Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets

 ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sample Collection
2.3. Extraction of Polyphenols and Determination of the Total Phenolic Content (TPC) from Feed
2.4. Extraction of polyphenols and Determination of the Total Phenolic Content (TPC) from Plasma and Organs (Liver, Spleen, Kidney, Mesenteric Lymph Nodes, Heart and Brain) and Longissimus Dorsi Muscle
2.5. Evaluation of the Antioxidant Activity Using the DPPH Method (AAR) in Organs (Liver, Spleen, Kidney, Mesenteric Lymph Nodes, Heart and Brain) and LONGISSIMUS DORSI MUSCLE
2.6. Preparation of Tissue Lysates
2.7. Measurement of Tissue Lipid Peroxidation—TBARS-MDA (Liver, Spleen and Kidneys)
2.8. Measurement of Total Antioxidant Status (TAS) (Liver, Spleen and Kidneys)
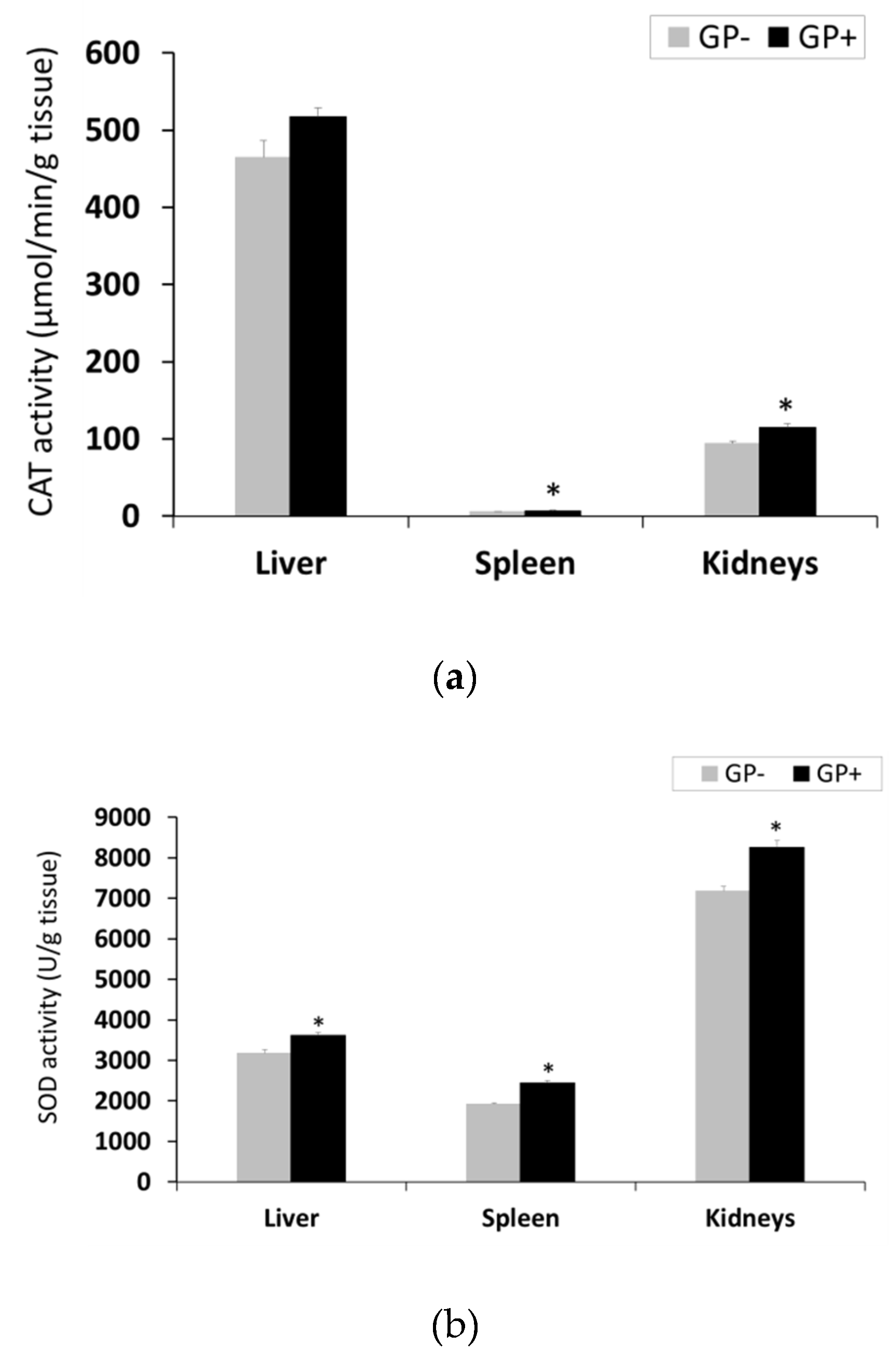
2.9. Measurement of Tissue Catalase (CAT), Superoxide Dismutase (SOD) and Glutathione Peroxidase (GPx) Activity (Liver, Spleen and Kidneys)
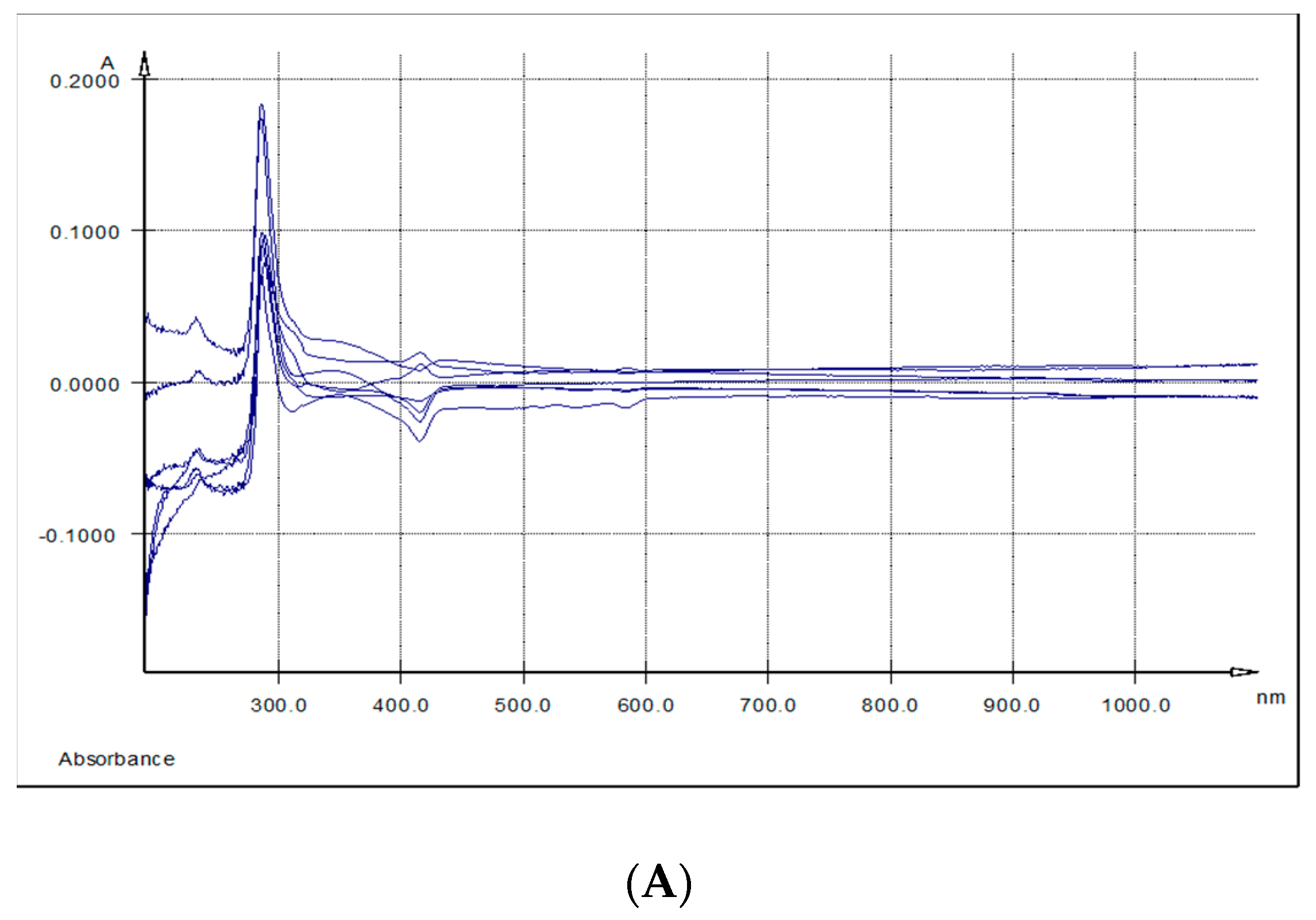
2.10. Measurement of UV-VIS Spectra from Liver, Spleen and Kidneys Samples
2.11. Measurement of Plasma Biochemical Parameters
2.12. Statistical Analysis
3. Results
3.1. Total Polyphenols Content (TPC) and Antioxidant Activity (AAR) in Feed Compound and Its Components
3.2. Effect of Dietary GP on Performance
3.3. Effect of Dietary GP on Health Status (Plasma Biochemical Parameters)
3.4. Total Polyphenols Content (TPC) in Blood and Its Fractions
3.5. Total Polyphenols Content (TPC) and Antiradical (DPPH) Activity (AAR) in Organs (Liver, Spleen, Kidney, Mesenteric Lymph Nodes, Heart and Brain) and LONGISSIMUS DORSI MUSCLE
3.6. Effect of GP Diet on Antioxidant Status in Liver, Spleen and Kidney
3.7. Qualitative Assessment of Polyphenols Absorption in Liver, Spleen and Kidney by UV-Vis Spectra Measurement
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wadhwa, M.; Bakshi, M.P.; Makkar, H.P. Waste to worth: Fruit wastes and by-products as animal feed. CAB Rev. 2015, 10, 1–26. [Google Scholar] [CrossRef]
- Llobera, A.; Cañellas, J. Dietary fibre content and antioxidant activity of Manto Negro red grape (Vitis vinifera): Pomace and stem. Food chem. 2007, 101, 659–666. [Google Scholar] [CrossRef]
- Cotoras, M.; Vivanco, H.; Melo, R.; Aguirre, M.; Silva, E.; Mendoza, L. In vitro and in vivo evaluation of the antioxidant and prooxidant activity of phenolic compounds obtained from grape (Vitis vinifera) pomace. Molecules 2014, 19, 21154–21167. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Chamorro, S.; Ignacio, A. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Goñí, I.; Brenes, A.; Centeno, C.; Viveros, A.; Saura-Calixto, F.; Rebole, A.; Arija, I.; Estevez, R. Effect of dietary grape pomace and vitamin E on growth performance, nutrient digestibility, and susceptibility to meat lipid oxidation in chickens. Poult. Sci. 2007, 86, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef]
- Choi, C.-S.; Chung, H.-K.; Choi, M.-K.; Kang, M.-H. Effects of grape pomace on the antioxidant defense system in diet-induced hypercholesterolemic rabbits. Nutr. Res. Pract. 2010, 4, 114–120. [Google Scholar] [CrossRef]
- Santos, N.; Santos, G.T.D.; Silva-Kazama, D.C.; Grande, P.A.; Pintro, P.M.; De Marchi, F.E.; Jobim, C.C.; Petit, H.V. Production, composition and antioxidants in milk of dairy cows fed diets containing soybean oil and grape residue silage. Livest. Sci. 2014, 159, 37–45. [Google Scholar] [CrossRef]
- Chedea, V.S.; Pelmus, R.S.; Lazar, C.; Pistol, G.C.; Calin, L.G.; Toma, S.M.; Dragomir, C.; Taranu, I. Effects of a diet containing dried grape pomace on blood metabolites and milk composition of dairy cows. J. Sci Food Agric. 2017, 97, 2516–2523. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Rivas, C.; Gallardo, B.; Mantecón, Á.R.; del Álamo-Sanza, M.; Manso, T. Evaluation of grape pomace from red wine by-product as feed for sheep. J. Sci. Food Agric. 2017, 6, 1885–1893. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Kim, I. Effect of dietary grape pomace fermented by Saccharomyces boulardii on the growth performance, nutrient digestibility and meat quality in finishing pigs. Asian-Australas. J. Anim. Sci. 2011, 24, 1763–1770. [Google Scholar] [CrossRef]
- Cho, S.; Cho, J.H.; Hwang, O.H.; Yang, S.H.; Park, K.H.; Choi, D.Y.; Yoo, Y.H.; Kim, I.H. Effects of fermented diets including grape and apple pomace on amino acid digestibility, nitrogen balance and volatile fatty acid (VFA) emission in finishing pigs. J. Anim. Vet. Adv. 2012, 11, 3444–3451. [Google Scholar] [CrossRef]
- Fiesel, A.; Fiesel, A.; Gessner, D.K.; Most, E.; Eder, K. Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet. Res. 2014, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, H.J.; Mantovani, G.; Alborali, G.L.; Caputo, J.M.; Savoini, G.; Dell’Orto, V.; Bontempo, V. The effect of plant polyphenols on the antioxidant defence system of weaned piglets subjected to an Escherichia coli challenge. J. Anim. Feed Sci. 2014, 663, 127. [Google Scholar] [CrossRef]
- Zhang, H.J.; Jiang, X.R.; Mantovani, G.; Lumbreras, A.E.V.; Comi, M.; Alborali, G.; Savoini, G.; Dell’Orto, V.; Bontempo, V. Modulation of plasma antioxidant activity in weaned piglets by plant polyphenols. Ital. J. Anim. Sci. 2014, 13, 3242. [Google Scholar] [CrossRef]
- Fraga, C.G.; Oteiza, P.I. Dietary flavonoids: Role of (−)-epicatechin and related procyanidins in cell signaling. Free Rad. Biol. Med. 2011, 51, 813–823. [Google Scholar] [CrossRef]
- Spencer, J.P.; Chaudry, F.; Pannala, A.S.; Srai, S.K.; Debnam, E.; Rice-Evans, C. Decomposition of cocoa procyanidins in the gastric milieu. Biochem. Biophys. Res. Comm. 2000, 272, 236–241. [Google Scholar] [CrossRef]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves performance, antioxidant status, fecal microbiota and meat quality of piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Gerasopoulos, K.; Stagos, D.; Petrotos, K.; Kokkas, S.; Kantas, D.; Goulas, P.; Kouretas, D. Feed supplemented with polyphenolic byproduct from olive mill wastewater processing improves the redox status in blood and tissues of piglets. Food Chem. Toxicol. 2015, 86, 319–327. [Google Scholar] [CrossRef]
- Chedea, V.S.; Palade, L.; Marin, D.; Pelmus, R.; Habeanu, M.; Rotar, M.; Gras, M.; Pistol, G.; Taranu, I. Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients 2018, 10, 588. [Google Scholar] [CrossRef]
- Burlacu, G.; Cavache, A.; Burlacu, R. Potentialul Productiv al Nutreturilor si Utilizarea Lor; Editura Ceres: București, Romania, 2002. [Google Scholar]
- Sehm, J.; Treutter, D.; Lindermayer, H.; Meyer, H.H.D.; Pfaffl, M.W. The influence of apple-or red-grape pomace enriched piglet diet on blood parameters, bacterial colonisation, and marker gene expression in piglet white blood cells. Food Nutr. Sci. 2011, 2, 366. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vit. 1965, 16, 144–158. [Google Scholar]
- Arnous, A.; Makris, D.P.; Kefalas, P. Effect of principal polyphenolic components in relation to antioxidant characteristics of aged red wines. J. Agric. Food Chem. 2001, 49, 5736–5742. [Google Scholar] [CrossRef]
- Wettasinghe, M.; Shahidi, F. Scavenging of reactive-oxygen species and DPPH free radicals by extracts of borage and evening primrose meals. Food Chem. 2000, 70, 17–26. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Chedea, V.S.; Braicu, C.; Socaciu, C. Antioxidant/prooxidant activity of a polyphenolic grape seed extract. Food Chem. 2010, 121, 132–139. [Google Scholar] [CrossRef]
- Chamorro, S.; Viveros, A.; Centeno, C.; Romero, C.; Arija, I.; Brenes, A. Effects of dietary grape seed extract on growth performance, amino acid digestibility and plasma lipids and mineral content in broiler chicks. Animal 2013, 7, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Fiesel, A.; Ehrmann, M.; Geßner, D.K.; Most, E.; Eder, K. Effects of polyphenol-rich plant products from grape or hop as feed supplements on iron, zinc and copper status in piglets. Arch. Anim. Nutr. 2015, 69, 276–284. [Google Scholar] [CrossRef]
- Rauw, W.; Corella, D.; Soler, J.; Reixach, J.; Tibau, J.; Prat, J.M.; Diaz, I.; Gómez-Raya, L. Behaviour influences cholesterol plasma levels in a pig model. Animal 2007, 1, 865–871. [Google Scholar] [CrossRef]
- Innis, S.M. The colostrum-deprived piglet as a model for study of infant lipid nutrition. J. Nutr. 1993, 123 (Suppl. 2), 386–390. [Google Scholar] [CrossRef]
- Burke, A.; Weiler, H. The effect of prostaglandin E2 (PGE2) and long-chain polyunsaturated fatty acids (LC PUFA) on bone formation in piglets: A model for bone growth in nutritional investigation. Prostaglandins Leukot Essent Fat. Acids 2002, 67, 229–235. [Google Scholar] [CrossRef]
- Guilloteau, P.; Zabielski, R.; Hammon, H.M.; Metges, C.C. Nutritional programming of gastrointestinal tract development. Is the pig a good model for man? Nutr. Res. Rev. 2010, 23, 4–22. [Google Scholar] [CrossRef]
- Ray, S.D.; Parmar, M.; Syed, I.; Rathod, J.; Zinkovsky, D.; Bulku, E.; Gigliotti, J.; Hackman, R.M.; Stohs, S.J. Long term exposure effect of a unique metabolic nutrition system containing a diverse group of phytochemicals on serum chemistry and genomic and non-genomic changes in the liver of female B6C3F1 mice. Phytother. Res. 2008, 22, 458–471. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Barbosa, S.; Silva, M.; Monteiro, D.; Pinheiro, V.; Mourão, J.L.; Fernandes, J.; Rocha, S.; Belo, L.; Santos-Silva, A. The effect of olive leaf supplementation on the constituents of blood and oxidative stability of red blood cells. J. Funct. Foods 2014, 9, 271–279. [Google Scholar] [CrossRef]
- Ginsburg, I.; Kohen, R.; Koren, E. Quantifying oxidant-scavenging ability of blood. N. Engl. J. Med. 2011, 364, 883–885. [Google Scholar] [CrossRef]
- Ginsburg, I.; Kohen, R.; Koren, E. Microbial and host cells acquire enhanced oxidant-scavenging abilities by binding polyphenols. Arch. Biochem. Biophys. 2011, 506, 12–23. [Google Scholar] [CrossRef]
- Serra, A.; Macià, A.; Romero, M.-P.; Anglès, N.; Morelló, J.R.; Motilva, M.-J. Distribution of procyanidins and their metabolites in rat plasma and tissues after an acute intake of hazelnut extract. Food Funct. 2011, 2, 562–568. [Google Scholar] [CrossRef]
- Chedea, V.S.; Choueiri, L.; Jisaka, M.; Kefalas, P. o-Quinone involvement in the prooxidant tendency of a mixture of quercetin and caffeic acid. Food Chem. 2012, 135, 1999–2004. [Google Scholar] [CrossRef]
- Serra, A.; Macià, A.; Romero, M.-P.; Piñol, C.; Motilva, M.-J. Rapid methods to determine procyanidins, anthocyanins, theobromine and caffeine in rat tissues by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2011, 879, 1519–1528. [Google Scholar] [CrossRef]
- Tsang, C.; Auger, C.; Mullen, W.; Bornet, A.; Rouanet, J.-M.; Crozier, A.; Teissedre, P.-L. The absorption, metabolism and excretion of flavan−3-ols and procyanidins following the ingestion of a grape seed extract by rats. Br. J. Nutr. 2005, 94, 170–181. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef]
- Fragou, S.; Fegeros, K.; Xylouri, E.; Baldi, A.; Politis, I. Effect of vitamin E supplementation on various functional properties of macrophages and neutrophils obtained from weaned piglets. Transbound. Emerg. Dis. 2004, 51, 178–183. [Google Scholar] [CrossRef]
- Eicher, S.; McKee, C.A.; Carroll, J.A.; Pajor, E.A. Supplemental vitamin C and yeast cell wall β-glucan as growth enhancers in newborn pigs and as immunomodulators after an endotoxin challenge after weaning. J. Anim. Sci. 2006, 84, 2352–2360. [Google Scholar] [CrossRef]
- Ali Rajput, S.; Sun, L.; Zhang, N.; Mohamed Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.; Zhang, J.; Qi, D. Ameliorative effects of grape seed proanthocyanidin extract on growth performance, immune function, antioxidant capacity, biochemical constituents, liver histopathology and aflatoxin residues in broilers exposed to aflatoxin B1. Toxins 2017, 9, 371. [Google Scholar] [CrossRef]
- Di Giancamillo, A.; Rossi, R.; Pastorelli, G.; Deponti, D.; Carollo, V.; Casamassima, D.; Domeneghini, C.; Corino, C. The effects of dietary verbascoside on blood and liver oxidative stress status induced by a high n-6 polyunsaturated fatty acids diet in piglets. J. Anim. Sci. 2015, 93, 2849–2859. [Google Scholar] [CrossRef]
- Gessner, D.K.; Bonarius, M.; Most, E.; Fiesel, A.; Eder, K. Effects of polyphenol-rich plant products from grape or hop as feed supplements on the expression of inflammatory, antioxidative, cytoprotective and endoplasmic reticulum stress-related genes and the antioxidative status in the liver of piglets. J. Anim. Physiol. Anim. Nutr. 2016, 101, e185–e194. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Control Diet | GP+ Diet |
|---|---|---|
| Corn | 57.32 | 53.90 |
| Rice meal | - | - |
| Wheat | 10.00 | 8.00 |
| Sunflower meal (31.94% CP) | 5.00 | 5.00 |
| Soybean meal (44% CP) | 16.00 | 16.00 |
| Sunflower oil | 0.20 | 0.70 |
| Milk powder | 3.00 | 3.00 |
| Corn gluten meal (60% CP) | 4.00 | 4.00 |
| Grape pomace | - | 5.00 |
| Monocalcium phosphate | 1.25 | 1.25 |
| Limestone | 1.56 | 1.48 |
| NaCl | 0.20 | 0.20 |
| DL-Methionine | 0.03 | 0.05 |
| L-Lysine HCL | 0.34 | 0.32 |
| Choline | 0.10 | 0.10 |
| Mineral–vitamin premix | 1.00 | 1.00 |
| Calculated nutrient content according to [21] | ||
| Dry matter (%) | 88.31 | 88.01 |
| Crude Protein (%) | 18.29 | 18.36 |
| Digestible crude protein (%) | 14.98 | 15.06 |
| Fat (%) | 2.85 | 2.96 |
| Crude fiber (%) | 4.07 | 5.30 |
| Metabolizable energy (Kcal/kg) | 3169 | 3147 |
| Lysine (%) | 1.08 | 1.08 |
| Digestible Lysine (%) | 0.92 | 0.92 |
| Met + Cys (%) | 0.65 | 0.65 |
| Calcium (%) | 0.90 | 0.90 |
| Phosphorus (%) | 0.65 | 0.65 |
| Sample | Acetone Extraction | Methanol Extraction | ||
| Total Polyphenols (mg GAE/100g raw material) | AAR (DPPH Assay) (μM Trolox equivalents) | Total Polyphenols (mg GAE/100g raw material) | AAR (DPPH Assay) (μM Trolox Equivalents) | |
| Ingredients | ||||
| Corn Grains | 694.48 ± 3.02 g,† | 237.39 ± 6.62 g,† | 682.24 ± 6.42 f,† | 253.68 ± 6.98 f,‡ |
| Wheat Grains | 697.39 ± 11.64 f,g,† | 297.74 ± 3.99 f,† | 719.1 ± 6.89 e,‡ | 273.92 ± 4.38 e,f,‡ |
| Sunflower Meal | 1588.55 ± 69.96 a,† | 683.10 ± 4.37 a,† | 942.5 ± 27.23 a,‡ | 678.85 ± 4.87 a,† |
| Soybean Meal | 839.07 ± 14.78 c,d,e,† | 644.64 ± 9.79 b,† | 746.14 ± 6.1 d,e,‡ | 466.8 9 ± 7.79 c,‡ |
| Dried Grape Pomace | 853.32 ± 19.75 b,c,d,† | 585.57 ± 3.61 c,† | 781.37 ± 7.26 b,‡ | 482.99 ± 4.87 b,‡ |
| Compound feeds | ||||
| Control (GP−) | 743.97 ± 10.66 e,f,g,† | 417.51 ± 2.46 e,† | 677.22 ± 7.44 g,‡ | 233.78 ± 10.25 g,‡ |
| Grape Pomace (GP+) | 810.60 ± 26.40 d,e,† | 454.36 ± 12.88 d,† | 749.95 ± 2.22 c,d,† | 410.52 ± 10.46 d,‡ |
| Items | 15 Days | 36 Days | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GP− | SEM | GP+ | SEM | GP− | SEM | GP+ | SEM | Diet | Time | D × T | |
| Glucose (mg/dL) | 96.81 | 4.58 | 100.05 | 3.73 | 90.24 | 1.73 | 80.76 | 3.40 | 0.826 | 0.09 | 0.335 |
| Total Cholesterol (mg/dL) | 83.32 | 3.49 | 76.89 | 2.18 | 83.30 | 1.43 | 85.65 | 1.67 | 0.388 | 0.069 | 0.068 |
| Triglycerides (mg/dL) | 65.91a | 0.76 | 63.48a | 2.90 | 62.95a | 3.85 | 48.18b | 3.46 | 0.07 | 0.040 | 0.046 |
| Phosphorus (mg/dL) | 7.77c | 0.34 | 7.82 b,c | 0.21 | 8.62 a,b | 0.12 | 8.69a | 0.13 | 0.778 | 0.000 | 0.982 |
| Calcium (mg/dL) | 9.52 a,b | 0.24 | 10.16a | 0.27 | 8.97b | 0.33 | 9.20 a,b | 0.30 | 0.141 | 0.012 | 0.479 |
| Iron (ug/dl) | 134.77 | 9.53 | 147.25 | 12.19 | 130.24 | 8.63 | 127.39 | 2.99 | 0.595 | 0.183 | 0.399 |
| Magnesium (mg/dL) | 1.36 | 0.05 | 1.67 | 0.18 | 1.49 | 0.06 | 1.46 | 0.03 | 0.155 | 0.703 | 0.103 |
| Total protein (mg/dL) | 5.13 | 0.19 | 5.23 | 0.12 | 5.11 | 0.19 | 5.24 | 0.17 | 0.515 | 0.963 | 0.948 |
| Albumin (g/L) | 2.99c | 0.10 | 3.39b | 0.09 | 3.97a | 0.02 | 3.99a | 0.01 | 0.004 | 0.000 | 0.008 |
| Bilirubin (mg/dL) | 0.12 | 0.013 | 0.11 | 0.010 | 0.13 | 0.015 | 0.12 | 0.013 | 0.451 | 0.451 | 1.000 |
| Urea (mg/dL) | 20.03 | 1.34 | 17.38 | 1.33 | 21.08 | 1.3 | 19.19 | 1.65 | 0.117 | 0.319 | 0.790 |
| Creatinine (mg/dL) | 0.94 | 0.03 | 0.98 | 0.035 | 0.94 | 0.04 | 1.02 | 0.04 | 0.123 | 0.576 | 0.672 |
| ALP (U/L) | 127.30 | 5.97 | 155.50 | 9.92 | 138.04 | 8.36 | 140.48 | 5.13 | 0.779 | 0.051 | 0.098 |
| ALT (U/L) | 50.43 | 2.43 | 53.92 | 4.89 | 52.31 | 2.53 | 56.56 | 4.13 | 0.296 | 0.540 | 0.918 |
| AST (U/L) | 49.17 | 4.02 | 45.64 | 3.36 | 47.33 | 2.70 | 50.06 | 3.05 | 0.976 | 0.843 | 0.354 |
| GGT (U/L) | 25.23 | 1.79 | 31.16 | 3.69 | 26.12 | 2.21 | 26.88 | 2.46 | 0.213 | 0.524 | 0.333 |
| Items | After 15 Days | After 36 Days | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GP− | SEM | GP+ | SEM | GP− | SEM | GP+ | SEM | Diet (D) | Time (T) | D × T | |
| plasma | 592.62b | 2.30 | 592.90b | 5.30 | 594.81b | 3.74 | 615.52a | 5.30 | 0.022 | 0.008 | 0.026 |
| cell sediment | 600.49 | 3.65 | 614.41 | 3.41 | 602.93 | 10.24 | 614.70 | 9.43 | 0.084 | 0.852 | 0.882 |
| blood | 635.28 | 8.10 | 643.57 | 2.94 | 649.37 | 3.70 | 657.46 | 7.01 | 0.169 | 0.022 | 0.987 |
| Items | Total Polyphenols (mgGAE/100g raw material) | Antioxidant Activity (AAR by DPPH assay) (μM Trolox equivalents) | ||||
| GP− | GP+ | p-Value | GP− | GP+ | p-Value | |
| Organs | ||||||
| Liver | 175.04 ± 1.39 | 171.99 ± 1.24 | 0.119 | 109.64 ± 9.86 b | 144.41 ± 7.02 a | 0.009 |
| Spleen | 155.32 ± 3.85 | 147.56 ± 1.68 | 0.072 | 99.87 ± 6.88 | 98.48 ± 9.53 | 0.909 |
| Kidney | 143.74 ± 0.59 b | 147.80 ± 1.26 a | 0.019 | 35.38 ± 6.57 | 41.90 ± 6.80 | 0.506 |
| Mesenteric lymph nodes | 141.69 ± 1.05 | 140.40 ± 1.37 | 0.464 | 110.36 ± 7.80 | 112.36 ± 10.96 | 0.886 |
| Heart | 140.10 ± 1.26 | 140.35 ± 1.08 | 0.882 | 8.51 ± 2.13 | 11.27 ± 2.80 | 0.474 |
| Brain | 153.28 ± 1.81 | 150.90 ± 2.00 | 0.394 | 63.29 ± 8.65 | 59.75 ± 9.12 | 0.782 |
| Muscle | ||||||
| L. dorsi muscle | 122.93 ± 1.18 | 124.71 ± 1.03 | 0.270 | 21.53 ± 3.40 | 21.17 ± 2.66 | 0.935 |
| Samples | λ max (nm) | ||
|---|---|---|---|
| Liver | - | 328 | 430 |
| Spleen | 286.5 | 344 | 415 |
| Kidney | 284.6-289 | 336 | 443 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chedea, V.S.; Palade, L.M.; Pelmus, R.S.; Dragomir, C.; Taranu, I. Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets. Animals 2019, 9, 149. https://doi.org/10.3390/ani9040149
Chedea VS, Palade LM, Pelmus RS, Dragomir C, Taranu I. Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets. Animals. 2019; 9(4):149. https://doi.org/10.3390/ani9040149
Chicago/Turabian StyleChedea, Veronica Sanda, Laurentiu Mihai Palade, Rodica Stefania Pelmus, Catalin Dragomir, and Ionelia Taranu. 2019. "Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets" Animals 9, no. 4: 149. https://doi.org/10.3390/ani9040149
APA StyleChedea, V. S., Palade, L. M., Pelmus, R. S., Dragomir, C., & Taranu, I. (2019). Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets. Animals, 9(4), 149. https://doi.org/10.3390/ani9040149