
Persistent Spatial Patterns of Listeria monocytogenes and Salmonella enterica Concentrations in Surface Waters: Empirical Orthogonal Function Analysis of Data from Maryland

 ,
,  ,
, 
 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Data
2.2. Empirical Orthogonal Function Analysis and Statistical Computations
3. Results and Discussions
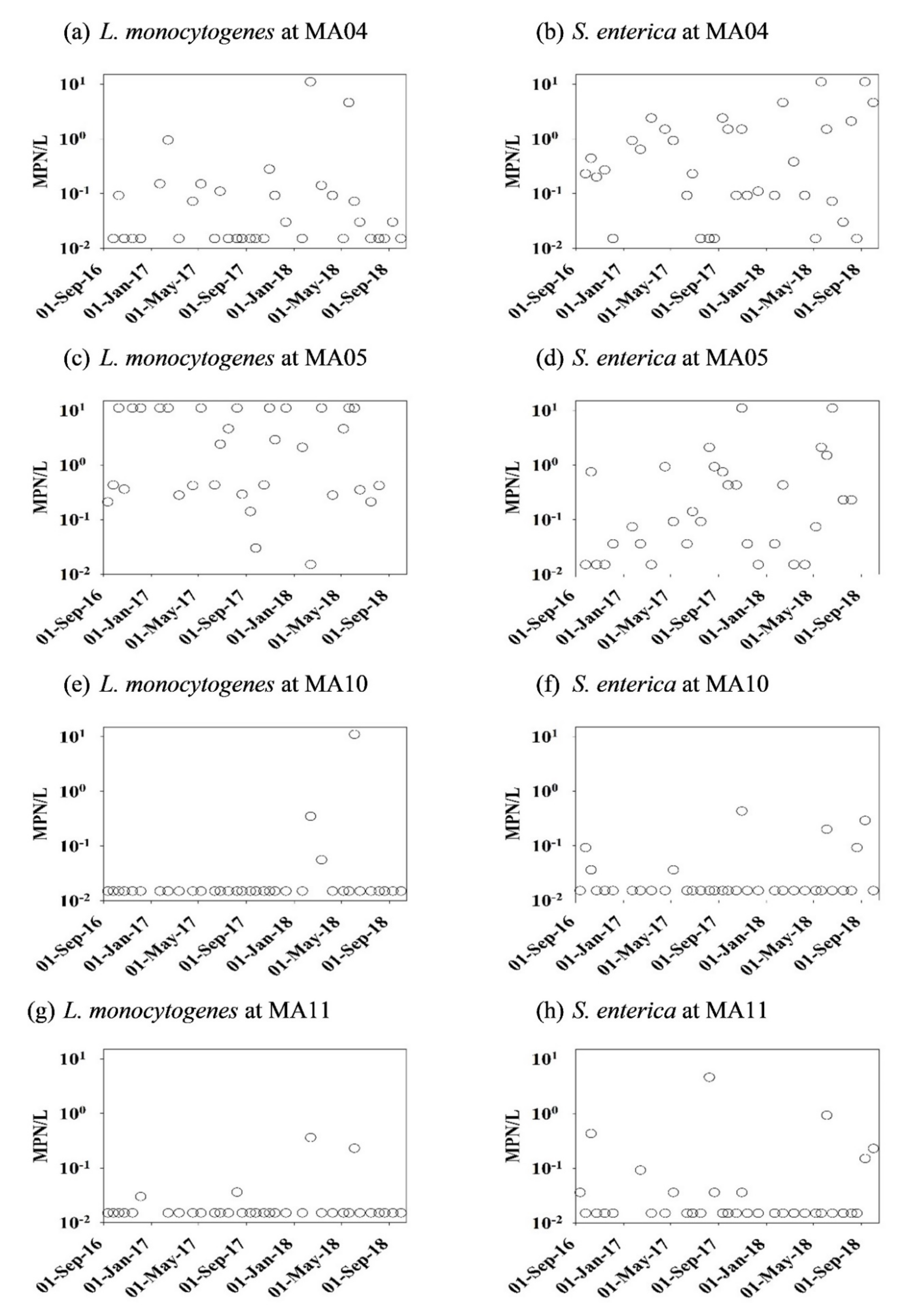
3.1. The Dataset Overview
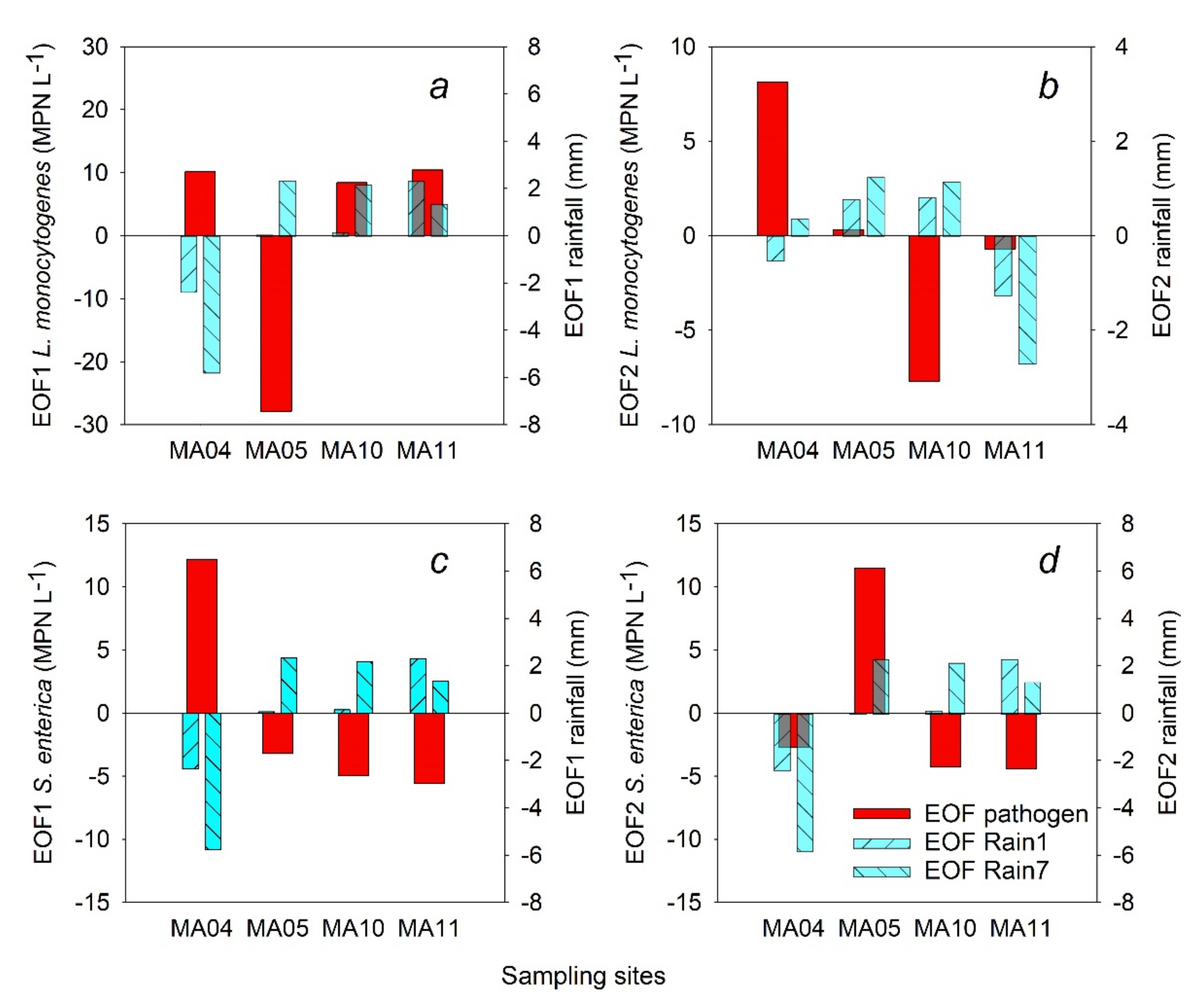
3.2. Empirical Orthogonal Functions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J. Foodborne illness acquired in the USA—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Greene, S.K.; Daly, E.R.; Talbot, E.A.; Demma, L.J.; Holzbauer, S.; Patel, N.J.; Hill, A.; Walderhaug, M.O.; Hoekstra, R.M.; Painter, J.A. Recurrent multistate outbreak of Salmonella Newport associated with tomatoes from contami-nated fields. Epidemiol. Infect. 2008, 136, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Angelo, K.M.; Chu, A.; Anand, M.; Nguyen, T.A.; Bottichio, L.; Wise, M.; Williams, I.; Seelman, S.; Bell, R.; Fatica, M.; et al. Outbreak of Salmonella Newport infections linked to cucumbers—United States. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 144. [Google Scholar]
- Wiedmann, M.; Sauders, B. Ecology of Listeria Species and L. monocytogenes in the Natural Environment. In Food Science and Technology; Marcel Dekker: New York, NY, USA, 2007; Volume 161, p. 21. [Google Scholar]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria Species in Urban and Natural Environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.B.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes Persistence in Food-Associated Environments: Epidemiology, Strain Characteristics, and Implications for Public Health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef]
- Linke, K.; Rückerl, I.; Brugger, K.; Karpiskova, R.; Walland, J.; Muri-Klinger, S.; Tichy, A.; Wagner, M.; Stessl, B. Reservoirs of Listeria Species in Three Environmental Ecosystems. Appl. Environ. Microbiol. 2014, 80, 5583–5592. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, V.S.; Janes, M.E.; King, J.M.; Doerrler, W.; Adhikari, A. Effect of residual chlorine and organic acids on survival and attachment of Escherichia coli O157: H7 and Listeria monocytogenes on spinach leaves during storage. LWT 2019, 105, 298–305. [Google Scholar] [CrossRef]
- McCollum, J.T.; Cronquist, A.B.; Silk, B.J.; Jackson, K.A.; O’Connor, K.A.; Cosgrove, S.; Gossack, J.P.; Parachini, S.S.; Jain, N.S.; Ettestad, P.; et al. Multistate Outbreak of Listeriosis Associated with Cantaloupe. N. Engl. J. Med. 2013, 369, 944–953. [Google Scholar] [CrossRef]
- Vereecken, H.; Pachepsky, Y.; Simmer, C.; Rihani, J.; Kunoth, A.; Korres, W.; Graf, A.; Franssen, H.-H.; Thiele-Eich, I.; Shao, Y. On the role of patterns in understanding the functioning of soil-vegetation-atmosphere systems. J. Hydrol. 2016, 542, 63–86. [Google Scholar] [CrossRef]
- Hannachi, A.; Jolliffe, I.T.; Stephenson, D.B. Empirical orthogonal functions and related techniques in atmos-pheric science: A review. Int. J. Climatol. 2007, 27, 1119–1152. [Google Scholar] [CrossRef]
- Navarra, A.; Simoncini, V. A Guide to Empirical Orthogonal Functions for Climate Data Analysis; Springer Science & Business Media: Berlin/Heilderberg, Germany, 2010. [Google Scholar] [CrossRef]
- Roundy, P.E. On the Interpretation of EOF Analysis of ENSO, Atmospheric Kelvin Waves, and the MJO. J. Clim. 2015, 28, 1148–1165. [Google Scholar] [CrossRef]
- Lian, T.; Chen, D. An Evaluation of Rotated EOF Analysis and Its Application to Tropical Pacific SST Variability. J. Clim. 2012, 25, 5361–5373. [Google Scholar] [CrossRef]
- Yue, W.; Meng, K.; Hou, K.; Zuo, R.; Zhang, B.T.; Wang, G. Evaluating climate and irrigation effects on spati-otemporal variabilities of regional groundwater in an arid area using EOFs. Sci. Total Environ. 2020, 709, 136147. [Google Scholar] [CrossRef]
- Korres, W.; Koyama, C.N.; Fiener, P.; Schneider, K. Analysis of surface soil moisture patterns in agricultural landscapes using Empirical Orthogonal Functions. Hydrol. Earth Syst. Sci. 2010, 14, 751–764. [Google Scholar] [CrossRef]
- Kim, S.; Daughtry, C.; Russ, A.; Pedrera-Parrilla, A.; Pachepsky, Y. Analysis of Spatiotemporal Variability of Corn Yields Using Empirical Orthogonal Functions. Water 2020, 12, 3339. [Google Scholar] [CrossRef]
- Jeon, D.J.; Pachepsky, Y.; Harriger, D.; Picard, R.; Coppock, C.; Wells, E.; Hong, E.-M. Analysis of Escherichia coli and Enterococci Concentrations Patterns in a Pennsylvania Creek Using Empirical Orthogonal Functions. J. Environ. Qual. 2019, 48, 1703–1710. [Google Scholar] [CrossRef]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, Diversity, and Seasonality of Waterborne Salmonellae in a Rural Watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar] [CrossRef]
- Walters, S.P.; Thebo, A.L.; Boehm, A.B. Impact of urbanization and agriculture on the occurrence of bacterial pathogens and stx genes in coastal waterbodies of central California. Water Res. 2011, 45, 1752–1762. [Google Scholar] [CrossRef]
- Strawn, L.K.; Fortes, E.D.; Bihn, E.A.; Nightingale, K.K.; Gröhn, Y.T.; Worobo, R.W.; Wiedmann, M.; Bergholz, P.W. Landscape and Meteorological Factors Affecting Prevalence of Three Food-Borne Pathogens in Fruit and Vegetable Farms. Appl. Environ. Microbiol. 2013, 79, 588–600. [Google Scholar] [CrossRef]
- Harris, C.S.; Tertuliano, M.; Rajeev, S.; Vellidis, G.; Levy, K. Impact of storm runoff on Salmonella and Escherichia coli prevalence in irrigation ponds of fresh produce farms in southern Georgia. J. Appl. Microbiol. 2018, 124, 910–921. [Google Scholar] [CrossRef]
- Lipp, E.K.; Schmidt, N.; Luther, M.E.; Rose, J.B. Determining the Effects of El Nino-Southern Oscillation Events on Coastal Water Quality. Estuaries 2001, 24, 491–497. [Google Scholar] [CrossRef]
- Martinez-Urtaza, J.; Saco, M.; de Novoa, J.; Perez-Piñeiro, P.; Peiteado, J.; Lozano-Leon, A.; Garcia-Martin, O. Influence of environmental factors and human activity on the presence of Salmonella serovars in a marine environment. Appl. Environ. Microbiol. 2004, 70, 2089–2097. [Google Scholar] [CrossRef]
- Crabill, C.; Donald, R.; Snelling, J.; Foust, R.; Southam, G. The impact of sediment fecal coliform reservoirs on seasonal water quality in Oak Creek, Arizona. Water Res. 1999, 33, 2163–2171. [Google Scholar] [CrossRef]
- Pandey, P.; Soupir, M.L.; Wang, Y.; Cao, W.; Biswas, S.; Vaddella, V.; Pasternack, G. Water and sediment microbial quality of mountain and agricultural streams. J. Environ. Qual. 2018, 47, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Callahan, M.T.; Van Kessel, J.A.; Micallef, S.A. Salmonella enterica recovery from river waters of the Maryland Eastern Shore reveals high serotype diversity and some multidrug resistance. Environ. Res. 2019, 168, 7–13. [Google Scholar] [CrossRef]
- Sharma, M.; Handy, E.T.; East, C.L.; Kim, S.; Jiang, C.; Callahan, M.T.; Allard, S.M.; Micallef, C.; Anderson-Coughlin, B.; Gartley, S.; et al. Prevalence of Salmonella and Listeria monocytogenes in non-traditional irrigation waters in the Mid-Atlantic United States is affected by water type, season, and recovery method. PLoS ONE 2020, 5, e0229365. [Google Scholar] [CrossRef]
- Truitt, L.N.; Vazquez, K.M.; Pfuntner, R.C.; Rideout, S.L.; Havelaar, A.H.; Strawn, L.K. Microbial Quality of Agricultural Water Used in Produce Preharvest Production on the Eastern Shore of Virginia. J. Food Prot. 2018, 81, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Haymaker, J.R.; Sharma, M.; Parveen, S.; Hashem, F.; May, E.B.; Handy, E.T.; White, C.; East, C.; Bradshaw, R.; Micallef, S.A.; et al. Prevalence of shi-ga-toxigenic and atypical enteropathogenic Escherichia coli in untreated surface water and reclaimed water in the Mid-Atlantic U.S. Environ. Res. 2019, 172, 630–636. [Google Scholar] [CrossRef]
- Sultana, S.; Theresa, C.M.; Chengsheng, J.; Eric, H.; Cheryl, E.; Joseph, H.; Bui, A.; Craddock, H.; Murray, R.; Kulkurni, P.; et al. A longitudinal assessment of Escherichia coli, total coliforms, Enterococcus and Aeromonas spp. dynamics in alternative irrigation water sources: A CONSERVE study. Appl. Environ. Microbiol. 2020, 86, 1098–5336. [Google Scholar]
- Cropland Data Layer Database, USDA-NASS 2020. Available online: https://nassgeodata.gmu.edu/CropScape/ (accessed on 29 May 2022).
- Glass, G.V.; Hopkins, K.D. Statistical Methods in Education and Psychology, 3rd ed.; Allyn & Bacon: Boston, MA, USA, 1995. [Google Scholar]
- Luo, Z.; Gu, G.; Ginn, A.; Giurcanu, M.C.; Adams, P.; Vellidis, G.; van Bruggen, A.H.C.; Danyluk, M.D.; Wright, A.C. Distribution and Characterization of Salmonella enterica Isolates from Irrigation Ponds in the Southeastern United States. Appl. Environ. Microbiol. 2015, 81, 4376–4387. [Google Scholar] [CrossRef] [PubMed]
- Stea, E.C.; Purdue, L.M.; Jamieson, R.C.; Yost, C.K.; Hansen, L.T. Comparison of the Prevalences and Diversities of Listeria Species and Listeria monocytogenes in an Urban and a Rural Agricultural Watershed. Appl. Environ. Microbiol. 2015, 81, 3812–3822. [Google Scholar] [CrossRef]
- Cooley, M.B.; Quiñones, B.; Oryang, D.; Mandrell, R.E.; Gorski, L. Prevalence of shiga toxin producing Escherichia coli, Salmonella enterica, and Listeria monocytogenes at public access watershed sites in a California Central Coast agricultural region. Front. Cell. Infect. Microbiol. 2014, 4, 30. [Google Scholar] [CrossRef]
- Lu, Z.; Lapen, D.; Scott, A.; Dang, A.; Topp, E. Identifying Host Sources of Fecal Pollution: Diversity of Escherichia coli in Confined Dairy and Swine Production Systems. Appl. Environ. Microbiol. 2005, 71, 5992–5998. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, K.K.; Schukken, Y.H.; Nightingale, C.R.; Fortes, E.D.; Ho, A.J.; Her, Z.; Grohn, Y.T.; McDonough, P.L.; Wiedmann, M. Ecology and Transmission of Listeria monocytogenes Infecting Ruminants and in the Farm Environment. Appl. Environ. Microbiol. 2004, 70, 4458–4467. [Google Scholar] [CrossRef] [PubMed]
- Gorski, L.; Parker, C.T.; Liang, A.; Cooley, M.B.; Jay-Russell, M.T.; Gordus, A.G.; Atwill, E.R.; Mandrell, R.E. Prevalence, Distribution, and Diversity of Salmonella enterica in a Major Produce Region of California. Appl. Environ. Microbiol. 2011, 77, 2734–2748. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, G.; Edge, T.; Gannon, V.; Jokinen, C.; Lyautey, E.; Neumann, N.; Ruecker, N.; Scott, A.; Sunohara, M.; Topp, E.; et al. Associations among pathogenic bacteria, parasites, and environmental and land use factors in multiple mixed-use watersheds. Water Res. 2011, 45, 5807–5825. [Google Scholar] [CrossRef]
- Weindl, L.; Frank, E.; Ullrich, U.; Heurich, M.; Kleta, S.; Ellerbroek, L.; Gareis, M. Listeria monocytogenes in Different Specimens from Healthy Red Deer and Wild Boars. Foodborne Pathog. Dis. 2016, 13, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Lyautey, E.; Lu, Z.; Lapen, D.R.; Wilkes, G.; Scott, A.; Berkers, T.; Topp, E. Distribution and diversity of Escherichia coli populations in the South Nation River drainage basin, eastern Ontario, Canada. Appl. Environ. Microbiol. 2010, 76, 1486–1496. [Google Scholar] [CrossRef]
- Baudart, J.; Grabulos, J.; Barusseau, J.P.; Lebaron, P. Salmonella spp. and fecal coliform loads in coastal waters from a point vs. nonpoint source of pollution. J. Environ. Qual. 2000, 29, 241–250. [Google Scholar] [CrossRef]
- Weller, D.; Shiwakoti, S.; Bergholz, P.; Grohn, Y.; Wiedmann, M.; Strawn, L.K. Validation of a Previously Developed Geospatial Model That Predicts the Prevalence of Listeria monocytogenes in New York State Produce Fields. Appl. Environ. Microbiol. 2016, 82, 797–807. [Google Scholar] [CrossRef]
- Chapin, T.K.; Nightingale, K.K.; Worobo, R.W.; Wiedmann, M.; Strawn, L.K. Geographical and Meteorological Factors Associated with Isolation of Listeria Species in New York State Produce Production and Natural Environments. J. Food Prot. 2014, 77, 1919–1928. [Google Scholar] [CrossRef] [PubMed]
- Deaven, A.M.; Ferreira, C.M.; Reed, E.A.; See, J.R.C.; Lee, N.A.; Almaraz, E.; Rios, P.C.; Marogi, J.G.; Lamendella, R.; Zheng, J.; et al. Salmonella Genomics and Population Analyses Reveal High Inter- and Intraserovar Diversity in Freshwater. Appl. Environ. Microbiol. 2021, 87, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Patchanee, P.; Molla, B.; White, N.; Line, D.E.; Gebreyes, W.A. Tracking Salmonella Contamination in Various Watersheds and Phenotypic and Genotypic Diversity. Foodborne Pathog. Dis. 2010, 7, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Tauxe, R.; Kruse, H.; Hedberg, C.; Potter, M.; Madden, J.; Wachsmuth, K. Microbial Hazards and Emerging Issues Associated with Produce† A Preliminary Report to the National Advisory Committee on Microbiologic Criteria for Foods. J. Food Prot. 1997, 60, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.K.; Bradshaw, R.; Nyarko, E.; Millner, P.D.; Neher, D.; Weicht, T.; Bergholz, T.M.; Sharma, M. Survival and Growth of Wild-Type and rpoS-Deficient Salmonella Newport Strains in Soil Extracts Prepared with Heat-Treated Poultry Pellets. J. Food Prot. 2019, 82, 501–506. [Google Scholar] [CrossRef]
- Litt, P.K.; Kelly, A.; Omar, A.; Johnson, G.; Vinyard, B.T.; Kniel, K.E.; Sharma, M. Temporal and Agricultural Factors Influence Escherichia coli Survival in Soil and Transfer to Cucumbers. Appl. Environ. Microbiol. 2021, 87, 1–14. [Google Scholar] [CrossRef]
- Libby, S.J.; Lesnick, M.; Hasegawa, P.; Kurth, M.; Belcher, C.; Fierer, J.; Guiney, D.G. Characterization of the spv Locus in Salmonella enterica Serovar Arizona. Infect. Immun. 2002, 70, 3290–3294. [Google Scholar] [CrossRef]
- Ferguson, C.; Husman, A.M.D.R.; Altavilla, N.; Deere, D.; Ashbolt, N. Fate and Transport of Surface Water Pathogens in Watersheds. Crit. Rev. Environ. Sci. Technol. 2003, 33, 299–361. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pathogen | Season | Site | |||
|---|---|---|---|---|---|
| MA04 | MA05 | MA10 | MA11 | ||
| L. monocytogenes | Non-growing | 0.76 ± 0.15 1 (58.8%) 2 (10/17) 3 | 5.92 ± 0.32 (93.8%) (15/16) | 0.03 ± 0.00 (11.1%) (2/18) | 0.03 ± 0.00 (12.5%) (2/16) |
| L. monocytogenes | Growing | 0.30 ± 0.05 (35.3%) (6/17) | 3.63 ± 0.27 (100%) (16/16) | 0.62 ± 0.14 (5.6%) (1/18) | 0.02 ± 0.00 (11.1%) (2/18) |
| S. enterica | Non-growing | 1.05 ± 0.08 (94.1%) (16/17) | 0.86 ± 0.16 (62.5%) (10/16) | 0.04 ± 0.00 (11.8%) (2/17) | 0.06 ± 0.00 (25.0%) (4/16) |
| S. enterica | Growing | 1.83 ± 0.20 (70.6%) (12/17) | 1.31 ± 0.16 (93.3%) (14/15) | 0.05 ± 0.00 (27.8%) (5/18) | 0.33 ± 0.05 (33.3%) (6/18) |
| Correlated variables | MA04 (Stream) | MA05 (Stream) | MA10 (Pond) | MA11 (Pond) | ||||
|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| Rain1 vs. L. monocytogenes | 0.003 1 | 0.987 | −0.634 | 0.003 | 0.589 | 0.001 | 0.725 | <0.001 |
| Rain1 vs. S. enterica | 0.707 | <0.001 | 0.31 | 0.115 | 0.371 | 0.056 | 0.14 | 0.485 |
| Rain7 vs. L. monocytogenes | 0.085 | 0.67 | −0.562 | 0.002 | 0.447 | 0.019 | 0.334 | 0.087 |
| Rain7 vs. S. enterica | 0.107 | 0.593 | 0.448 | 0.019 | 0.208 | 0.296 | 0.108 | 0.591 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Paul, M.; Negahban-Azar, M.; Micallef, S.A.; Rosenberg Goldstein, R.E.; Hashem, F.; Parveen, S.; Sapkota, A.; Kniel, K.; Sapkota, A.R.; et al. Persistent Spatial Patterns of Listeria monocytogenes and Salmonella enterica Concentrations in Surface Waters: Empirical Orthogonal Function Analysis of Data from Maryland. Appl. Sci. 2022, 12, 7526. https://doi.org/10.3390/app12157526
Kim S, Paul M, Negahban-Azar M, Micallef SA, Rosenberg Goldstein RE, Hashem F, Parveen S, Sapkota A, Kniel K, Sapkota AR, et al. Persistent Spatial Patterns of Listeria monocytogenes and Salmonella enterica Concentrations in Surface Waters: Empirical Orthogonal Function Analysis of Data from Maryland. Applied Sciences. 2022; 12(15):7526. https://doi.org/10.3390/app12157526
Chicago/Turabian StyleKim, Seongyun, Manashi Paul, Masoud Negahban-Azar, Shirley A. Micallef, Rachel E. Rosenberg Goldstein, Fawzy Hashem, Salina Parveen, Amir Sapkota, Kalmia Kniel, Amy R. Sapkota, and et al. 2022. "Persistent Spatial Patterns of Listeria monocytogenes and Salmonella enterica Concentrations in Surface Waters: Empirical Orthogonal Function Analysis of Data from Maryland" Applied Sciences 12, no. 15: 7526. https://doi.org/10.3390/app12157526
APA StyleKim, S., Paul, M., Negahban-Azar, M., Micallef, S. A., Rosenberg Goldstein, R. E., Hashem, F., Parveen, S., Sapkota, A., Kniel, K., Sapkota, A. R., Pachepsky, Y., & Sharma, M. (2022). Persistent Spatial Patterns of Listeria monocytogenes and Salmonella enterica Concentrations in Surface Waters: Empirical Orthogonal Function Analysis of Data from Maryland. Applied Sciences, 12(15), 7526. https://doi.org/10.3390/app12157526