
Serum Norepinephrine and Cholesterol Concentrations as Novel Diagnostic Biomarkers for Vitamin E Deficiency in Holstein Cows


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Blood Collection
2.3. Serum Detection
2.4. Sample Preparation
2.5. GC-TOF-MS Analysis
2.6. Data Preprocessing
2.7. Multivariate Statistical Analysis
2.8. Metabolite Identification and Pathway Analysis
2.9. Targeted Metabolomics Assays
2.10. Statistical Analysis
3. Results
3.1. Background Attributes and Serum Biochemical Profiles
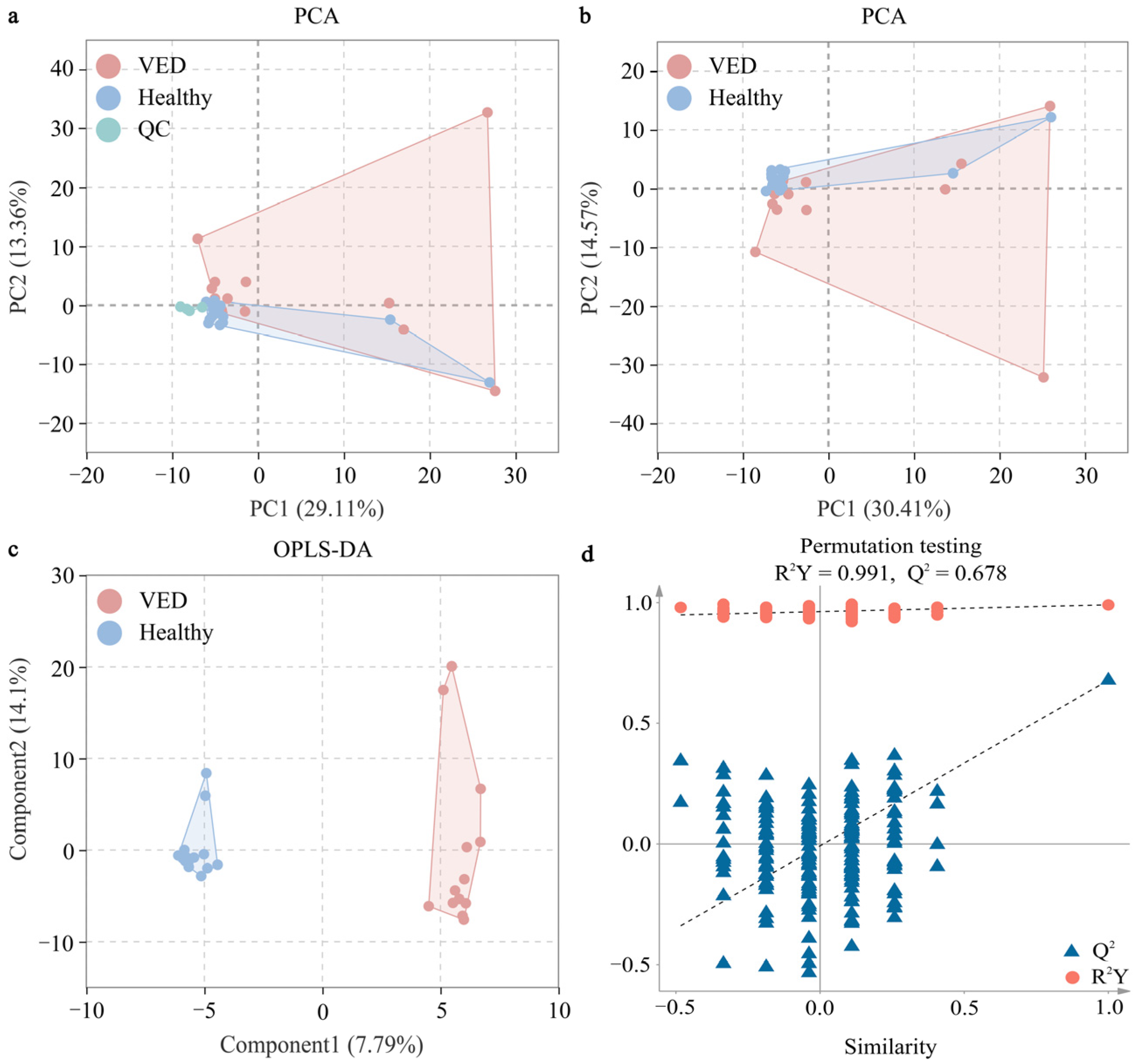
3.2. DMs Identified by Untargeted GC-TOF-MS
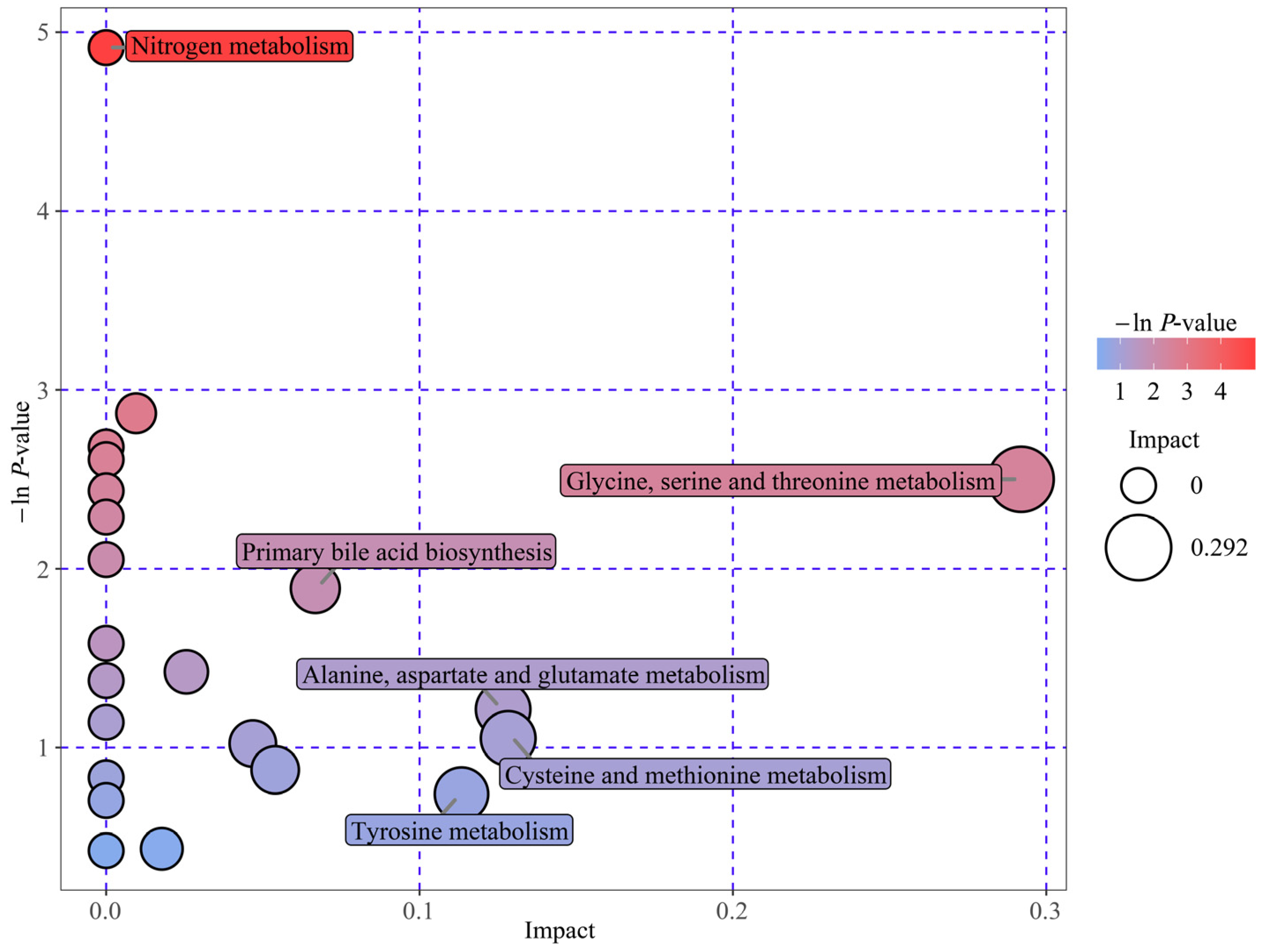
3.3. Analysis and Identification of Key Metabolic Pathways
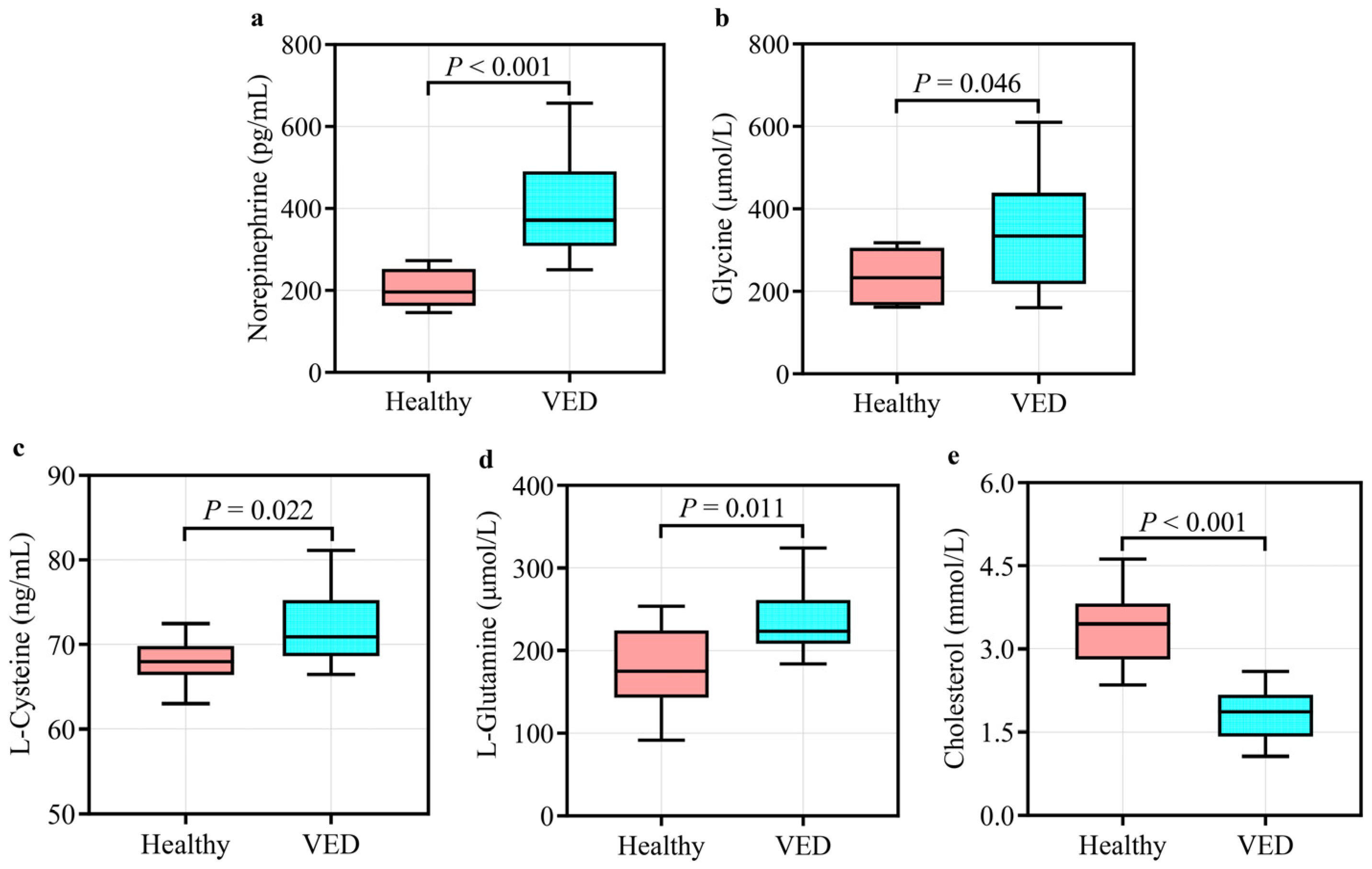
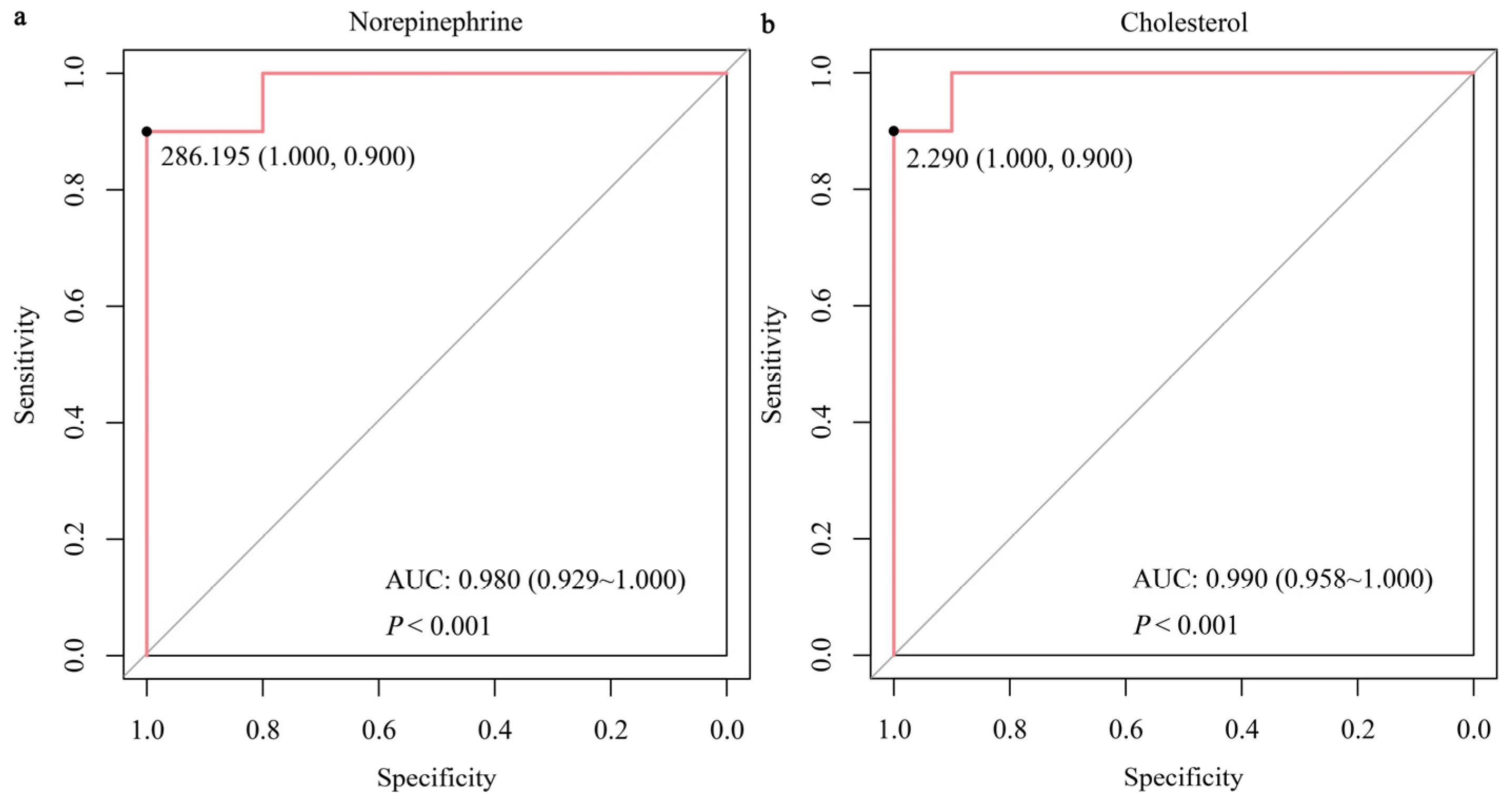
3.4. Validation of the Important DMs and Screening for Novel Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beeckman, A.; Vicca, J.; Van Ranst, G.; Janssens, G.P.; Fievez, V. Monitoring of vitamin E status of dry, early and mid-late lactating organic dairy cows fed conserved roughages during the indoor period and factors influencing forage vitamin E levels. J. Anim. Physiol. Anim. Nutr. 2010, 94, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.P. A 100-Year Review: From ascorbic acid to zinc—Mineral and vitamin nutrition of dairy cows. J. Dairy Sci. 2017, 100, 10045–10060. [Google Scholar] [CrossRef]
- Haga, S.; Ishizaki, H.; Roh, S. The Physiological Roles of Vitamin E and Hypovitaminosis E in the Transition Period of High-Yielding Dairy Cows. Animals 2021, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- Moghimi-Kandelousi, M.; Alamouti, A.A.; Imani, M.; Zebeli, Q. A meta-analysis and meta-regression of the effects of vitamin E supplementation on serum enrichment, udder health, milk yield, and reproductive performance of transition cows. J. Dairy Sci. 2020, 103, 6157–6166. [Google Scholar] [CrossRef]
- Kuhn, M.J.; Sordillo, L.M. Vitamin E analogs limit in vitro oxidant damage to bovine mammary endothelial cells. J. Dairy Sci. 2021, 104, 7154–7167. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.M.; Wisnieski, L.; Herdt, T.H.; Sordillo, L.M. Serum retinol, β-carotene, and α-tocopherol as biomarkers for disease risk and milk production in periparturient dairy cows. J. Dairy Sci. 2021, 104, 915–927. [Google Scholar] [CrossRef]
- Quigley, J.D.; Hill, T.M.; Dennis, T.S.; Suarez-Mena, F.X.; Hu, W.; Kahl, S.; Elsasser, T.H. Effects of mixed tocopherols added to milk replacer and calf starter on intake, growth, and indices of stress. J. Dairy Sci. 2021, 104, 9769–9783. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Qian, W.; Yu, H.; Zhang, C.; Zhang, H.; Fu, S.; Xia, C. plasma proteomics characteristics of subclinical vitamin e deficiency of dairy cows during early lactation. Front. Vet. Sci. 2021, 8, 723898. [Google Scholar] [CrossRef]
- Song, Y.; Wang, H.; Sun, R.; Chang, J.; Tang, J.; Bai, Y.; Xia, C. Serum metabolic characterization of vitamin E deficiency in Holstein cows during the transition period based on proton nuclear magnetic resonance spectroscopy. Animals 2023, 13, 2957. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, L.; Xu, C.; Xia, C.; Sun, L.; Shu, S. Plasma metabolomic profiling of dairy cows affected with ketosis using gas chromatography/mass spectrometry. BMC Vet. Res. 2013, 9, 186. [Google Scholar] [CrossRef]
- Liu, L.; Shen, T.; Yang, W.; Yu, H.; Gao, S.; Huang, B.; Xu, C. Ketotic cows display a different serum nonesterified fatty acid composition. J. Dairy Res. 2020, 87, 52–55. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC-MS metabolomics identifies metabolite alterations that precede subclinical mastitis in the blood of transition dairy cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, H.; Zhang, Y.; Xiong, B.; Jiang, L. Microbiome and metabolome analyses of milk from dairy cows with subclinical streptococcus agalactiae mastitis-potential biomarkers. Front. Microbiol. 2019, 10, 2547. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Song, Y.; Zhang, J.; Fu, S.; Wu, L.; Xia, C.; Xu, C. GC/MS and LC/MS based serum metabolomic analysis of dairy cows with ovarian inactivity. Front. Vet. Sci. 2021, 8, 678388. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Hu, P.; Bai, Y.L.; Xia, C. Plasma metabolic differences in cows affected by inactive ovaries or normal ovarian function post partum. Pol. J. Vet. Sci. 2020, 23, 59–67. [Google Scholar] [CrossRef]
- Figueiredo, C.C.; Balzano-Nogueira, L.; Bisinotto, D.Z.; Ruiz, A.R.; Duarte, G.A.; Conesa, A.; Galvão, K.N.; Bisinotto, R.S. Differences in uterine and serum metabolome associated with metritis in dairy cows. J. Dairy Sci. 2023, 106, 3525–3536. [Google Scholar] [CrossRef]
- Casaro, S.; Prim, J.G.; Gonzalez, T.D.; Figueiredo, C.C.; Bisinotto, R.S.; Chebel, R.C.; Santos, J.E.P.; Nelson, C.D.; Jeon, S.J.; Bicalho, R.C.; et al. Blood metabolomics and impacted cellular mechanisms during transition into lactation in dairy cows that develop metritis. J. Dairy Sci. 2023, 106, 8098–8109. [Google Scholar] [CrossRef]
- de Oliveira, E.B.; Monteiro, H.F.; Pereira, J.M.V.; Williams, D.R.; Pereira, R.V.; Silva Del Rio, N.; Menta, P.R.; Machado, V.S.; Lima, F.S. Changes in uterine metabolome associated with metritis development and cure in lactating Holstein cows. Metabolites 2023, 13, 1156. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.M.G.; Cunha, A.L.R.R.; David, L.C.; Camelo, Í.N.; Martins, N.M.; Galvão, F.S.; Mendonça, D.R.; Venâncio, M.T.; Cunha, R.D.S.; Filho, A.R.C.; et al. A veterinary cerumenomic assay for bovine laminitis identification. Vet. Res. Commun. 2024, 48, 1003–1013. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Serum metabolic fingerprinting of pre-lameness dairy cows by GC-MS reveals typical profiles that can identify susceptible cows. J. Proteom. 2020, 213, 103620. [Google Scholar] [CrossRef]
- Kim, T.J.; Park, J.G.; Ahn, S.K.; Kim, K.W.; Choi, J.; Kim, H.Y.; Ha, S.H.; Seo, W.D.; Kim, J.K. Discrimination of adzuki bean (Vigna angularis) geographical origin by targeted and non-targeted metabolite profiling with gas chromatography time-of-flight mass spectrometry. Metabolites 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jiang, D.; Wang, C.; An, G.; Zhu, L.; Cui, C. Comprehensive analysis of metabolic changes in male mice exposed to sodium valproate based on GC-MS analysis. Drug Des. Dev. Ther. 2022, 16, 1915–1930. [Google Scholar] [CrossRef] [PubMed]
- Putman, A.K.; Brown, J.L.; Gandy, J.C.; Wisnieski, L.; Sordillo, L.M. Changes in biomarkers of nutrient metabolism, inflammation, and oxidative stress in dairy cows during the transition into the early dry period. J. Dairy Sci. 2018, 101, 9350–9359. [Google Scholar] [CrossRef]
- Edmonson, A.J.; Lean, I.J.; Weaver, L.D.; Farver, T.; Webster, G. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 1989, 72, 68–78. [Google Scholar] [CrossRef]
- Siomek, A.; Gackowski, D.; Rozalski, R.; Dziaman, T.; Szpila, A.; Guz, J.; Olinski, R. Higher leukocyte 8-oxo-7,8-dihydro-2′-deoxyguanosine and lower plasma ascorbate in aging humans? Antioxid. Redox Signal. 2007, 9, 143–150. [Google Scholar] [CrossRef]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Luo, Z.Z.; Shen, L.H.; Jiang, J.; Huang, Y.X.; Bai, L.P.; Yu, S.M.; Yao, X.P.; Ren, Z.H.; Yang, Y.X.; Cao, S.Z. Plasma metabolite changes in dairy cows during parturition identified using untargeted metabolomics. J. Dairy Sci. 2019, 102, 4639–4650. [Google Scholar] [CrossRef]
- Basoglu, A.; Baspinar, N.; Tenori, L.; Licari, C.; Gulersoy, E. Nuclear magnetic resonance (NMR)-based metabolome profile evaluation in dairy cows with and without displaced abomasum. Vet. Q. 2020, 40, 1–15. [Google Scholar] [CrossRef]
- Wathes, D.C.; Fenwick, M.; Cheng, Z.; Bourne, N.; Llewellyn, S.; Morris, D.G.; Kenny, D.; Murphy, J.; Fitzpatrick, R. Influence of negative energy balance on cyclicity and fertility in the high producing dairy cow. Theriogenology 2007, 68, S232–S241. [Google Scholar] [CrossRef]
- Daros, R.R.; Havekes, C.D.; DeVries, T.J. Body condition loss during the dry period: Insights from feeding behavior studies. J. Dairy Sci. 2021, 104, 4682–4691. [Google Scholar] [CrossRef]
- Haga, S.; Miyaji, M.; Nakano, M.; Ishizaki, H.; Matsuyama, H.; Katoh, K.; Roh, S. Changes in the expression of α-tocopherol-related genes in liver and mammary gland biopsy specimens of peripartum dairy cows. J. Dairy Sci. 2018, 101, 5277–5293. [Google Scholar] [CrossRef] [PubMed]
- Macrae, A.I.; Burrough, E.; Forrest, J.; Corbishley, A.; Russell, G.; Shaw, D.J. Prevalence of excessive negative energy balance in commercial United Kingdom dairy herds. Vet. J. 2019, 248, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.G.; Waters, S.M.; McCarthy, S.D.; Patton, J.; Earley, B.; Fitzpatrick, R.; Murphy, J.J.; Diskin, M.G.; Kenny, D.A.; Brass, A.; et al. Pleiotropic effects of negative energy balance in the postpartum dairy cow on splenic gene expression: Repercussions for innate and adaptive immunity. Physiol. Genom. 2009, 39, 28–37. [Google Scholar] [CrossRef]
- Ghavipanje, N.; Fathi Nasri, M.H.; Farhangfar, S.H.; Ghiasi, S.E.; Vargas-Bello-Pérez, E. The impact of dietary berberine supplementation during the transition period on blood parameters, antioxidant indicators and fatty acids profile in colostrum and milk of dairy goats. Vet. Sci. 2022, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.W.; Di Lucente, J.; Ruiz Mendiola, U.; Suthprasertporn, N.; Tomilov, A.; Cortopassi, G.; Kim, K.; Ramsey, J.J.; Maezawa, I. The ketone body β-hydroxybutyrate shifts microglial metabolism and suppresses amyloid-β oligomer-induced inflammation in human microglia. FASEB J. 2023, 37, e23261. [Google Scholar] [CrossRef]
- Ore, A.; Olayinka, E.T. Influence of moxifloxacin on hepatic redox status and plasma biomarkers of hepatotoxicity and nephrotoxicity in rat. Biochem. Res. Int. 2015, 2015, 192724. [Google Scholar] [CrossRef]
- Zhang, J.; Head, B.; Leonard, S.W.; Choi, J.; Tanguay, R.L.; Traber, M.G. Vitamin E deficiency dysregulates thiols, amino acids and related molecules during zebrafish embryogenesis. Redox Biol. 2021, 38, 101784. [Google Scholar] [CrossRef]
- Carracedo, J.; Ramírez-Carracedo, R.; Martínez de Toda, I.; Vida, C.; Alique, M.; De la Fuente, M.; Ramírez-Chamond, R. Protein carbamylation: A marker reflecting increased age-related cell oxidation. Int. J. Mol. Sci. 2018, 19, 1495. [Google Scholar] [CrossRef]
- He, B.; Li, H.; Hu, M.; Dong, W.; Wei, Z.; Li, J.; Yao, W.; Guo, X. Association between serum interleukin-17A level and high-altitude deacclimatization syndrome. Mediat. Inflamm. 2016, 2016, 1732352. [Google Scholar] [CrossRef]
- Zhou, X.Q.; Yao, Z.W.; Peng, Y.; Mao, S.S.; Xu, D.; Qin, X.F.; Zhang, R.J. PQQ ameliorates D-galactose induced cognitive impairments by reducing glutamate neurotoxicity via the GSK-3β/Akt signaling pathway in mouse. Sci. Rep. 2018, 8, 8894. [Google Scholar] [CrossRef]
- Chen, Y.W.; Das, M.; Oyarzabal, E.A.; Cheng, Q.; Plummer, N.W.; Smith, K.G.; Jones, G.K.; Malawsky, D.; Yakel, J.L.; Shih, Y.I.; et al. Genetic identification of a population of noradrenergic neurons implicated in attenuation of stress-related responses. Mol. Psychiatry 2019, 24, 710–725. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Zhong, H.; Yanamadala, S.; Campese, V.M. Oxidative stress mediates the stimulation of sympathetic nerve activity in the phenol renal injury model of hypertension. Hypertension 2006, 48, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Egger, F.; Jakab, M.; Fuchs, J.; Oberascher, K.; Brachtl, G.; Ritter, M.; Kerschbaum, H.H.; Gaisberger, M. Effect of glycine on BV-2 microglial cells treated with interferon-γ and lipopolysaccharide. Int. J. Mol. Sci. 2020, 21, 804. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Acosta, O.L. Glycine increases insulin sensitivity and glutathione biosynthesis and protects against oxidative stress in a model of sucrose-induced insulin resistance. Oxid. Med. Cell. Longev. 2018, 2018, 2101562. [Google Scholar]
- Yin, J.; Ren, W.; Yang, G.; Duan, J.; Huang, X.; Fang, R.; Li, C.; Li, T.; Yin, Y.; Hou, Y.; et al. L-Cysteine metabolism and its nutritional implications. Mol. Nutr. Food Res. 2016, 60, 134–146. [Google Scholar] [CrossRef]
- Haisa, M. Glutamine is a key regulator for amino acid-controlled cell growth through the mTOR signaling pathway in rat intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2005, 326, 174–180. [Google Scholar]
- Altman, B.J.; Stine, Z.E.; Dang, C.V. From Krebs to clinic: Glutamine metabolism to cancer therapy. Nat. Rev. Cancer 2016, 16, 619–634. [Google Scholar] [CrossRef]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef]
- Fukui, K.; Nakamura, K.; Shirai, M.; Hirano, A.; Takatsu, H.; Urano, S. Long-term vitamin E-deficient mice exhibit cognitive dysfunction via elevation of brain oxidation. J. Nutr. Sci. Vitaminol. 2015, 61, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Hartl, J.; Kiefer, P.; Kaczmarczyk, A.; Mittelviefhaus, M.; Meyer, F.; Vonderach, T.; Hattendorf, B.; Jenal, U.; Vorholt, J.A. Untargeted metabolomics links glutathione to bacterial cell cycle progression. Nat. Metab. 2020, 2, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Kohler, I.; Hankemeier, T.; van der Graaf, P.H.; Knibbe, C.A.J.; van Hasselt, J.G.C. Integrating clinical metabolomics-based biomarker discovery and clinical pharmacology to enable precision medicine. Eur. J. Pharm. Sci. 2017, 109S, S15–S21. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.Y.; Tan, J.; Li, L.X.; Wang, Y.; Liu, M.; Jiang, L.S.; Zhao, Y.C. Longitudinal characterization of serum metabolome and lipidome reveals that the ceramide profile is associated with metabolic health in early postpartum cows experiencing different lipolysis. J. Dairy Sci. 2024, 107, 7446–7468. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Healthy (n = 14) | VED (n = 13) | p-Value |
|---|---|---|---|
| Age | 3.55 ± 0.95 | 3.79 ± 0.81 | 0.488 |
| Parity | 2.73 ± 0.88 | 2.60 ± 1.05 | 0.730 |
| BCS | 3.43 ± 0.41 | 3.06 ± 0.31 | 0.014 |
| Milk yield (kg/d) | 38.37 ± 7.97 | 36.29 ± 7.66 | 0.496 |
| DMI (kg/d) | 16.52 ± 0.37 | 15.46 ± 0.72 | <0.001 |
| α-Tocopherol (μg/mL) | 6.73 ± 0.84 | 2.12 ± 0.66 | <0.001 |
| NEFA (mmol/L) | 0.65 ± 0.18 | 0.79 ± 0.22 | 0.081 |
| BHB (mmol/L) | 0.98 ± 0.36 | 1.23 ± 0.29 | 0.059 |
| Glucose (mmol/L) | 7.67 ± 2.03 | 6.42 ± 2.05 | 0.124 |
| T-AOC (mmol/L) | 0.38 ± 0.05 | 0.29 ± 0.04 | <0.001 |
| SOD (U/mL) | 153.09 ± 16.24 | 145.06 ± 20.68 | 0.271 |
| Catalase (U/mL) | 20.98 ± 1.34 | 22.02 ± 1.49 | 0.068 |
| GSH-Px (U/mL) | 128.55 ± 16.54 | 119.42 ± 37.28 | 0.425 |
| GSH (μmol/L) | 83.22 ± 20.86 | 61.84 ± 23.82 | 0.020 |
| MDA (mmol/mL) | 2.67 ± 0.67 | 3.98 ± 0.72 | <0.001 |
| •OH (U/mL) | 613.64 ± 69.81 | 653.12 ± 50.09 | 0.106 |
| No. | KEGG ID | Metabolites | RT (min) | VIP | p-Value | FC | VED vs. Healthy |
|---|---|---|---|---|---|---|---|
| 1 | C00547 | Noradrenaline | 12.01 | 1.60 | 0.004 | 4.30 | Up |
| 2 | C00178 | Thymine | 7.84 | 1.15 | <0.001 | 4.26 | Up |
| 3 | C00584 | Prostaglandin E2 | 14.78 | 1.88 | 0.020 | 4.18 | Up |
| 4 | C07326 | 1,5-Anhydroglucitol | 10.48 | 2.46 | 0.006 | 3.26 | Up |
| 5 | C00037 | Glycine | 7.19 | 2.85 | <0.001 | 2.97 | Up |
| 6 | C00451 | D-threo-Isocitric acid | 10.38 | 1.76 | 0.009 | 2.93 | Up |
| 7 | — | Diglycerol | 10.05 | 2.06 | 0.014 | 2.50 | Up |
| 8 | C00476 | Lyxose | 9.48 | 1.87 | 0.013 | 2.37 | Up |
| 9 | C06555 | Biuret | 10.04 | 2.64 | <0.001 | 2.31 | Up |
| 10 | C01712 | Elaidic acid | 12.30 | 1.86 | 0.033 | 1.97 | Up |
| 11 | C01733 | Racemethionine | 8.61 | 1.57 | 0.013 | 1.87 | Up |
| 12 | C00805 | Salicylic acid | 8.58 | 1.21 | 0.016 | 1.83 | Up |
| 13 | C00085 | D-Fructose-6-phosphate | 12.71 | 2.22 | 0.015 | 1.74 | Up |
| 14 | C00097 | L-Cysteine | 8.81 | 1.61 | 0.009 | 1.62 | Up |
| 15 | C02057 | Phenylalanine | 9.29 | 1.60 | 0.034 | 1.49 | Up |
| 16 | C01877 | 4-Oxoproline | 8.65 | 1.72 | 0.030 | 1.43 | Up |
| 17 | C01073 | N-Acetyl-beta-alanine | 7.82 | 1.50 | 0.029 | 1.38 | Up |
| 18 | C01571 | Capric acid | 8.23 | 1.61 | 0.028 | 1.37 | Up |
| 19 | — | Adipamide | 9.56 | 1.49 | 0.039 | 1.36 | Up |
| 20 | C00064 | L-Glutamine | 8.74 | 1.48 | 0.040 | 1.34 | Up |
| 21 | C00093 | D-Glycerol 1-phosphate | 9.96 | 2.03 | 0.026 | 0.65 | Down |
| 22 | C00665 | D-Fructose 2,6-biphosphate | 12.21 | 1.87 | 0.003 | 0.64 | Down |
| 23 | C00392 | Mannitol | 10.91 | 1.28 | 0.047 | 0.62 | Down |
| 24 | — | Allylmalonic acid | 7.41 | 1.24 | 0.007 | 0.51 | Down |
| 25 | C00187 | Cholesterol | 16.77 | 2.53 | <0.001 | 0.48 | Down |
| 26 | C06730 | 4-Methylcatechol | 7.78 | 1.73 | 0.018 | 0.46 | Down |
| 27 | C02477 | alpha-Tocopherol | 16.53 | 2.29 | 0.011 | 0.40 | Down |
| 28 | C00180 | Benzoic acid | 6.79 | 2.01 | 0.033 | 0.39 | Down |
| 29 | C02591 | Sucrose-6-phosphate | 15.75 | 1.96 | 0.004 | 0.30 | Down |
| 30 | — | Methyl phosphate | 6.26 | 3.08 | <0.001 | 0.25 | Down |
| 31 | — | Hesperitin | 15.63 | 2.00 | 0.001 | 0.17 | Down |
| No. | Pathway (Metabolism) Name | Total a | Hits b | Raw p c | Holm p d | −ln(p) e | Impact f |
|---|---|---|---|---|---|---|---|
| 1 | Nitrogen | 9 | 2 | 0.007 | 0.594 | 4.92 | 0.00 |
| 2 | Glycine, serine, and threonine | 32 | 2 | 0.082 | 1 | 2.50 | 0.29 |
| 3 | Alanine, aspartate, and glutamate | 23 | 1 | 0.297 | 1 | 1.21 | 0.13 |
| 4 | Cysteine and methionine | 28 | 1 | 0.350 | 1 | 1.05 | 0.13 |
| 5 | Tyrosine | 42 | 1 | 0.477 | 1 | 0.74 | 0.11 |
| 6 | Primary bile acid biosynthesis | 46 | 2 | 0.151 | 1 | 1.89 | 0.07 |
| Parameters | Mean (n = 20) | SD | R-Value | p-Value |
|---|---|---|---|---|
| Norepinephrine (pg/mL) | 306.39 | 140.75 | 0.832 ** | <0.001 |
| Glycine (μmol/L) | 285.26 | 119.31 | 0.416 | 0.068 |
| L-Cysteine (ng/mL) | 70.00 | 4.31 | 0.503 * | 0.024 |
| L-Glutamine (μmol/L) | 207.76 | 55.03 | 0.538 * | 0.014 |
| Cholesterol (mmol/L) | 2.61 | 0.99 | −0.850 ** | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Jiang, X.; Hao, Y.; Sun, R.; Bai, Y.; Xu, C.; Xia, C. Serum Norepinephrine and Cholesterol Concentrations as Novel Diagnostic Biomarkers for Vitamin E Deficiency in Holstein Cows. Animals 2025, 15, 1333. https://doi.org/10.3390/ani15091333
Song Y, Jiang X, Hao Y, Sun R, Bai Y, Xu C, Xia C. Serum Norepinephrine and Cholesterol Concentrations as Novel Diagnostic Biomarkers for Vitamin E Deficiency in Holstein Cows. Animals. 2025; 15(9):1333. https://doi.org/10.3390/ani15091333
Chicago/Turabian StyleSong, Yuxi, Xuejie Jiang, Yu Hao, Rui Sun, Yunlong Bai, Chuang Xu, and Cheng Xia. 2025. "Serum Norepinephrine and Cholesterol Concentrations as Novel Diagnostic Biomarkers for Vitamin E Deficiency in Holstein Cows" Animals 15, no. 9: 1333. https://doi.org/10.3390/ani15091333
APA StyleSong, Y., Jiang, X., Hao, Y., Sun, R., Bai, Y., Xu, C., & Xia, C. (2025). Serum Norepinephrine and Cholesterol Concentrations as Novel Diagnostic Biomarkers for Vitamin E Deficiency in Holstein Cows. Animals, 15(9), 1333. https://doi.org/10.3390/ani15091333