

Effects of Exposure to Urban Atmospheric Particulate Matter Suspended in Seawater on the Mussel Mytilus galloprovincialis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of PM Stock Solution
2.2. Biological Model
2.3. Exposure Assays
2.4. Total Protein Determination
2.5. Antioxidant Enzyme Biomarkers
2.5.1. Catalase (CAT)
2.5.2. Superoxide Dismutase (SOD)
2.5.3. Glutathione Peroxidase (GPx)
2.5.4. Glutathione S-Transferase (GST)
2.6. Lipid Peroxidation (LPO)
2.7. Ubiquitin (Ubi) Assay
2.8. Statistical Analysis
2.9. Ethics
3. Results
3.1. Mortality
3.2. Enzymatic Biomarkers
3.2.1. Catalase (CAT)
3.2.2. Superoxide Dismutase (SOD)
3.2.3. Glutathione Peroxidase (GPx)
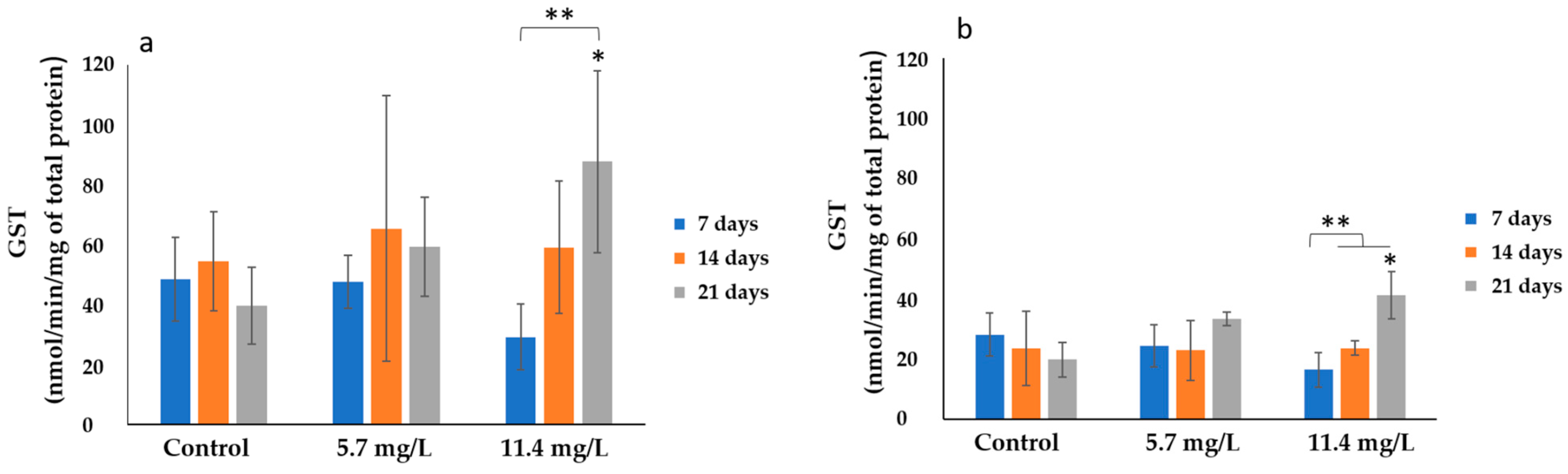
3.2.4. Glutathione-S-Transferase (GST)
3.3. Lipid Peroxidation (LPO)
3.4. Total Ubiquitin
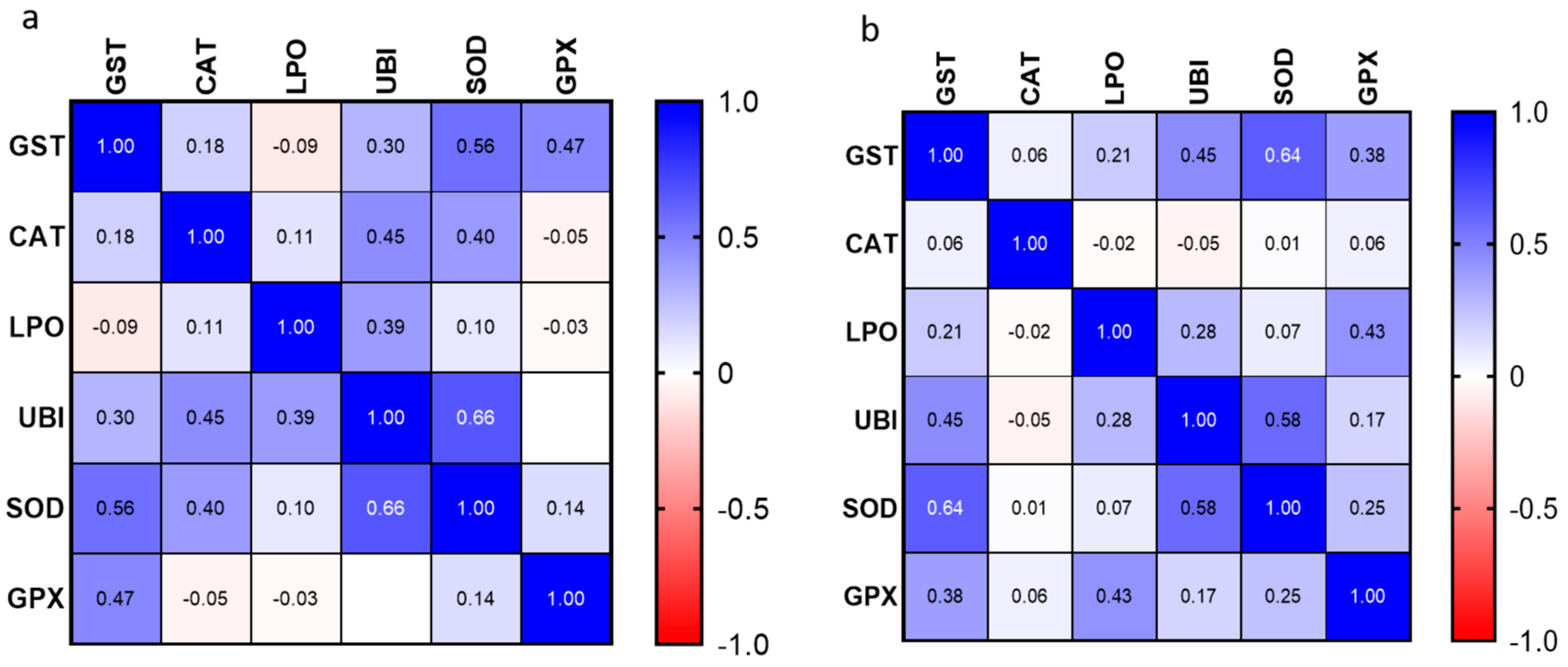
3.5. Correlation Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buis, A. Getting to the Heart of the Particulate Matter. Earth Science Communications Team at NASA’s Jet Propulsion Laboratory|California Institute of Technology. Available online: https://climate.nasa.gov/news/3027/getting-to-the-heart-of-the-particulate-matter/ (accessed on 1 July 2022).
- He, M.; Sun, Y.L.X.; Yang, Z. Distribution patterns of nitrobenzenes and polychlorinated biphenyls in water, suspended particulate matter and sediment from mid- and downstream of the Yellow River (China). Chemosphere 2006, 65, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Jakovljević, I.; Pehnec, G.; Vađić, V.; Čačković, M.; Tomašić, V.; Jelin, J.D. Polycyclic aromatic hydrocarbons in PM10, PM2.5 and PM1 particle fractions in an urban area. Air Qual. Atmos. Health 2018, 11, 843–854. [Google Scholar] [CrossRef]
- Vithanage, M.; Bandara, P.C.; Novo, L.A.B.; Kumar, A.; Ambade, B.; Naveendrakumar, G.; Ranagalage, M.; Magana-Arachchi, D.N. Deposition of trace metals associated with atmospheric particulate matter: Environmental fate and health risk assessment. Chemosphere 2022, 303, 13505. [Google Scholar] [CrossRef] [PubMed]
- Świsłowski, P.; Hrabák, P.; Wacławek, S.; Liskova, K.; Antos, V.; Rajfur, M.; Ząbkowska-Wacławek, M. The Application of Active Biomonitoring with the Use of Mosses to Identify Polycyclic Aromatic Hydrocarbons in an Atmospheric Aerosol. Molecules 2021, 26, 7258. [Google Scholar] [CrossRef] [PubMed]
- Duarte, R.; Duarte, A. Urban Atmospheric Aerosols: Sources, Analysis, and Effects. Atmosphere 2020, 11, 1221. [Google Scholar] [CrossRef]
- Duarte, R.M.B.O.; Duarte, A.C. Health Effects of Urban Atmospheric Aerosols. Atmosphere 2023, 14, 309. [Google Scholar] [CrossRef]
- Abbas, I.; Badran, G.; Verdin, A.; Ledoux, F.; Roumié, M.; Courcot, D.; Garçon, G. Polycyclic aromatic hydrocarbon derivatives in airborne particulate matter: Sources, analysis and toxicity. Environ. Chem. Lett. 2018, 16, 439–475. [Google Scholar] [CrossRef]
- Mulder, M.; Dumanoglu, Y.; Efstathiou, C.; Kukučka, P.; Matejovičová, J.; Maurer, C.; Přibylová, P.; Prokeš, R.; Sofuoglu, A.; Sofuoglu, S.C.; et al. Fast formation of Nitro-PAHs in the Marine Atmosphere Constrained in a Regional Scale Lagragian Field Experiment. Environ. Sci. Technol. 2019, 53, 8914–8924. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, D.; Cui, B.; Ding, R.; Shi, X.; He, P. Association between particulate matter air pollution and lung cancer. Thorax 2020, 5, 85–87. [Google Scholar] [CrossRef]
- Flood-Garibay, J.A.; Angulo-Molina, A.; Mendez-Rojas, M.A. Particulate matter and ultrafine particles in urban air pollution and their effect on the nervous system. Environ. Sci. Process. Impacts 2023, 25, 704. [Google Scholar] [CrossRef]
- Almeida, A.S.; Ferreira, R.M.P.; Silva, A.M.S.; Duarte, A.C.; Neves, B.M.; Duarte, R.M.B.O. Structural Features and Pro-Inflammatory Effects of Water-Soluble Organic Matter in Inhalable Fine Urban Air Particles. Environ. Sci. Technol. 2020, 54, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Guevara, M. Emissions of Airborne Particulate Matter. In Issues in Environmental Science and Technology Nº42, Airborne Particulate Matter: Sources, Atmospheric Processes and Health, 1st ed.; Edited by Hester, R.E., Harrison, R.M., Querol, X., Eds.; The Royal Society of Chemistry: London, UK, 2016; pp. 1–22. Available online: https://api.pageplace.de/preview/DT0400.9781782626589_A28401559/preview-9781782626589_A28401559.pdf (accessed on 14 October 2022).
- Wei, J.; Wang, M.; Jiang, L.; Yu, X.; Mikelsons, K.; Shen, F. Global estimation of suspended particulate matter from satellite ocean color imagery. J. Geophys. Res. Oceans 2021, 126, e2021JC017303. [Google Scholar] [CrossRef]
- E.U. Copernicus Marine Service Information (CMEMS). Marine Data Store (MDS). 2023. Available online: https://data.marine.copernicus.eu/product/OCEANCOLOUR_NWS_BGC_HR_L3_NRT_009_203/contacts (accessed on 24 July 2023).
- Beyer, J.; Green, N.W.; Brooks, S.; Allan, I.J.; Ruus, A.; Gomes, T.; Bråte, I.L.N.; Schøyen, M. Blue mussels (Mytilus edulis spp.) as sentinel organisms in coastal pollution monitoring: A review. Mar. Environ. Res. 2017, 130, 338–365. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lusher, A.L.; Rotchell, J.M.; Deudero, S.; Turra, A.; Lise, I.; Bråte, N.; Sun, C.; Hossain, M.S.; Li, Q.; et al. Using mussel as a global bioindicator of coastal microplastic Pollution. Environ. Pollut. 2019, 244, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Nekhoroshkov, P.; Bezuidenhout, J.; Zinicovscaia, I.; Yushin, N.; Vergel, K.; Frontasyeva, M. Levels of Elements in Typical Mussels from the Southern Coast of Africa (Namibia, South Africa, Mozambique): Safety Aspect. Water 2021, 13, 3238. [Google Scholar] [CrossRef]
- Farrington, J.W.; Tripp, B.W.; Tanabe, S.; Subramanian, A.; Sericano, J.L.; Wade, T.L.; Knap, A.H.; Edward, D. Goldberg’s proposal of “the Mussel Watch”: Reflections after 40 years. Mar. Pollut. Bull. 2016, 110, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Mikhailov, A.T.; Torrado, M.; Méndez, J. Mussel Mytilus as model organisms in marine biotechnology: Polypeptide markers of development and sexual differentiation of the reproductive system. In New Development in Marine Biotechnology; Le Gal, Y., Halvorson, H.O., Eds.; Plenum Press: New York, NY, USA, 1998; pp. 259–263. [Google Scholar] [CrossRef]
- Pinto, J.; Costa, M.; Leite, C.; Borges, C.; Coppola, F.; Henriques, B.; Monteiro, R.; Russo, T.; Di Cosmo, A.; Soares, A.M.V.M.; et al. Ecotoxicological effects of lanthanum in Mytilus galloprovincialis. Biochemical and histopathological impacts. Aquat. Toxicol. 2019, 211, 181–192. [Google Scholar] [CrossRef]
- Curpan, A.S.; Impellitteri, F.; Plavan, G.; Ciobica, A.; Faggio, C. Review: Mytilus galloprovincialis: An essential, low-cost model organism for the impact of xenobiotics on oxidative stress and public health. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 256, 109302. [Google Scholar] [CrossRef]
- Lam, P.K.S.; Gray, J.S. The use of biomarkers in environmental monitoring programmes. Mar. Pollut. Bull. 2003, 46, 182–186. [Google Scholar] [CrossRef]
- Borković, S.; Saponjić, J.; Pavlović, S.; Blagojević, D.; Milosević, S.; Kovacević, T.; Radojičić, R.M.; Spasić, M.B.; Žikić, R.V.; Saicić, Z. The activity of antioxidant defence enzymes in the mussel Mytilus galloprovincialis from the Adriatic Sea. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 141, 366–374. [Google Scholar] [CrossRef]
- Matozzo, V.; Munari, M.; Masiero, L. Ecotoxicological hazard of a mixture of glyphosate and aminomethylphosphonic acid to the mussel Mytilus galloprovincialis (Lamarck 1819). Sci. Rep. 2019, 9, 14302. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Chora, S.; McDonagh, B.; Sheehan, D.; Starita-Geribaldi, M.; Roméo, M.; Bebianno, M. Ubiquitination and carbonylation as markers of oxidative-stress in Ruditapes decussatus. Mar. Environ. Res. 2008, 66, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Umeno, A.; Shichiri, M. Lipid peroxidation biomarkers for evaluating oxidative stress and assessing antioxidant capacity in vivo. J. Clin. Biochem. Nutr. 2013, 52, 9–16. [Google Scholar] [CrossRef] [PubMed]
- NIST Office of Reference Materials. Standard Reference Material® 1648a-Urban Particulate Matter, USA. National Institute of Standards & Technology-Certificate of Analysis. Available online: https://tsapps.nist.gov/srmext/certificates/1648a.pdf (accessed on 31 March 2022).
- Coelho, J.; Court, M.; Otjacques, E.; Lopes, V.; Paula, J.; Repolho, T.; Diniz, M.; Rosa, R. Effects of tidal emersion and marine heatwaves on cuttlefish early ontogeny. Mar. Biol. 2022, 170, 3. [Google Scholar] [CrossRef]
- Cid, A.; Picado, A.; Correia, J.; Chaves, R.; Silva, H.; Caldeira, J.; Matos, A.P.A.; Diniz, M. Oxidative stress and histological changes following exposure to diamond nanoparticles in the freshwater Asian clam Corbicula fluminea (Müller, 1774). J. Hazard. Mater. 2014, 284, 27–34. [Google Scholar] [CrossRef]
- Matos, B.; Martins, M.; Samamed, A.; Sousa, D.; Ferreira, I.; Diniz, M. Toxicity Evaluation of Quantum Dots (ZnS and CdS) Singly and Combined in Zebrafish (Danio rerio). Int. J. Environ. Res. Public Health 2019, 17, 232. [Google Scholar] [CrossRef]
- Larguinho, M.; Correia, D.; Diniz, M.; Batista, P. Evidence of one-way flow bioaccumulation of gold nanoparticles across two trophic levels. J. Nanoparticle Res. 2014, 16, 2549. [Google Scholar] [CrossRef]
- Diniz, M.; Matos, A.; Lourenço, J.; Castro, L.; Peres, I.; Mendonça, E.; Picado, A. Liver Alterations in Two Freshwater Fish Species (Carassius auratus and Danio rerio) Following Exposure to Different TiO2 Nanoparticle Concentrations. Microsc. Microanal. 2013, 19, 1131–1140. [Google Scholar] [CrossRef]
- European Animal Research Association. EU Regulations on Animal Research. Available online: https://www.eara.eu/animal-research-law (accessed on 14 December 2023).
- NSW Department of Primary Industries and Animal Research Review Panel. Three Rs. Available online: https://www.animalethics.org.au/three-rs (accessed on 14 December 2023).
- Smoot, J.; Padilla, S.; Farraj, A.K. The utility of alternative models in particulate matter air pollution toxicology. Curr. Res. Toxicol. 2022, 3, 100077. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Kong, F.; Li, M.; Xi, M.; Yu, Z. How do trophic magnification factors (TMFs) and biomagnification factors (BMFs) perform on toxic pollutant bioaccumulation estimation in coastal and marine food webs. Reg. Stud. Mar. Sci. 2021, 44, 101797. [Google Scholar] [CrossRef]
- Ostro, B.; Hu, J.; Goldberg, D.; Reynolds, P.; Hertz, A.; Bernstein, L.; Kleeman, M. Associations of Mortality with Long-Term Exposures to Fine and Ultrafine Particles, Species and Sources: Results from the California Teachers Study Cohort. Environ. Health Perspect. 2015, 123, 549–556. [Google Scholar] [CrossRef] [PubMed]
- EEA. Premature Deaths Due to Exposure to Fine Particulate Matter in Europe. European Environmental Agency. December 2022. Available online: https://www.eea.europa.eu/ims/health-impacts-of-exposure-to (accessed on 14 December 2022).
- Kim, J.; Son, J.; Kim, J.; Kim, M.; Jeong, S.; Park, T.; Son, S.W.; Ryu, H. Particulate matter (PM)2.5 affects keratinocytes via endoplasmic reticulum (ER) stress- mediated suppression of apoptosis. Mol. Cell. Toxicol. 2020, 16, 129–137. [Google Scholar] [CrossRef]
- Brunekreef, B.; Forsberg, B. Epidemiological evidence of effects of coarse airborne particles on health. Eur. Respir. J. 2005, 26, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Zeb, B.; Alam, K.; Sorooshian, A.; Blaschke, T.; Ahmad, I.; Shahid, I. On the Morphology and Composition of Particulate Matter in an Urban Environment. Aerosol. Air Qual. Res. 2018, 18, 1431–1447. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Hu, H.; Zhang, Y.; Feng, L.; Shi, Y.; Miller, M.; Sun, Z. Multi-organ toxicity induced by fine particulate matter PM2.5 in zebrafish (Danio rerio) model. Chemosphere 2017, 180, 24–32. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, Z.; Rajendran, R.S.; Zhu, C.; Wang, X.; Liu, K.; Cen, J. Exposure of particulate matter (PM10) induces neurodevelopmental toxicity in zebrafish embryos. Neurotoxicology 2021, 87, 208–218. [Google Scholar] [CrossRef]
- Cen, J.; Jia, Z.-L.; Zhu, C.-Y.; Wang, X.-F.; Zhang, F.; Chen, W.-Y.; Liu, K.-C.; Li, S.-Y.; Zhang, Y. Particulate matter (PM10) induces cardiovascular developmental toxicity in zebrafish embryos and larvae via the ERS, Nrf2 and wnt pathways. Chemosphere 2020, 250, 126288. [Google Scholar] [CrossRef]
- Ren, F.; Ji, C.; Huang, Y.; Aniagu, S.; Jiang, Y.; Chen, T. AHR-mediated ROS production contributes to the cardiac developmental toxicity of PM2.5 in zebrafish embryos. Sci. Total Environ. 2020, 719, 135097. [Google Scholar] [CrossRef]
- Zhao, P.; Lu, W.; Hong, Y.; Chena, J.; Dong, S.; Huang, Q. Long-term wet precipitation of PM2.5 disturbed the gut microbiome and inhibited the growth of marine medaka Oryzias melastigma. Sci. Total Environ. 2021, 755, 142512. [Google Scholar] [CrossRef]
- Hartono, D.; Lioe, B.; Zhang, Y.; Li, B.; Yu, J. Impacts of particulate matter (PM2.5) on the behavior of freshwater snail Parafossarulus striatulus. Sci. Rep. 2017, 7, 644. [Google Scholar] [CrossRef]
- Kurelec, B.; Pivčević, B. Evidence for a multixenobiotic resistance mechanism in the mussel Mytilus galloprovincialis. Aquat. Toxicol. 1991, 19, 291–301. [Google Scholar] [CrossRef]
- Franzellitti, S.; Capolupo, M.; Wathsala, R.; Valbonesi, P.; Fabbri, E. The Multixenobiotic resistance system as a possible protective response triggered by microplastic ingestion in Mediterranean mussels (Mytilus galloprovincialis): Larvae and adult stages. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 219, 50–58. [Google Scholar] [CrossRef]
- Galimany, E.; Ramón, M.; Ibarrola, I. Feeding behavior of the mussel Mytilus galloprovincialis (L.) in a Mediterranean estuary: A field study. Aquaculture 2011, 314, 236–243. [Google Scholar] [CrossRef]
- Almeida, E.A.; Bainy, A.C.D.; Loureiro, A.P.M.; Martinez, G.R.; Miyamoto, S.; Onuki, J.; Barbosa, L.F.; Garcia, C.C.M.; Prado, F.M.; Ronsein, G.E.; et al. Oxidative stress in Perna perna and other bivalves as indicators of environmental stress in the Brazilian marine environment: Antioxidants, lipid peroxidation and DNA damage. Comp. Biochem. Physiol. A 2007, 146, 588–600. [Google Scholar] [CrossRef]
- Adorno, H.; Souza, I.; Monferrán, M.; Wunderlin, D.; Fernandes, M.; Monteiro, D. A multi-biomarker approach to assess the sublethal effects of settleable atmospheric particulate matter from an industrial area on Nile tilapia (Oreochromis niloticus). Sci. Total Environ. 2023, 856, 159168. [Google Scholar] [CrossRef]
- Duarte, L.F.A.; Ortega, A.D.S.B.; Paço, M.S.; Sadauskas-Henrique, H.; Cesar-Ribeiro, C.; Souza, I.C.; Monteiro, R.; Monferrán, M.V.; Wunderlin, D.A.; Fernandes, M.N.; et al. Settleable atmospheric particulate matter harms a marine invertebrate: Integrating chemical and biological damage in a bivalve model. Sci. Total Environ. 2023, 881, 163380. [Google Scholar] [CrossRef]
- Monteiro, R.; Souza, I.C.; Morozesk, M.; Pereira Soares, M.; De Angelis, C.F.; Vieira, N.S.; Bendhack, F.; Monferran, M.V.; Wunderlin, D.A.; Fernandes, M.N. Metalliferous atmospheric settleable particulate matter action on the fat snook fish (Centropomus parallelus): Metal bioaccumulation, antioxidant responses and histological changes in gills, hepatopancreas and kidneys. Chemosphere 2023, 330, 138715. [Google Scholar] [CrossRef]
- Moron, M.; Depierre, J.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Revel, M.; Lagarde, F.; Perrein-Ettajani, H.; Bruneau, M.; Akcha, F.; Sussarellu, R.; Rouxel, J.; Costil, K.; Decottignies, P.; Cognie, B.; et al. Tissue-Specific Biomarker Responses in the Blue Mussel Mytilus spp. Exposed to a Mixture of Microplastics at Environmentally Relevant Concentrations. Front. Environ. Sci. 2019, 7, 33. [Google Scholar] [CrossRef]
- Dobal, V.; Suárez, P.; Ruiz, Y.; García-Martín, O.; Juan, F. Activity of antioxidant enzymes in Mytilus galloprovincialis exposed to tar: Integrated response of different organs as pollution biomarker in aquaculture areas. Aquaculture 2022, 548, 737638. [Google Scholar] [CrossRef]
- Wilczek, G.; Kramarz, P.; Babczyńska, A. Activity of carboxylesterase and glutathione S-transferase in different life-stages of carabid beetle (Poecilus cupreus) exposed to toxic metal concentrations. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 134, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, H.; Zhao, J.; Zhang, W.; Zhang, L.; Sun, S.; Yang, D.; Cheng, B.; Wang, Q. The role of GST omega in metabolism and detoxification of arsenic in clam Ruditapes philippinarum. Aquat. Toxicol. 2018, 204, 9–18. [Google Scholar] [CrossRef]
- Han, J.; Park, Y.; Jeong, H.; Park, C.J. Effects of particulate matter (PM2.5) on life history traits, oxidative stress, and defensome system in the marine copepod Tigriopus japonicus. Mar. Pollut. Bull. 2022, 178, 113588. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, I.; Ferreira, I.J.; Duarte, R.M.B.O.; Diniz, M. Effects of Exposure to Urban Atmospheric Particulate Matter Suspended in Seawater on the Mussel Mytilus galloprovincialis. Environments 2024, 11, 12. https://doi.org/10.3390/environments11010012
Rodrigues I, Ferreira IJ, Duarte RMBO, Diniz M. Effects of Exposure to Urban Atmospheric Particulate Matter Suspended in Seawater on the Mussel Mytilus galloprovincialis. Environments. 2024; 11(1):12. https://doi.org/10.3390/environments11010012
Chicago/Turabian StyleRodrigues, Inês, Inês João Ferreira, Regina M. B. O. Duarte, and Mário Diniz. 2024. "Effects of Exposure to Urban Atmospheric Particulate Matter Suspended in Seawater on the Mussel Mytilus galloprovincialis" Environments 11, no. 1: 12. https://doi.org/10.3390/environments11010012
APA StyleRodrigues, I., Ferreira, I. J., Duarte, R. M. B. O., & Diniz, M. (2024). Effects of Exposure to Urban Atmospheric Particulate Matter Suspended in Seawater on the Mussel Mytilus galloprovincialis. Environments, 11(1), 12. https://doi.org/10.3390/environments11010012