Comparative Studies on the Antioxidant Profiles of Curcumin and Bisdemethoxycurcumin in Erythrocytes and Broiler Chickens



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Chicken Erythrocytes Preparation
2.3. Determination of Hemolysis
2.4. Determination of Superoxide Dismutase Activity and Malondialdehyde Levels in Erythrocytes
2.5. Animals, Diets, and Experimental Design
2.6. Sample Collection
2.7. Bird Performance
2.8. Measurement of Serum Antioxidant Index
2.9. Glutathione Redox Potential of Serum, Liver, Duodenum, Jejunum and Ileum
2.10. Real-Time Polymerase Chain Reaction (PCR) Analysis
2.11. Measurement of the ABTS Scavenging Activity and FRAP Value
2.12. Statistical Analysis
3. Results
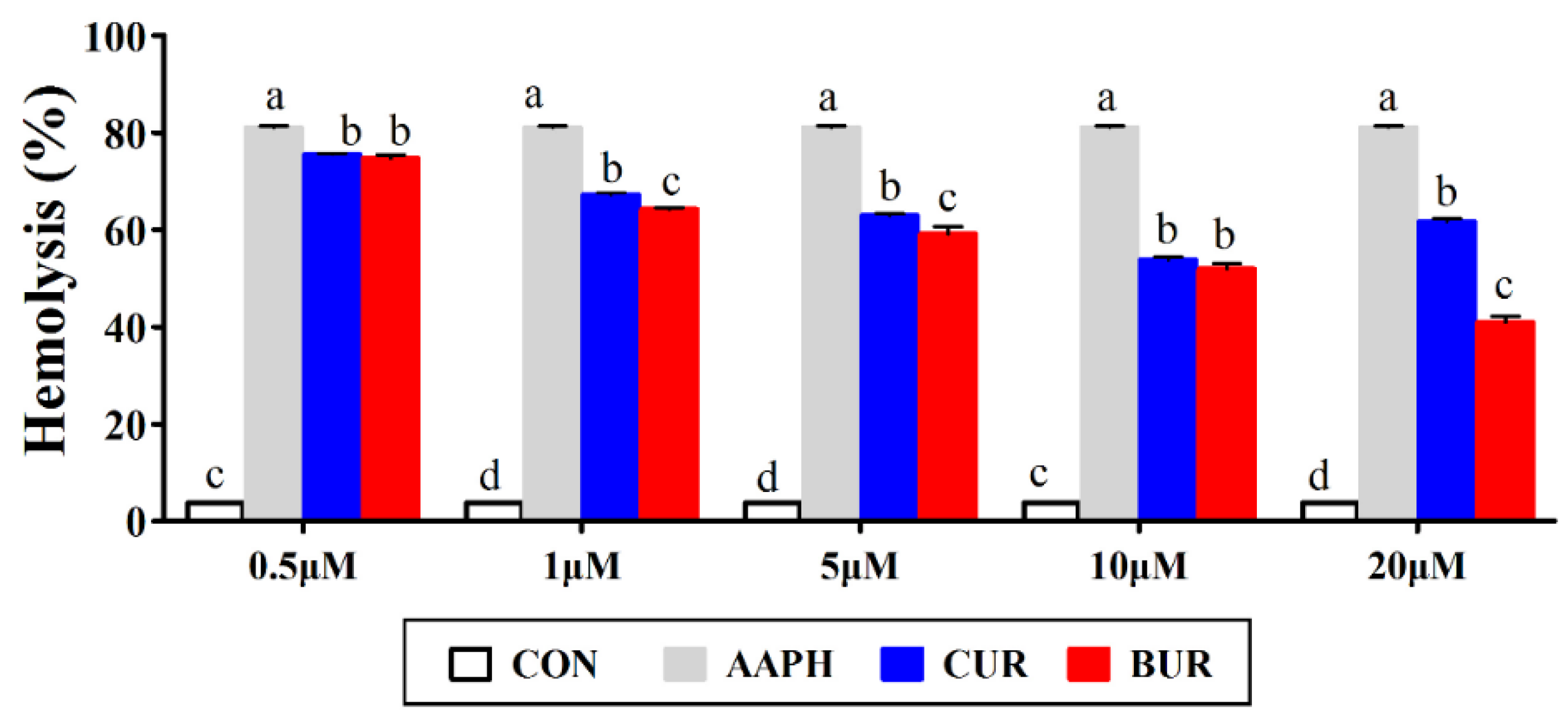
3.1. Erythrocytes Hemolysis (Experiment 1)
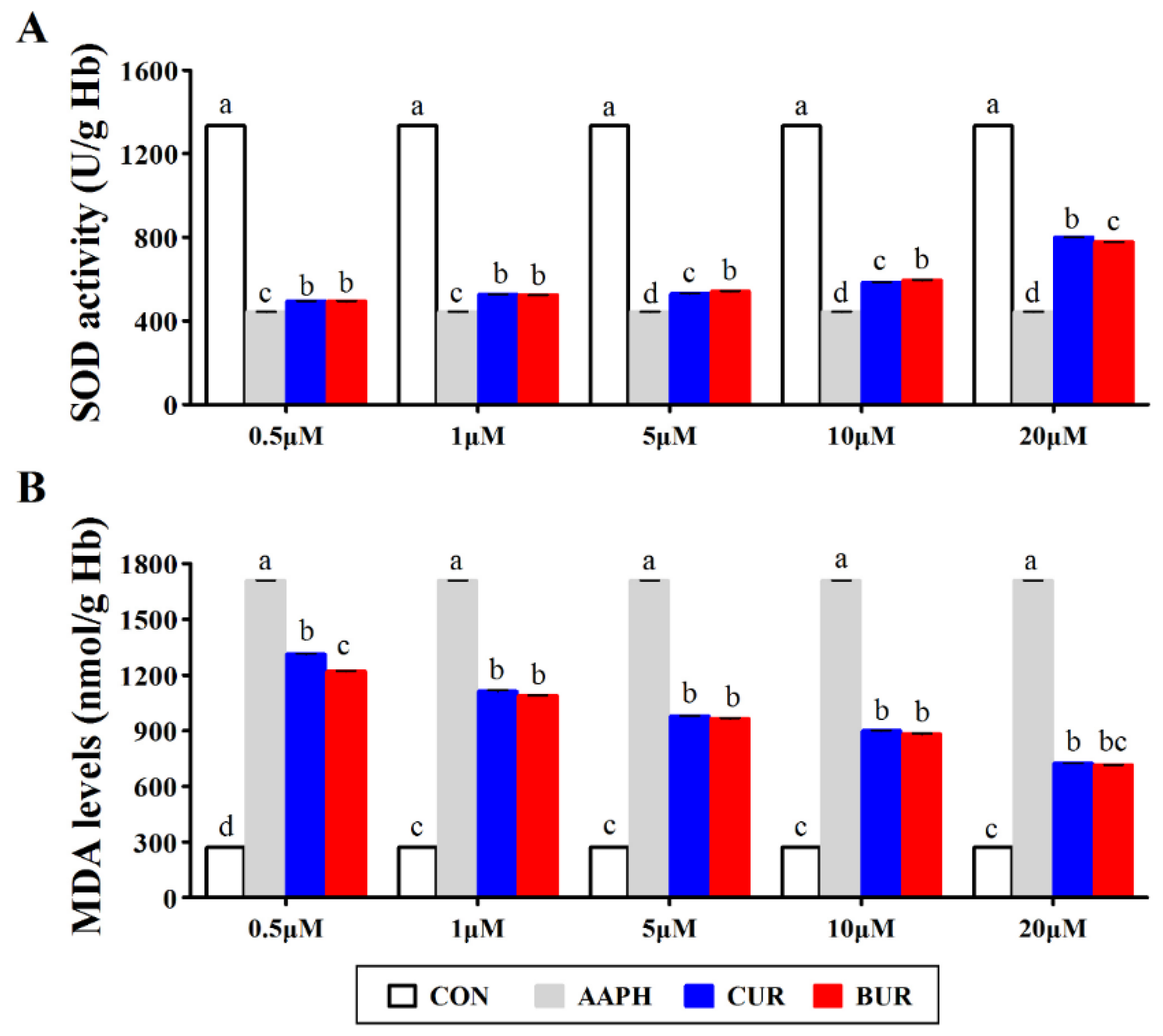
3.2. SOD Activity and MDA Level in Erythrocytes (Experiment 1)
3.3. Growth Performance (Experiment 2)
3.4. Serum Antioxidant Index (Experiment 2)
3.5. Glutathione Redox Potential in Different Tissues (Experiment 2)
3.6. Antioxidant-Related Gene Expression (Experiment 2)
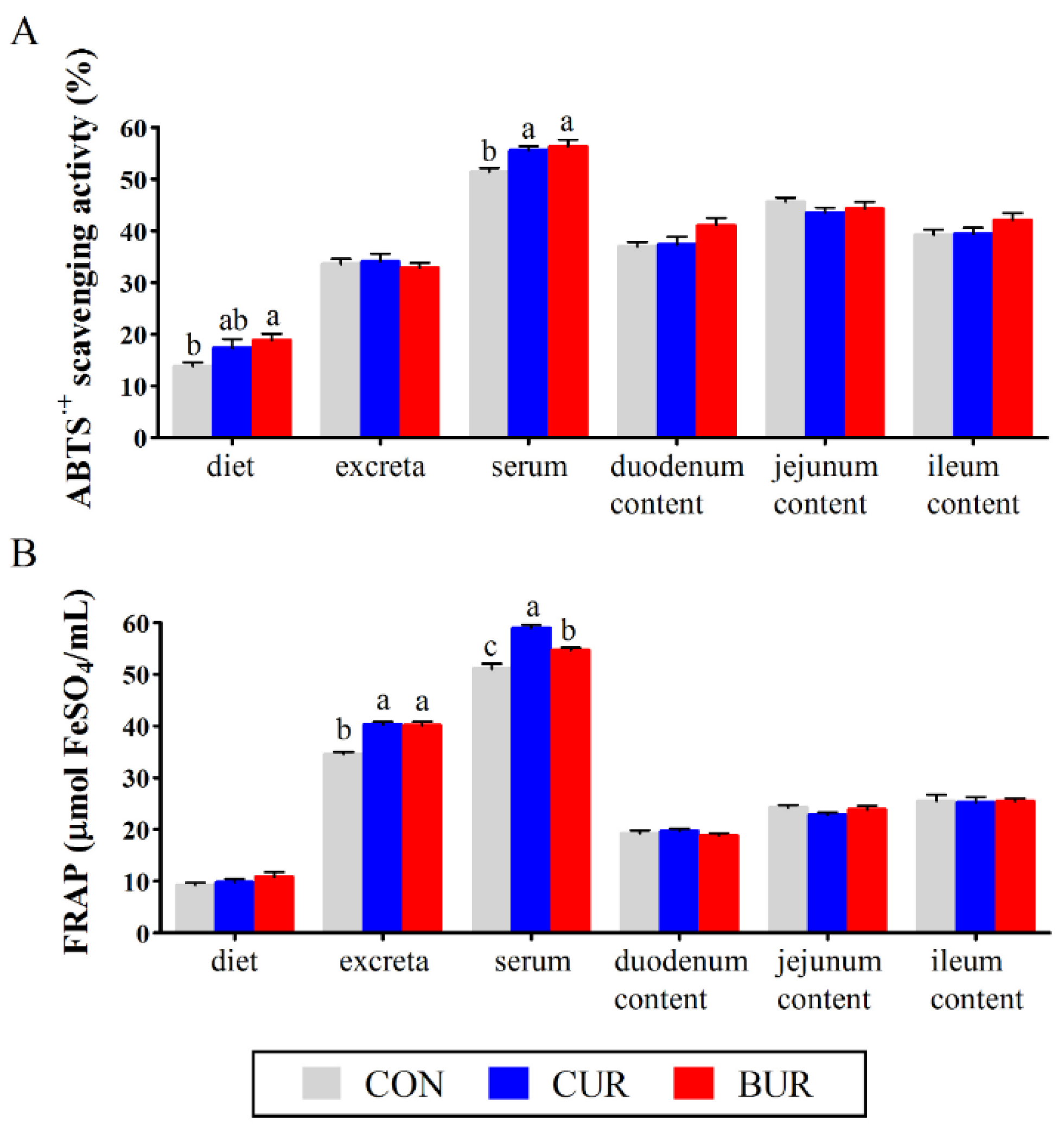
3.7. Antioxidant Activity in Diets, Excreta, Serum, and Small Intestine Contents
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Anand, P.; Thomas, S.G.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Sung, B.; Tharakan, S.T.; Misra, K.; Priyadarsini, I.K.; Rajasekharan, K.N. Biological activities of curcumin and its analogues (Congeners) made by man and Mother Nature. Biochem. Pharmacol. 2008, 76, 1590–1611. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. From exotic spice to modern drug? Cell 2007, 130, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Nardo, L.; Andreoni, A.; Masson, M.; Haukvik, T.; Tønnesen, H.H. Studies on Curcumin and Curcuminoids. XXXIX. Photophysical Properties of Bisdemethoxycurcumin. J. Fluoresc. 2011, 21, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.N. Curcuminoids as potent inhibitors of lipid peroxidation. J. Pharm. Pharmacol. 2011, 46, 1013–1016. [Google Scholar]
- Jayaprakasha, G.; Jaganmohan Rao, L.; Sakariah, K. Antioxidant activities of curcumin, demethoxycurcumin and bisdemethoxycurcumin. Food Chem. 2006, 98, 720–724. [Google Scholar] [CrossRef]
- Ahsan, H.; Parveen, N.; Khan, N.U.; Hadi, S. Pro-oxidant, anti-oxidant and cleavage activities on DNA of curcumin and its derivatives demethoxycurcumin and bisdemethoxycurcumin. Chem.-Biol. Interact. 1999, 121, 161–175. [Google Scholar] [CrossRef]
- Somparn, P.; Phisalaphong, C.; Nakornchai, S.; Unchern, S.; Morales, N.P. Comparative antioxidant activities of curcumin and its demethoxy and hydrogenated derivatives. Biol. Pharm. Bull. 2007, 30, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, C.; Zhao, S.; Yuan, D.; Lian, G.; Wang, X.; Wang, L.; Yang, J. Demethoxycurcumin, a natural derivative of curcumin attenuates LPS-induced pro-inflammatory responses through down-regulation of intracellular ROS-related MAPK/NF-κB signaling pathways in N9 microglia induced by lipopolysaccharide. Int. Immunopharmacol. 2010, 10, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Z.; Lu, C.; Bai, K.; Zhang, L.; Tian, W. Effect of various levels of dietary curcumin on meat quality and antioxidant profile of breast muscle in broilers. J. Agric. Food Chem. 2015, 63, 3880–3886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Hou, X.; Ahmad, H.; Zhang, H.; Zhang, L.L.; Wang, T. Assessment of free radicals scavenging activity of seven natural pigments and protective effects in AAPH-challenged chicken erythrocytes. Food Chem. 2014, 145, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, H.; Parvez, S.; Pasha, S.T.; Banerjee, B.D.; Raisuddin, S. Protective effect of lipoic acid against methotrexate-induced oxidative stress in liver mitochondria. Food Chem. Toxicol. 2010, 48, 1973–1979. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Kim, S.J.; Manchester, L.C.; Qi, W.; Garcia, J.J.; Cabrera, J.C.; El-Sokkary, G.; Rouvier-Garay, V. Augmentation of indices of oxidative damage in life-long melatonin-deficient rats. Mech. Ageing Dev. 1999, 110, 157–173. [Google Scholar] [CrossRef]
- Nkabyo, Y.S.; Gu, L.H.; Jones, D.P.; Ziegler, T.R. Thiol/disulfide redox status is oxidized in plasma and small intestinal and colonic mucosa of rats with inadequate sulfur amino acid intake. J. Nutr. 2006, 136, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Viveros, A.; Goni, I.; Centeno, C.; Sáyago-Ayerdy, S.; Arija, I.; Saura-Calixto, F. Effect of grape pomace concentrate and vitamin E on digestibility of polyphenols and antioxidant activity in chickens. Poult. Sci. 2008, 87, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Siriwan, P.; Bryden, W.L.; Mollah, Y.; Annison, E.F. Measurement of endogenous amino acid losses in poultry. Brit. Poult. Sci. 1993, 34, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.; Kikusato, M.; Maekawa, T.; Shirakawa, H.; Toyomizu, M. Metabolic characteristics and oxidative damage to skeletal muscle in broiler chickens exposed to chronic heat stress. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, K.W.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mitochondrial thioredoxin system in heat-stressed broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.M.; Jones, D.P. Redox control systems in the nucleus: Mechanisms and functions. Antioxid. Redox Signal. 2010, 13, 489–509. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Radical-free biology of oxidative stress. Am. J. Physiol. Cell Physiol. 2008, 295, C849–C868. [Google Scholar] [CrossRef] [PubMed]
- Stasiuk, M.; Kijanka, G.; Kozubek, A. Transformations of erythrocytes shape and its regulation. Postepy Biochem. 2009, 55, 425–433. [Google Scholar] [PubMed]
- Kumar, R.; Lal, N.; Nemaysh, V.; Luthra, P.M. Demethoxycurcumin mediated targeting of Mn-SOD leading to activation of apoptotic pathway and inhibition of Akt/NF-κB survival signalling in human glioma U87 MG cells. Toxicol. Appl. Pharmacol. 2018, 345, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ramirez, I.; Santamaria, A.; Tobon-Velasco, J.C.; Orozco-Ibarra, M.; Gonzalez-Herrera, I.G.; Pedraza-Chaverri, J.; Maldonado, P.D. Curcumin restores Nrf2 levels and prevents quinolinic acid-induced neurotoxicity. J. Nutr. Biochem. 2013, 24, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Amichand, D.; Limson, J.L.; Watkins, G.M.; Edith, A.; Santy, D. Curcuminoids, curcumin, and demethoxycurcumin reduce lead-induced memory deficits in male Wistar rats. J. Agric. Food Chem. 2007, 55, 1039–1044. [Google Scholar]
- Zhang, J.F.; Hu, Z.P.; Lu, C.H.; Yang, M.X.; Zhang, L.L.; Wang, T. Dietary curcumin supplementation protects against heat-stress-impaired growth performance of broilers possibly through a mitochondrial pathway. J. Anim. Sci. 2015, 93, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.N.; Lee, Y.J.; Song, J.H.; Jang, H.D. Curcumin Ameliorates AAPH-induced Oxidative Stress in HepG2 Cells by Activating Nrf2. Food Sci. Biotechnol. 2013, 22, 241–247. [Google Scholar] [CrossRef]
- Anna, K.; Wookwang, J.; Jungjun, L.; Byungchul, K. Up-regulation of heme oxygenase-1 expression through CaMKII-ERK1/2-Nrf2 signaling mediates the anti-inflammatory effect of bisdemethoxycurcumin in LPS-stimulated macrophages. Free Radic. Biol. Med. 2010, 49, 323–331. [Google Scholar]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Vomhof-Dekrey, E.E.; Picklo, M.J., Sr. The Nrf2-antioxidant response element pathway: A target for regulating energy metabolism. J. Nutr. Biochem. 2012, 23, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Pugazhenthi, S.; Akhov, L.G.; Wang, M.; Alam, J. Regulation of heme oxygenase-1 expression by demethoxy curcuminoids through Nrf2 by a PI3-kinase/Akt-mediated pathway in mouse beta-cells. Am. J. Physiol. 2007, 293, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Antioxidant function of thioredoxin and glutaredoxin systems. Antioxid. Redox Signal. 2000, 2, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Uys, J.D.; Mulholland, P.J.; Townsend, D.M. Glutathione and redox signaling in substance abuse. Biomed. Pharmacother. 2014, 68, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in redox homeostasis. J. Free Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Bray, T.M. Dietary antioxidants and assessment of oxidative stress. J. Nutr. 2000, 16, 578–581. [Google Scholar] [CrossRef]
- Salami, S.A.; Majoka, M.A.; Saha, S.; Garber, A.; Gabarrou, J.F. Efficacy of dietary antioxidants on broiler oxidative stress, performance and meat quality: Science and market. Avian Biol. Res. 2015, 8, 65–78. [Google Scholar] [CrossRef]
- Singhal, S.S.; Awasthi, S.; Pandya, U.; Piper, J.T.; Saini, M.K.; Cheng, J.Z.; Awasthi, Y.C. The effect of curcumin on glutathione-linked enzymes in K562 human leukemia cells. Toxicol. Lett. 1999, 109, 87–95. [Google Scholar] [CrossRef]
- Zheng, S.; Yumei, F.; Chen, A. De novo synthesis of glutathione is a prerequisite for curcumin to inhibit hepatic stellate cell (HSC) activation. Free Radic. Biol. Med. 2007, 43, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Guo, C.; Yang, J.; Wei, J.; Li, Y.; Xu, J.; Jiang, Y. Antioxidant activities of peel, pulp and seed fractions of common fruits as determined by FRAP assay. Nutr. Res. Rev. 2003, 23, 1719–1726. [Google Scholar] [CrossRef]
- Holder, G.M.; Plummer, J.L.; Ryan, A.J. The metabolism and excretion of curcumin (1, 7-bis-(4-hydroxy-3-methoxyphenyl)-1, 6-heptadiene-3, 5-dione) in the rat. Xenobiotica 1978, 8, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin—From molecule to biological function. Angew. Chem. Int. Ed. Engl. 2012, 51, 5308–5332. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, T.; Satoh, K.; Kadoma, Y.; Murakami, Y.; Unten, S.; Atsumi, T.; Sakagami, H.; Fujisawa, S. Radical scavenging activity and cytotoxicity of ferulic acid. Anticancer Res. 2002, 22, 2711–2717. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items | 1–21 Days | 22–42 Days |
|---|---|---|
| Ingredient (%) | ||
| Corn | 57.0 | 61.9 |
| Soybean meal (44.2%, crude protein) | 31.3 | 25.6 |
| Corn gluten meal (60%, crude protein) | 3.9 | 4.3 |
| Soybean oil | 3.1 | 3.8 |
| Dicalcium phosphate | 1.8 | 1.6 |
| Limestone | 1.3 | 1.2 |
| L-lysine | 0.15 | 0.2 |
| DL-methionine | 0.15 | 0.1 |
| Premix 1 | 1 | 1 |
| Salt | 0.3 | 0.3 |
| Total | 100 | 100 |
| Calculation of nutrients | ||
| Metabolizable energy, MJ/kg | 12.69 | 13.10 |
| Crude protein, % | 21.52 | 19.71 |
| lysine, % | 1.14 | 1.04 |
| methionine, % | 0.50 | 0.43 |
| Calcium, % | 1.00 | 0.90 |
| Available phosphorus, % | 0.46 | 0.42 |
| Arginine, % | 1.36 | 1.19 |
| Methionine+Cystine, % | 0.85 | 0.76 |
| Genbank ID | Gene Name 1 | Sequence (5′→3′) 1 | Product Length |
|---|---|---|---|
| NM_205518 | β-Actin | forward: TGCTGTGTTCCCATCTATCG | 150 |
| reverse: TTGGTGACAATACCGTGTTCA | |||
| NM_001031215.1 | CAT | forward: GGTTCGGTGGGGTTGTCTTT | 211 |
| reverse: CACCAGTGGTCAAGGCATCT | |||
| NM_205064.1 | CuZnSOD | forward: CCGGCTTGTCTGATGGAGAT | 124 |
| reverse: TGCATCTTTTGGTCCACCGT | |||
| NM_001277853.1 | GPx | forward: GACCAACCCGCAGTACATCA | 205 |
| reverse: GAGGTGCGGGCTTTCCTTTA | |||
| HM237181.1 | HO-1 | forward: GGTCCCGAATGAATGCCCTTG | 138 |
| reverse: ACCGTTCTCCTGGCTCTTGG | |||
| NM_205117.1 | Nrf2 | forward: GATGTCACCCTGCCCTTAG | 215 |
| reverse: CTGCCACCATGTTATTCC |
| Items | CON 2 | CUR | BUR | p-Value | |
|---|---|---|---|---|---|
| 1-21 d | ADG, g/bird per day | 34.85 ± 0.68 | 35.67 ± 0.64 | 35.41 ± 0.30 | 0.586 |
| ADFI, g/bird per day | 49.78 ± 0.66 | 49.73 ± 0.93 | 48.89 ± 0.48 | 0.617 | |
| FCR | 1.43 ± 0.02 | 1.39 ± 0.02 | 1.38 ± 0.01 | 0.133 | |
| 21-42 d | ADG, g/bird per day | 89.93 ± 1.570 b | 96.79 ± 1.55 a | 93.35 ± 2.25 ab | 0.047 |
| ADFI, g/bird per day | 164.31 ± 2.770 | 173.80 ± 2.88 | 167.81 ± 4.65 | 0.184 | |
| FCR | 1.83 ± 0.020 | 1.80 ± 0.03 | 1.80 ± 0.03 | 0.607 | |
| 1-42 d | ADG, g/bird per day | 59.42 ± 0.90 b | 63.08 ± 0.96 a | 61.31 ± 1.02 ab | 0.044 |
| ADFI, g/bird per day | 100.09 ± 1.44 | 104.24 ± 1.68 | 100.95 ± 2.00 | 0.221 | |
| FCR | 1.69 ± 0.02 | 1.65 ± 0.02 | 1.65 ± 0.01 | 0.278 | |
| Items | CON 2 | CUR | BUR | p-Value |
|---|---|---|---|---|
| CAT, U/mL | 1.71 ± 0.24 | 1.95 ± 0.31 | 1.94 ± 0.45 | 0.854 |
| T-SOD, U/mL | 182.23 ± 12.05 | 202.78 ± 31.78 | 191.16 ± 12.88 | 0.788 |
| GPx, U/mL | 223.64 ± 12.71 | 231.71 ± 6.43 | 239.65 ± 16.59 | 0.674 |
| T-AOC, U/mL | 7.38 ± 0.67 b | 9.48 ± 0.45 ab | 10.15 ± 0.77 a | 0.016 |
| T-SH, µmol/L | 136.72 ± 11.13 | 139.94 ± 16.21 | 144.76 ± 10.25 | 0.905 |
| MDA, nmol/mL | 1.94 ± 0.08 a | 1.63 ± 0.08 b | 1.78 ± 0.05 ab | 0.018 |
| PC, nmol/mg prot | 1.11 ± 0.11 | 1.30 ± 0.17 | 1.14 ± 0.17 | 0.649 |
| Items 3 | CON 2 | CUR | BUR | p-Value |
|---|---|---|---|---|
| Glutathione redox potential Eh, mV | ||||
| serum | −81.91 ± 4.95 | −90.57 ± 4.15 | −87.49 ± 8.30 | 0.600 |
| liver | −141.43 ± 1.05 | −145.05 ± 1.61 | −146.28 ± 1.45 | 0.057 |
| duodenum | −121.46 ± 3.88 | −122.79 ± 4.97 | −127.23 ± 2.47 | 0.560 |
| jejunum | −108.23 ± 3.57 a | −121.79 ± 2.15 b | −124.36 ± 0.81 b | <0.001 |
| ileum | −126.28 ± 1.40 a | −131.02 ± 2.60 ab | −134.86 ± 3.08 b | 0.060 |
| r-GSH, μmol/L | ||||
| serum | 1.04 ± 0.10 | 1.34 ± 0.12 | 1.33 ± 0.24 | 0.354 |
| liver | 124.41 ± 3.47 b | 135.46 ± 5.32 ab | 141.99 ± 4.96 a | 0.044 |
| duodenum | 77.44 ± 10.30 | 78.20 ± 10.36 | 83.60 ± 5.77 | 0.872 |
| jejunum | 58.14 ± 7.08 b | 80.39 ± 4.85 a | 91.47 ± 3.17 a | 0.001 |
| ileum | 103.20 ± 5.15 | 120.45 ± 8.27 | 131.47 ± 11.46 | 0.081 |
| o-GSH, μmol/L | ||||
| serum | 0.28 ± 0.05 | 0.23 ± 0.03 | 0.24 ± 0.05 | 0.672 |
| liver | 40.62 ± 1.60 | 36.50 ± 2.03 | 36.48 ± 1.74 | 0.196 |
| duodenum | 64.80 ± 2.68 | 57.69 ± 4.25 | 54.16 ± 4.47 | 0.167 |
| jejunum | 101.91 ± 3.06 a | 75.68 ± 4.97 b | 81.12 ± 2.92 b | <0.001 |
| ileum | 88.04 ± 2.04 | 83.27 ± 5.49 | 72.97 ± 5.73 | 0.060 |
| Items | CON 2 | CUR | BUR |
|---|---|---|---|
| Liver, % | |||
| CAT3 | 100.00 ± 8.895 | 111.88 ± 8.681 | 113.85 ± 11.346 |
| HO-1 | 100.00 ± 9.972 | 121.59 ± 6.569 | 129.41 ± 13.854 |
| GPx | 100.00 ± 9.930 | 112.21 ± 8.063 | 120.34 ± 7.935 |
| CuZnSOD | 100.00 ± 9.643 | 97.89 ± 4.701 | 110.61 ± 6.515 |
| Nrf2 | 100.00 ± 4.032 b | 118.31 ± 7.744 ab | 121.32 ± 3.780 a |
| jejunum, % | |||
| CAT | 100.00 ± 5.808 | 132.07 ± 8.004 | 144.11 ± 20.052 |
| HO-1 | 100.00 ± 7.778 b | 139.24 ± 20.163 ab | 155.82 ± 15.376 a |
| GPx | 100.00 ± 12.405 | 127.60 ± 20.735 | 101.17 ± 7.485 |
| CuZnSOD | 100.00 ± 8.584 | 96.88 ± 6.349 | 124.40 ± 12.215 |
| Nrf2 | 100.00 ± 7.663 | 105.65 ± 9.953 | 110.16 ± 8.740 |
| ileum, % | |||
| CAT | 100.00 ± 8.136 | 97.93 ± 4.696 | 101.66 ± 4.998 |
| HO-1 | 100.00 ± 6.306 | 99.76 ± 6.640 | 100.62 ± 9.090 |
| GPx | 100.00 ± 5.003 | 96.71 ± 3.639 | 98.62 ± 5.586 |
| CuZnSOD | 100.00 ± 8.776 b | 161.82 ± 11.127 a | 141.22 ± 9.167 a |
| Nrf2 | 100.00 ± 5.907 b | 122.06 ± 6.876 a | 117.82 ± 5.374 ab |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Han, H.; Shen, M.; Zhang, L.; Wang, T. Comparative Studies on the Antioxidant Profiles of Curcumin and Bisdemethoxycurcumin in Erythrocytes and Broiler Chickens. Animals 2019, 9, 953. https://doi.org/10.3390/ani9110953
Zhang J, Han H, Shen M, Zhang L, Wang T. Comparative Studies on the Antioxidant Profiles of Curcumin and Bisdemethoxycurcumin in Erythrocytes and Broiler Chickens. Animals. 2019; 9(11):953. https://doi.org/10.3390/ani9110953
Chicago/Turabian StyleZhang, Jingfei, Hongli Han, Mingming Shen, Lili Zhang, and Tian Wang. 2019. "Comparative Studies on the Antioxidant Profiles of Curcumin and Bisdemethoxycurcumin in Erythrocytes and Broiler Chickens" Animals 9, no. 11: 953. https://doi.org/10.3390/ani9110953
APA StyleZhang, J., Han, H., Shen, M., Zhang, L., & Wang, T. (2019). Comparative Studies on the Antioxidant Profiles of Curcumin and Bisdemethoxycurcumin in Erythrocytes and Broiler Chickens. Animals, 9(11), 953. https://doi.org/10.3390/ani9110953