Abstract
Nanoparticles are components of many vaccines, helping to make them more stable and immunogenic. They protect antigens—or the genetic material encoding them—from degradation, target them to particular tissues or cells, promote their uptake into antigen-presenting cells, and activate the immune response (in the form of adjuvants). Nanoparticles come in many different kinds, some with uniform composition and some with elaborate core-and-shell structures, including lipid membranes. The antigen is usually retained inside, and the surface can be functionalized by targeting or activating proteins and carbohydrates. This minireview provides a general introductory overview to vaccination and a survey of nanoparticles, their types, production, characteristics, and individual applications in vaccines, and finally, a brief look into the world of artificial antigen-presenting cells.
1. Vaccination
In the current pandemic caused by the novel coronavirus, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) (Abbreviations: aAPC, artificial APC; ABC, amphiphilic block copolymer; APC, antigen-presenting cell; AuNP, gold nanoparticle; BCR, B cell receptor; CAR, chimeric antigen receptor; CTL, cytotoxic T lymphocyte; DCs, dendritic cells; HA, hemagglutinin; HBV, hepatitis B virus; HIV, human immunodeficiency virus; HPV, human papillomavirus; ISCOM, immunostimulating complex; LPN, liposome-polycation nanoparticle; MHC, major histocompatibility complex; MSN, mesoporous silica nanoparticles; NFAT, nuclear factor of activated T cells; NP, nanoparticle; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; SiNP, silica nanoparticles; SVP, subviral particle; TCR, T cell receptor; Treg, regulatory (inhibitory) T cell; VLP, virus-like particles; VSP, variant-specific surface protein; ZIKV, ZIKA virus), the need for the development of effective vaccines has again come to the forefront of public attention [1]. Of course, vaccines against infectious diseases or cancer have been a predominant medical concern throughout history and will remain so in the future [2]. This review focuses on the contribution of nanoparticle technology to vaccine development in recent years, providing introductions into both fields for the non-specialized reader. For deeper information, the reader is referred to the considerable body of detailed recent reviews [3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19].
The mammalian immune system includes stationary cells such as macrophages and dendritic cells (DCs) in the skin and the mucosal tissues of the airways and intestines that act as guards wherever pathogens (mainly bacteria, viruses, or fungi) can enter the body [3,20]. Should this happen, the macrophages trigger an inflammation, which constitutes a so-called ‘innate’ (rapid but low-specificity) immune response (Figure 1). Such an inflammatory reaction usually requires whole (entire) pathogens, whose typical surface patterns of cell wall molecules are recognized by the innate receptors of macrophages. These then endocytose the pathogens or attack them with reactive oxygen species and other substances, simultaneously secreting cytokines to attract more immune cells. At the same time, DCs also become activated through innate receptors, take up the pathogens by receptor-mediated or fluid-phase endocytosis, and travel to lymph nodes to present processed protein antigens, i.e., fragments of the pathogen proteins, to the T cells of the adaptive immune system. Activated T helper cells then activate macrophages (to kill endocytosed pathogens) or B cells (to secrete antibodies), and activated cytotoxic T lymphocytes (CTL) kill virus-infected or tumor cells. The generation of activated effector B and T cells is called the adaptive immune response [20].
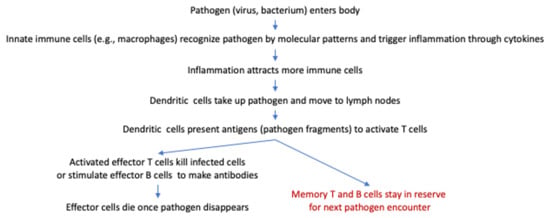
Figure 1.
The course of an antiviral or antibacterial immune response is simplified. The pathogen enters the body and is recognized by the innate immune cells through its specific molecular patterns, the cytokines then triggering an inflammation. This inflammation attracts more immune cells, with the pathogen being taken up by dendritic cells and moved to the lymph nodes. The dendritic cells then present antigens that activate effector T cells and memory T cells. The effector T cells either kill the infected cells or stimulate effector B cells to make antibodies, with the effector cells dying after the pathogen is eliminated. In contrast, memory T and B cells remain in reserve to be used in the future encounter of the same pathogen.
Antibodies are soluble proteins that are produced by activated B cells (the humoral adaptive immune response), and they bind to epitopes (molecular regions) on antigens, which can be proteins, nucleic acids, or lipids of the pathogen [20]. Therefore, antibodies are especially suitable for detecting pathogens in the blood, the extracellular space, and the interior (mucosal) surfaces of the body, such as the lungs or the intestines [20,21]. The binding of antibodies to their epitopes on the surface of pathogens leads to their inactivation, cell death, or phagocytosis by immune cells [20]. Antibodies can only survey the external surface of a pathogen or an infected cell, but not its interior [22].
Activated cytotoxic T lymphocytes (CTL), in contrast, recognize peptide fragments of intracellular proteins presented to them by MHC (major histocompatibility complex) class I proteins (the cellular adaptive immune response). Such MHC class I epitope peptides can originate from any protein that is synthesized in the cell, including nuclear or cytoplasmic proteins, and so CTL are especially suitable for detecting pathogen-infected or tumor cells [4]. Recognition by a CTL leads to the death of the presenting cell by induced apoptosis, which limits the spread of the virus or the propagation of the tumor. CTL responses against tumor cells are in principle similar to those against virus-infected cells, but inefficient initiation of the response (in the absence of inflammation) and the immunosuppressive microenvironment of the tumor (which employs suppressive signaling or mechanical barriers) imposes additional barriers.
After clearing the pathogens or tumor cells, T and B cells undergo programmed cell death, leaving the immune system with a small number of antigen-specific memory cells that can be rapidly reactivated and expanded if the same pathogen is encountered again. The purpose of a vaccine is, therefore, to present the antigens to the adaptive immune system such that it will generate memory B and T cells that warrant a rapid immune response [3,20,23]. For this, the vaccine formulation must contain the right antigen; for example, if the purpose of the vaccine is to generate antibodies to the surface proteins of a virus, then these surface proteins, or their most exposed parts that are most likely to be the epitope of antibodies, are the antigenic epitopes to be used in the vaccine. These antigens (or indeed their genetic information, as in a nucleic acid vaccine) must be contained in a vaccine in a physical form that is effectively taken up, processed, and presented by the DCs [24,25]. The vaccine should also contain an adjuvant that causes mild inflammation in order to activate the DCs in the first place; such adjuvants are mineral salts, saponins, emulsions, polymers, cytokines, liposomes, or nanoparticles (see below) [4,26]. Sometimes, the antigen itself can function as an adjuvant [3,27]. The vaccine may be formulated to ensure a slow and sustained release of the antigen since a strong adaptive response might take days to weeks to develop [27].
Many successful vaccines have been designed, especially against pathogens that cause acute infections such as tetanus, polio, smallpox, and measles [3,25]; in contrast, there are no effective vaccines yet for diseases that cause chronic infections such as HIV [25]. Another focus of development is therapeutic vaccines for cancer or autoimmune diseases and allergies [25]. In the past, the development of vaccines that contained the actual pathogen often took many years; for therapy or to rapidly deal with newly emerging pathogens, faster development cycles and universally usable technical platforms are highly desirable, and with synthetic vaccine formulations, these aims can be achieved, as the many vaccines against SARS-CoV-2 demonstrate [1]; in this endeavor, artificial nanoparticles play a central role.
2. Forms of Antigen in a Vaccine
In principle, the best immune response is elicited by a vaccine that is as similar as possible to the actual pathogen and that mimics its route and mechanism of infection [5]. Thus, many successful vaccines against viruses and bacteria consist of inactivated (killed) or attenuated (chemically or genetically disabled) pathogens [5,15]. Such formulations have drawbacks, though: their development (e.g., by a repeated passage in non-human host organisms or cell lines) might take many years, and their production can be difficult and time-consuming [5,28].
For these reasons, vaccines have been developed that present the most important epitopes, or their genetic information, to the immune system by other means. Subunit vaccines use purified antigen molecules such as proteins or bacterial cell wall polysaccharides [29], though the latter can only induce antibodies but not CTL [30]. Nucleotide vaccines do not contain actual antigen molecules but instead RNA or DNA that encodes them, leaving it to the cells in the vicinity of the injected vaccine to produce the antigenic protein; macrophages and/or dendritic cells then process the antigen and initiate an immune response [1].
Since in an infection, the immune system usually encounters whole pathogens, i.e., particles that may contain many copies of the antigens on its surface or in its interior [17], the innate immune response is naturally optimized for particulate antigens. Compared to soluble antigens, they are phagocytosed better by the immune cells and give stronger responses. Particulate antigens such as whole bacteria more strongly activate the intracellular signaling pathways of the innate immune cells, such as the inflammasome and the calcineurin/NFAT pathway, resulting in better cytokine production and immune stimulation. Starting the humoral adaptive immune response, in turn, requires endocytosis of antigen by B cells, which is more efficient with particulate antigens since the crosslinking (induced proximity) of several B cell receptor molecules by a multivalent particulate antigen yields stronger B cell activation with better antigen presentation to T helper cells [17].
Pathogens have used such strong immune reactions to particulate antigens for their own purpose, namely to subvert the immune response directed against them. For example, hepatitis B virus (HBV) triggers infected cells to produce virus-like entities called subviral particles (SVPs) of about 25 nm diameter (compared to 42 nm for the HBV virion) that display the viral surface antigen (sAg) but contain no viral genome. SVPs vastly outnumber the real virions, and being highly immunogenic, they deflect the immune response, enabling chronic HBV infection [31]. Because of their strong immunogenicity, these SVPs have indeed been used as carriers for vaccine epitopes from other pathogens [7] as a kind of natural vaccine nanoparticle. The HBsAg SVPs are glycosylated lipoprotein particles stabilized by a network of intramolecular and intermolecular disulfide bonds, and they allow targeted modifications of both the glycan content and the level of disulfide bonding. HBsAg SVPs accept and present foreign antigenic sequences to yield a chimeric particulate delivery platform that led to the development of the Mosquirix vaccine against malaria and other potential preclinical vaccines against infectious diseases [7].
In the development of vaccines with all-synthetic components, it makes sense to use synthetic nanoparticles (NPs) to emulate the strong immunogenicity of polyvalent bacteria, viruses, and SVPs [3,5,6,8,16,21,32,33]. In principle, synthetic NPs might contain the antigen on the inside or on the outside, and they might be modified with additional molecules at their surface; thus, they might be constructed to deliver peptides or proteins to specific tissues and generate location-specific immune responses [3,23,32,34]; to enter innate immune cells through fluid-phase endocytosis [35]; to protect antigens from enzymatic degradation, which is particularly important in case of mucosal vaccination [18,21,35]; to carry several components in one particle, such as both the antigen and the adjuvant; to be made biodegradable, biocompatible (i.e., not producing an adverse reaction in the living organism), and easy to produce [3,5,6,8,16,21,23,26,32,34,35]. In prophylactic and therapeutic studies, synthetic NPs are now used as immunostimulant adjuvants in order to activate or enhance immunity or as delivery systems to enhance antigen processing [32]. Through their influence on innate and adaptive immune responses, NPs might also be designed to boost the production of memory T and B cells, which is vital for a vaccine to be successful (see above) [35].
In principle, therapeutic nanoparticles can be used both in the prophylactic and in the therapeutic context. While therapeutic nanoparticle vaccines are mainly used in cancer treatments and more recently also for the treatment of other diseases such as hypertension or Alzheimer’s, prophylactic nanoparticle vaccines have been applied to the prevention of various infections such as human papillomavirus (HPV) infections [7,35,36,37].
3. Types and Uses of Nanoparticles in Vaccines
The nanoparticles currently used in vaccine development can be loosely grouped into six categories (Table 1, Figure 2) [5,35].

Table 1.
Summary of the six types of nanoparticles used in vaccine development, their main characteristics and sizes.
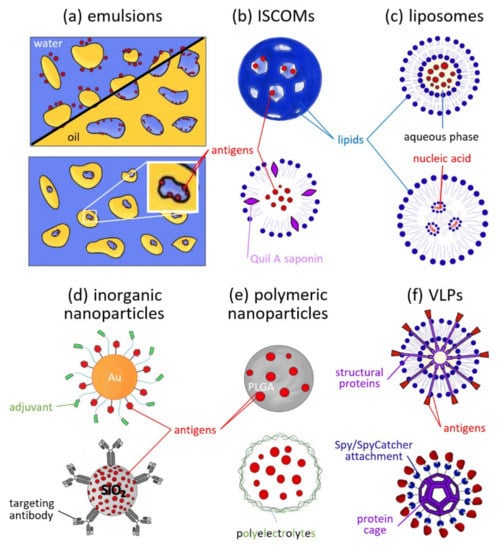
Figure 2.
Types of nanoparticles used in vaccines. The images are schematic and do not depict the size relationships of the particles to their components. (a) Oil-in-water (top left), water-in-oil (top right), and water-in-oil-in-water emulsion (bottom, with reverse micelle enlarged). The antigen is usually adsorbed to the phase interface that is stabilized by the emulsifier (not shown). (b) Three-dimensional view (top) and cross-section schematic (bottom) of an ISCOM. (c) Liposome with protein antigens in the aqueous center (top) and with nucleic acids complexed to cationic lipids inside (bottom). (d) Gold nanoparticles decorated with antigen and adjuvants (top) and mesoporous silica nanoparticle functionalized with antibodies for targeting (bottom). (e) Isotropic PLGA nanoparticle (top) and layer-by-layer polyelectrolyte microcapsule (bottom). (f) Virus-like particles with (top) and without (bottom) enveloping membrane.
3.1. Emulsions
Emulsions are the simplest two-phase structures used in vaccine delivery. Though they are entirely made of liquids, they share some properties with solid particles. In addition to the aqueous phase, they usually consist of a hydrophobic oil phase and an amphiphilic emulsifier detergent that stabilizes the phase boundary, for example, the fatty acid ester of sugar alcohol. There are oil-in-water, water-in-oil, and water-in-oil-in-water emulsions, with droplet sizes between 50 and 600 nm [11,19,24]. The antigens are usually adsorbed to the phase boundary, and from their slow and sustained release into the tissue, emulsions provide a strong adjuvant effect. [19]. Examples comprise MF59TM, an oil-in-water emulsion that has been used as an adjuvant and studied for its potential uses in influenza vaccines [38], and MontanideTM, a family of oil-in-water and water-in-oil emulsions that have been used in malaria vaccines [39]. Oil-in-water nanoemulsions can be complemented with biosurfactant peptides and proteins that can serve as anchors for functionalization with other biomolecules, such that polyethylene glycol and/or a receptor-specific antibody (for targeting) can be selectively attached to the droplet–water interface [40].
3.2. ISCOMs
Immunostimulating complexes (ISCOMs) are spherical, perforated, cage-like, hollow micelles with a size of 40–60 nm that are made of the saponin adjuvant Quil-A®, phospholipids, cholesterol, and the protein or peptide antigens, which are trapped in the center through nonpolar interactions [5,19,24] [32]. ISCOMS can bind and penetrate cellular membranes and carry the antigen and the adjuvant into the cytosol of the antigen-presenting cells (APCs) such as DCs so that the antigen is transported—with activated DC migration—to the lymphnodes [19,24]. ISCOMs are produced with the help of non-ionic detergents that are then removed by dialysis [19]. The antigens that are currently used with ISCOMs are derived from the Newcastle disease virus, herpes simplex virus, and influenza, among others [24].
3.3. Liposomes
Liposomes for vaccination purposes are prepared from biodegradable and nontoxic phospholipids with a reverse-phase evaporation process by dissolving phospholipids in an organic solvent, adding water and antigen, evaporating the solvent, and breaking up the resulting lipid aggregates into liposomes of defined size by sonication or by passage through filters, or both. Alternatively, they can be formed by dissolving lipids in a detergent and then removing the detergent by absorption (e.g., onto hydrophobic beads) or dialysis (if the critical micelle concentration is sufficiently high) [11,19,24]. Liposomes can encapsulate hydrophilic antigens into their aqueous core, and their membranes can incorporate viral envelope glycoproteins to form virosomes [9,19,24]. Inflexal® V for influenza [41] and Epaxal® for hepatitis A [42] are such virosome systems that are approved for human use.
For the delivery of nucleic acid vaccines by liposomes, polycations such as polyethylenimine or cationic lipids can, on the one hand, help to condense the nucleic acid inside the respective liposome and, on the other hand, disrupt the cellular endosomal membranes, leading to more efficient delivery [9,19,24]. Such polycations can be combined with a calcium phosphate core that may contain the nucleic acid as a co-precipitate [43]. As they function as messenger RNA (mRNA) protectors and carriers, such cation-containing liposomes have been used in the development of SARS-CoV-2 vaccines by BioNTech/Pfizer and Moderna companies [31].
As a synthetic alternative to phospholipids, liposomes from amphiphilic block copolymers (ABCs) were studied for the delivery of a DNA vaccine against the Zika virus (ZIKV). Therefore, plasmid DNA encoding a ZIKV antigen was enveloped with tetrafunctional ABCs made of four amphiphilic diblocks that were centered on an ethylenediamine moiety. Delivery was effective in a variety of cell and tissue types such as heart, lung, and muscle [27].
3.4. Inorganic Nanoparticles
Inorganic nanoparticles that are made, for example, from gold, silica, or carbon, were used in the development of vaccines due to their rigid structure and tightly controllable synthesis [11,19,24]. Since they are not biodegradable, though, they might cause inflammation and even fibrosis in some organs such as the lower lungs [11,17]. Inorganic NPs are usually isotropic (internally unstructured); they can be produced by first using a reducing agent to form small particles of the respective material, followed by using silver acetate and cetyltrimethylammonium bromide to bring them into the desired size and shape [19].
Gold nanoparticles (AuNPs) can be easily manufactured in different shapes (cubic, spherical, etc.) with sizes between 2 nm and 150 nm, and thus, they are used in vaccine delivery with gold nanorods conjugating the protein antigen to the surface [44,45]. They can be modified at their surface with multivalent carbohydrates that interact with lectins (carbohydrate-binding proteins of the cell surfaces), with a potential for tissue-specific targeting [30]. Various types of AuNPs were used both to carry antigens derived from viruses, such as the influenza virus, and DNA of the human immunodeficiency virus (HIV) [19,24].
Carbon nanoparticles were studied for vaccine delivery because they are biocompatible and can be produced in different shapes, e.g., nanotubes and mesoporous spheres. Carbon nanotubes lead to intense IgG responses since they can carry several copies of protein and peptide antigens [24].
Nanoparticles based on silica (SiO2, SiNPs) are biocompatible and water-soluble, with long-lasting blood circulation and vaccine delivery, which make them one of the most encouraging inorganic materials used in nanovaccines and delivery system design [26,46]. They can be mesoporous and even hollow, with very high antigen loading capacity. The surface silanol groups allow functionalization with antibodies or other proteins for cell recognition, absorption of additional biomolecules, and strengthened cellular uptake [47,48]. SiNPs can be fabricated with a luminescent core and a paramagnetic coat, allowing for multimodal imaging by microscopy and magnetic resonance [11,49].
3.5. Polymeric Nanoparticles
Polymeric NPs are made from synthetic polymers, with poly(lactide-co-glycolide) (PLG) and poly(lactic-co-glycolic acid) (PLGA) studied the most because of their good biocompatibility and biodegradability [4,32,35,50]. They are isotropic, i.e., without a shell and with the antigen distributed evenly throughout the particle. One advantage of polymeric NPs is that they afford control over the release kinetics of the loaded materials by altering the polymer such as changing its functional chemical groups, monomer ratio, or molecular weight [32,35,51].
Polymeric NPs are often synthesized by the double emulsion solvent evaporation technique: the polymer is dissolved in an organic solvent such as ethyl acetate, and a protein antigen is added together with an emulsifying agent such as polyvinyl alcohol. Then, by vortexing, a water-in-oil-in-water emulsion is formed, and the polymer precipitates around the antigen. The product is dried by solvent evaporation to prevent polymer degradation caused by ester hydrolysis [32]. Another synthesis method uses amphiphilic copolymers that self-assemble through hydrophobic interactions [32,51], enclosing the antigen. Polymeric NPs enable antigen delivery to immune cells and sustained antigen release through their slow biodegradation [32,35]. In recent years, PLGA has been utilized to carry antigens derived from diverse pathogens such as Plasmodium vivax (using the mono-phosphoryl lipid A as the adjuvant), hepatitis B virus (HBV), and experimental model antigens such as chicken ovalbumin and tetanus toxoid [4,32,35,50]. Co-encapsulation of a lipid that stimulates invariant natural killer T cells (iNKT cells) further enhanced T and B cell responses to PLGA nanoparticles [52].
The amphiphilic poly(γ-glutamic acid) (γ-PGA) also self-assembles into nano-micelles with a hydrophilic outer shell and a hydrophobic core [53,54]. The γ-PGA NPs are mostly used to encapsulate hydrophobic antigens [32]. Polymeric NPs that are made of naturally polymeric polysaccharides (such as chitosan, pullulan, inulin, or alginate) can incorporate antigens and can also themselves act as adjuvants [4,8,26,32,35]. Such polysaccharide-based NPs are produced by covalent crosslinking, ionic crosslinking, polyelectrolyte complexes, or by self-assembly of hydrophobically modified polysaccharides [8,32]. Chitosan-based NPs were especially well investigated due to their biocompatibility, biodegradability, their ability to be easily adapted according to preferred variable size (20–600 nm), and their nontoxic nature [4,5,26,32,35]. They were used in the development of several vaccines against HBV [7,32,35]. In vaccines against both HBV and influenza, Advax, an NP adjuvant made from inulin (an activator of complement through the alternative pathway), was used [5].
Polyelectrolytes, i.e., polymers with repeating units that carry either positive (polycations) or negative (polyanions) charges, are another important subclass of polymeric NPs in practical use [55,56]. While they are mainly used as carriers in the delivery of bioactive substances, they have recently been studied as components of peptide-based subunit vaccines against group A streptococcus [55,56]. They can be made water-soluble, biocompatible, and slowly biodegradable [56]. Polyelectrolyte complex nanoparticles (PECNs) are produced using techniques such as ionic gelation (crosslinking) and coacervation (phase separation) [56]. Some approaches use hyaluronic acid and chondroitin sulfate, but the most commonly used natural polymer is chitosan, as it is mucoadhesive, has enhanced tissue permeation, and enables the production of NPs under mild conditions [56]. Chitosan, a hydrolysate of fungal or insect chitin, bears 0.5 positive charges per saccharide unit at physiological pH and can be complexed with hyaluronic acid to form PECNs [56]. Hyaluronic acid naturally occurs in the extracellular matrix of connective tissues and interacts with cell-surface receptors such as CD44; together with chitosan, it increases mucoadhesion [56]. Polyelectrolyte capsules can also be manufactured larger than 1 µM when assembled in a core-shell format with a layer-by-layer buildup technique [57], with the active substances either inside the core or integrated into the shell, with the additional option of modifying the surface with receptors that target particular cells or tissues [58,59]. Such core-shell microcapsules might also be used in vaccine development.
3.6. VLPs and Self-Assembled Protein Cages
Virus-like particles (VLPs) are capsid-like NPs (20–200 nm) made up of viral proteins that can self-assemble into regular structures. They lack a viral genome and thus cannot replicate [4,10,13,32,35,60,61]. When they are produced in eukaryotic cells (e.g., mammalian or insect cell lines, plant [62,63], or yeast cells), VLPs can have an envelope membrane through which the antigens protrude as transmembrane proteins, making them—from the outside—similar to liposomes. Because of their highly repetitive structure, VLPs sometimes generate powerful immune responses even without adjuvant [26,64]. The VLP vaccine for HBV (Engerix) was the first virus-like particle vaccine to reach the wider market, followed by prophylactic VLP vaccines for the human papillomavirus (Cervarix) and malaria (Mosquirix) [9,65,66]; influenza VLP vaccines are in clinical trials [67].
The surfaces of VLPs, or the extra-particular parts of their transmembrane proteins, can be modified to attach other proteins for targeting or for adjuvant functions. This is achieved, for example, by genetic engineering [68], the Spy/SpyCatcher reaction [69], or by chemical crosslinking [70]. One such approach uses genetic fusions of the transmembrane and cytosolic portions of the vesicular stomatitis virus G protein (which makes the VLPs) with variant-specific surface proteins from protozoa (e.g., Giardia), which act as very strong mucosa-specific adjuvants [60].
Self-assembling cage-like protein particles similar to VLPs were also designed de novo. In one study, a trimeric aldolase was engineered with RosettaDesign to fit an icosahedral symmetry with novel interfaces between the trimers optimized for close packing without steric clashes [36]. This self-assembling cage, produced in E. coli, was then functionalized with the recombinant receptor-binding domain of the SARS-CoV-2 Spike protein by the Spy/SpyCatcher reaction for eventual use as a VLP vaccine, using additional adjuvant [71,72].
On the small side of self-assembling protein complexes, ferritin is an icositetramer (24-mer) with an almost spherical structure of 10 nm diameter. In the development of an influenza vaccine, a fusion protein of ferritin and hemagglutinin (HA) from the virus is therefore spontaneously assembled into an octahedral particle with eight trimeric HA spikes. This yielded a stronger immune response than the trivalent inactivated influenza vaccine by itself [35].
4. Nanoparticles as Artificial Antigen-Presenting Cells—The Future of Immunotherapy?
Traditionally, vaccines have aimed to bring about an immune response by stimulating DCs to present the provided antigen to T cells (see above). With a better understanding of the rules of T cell activation, it has recently become feasible to directly stimulate T cells using multivalent reagents that mimic activated dendritic cells, so-called artificial antigen-presenting cells (aAPCs). Because T cells only recognize processed antigen as peptide epitopes bound to an MHC molecule, the use of aAPCs is indicated where peptide-specific T cell stimulation is possible and desirable, for example, with T cells directed against a known tumor epitope. The aAPCs must carry at their surface the MHC molecule/peptide complexes that are needed to convey antigen specificity, and they must also display co-stimulatory molecules, especially ligands for the CD28 receptor on the T cells; they may also carry proteins or lipids that promote general cell adhesion, and they may contain activating cytokines for slow release [73,74]. Inorganic and polymeric nanoparticles of different sizes are being investigated for direct T cell stimulation. In addition, filaments such as dextran molecules or chemical analogs of peptides with a longer biological lifetime promise polyvalent backbones (though not strictly nanoparticles) [75]. The resulting possibilities for direct antigen-specific stimulation of individual T cell clones allow much more finely tuned manipulation of the desired immune response with boundless opportunities, including perhaps, in the future, the inactivation of regulatory (inhibitory) T cells (Tregs) or perhaps the generation of desirable Tregs to treat autoimmune diseases [76].
5. Conclusions
The development of novel vaccines is essential for public health, not just because of the current and potential future pandemics but also because of the increasing understanding of the immune response to cancer, which is beginning to enable prophylactic and therapeutic immunization. Modular vaccine platforms (i.e., that do not need to be completely redesigned for each application) will allow faster development than previously possible, some degree of standardization of their production, and they will perhaps also simplify the process of approval by regulatory authorities. Nanoparticles can, in principle, be such modular platforms.
Among the nanoparticles described above, synthetic nanoparticles, which are assembled from known ingredients, are better candidates for such standardization in development and production (as compared to natural particles such as HBSAg SVPs). They allow fine-tuning with respect to targeting, durability to degradation, the amount of antigen, and the degree of its protection, with the particle either containing the antigen on the inside or the outside of it and the potential of adding diverse molecules to its surface.
The recent impressive success of nucleic acid vaccines that are enveloped in liposomes has made it clear that such vaccines have an additional advantage with respect to standardization: their antigenic content, here as genetic information, can be easily and rapidly altered, with all other components of the vaccine (and even the size of the nucleic acid) remaining the same. This provides convincing answers to newly developing variants of a pathogenic virus and novel pathogens, but it may also be an essential condition of future cancer vaccines, which, because of the highly individual nature of tumor epitopes, may have to be personalized. For this, liposome-encapsulated nucleic acid vaccines may indeed be the only financially viable option.
Such liposomes may be functionalized with adjuvants to enhance their immunogenicity, e.g., in mucosal membranes, which might then enable their use as oral vaccines. This functionalization might occur with the help of transmembrane proteins, blurring the boundary between liposomes and VLPs. Initial approaches for the packaging and delivery of RNA with the help of VLP vesicles exist [77].
Development and optimal design of a standardized toolbox of nanoparticles for vaccine delivery still face several challenges, including our understanding of the physical properties of the different types of nanoparticles (and their ingredients), their biodistribution, biodegradability, and targeting. Therefore, the study and development of reproducible nanoparticles with advantageous properties continues to be of great importance, as well as discovering new administration, targeting, and delivery methods that will enable more efficient use of vaccine nanoparticles in the treatment of disease.
Author Contributions
Writing—original draft preparation, A.P.; writing—review and editing, S.S.; funding acquisition, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Nouria Jantz-Naeem for drawing Figure 2.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Beeraka, N.M.; Tulimilli, S.V.; Karnik, M.; Sadhu, S.P.; Pragada, R.R.; Aliev, G.; Madhunapantula, S.V. The Current Status and Challenges in the Development of Vaccines and Drugs against Severe Acute Respiratory Syndrome-Corona Virus-2 (SARS-CoV-2). BioMed Res. Int. 2021, 2021, 8160860. [Google Scholar] [CrossRef] [PubMed]
- Brisse, M.; Vrba, S.M.; Kirk, N.; Liang, Y.; Ly, H. Emerging Concepts and Technologies in Vaccine Development. Front. Immunol. 2020, 11, 2578. [Google Scholar] [CrossRef]
- Irvine, D.J.; Hanson, M.C.; Rakhra, K.; Tokatlian, T. Synthetic Nanoparticles for Vaccines and Immunotherapy. Chem. Rev. 2015, 115, 11109–11146. [Google Scholar] [CrossRef] [PubMed]
- Sulczewski, F.B.; Liszbinski, R.B.; Romão, R.T.; Rodrigues, L.C. Junior Nanoparticle vaccines against viral infections. Arch. Virol. 2018, 163, 2313–2325. [Google Scholar] [CrossRef]
- Gomes, A.C.; Mohsen, M.; Bachmann, M.F. Harnessing Nanoparticles for Immunomodulation and Vaccines. Vaccines 2017, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Gao, D.; Shen, J.; Wang, Q. A Review of Mesoporous Silica Nanoparticle Delivery Systems in Chemo-Based Combination Cancer Therapies. Front. Chem. 2020, 8, 1086. [Google Scholar] [CrossRef]
- Ho, J.K.-T.; Jeevan-Raj, B.; Netter, H.-J. Hepatitis B Virus (HBV) Subviral Particles as Protective Vaccines and Vaccine Platforms. Viruses 2020, 12, 126. [Google Scholar] [CrossRef]
- Liu, Z.; Jiao, Y.; Wang, Y.; Zhou, C.; Zhang, Z. Polysaccharides-based nanoparticles as drug delivery systems. Adv. Drug Deliv. Rev. 2008, 60, 1650–1662. [Google Scholar] [CrossRef]
- Cutts, F.T.; Franceschi, S.; Goldie, S.; Castellsague, X.D.; De Sanjose, S.; Garnett, G.; Edmunds, W.J.; Claeys, P.; Goldenthal, K.L.; Harperi, D.M.; et al. Human papillomavirus and HPV vaccines: A review. Bull. World Health Organ. 2007, 85, 719–726. [Google Scholar] [CrossRef]
- Donaldson, B.; Lateef, Z.; Walker, G.F.; Young, S.L.; Ward, V.K. Virus-like particle vaccines: Immunology and formulation for clinical translation. Expert Rev. Vaccines 2018, 17, 833–849. [Google Scholar] [CrossRef]
- Zheng, X.; Zeng, S.; Hu, J.; Wu, L.; Hou, X. Applications of silica-based nanoparticles for multimodal bioimaging. Appl. Spectrosc. Rev. 2018, 53, 377–394. [Google Scholar] [CrossRef]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef]
- López-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. J. 2016, 14, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Souza, G.; Rocha, R.; Gonçalves, R.; Ferreira, C.; Silva, B.; Castro, R.; Rodrigues, J.; Júnior, J.; Malaquias, L.; Abrahão, J.; et al. Nanoparticles as Vaccines to Prevent Arbovirus Infection: A Long Road Ahead. Pathogens 2021, 10, 36. [Google Scholar] [CrossRef]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of Action of Adjuvants. Front. Immunol. 2013, 4, 114. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Liu, X.; Zhang, L.; Liu, C.; Liu, G. Cell membrane-encapsulated nanoparticles for vaccines and immunotherapy. Particuology 2021, in press. [Google Scholar] [CrossRef]
- Snapper, C.M. Distinct Immunologic Properties of Soluble Versus Particulate Antigens. Front. Immunol. 2018, 9, 598. [Google Scholar] [CrossRef]
- Li, M.; Kaminskas, L.M.; Marasini, N. Recent advances in nano/microparticle-based oral vaccines. J. Pharm. Investig. 2021, 51, 425–438. [Google Scholar] [CrossRef]
- Petkar, K.; Patil, S.; Chavhan, S.; Kaneko, K.; Sawant, K.; Kunda, N.; Saleem, I. An Overview of Nanocarrier-Based Adjuvants for Vaccine Delivery. Pharmaceutics 2021, 13, 455. [Google Scholar] [CrossRef]
- Medina, K.L. Chapter 4—Overview of the immune system. In Handbook of Clinical Neurology; Pittock, S.J., Vincent, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 133, pp. 61–76. [Google Scholar] [CrossRef]
- Ghobadi, N.; Ghobadi, S.; Burkholder, M.B.; Habibipour, R. Nanoparticles development for pulmonary vaccination: Challenges and opportunities. Nanomed. J. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Zhang, C.; Ötjengerdes, R.M.; Roewe, J.; Mejias, R.; Marschall, A.L.J. Applying Antibodies Inside Cells: Principles and Recent Advances in Neurobiology, Virology and Oncology. Biodrugs 2020, 34, 435. [Google Scholar] [CrossRef] [PubMed]
- Kelly, H.G.; Kent, S.J.; Wheatley, A.K. Immunological basis for enhanced immunity of nanoparticle vaccines. Expert Rev. Vaccines 2019, 18, 269–280. [Google Scholar] [CrossRef]
- Saroja, C.; Lakshmi, P.K.; Bhaskaran, S. Recent trends in vaccine delivery systems: A review. Int. J. Pharm. Investig. 2011, 1, 64–74. [Google Scholar] [CrossRef]
- Vaillant, A.A.J.; Grella, M.J. Vaccine (Vaccination); StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: http://www.ncbi.nlm.nih.gov/books/NBK532895/ (accessed on 13 August 2021).
- Alfagih, I.M.; Aldosari, B.; AlQuadeib, B.; Almurshedi, A.; Alfagih, M.M. Nanoparticles as Adjuvants and Nanodelivery Systems for mRNA-Based Vaccines. Pharmaceutics 2020, 13, 45. [Google Scholar] [CrossRef]
- Clem, A.S. Fundamentals of Vaccine Immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, B. The contribution of vaccination to global health: Past, present and future. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130433. [Google Scholar] [CrossRef]
- Jarząb, A.; Skowicki, M.; Witkowska, D. Subunit vaccines—Antigens, carriers, conjugation methods and the role of adjuvants. Postepy Hig. Med. Dosw. 2013, 67, 1128–1143. [Google Scholar] [CrossRef] [PubMed]
- Khan, S. Conjugate vaccines and polysaccharide response. CMAJ Can. Med. Assoc. J. 2006, 174, 976–977. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ganem, D.; Prince, A.M. Hepatitis B virus infection—Natural history and clinical consequences. N. Engl. J. Med. 2004, 350, 1118–1129. [Google Scholar] [CrossRef]
- Gregory, A.E.; Williamson, D.; Titball, R. Vaccine delivery using nanoparticles. Front. Cell. Infect. Microbiol. 2013, 3, 13. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, X.; Lou, P.; Hu, Z.; Qu, P.; Li, D.; Li, Q.; Xu, Y.; Niu, J.; He, Y. A Nanoparticle-Based Hepatitis C Virus Vaccine With Enhanced Potency. J. Infect. Dis. 2020, 221, 1304–1314. [Google Scholar] [CrossRef]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle Vaccines Against Infectious Diseases. Front. Immunol. 2018, 9, 2224. [Google Scholar] [CrossRef]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.-X.; Mitter, N.; Yu, C.; Middelberg, A.P. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef]
- Briquez, P.S.; Hauert, S.; De Titta, A.; Gray, L.T.; Alpar, A.T.; Swartz, M.A.; Hubbell, J.A. Engineering Targeting Materials for Therapeutic Cancer Vaccines. Front. Bioeng. Biotechnol. 2020, 8, 19. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, S.; Wang, X.-Y.; Zhu, G. Nanovaccines for cancer immunotherapy. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1559. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; Ott, G.S.; Nest, G.V.; Rappuoli, R.; Giudice, G.D. The history of MF59® adjuvant: A phoenix that arose from the ashes. Expert Rev. Vaccines 2013, 12, 13–30. [Google Scholar] [CrossRef]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide ISA 720 and 51: A new generation of water in oil emulsions as adjuvants for human vaccines. Expert Rev. Vaccines 2002, 1, 111–118. [Google Scholar] [CrossRef]
- Zeng, B.J.; Chuan, Y.P.; O’Sullivan, B.; Caminschi, I.; Lahoud, M.H.; Thomas, R.; Middelberg, A.P.J. Receptor-specific delivery of protein antigen to dendritic cells by a nanoemulsion formed using top-down non-covalent click self-assembly. Small Weinh. Bergstr. Ger. 2013, 9, 3736–3742. [Google Scholar] [CrossRef]
- Mischler, R.; Metcalfe, I.C. Inflexal® V a trivalent virosome subunit influenza vaccine: Production. Vaccine 2002, 20, B17–B23. [Google Scholar] [CrossRef]
- Epaxal: A Virosomal Vaccine to Prevent Hepatitis A Infection—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/18844588/ (accessed on 20 September 2021).
- Zhang, J.; Sun, X.; Shao, R.; Liang, W.; Gao, J.; Chen, J. Polycation liposomes combined with calcium phosphate nanoparticles as a non-viral carrier for siRNA delivery. J. Drug Deliv. Sci. Technol. 2015, 30, 1–6. [Google Scholar] [CrossRef]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T.; et al. Gold nanoparticles as a vaccine platform: Influence of size and shape on immunological responses in vitro and in vivo. ACS Nano 2013, 7, 3926–3938. [Google Scholar] [CrossRef]
- Stone, J.W.; Thornburg, N.J.; Blum, D.L.; Kuhn, S.J.; Wright, D.W.; Crowe, J.E. Gold nanorod vaccine for respiratory syncytial virus. Nanotechnology 2013, 24, 295102. [Google Scholar] [CrossRef]
- An, M.; Li, M.; Xi, J.; Liu, H. Silica Nanoparticle as a Lymph Node Targeting Platform for Vaccine Delivery. ACS Appl. Mater. Interfaces 2017, 9, 23466–23475. [Google Scholar] [CrossRef]
- Ow, H.; Larson, D.R.; Srivastava, M.; Baird, B.A.; Webb, W.W.; Wiesner, U. Bright and stable core-shell fluorescent silica nanoparticles. Nano Lett. 2005, 5, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Niut, Y.; Popatt, A.; Yu, M.; Karmakar, S.; Gu, W.; Yu, C. Recent advances in the rational design of silica-based nanoparticles for gene therapy. Ther. Deliv. 2012, 3, 1217–1237. [Google Scholar] [PubMed]
- Morato, Y.L.; Paredes, K.O.; Chamizo, L.L.; Marciello, M.; Filice, M. Recent Advances in Multimodal Molecular Imaging of Cancer Mediated by Hybrid Magnetic Nanoparticles. Polymers 2021, 13, 2989. [Google Scholar] [CrossRef] [PubMed]
- PLGA-Particle Vaccine Carrying TLR3/RIG-I Ligand Riboxxim Synergizes with Immune Checkpoint Blockade for Effective Anti-Cancer Immunotherapy|Nature Communications. Available online: https://www.nature.com/articles/s41467-021-23244-3 (accessed on 13 August 2021).
- Hraber, P.; Bradfute, S.; Clarke, E.; Ye, C.; Pitard, B. Amphiphilic block copolymer delivery of a DNA vaccine against Zika virus. Vaccine 2018, 36, 6911–6917. [Google Scholar] [CrossRef]
- Dölen, Y.; Gileadi, U.; Chen, J.-L.; Valente, M.; Creemers, J.H.A.; Van Dinther, E.A.W.; van Riessen, N.K.; Jäger, E.; Hruby, M.; Cerundolo, V.; et al. PLGA Nanoparticles Co-encapsulating NY-ESO-1 Peptides and IMM60 Induce Robust CD8 and CD4 T Cell and B Cell Responses. Front. Immunol. 2021, 12, 641703. [Google Scholar] [CrossRef]
- Trimaille, T.; Verrier, B. Micelle-Based Adjuvants for Subunit Vaccine Delivery. Vaccines 2015, 3, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, A.; Phelip, C.; Coolen, A.-L.; Monge, C.; Boisgard, A.-S.; Paul, S.; Verrier, B. Biodegradable Polymeric Nanoparticles-Based Vaccine Adjuvants for Lymph Nodes Targeting. Vaccines 2016, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jin, W.; Cruz, J.G.; Marasini, N.; Khalil, Z.G.; Capon, R.J.; Hussein, W.M.; Skwarczynski, M.; Toth, I. Development of Polyelectrolyte Complexes for the Delivery of Peptide-Based Subunit Vaccines against Group A Streptococcus. Nanomaterials 2020, 10, 823. [Google Scholar] [CrossRef] [PubMed]
- Taber, L.; Umerska, A. Polyelectrolyte complexes as nanoparticulate drug delivery systems. Eur. Pharm. Rev. 2015, 20, 36–40. [Google Scholar]
- Sukhorukov, G.B.; Rogach, A.L.; Garstka, M.A.; Springer, S.; Parak, W.J.; Muñoz-Javier, A.; Kreft, O.; Skirtach, A.; Susha, A.S.; Ramaye, Y.; et al. Multifunctionalized polymer microcapsules: Novel tools for biological and pharmacological applications. Small Weinh. Bergstr. Ger. 2007, 3, 944–955. [Google Scholar] [CrossRef]
- Timin, A.S.; Gould, D.J.; Sukhorukov, G.B. Multi-layer microcapsules: Fresh insights and new applications. Expert Opin. Drug Deliv. 2017, 14, 583–587. [Google Scholar] [CrossRef]
- Protein A Functionalized Polyelectrolyte Microcapsules as a Universal Platform for Enhanced Targeting of Cell Surface Receptors|ACS Applied Materials & Interfaces. Available online: https://pubs.acs.org/doi/10.1021/acsami.7b01313 (accessed on 20 September 2021).
- Serradell, M.C.; Rupil, L.L.; Martino, R.A.; Prucca, C.G.; Carranza, P.G.; Saura, A.; Fernández, E.A.; Gargantini, P.R.; Tenaglia, A.H.; Petiti, J.P.; et al. Efficient oral vaccination by bioengineering virus-like particles with protozoan surface proteins. Nat. Commun. 2019, 10, 361. [Google Scholar] [CrossRef]
- Chu, K.-B.; Quan, F.-S. Virus-Like Particle Vaccines Against Respiratory Viruses and Protozoan Parasites. Curr. Top. Microbiol. Immunol. 2021, 433, 77–106. [Google Scholar] [CrossRef]
- Huang, X.; Wang, X.; Zhang, J.; Xia, N.; Zhao, Q. Escherichia coli-derived virus-like particles in vaccine development. NPJ Vaccines 2017, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Santi, L.; Huang, Z.; Mason, H. Virus like particles production in green plants. Methods 2006, 40, 66–76. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.F.; Zhou, J.; Chen, S.; Cai, L.L.; Bao, Q.Y.; Zheng, F.Y.; Lu, J.Q.; Padmanabha, J.; Hengst, K.; Malcolm, K.; et al. HPV6b virus like particles are potent immunogens without adjuvant in man. Vaccine 2000, 18, 1051–1058. [Google Scholar] [CrossRef]
- RTS,S Clinical Trials Partnership. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar] [CrossRef]
- André, F.E. Overview of a 5-year clinical experience with a yeast-derived hepatitis B vaccine. Vaccine 1990, 8, S74–S78. [Google Scholar] [CrossRef]
- Landry, N.; Ward, B.J.; Trepanier, S.; Montomoli, E.; Dargis, M.; Lapini, G.; Vézina, L.-P. Preclinical and Clinical Development of Plant-Made Virus-Like Particle Vaccine against Avian H5N1 Influenza. PLoS ONE 2010, 5, e15559. [Google Scholar] [CrossRef]
- Wetzel, D.; Rolf, T.; Suckow, M.; Kranz, A.; Barbian, A.; Chan, J.-A.; Leitsch, J.; Weniger, M.; Jenzelewski, V.; Kouskousis, B.; et al. Establishment of a yeast-based VLP platform for antigen presentation. Microb. Cell Factories 2018, 17, 17. [Google Scholar] [CrossRef]
- Thrane, S.; Janitzek, C.M.; Matondo, S.; Resende, M.; Gustavsson, T.; De Jongh, W.A.; Clemmensen, S.; Roeffen, W.; Van De Vegte-Bolmer, M.; Van Gemert, G.J.; et al. Bacterial superglue enables easy development of efficient virus-like particle based vaccines. J. Nanobiotechnol. 2016, 14, 30. [Google Scholar] [CrossRef]
- Jegerlehner, A.; Tissot, A.; Lechner, F.; Sebbel, P.; Erdmann, I.; Kündig, T.; Bächi, T.; Storni, T.; Jennings, G.; Pumpens, P.; et al. A molecular assembly system that renders antigens of choice highly repetitive for induction of protective B cell responses. Vaccine 2002, 20, 3104–3112. [Google Scholar] [CrossRef]
- Tan, T.K.; Rijal, P.; Rahikainen, R.; Keeble, A.H.; Schimanski, L.; Hussain, S.; Harvey, R.; Hayes, J.W.P.; Edwards, J.C.; McLean, R.K.; et al. A COVID-19 vaccine candidate using SpyCatcher multimerization of the SARS-CoV-2 spike protein receptor-binding domain induces potent neutralising antibody responses. Nat. Commun. 2021, 12, 542. [Google Scholar] [CrossRef]
- Hsia, Y.; Bale, J.; Gonen, S.; Shi, D.; Sheffler, W.; Fong, K.K.; Nattermann, U.; Xu, C.; Huang, P.-S.; Ravichandran, R.; et al. Design of a hyperstable 60-subunit protein icosahedron. Nature 2016, 535, 136–139. [Google Scholar] [CrossRef]
- Neal, L.R.; Bailey, S.; Wyatt, M.; Bowers, J.; Majchrzak, K.; Nelson, M.H.; Haupt, C.; Paulos, C.M.; Varela, J.C. The Basics of Artificial Antigen Presenting Cells in T Cell-Based Cancer Immunotherapies. J. Immunol. Res. Ther. 2017, 2, 68–79. [Google Scholar]
- Eggermont, L.J.; Paulis, L.E.; Tel, J.; Figdor, C.G. Towards efficient cancer immunotherapy: Advances in developing artificial antigen-presenting cells. Trends Biotechnol. 2014, 32, 456–465. [Google Scholar] [CrossRef]
- Hickey, J.W.; Kosmides, A.K.; Schneck, J. Chapter Six—Engineering Platforms for T Cell Modulation. In International Review of Cell and Molecular Biology; Galluzzi, L., Rudqvist, N.-P., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 341, pp. 277–362. [Google Scholar] [CrossRef]
- Horwitz, D.A.; Bickerton, S.; La Cava, A. Strategies to Use Nanoparticles to Generate CD4 and CD8 Regulatory T Cells for the Treatment of SLE and Other Autoimmune Diseases. Front. Immunol. 2021, 12, 681062. [Google Scholar] [CrossRef]
- Segel, M.; Lash, B.; Song, J.; Ladha, A.; Liu, C.C.; Jin, X.; Mekhedov, S.L.; Macrae, R.K.; Koonin, E.V.; Zhang, F. Mammalian retrovirus-like protein PEG10 packages its own mRNA and can be pseudotyped for mRNA delivery. Science 2021, 373, 882–889. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).