Funct. Foods 2019, 59, 329–336. [Google Scholar] [CrossRef]
- Martins, I.M.; Macedo, G.A.; Macedo, J.A. Biotransformed grape pomace as a potential source of anti-inflammatory polyphenolics: Effects in Caco-2 cells. Food Biosci. 2020, 35, 100607. [Google Scholar] [CrossRef]
- Prakash, M.D.; Stojanovska, L.; Feehan, J.; Nurgali, K.; Donald, E.L.; Plebanski, M.; Flavel, M.; Kitchen, B.; Apostolopoulos, V. Anti-cancer effects of polyphenol-rich sugarcane extract. PLoS ONE 2021, 16, e0247492. [Google Scholar] [CrossRef]
- Corral, L.G.; Haslett, P.A.J.; Muller, G.W.; Chen, R.; Wong, L.-M.; Ocampo, C.J.; Patterson, R.T.; Stirling, D.I.; Kaplan, G. Differential cytokine modulation and T cell activation by two distinct classes of thalidomide analogues that are potent inhibitors of TNF-α. J. Immunol. 1999, 163, 380–386. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.J.; Kim, T.-W.; Kim, S.; Kim, J.H.; Park, J.-T.; Lee, N.H.; Choi, Y.-S.; Kang, M.-C.; Song, K.-M. Immune-enhancing effects of polysaccharide extract of by-products of Korean liquor fermented by Saccharomyces cerevisiae. Int. J. Biol. Macromol. 2021, 188, 245–252. [Google Scholar] [CrossRef]
- Yin, M.; Zhang, Y.; Li, H. Advances in research on immunoregulation of macrophages by plant polysaccharides. Front. Immunol. 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Alarcon De La Lastra, C.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Chuang, C.-C.; McIntosh, M.K. Potential mechanisms by which polyphenol-rich grapes prevent obesity-mediated inflammation and metabolic diseases. Annu. Rev. Nutr. 2011, 31, 155–176. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Guo, Y.; Zhao, J.; Xu, X.; Wang, N.; Liu, Q. Ferulic Acid Ameliorates Lipopolysaccharide-Induced Barrier Dysfunction via MicroRNA-200c-3p-Mediated Activation of PI3K/AKT Pathway in Caco-2 Cells. Front. Pharmacol. 2020, 11, 376. [Google Scholar] [CrossRef] [PubMed]
- Romier-Crouzet, B.; Van De Walle, J.; During, A.; Joly, A.; Rousseau, C.; Henry, O.; Larondelle, Y.; Schneider, Y.-J. Inhibition of inflammatory mediators by polyphenolic plant extracts in human intestinal Caco-2 cells. Food Chem. Toxicol. 2009, 47, 1221–1230. [Google Scholar] [CrossRef]
- Batista, V.L.; De Jesus, L.C.L.; Tavares, L.M.; Barroso, F.L.A.; Fernandes, L.J.d.S.; Freitas, A.d.S.; Americo, M.F.; Drumond, M.M.; Mancha-Agresti, P.; Ferreira, E. Paraprobiotics and Postbiotics of Lactobacillus delbrueckii CIDCA 133 Mitigate 5-FU-Induced Intestinal Inflammation. Microorganisms 2022, 10, 1418. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).