
Metabolic Profiles of Serum and Ovarian Tissue in Taihe Black-Boned Silky Fowl During the Early and Peak Laying Periods


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animal Feeding and Sample Collection
2.3. Serum Hormone Measurement
2.4. Metabolite Extraction, Detection, and Analysis
2.5. Statistical Analysis
3. Results
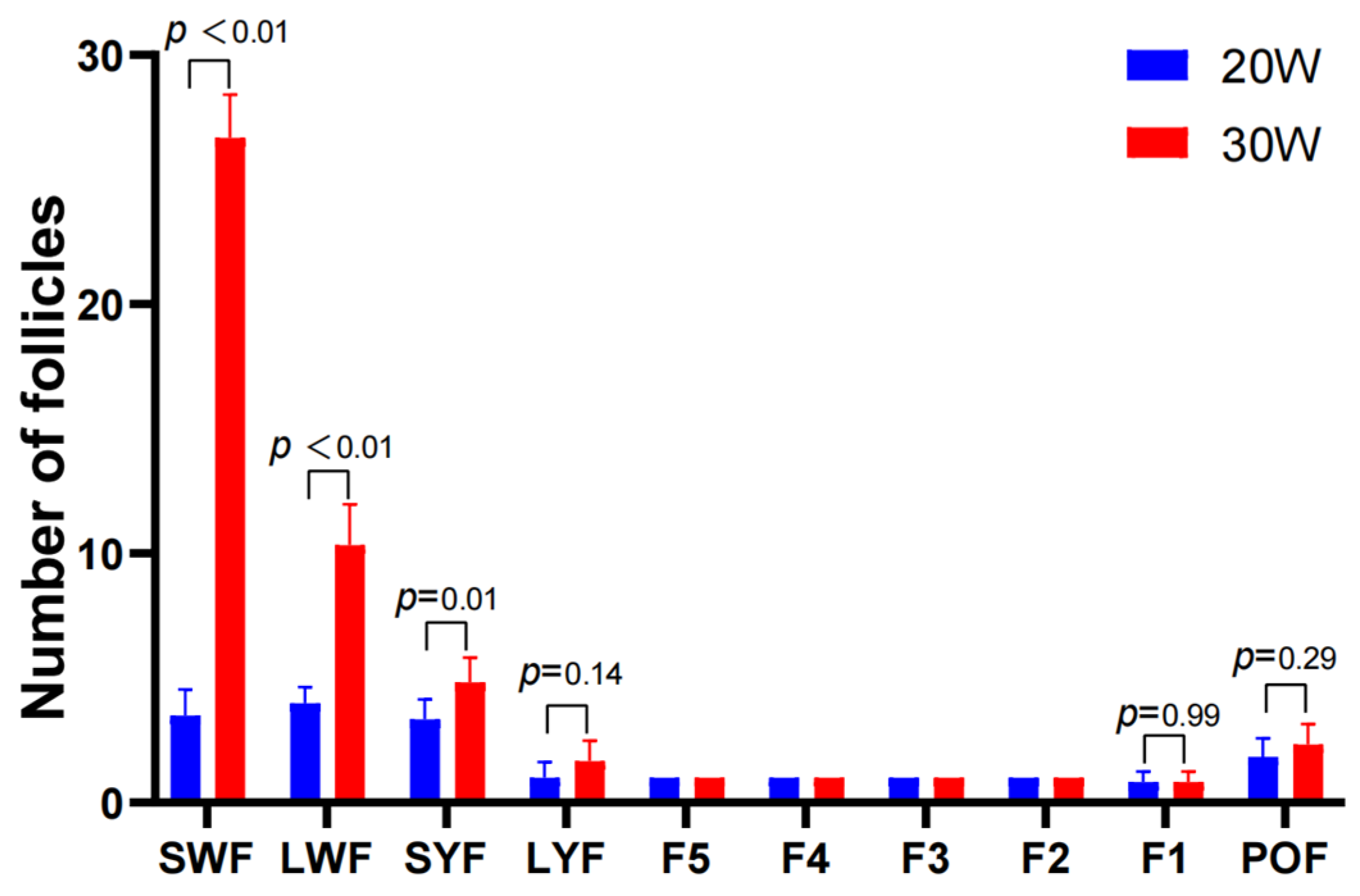
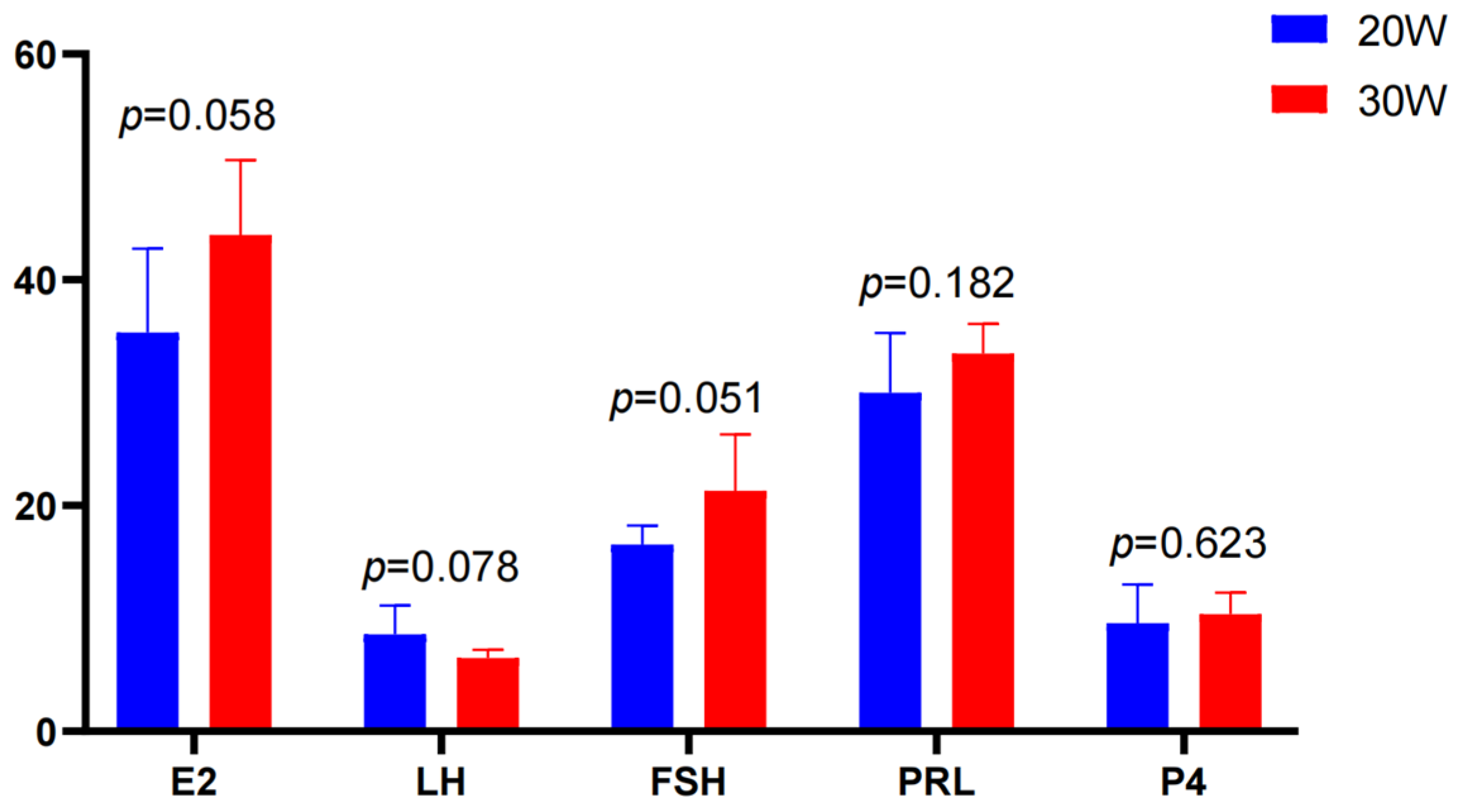
3.1. Follicle Counts and Serum Hormone Levels
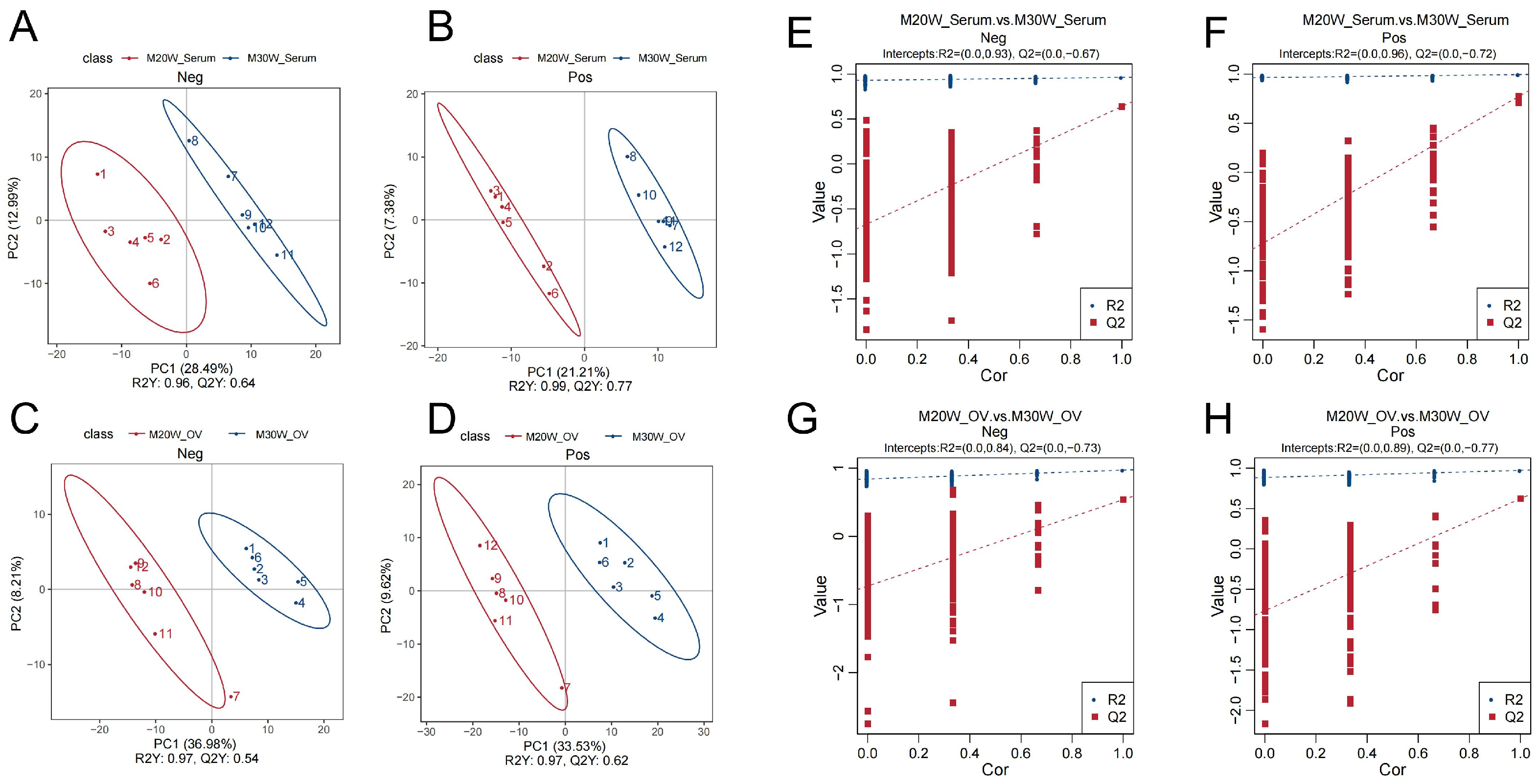
3.2. Metabolomics Quality Control
3.3. Correlation Analysis Between Serum Metabolites and Ovarian Tissue Metabolome
3.4. Metabolite Annotation
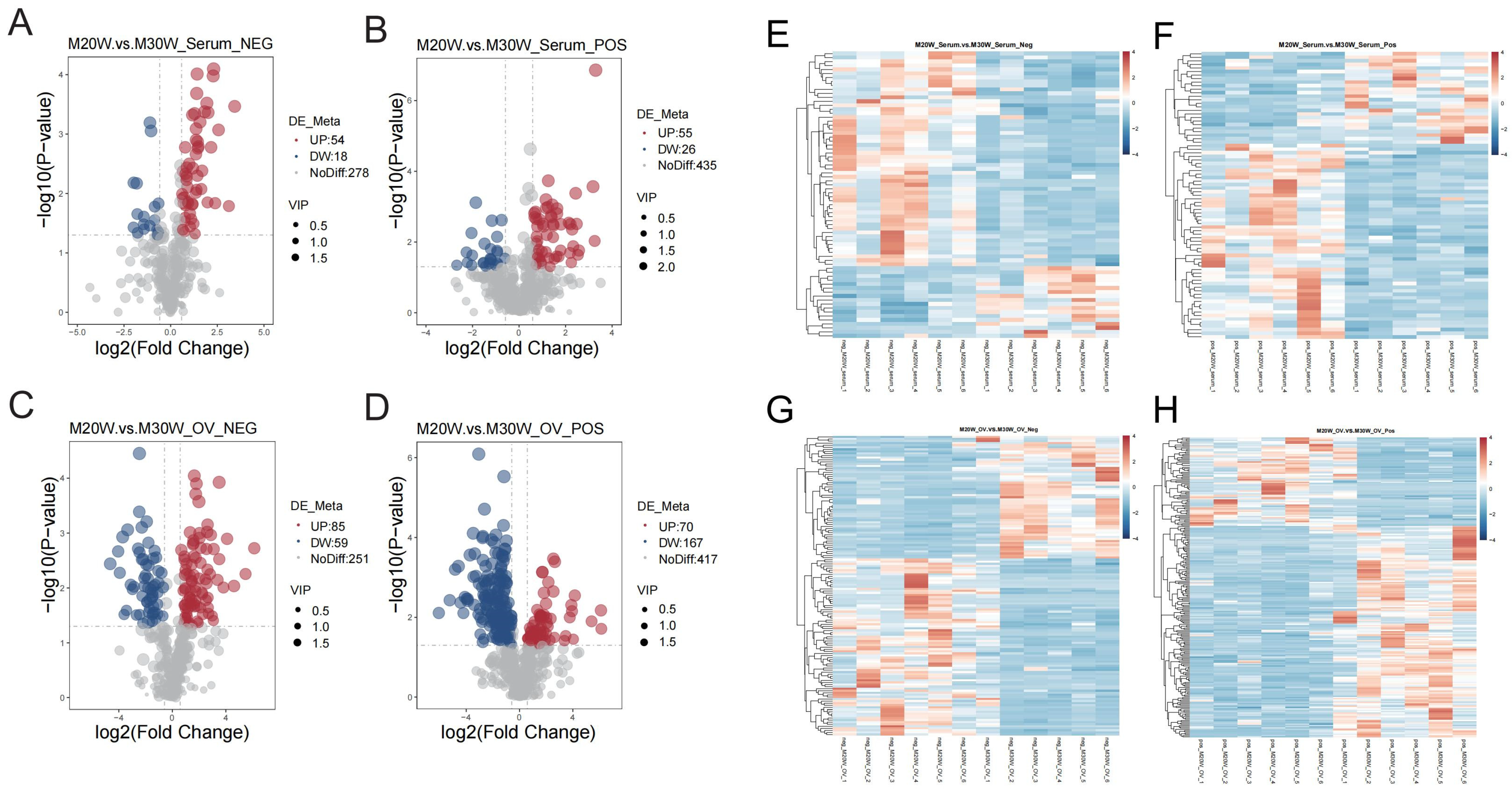
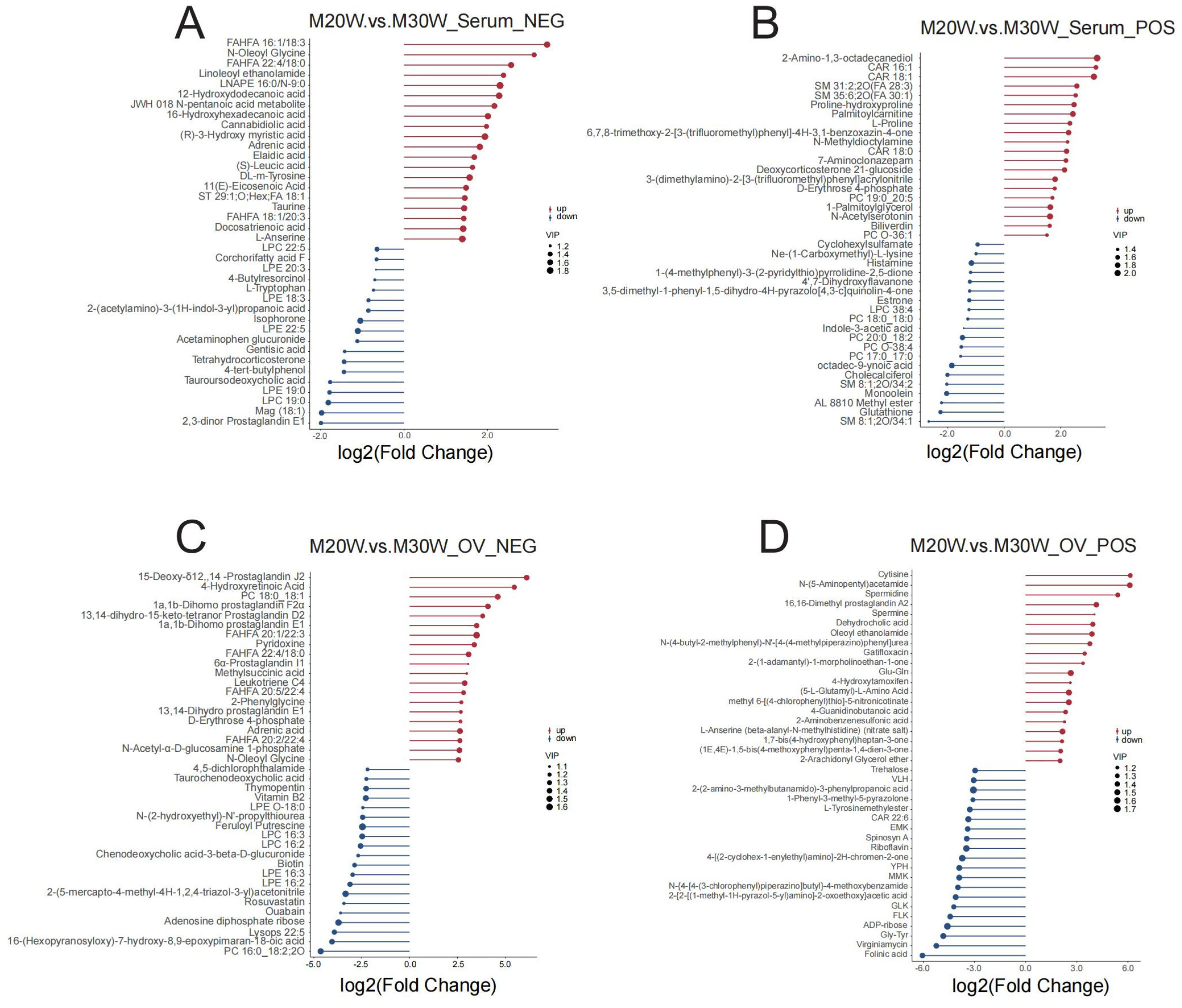
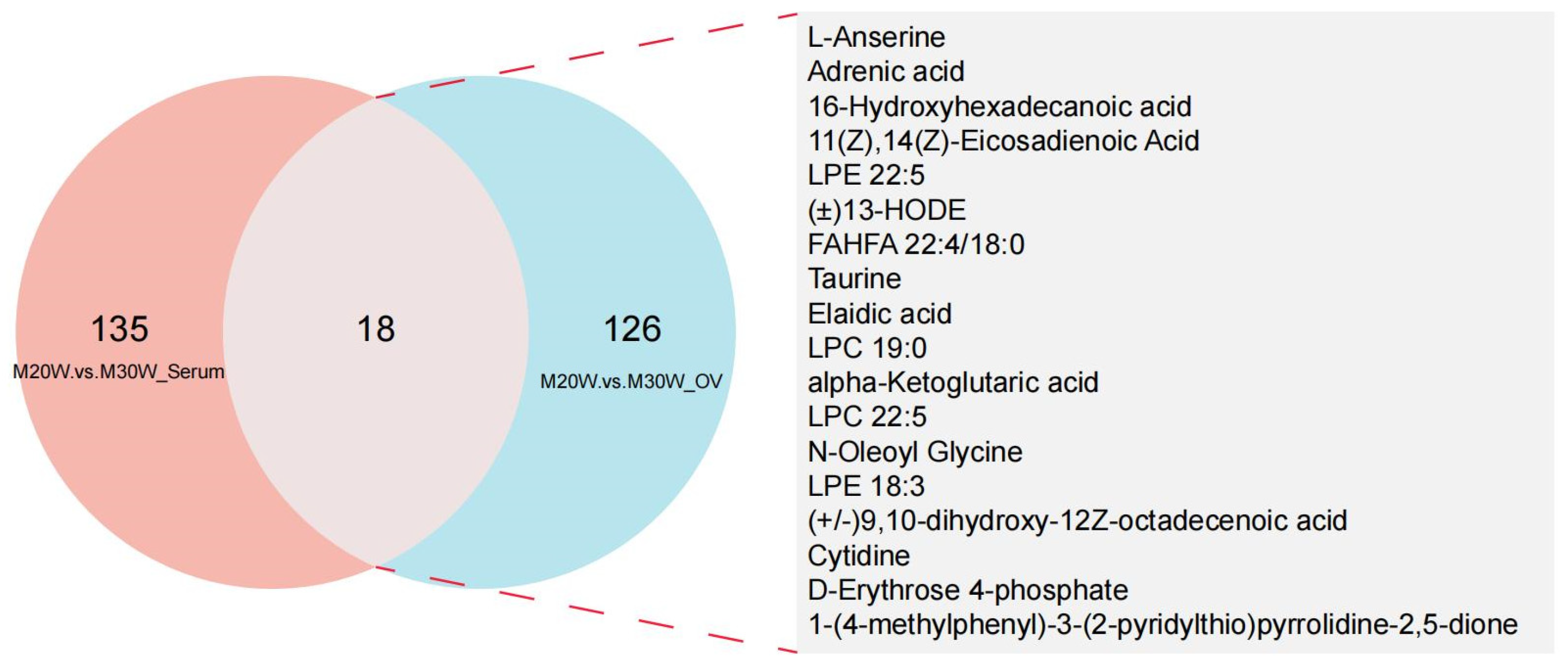
3.5. Differential Expressed Metabolites (DEMs) Characteristic
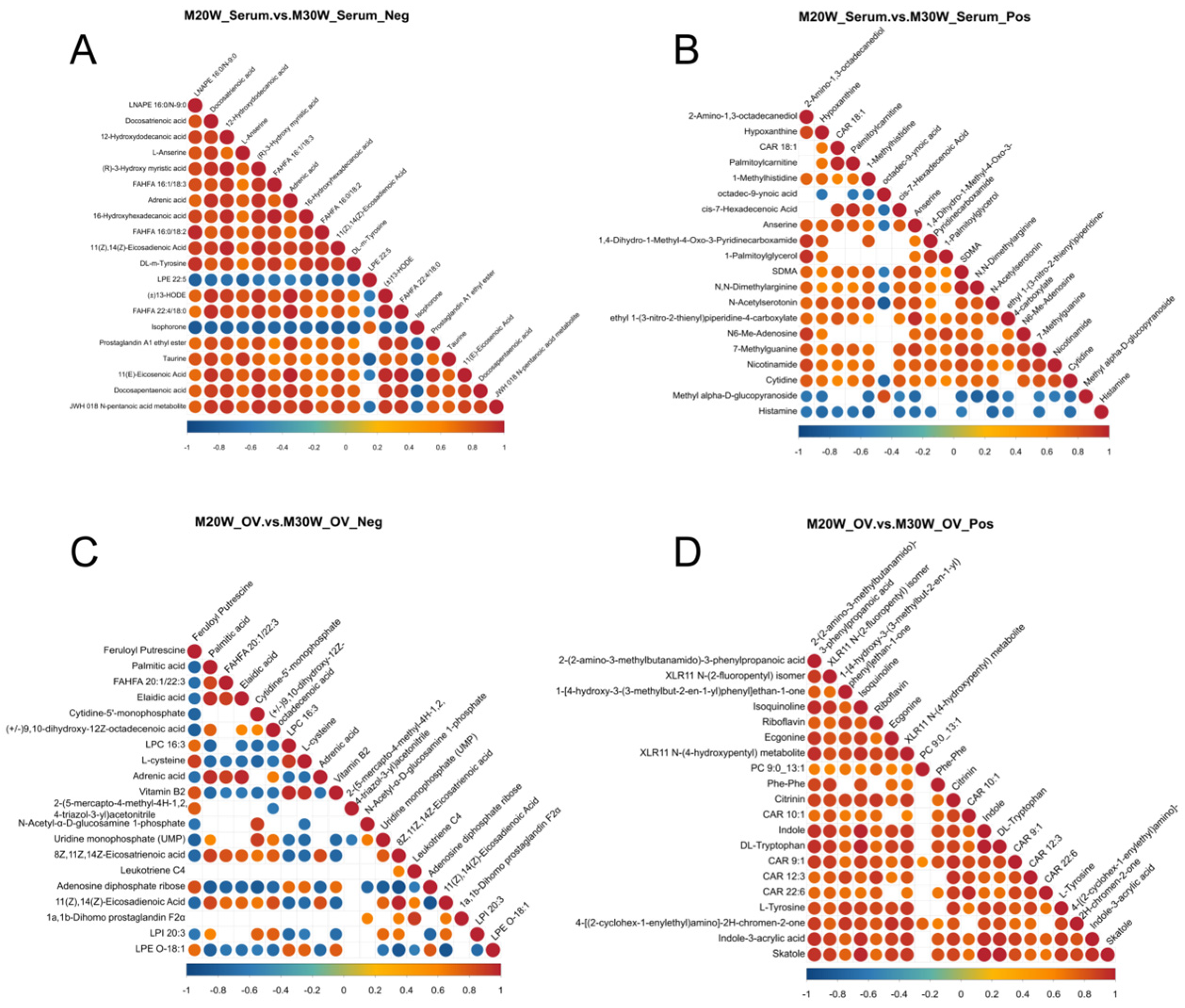
3.6. Correlation Analysis of Differential Metabolites
3.7. DEMs Functional Enrichment
4. Discussion
4.1. Spatiotemporal Heterogeneity in Systemic and Local Metabolic Regulation
4.2. Lipid Metabolic Dynamics Drive Ovulation and Hormone Synthesis
4.3. Crosstalk Between ECM Precursors and Polyamines in Ovarian Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 20W | 20-weeks-old |
| 30W | 30-weeks-old |
| COX | Cyclooxygenase |
| E2 | Estradiol |
| ECM | Extracellular matrix |
| FSH | Follicle-stimulating hormone |
| LH | Luteinizing hormone |
| mTOR | Mammalian target of rapamycin |
| PBS | Phosphate-buffered saline |
| PRL | Prolactin |
| P4 | Progesterone |
| TBSF | Taihe black-boned silky fowl |
References
- Robinson, F.E.; Renema, R.A.; Oosterhoff, H.H.; Zuidhof, M.J.; Wilson, J.L. Carcass Traits, Ovarian Morphology and Egg Laying Characteristics in Early versus Late Maturing Strains of Commercial Egg-Type Hens. Poult. Sci. 2001, 80, 37–46. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Li, Q.; Li, G.; Li, W.; Li, H.; Kang, X.; Tian, Y. Novel Regulatory Factors in the Hypothalamic-Pituitary-Ovarian Axis of Hens at Four Developmental Stages. Front. Genet. 2020, 11, 591672. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, W.; Cao, H.; Zhang, H.; Xiang, X.; Yin, Z. Ovarian Transcriptomic Analysis of Ninghai Indigenous Chickens at Different Egg-Laying Periods. Genes 2022, 13, 595. [Google Scholar] [CrossRef] [PubMed]
- Hao, E.-Y.; Wang, D.-H.; Chen, Y.-F.; Zhou, R.-Y.; Chen, H.; Huang, R.-L. The Relationship between the mTOR Signaling Pathway and Ovarian Aging in Peak-Phase and Late-Phase Laying Hens. Poult. Sci. 2021, 100, 334–347. [Google Scholar] [CrossRef]
- Hu, Z.; Liu, J.; Cao, J.; Zhang, H.; Liu, X. Ovarian Transcriptomic Analysis of Black Muscovy Duck at the Early, Peak and Late Egg-Laying Stages. Gene 2021, 777, 145449. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, Y.; Ruan, J.; Zhu, H.; Liang, H.; Cao, J.; Wei, Q.; Huang, J. Transcriptome and Chromatin Accessibility Landscape of Ovarian Development at Different Egg-Laying Stages in Taihe Black-Bone Silky Fowls. Poult. Sci. 2025, 104, 104864. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The Apogee of the Omics Trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Pan, B.; Chai, J.; Fei, K.; Zheng, T.; Jiang, Y. Dynamic Changes in the Transcriptome and Metabolome of Pig Ovaries across Developmental Stages and Gestation. BMC Genom. 2024, 25, 1193. [Google Scholar] [CrossRef]
- Yin, L.; Wang, X.; Zhang, D.; Lin, Z.; Wang, Y.; Yu, C.; Jie, H.; Xu, F.; Yang, C.; Liu, Y. The Proteome and Metabolome Changes Distinguish the Effect of Dietary Energy Levels on the Development of Ovary in Chicken during Sexual Maturity. Poult. Sci. 2024, 103, 104495. [Google Scholar] [CrossRef]
- Tu, Y.; Sun, Y.; Tian, Y.; Xie, M.; Chen, J. Physicochemical Characterisation and Antioxidant Activity of Melanin from the Muscles of Taihe Black-Bone Silky Fowl (Gallus Gallus Domesticus Brisson). Food Chem. 2009, 114, 1345–1350. [Google Scholar] [CrossRef]
- Qiu, Y.; Ying, J.; Yan, F.; Yu, H.; Zhao, Y.; Li, H.; Xia, S.; Chen, J.; Zhu, J. Novel Antiosteoporotic Peptides Purified from Protein Hydrolysates of Taihe Black-Boned Silky Fowl: By Larval Zebrafish Model and Molecular Docking. Food Res. Int. 2023, 169, 112850. [Google Scholar] [CrossRef]
- Huang, X.; Li, S.; Tan, Y.; Xu, C.; Huang, Y.; Yin, Z. Proteomic Analysis of Egg Production Peak and Senescence in the Ovaries of Taihe Black-Boned Silky Fowl (Gallus Gallus Domesticus Brisson). BMC Genom. 2025, 26, 17. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global Metabolic Profiling of Animal and Human Tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A Flexible and Comprehensive Software for Processing Metabolomics Data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [PubMed]
- Suhre, K.; Gieger, C. Genetic Variation in Metabolic Phenotypes: Study Designs and Applications. Nat. Rev. Genet. 2012, 13, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, X.; Wang, R.; Wei, Y.; Peng, H.; Wang, K.; Li, H.; Ji, Y. Metabolomic Profiling of Exosomes Reveals Age-Related Changes in Ovarian Follicular Fluid. Eur. J. Med. Res. 2024, 29, 4. [Google Scholar] [CrossRef]
- Seol, H.S.; Sato, K.; Murakami, H.; Toyomizu, M.; Akiba, Y. Changes in Gene Expression Involved in Energy Utilization during Chicken Follicle Development. Anim. Reprod. Sci. 2006, 95, 283–294. [Google Scholar] [CrossRef]
- Cui, Z.; Ning, Z.; Deng, X.; Du, X.; Amevor, F.K.; Liu, L.; Kang, X.; Tian, Y.; Wang, Y.; Li, D.; et al. Integrated Proteomic and Metabolomic Analyses of Chicken Ovary Revealed the Crucial Role of Lipoprotein Lipase on Lipid Metabolism and Steroidogenesis During Sexual Maturity. Front. Physiol. 2022, 13, 885030. [Google Scholar] [CrossRef]
- Blitek, A.; Szymanska, M. Expression Profiles of Fatty Acid Transporters and the Role of N-3 and n-6 Polyunsaturated Fatty Acids in the Porcine Endometrium. Int. J. Mol. Sci. 2024, 25, 11102. [Google Scholar] [CrossRef]
- Crawford, M.A.; Sinclair, A.J.; Hall, B.; Ogundipe, E. The Imperative of Arachidonic Acid in Human Reproduction. Prog. Lipid Res. 2023, 91, 101222. [Google Scholar] [CrossRef]
- Cebrián-Prats, A.; González-Lafont, À.; Lluch, J.M. Unraveling the Molecular Details of the Complete Mechanism That Governs the Synthesis of Prostaglandin G2 Catalyzed by Cyclooxygenase-2. ACS Omega 2019, 4, 2063–2074. [Google Scholar] [CrossRef]
- Strickland, S.; Beers, W.H. Studies on the Role of Plasminogen Activator in Ovulation. In Vitro Response of Granulosa Cells to Gonadotropins, Cyclic Nucleotides, and Prostaglandins. J. Biol. Chem 1976, 251, 5694–5702. [Google Scholar] [CrossRef] [PubMed]
- Höper, M.M.; Voelkel, N.F.; Bates, T.O.; Allard, J.D.; Horan, M.; Shepherd, D.; Tuder, R.M. Prostaglandins Induce Vascular Endothelial Growth Factor in a Human Monocytic Cell Line and Rat Lungs via cAMP. Am. J. Respir. Cell Mol. Biol. 1997, 17, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, J.; Johnson, A.L. Vascular Endothelial Growth Factor and Angiopoietins during Hen Ovarian Follicle Development. Gen. Comp. Endocrinol. 2016, 232, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Chen, X.; Mao, Y.; Kang, L.; Ma, X.; Jiang, Y. Characterization of Annexin A2 in Chicken Follicle Development: Evidence for Its Involvement in Angiogenesis. Anim. Reprod. Sci. 2015, 161, 104–111. [Google Scholar] [CrossRef]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-Delta 12,14-Prostaglandin J2 Is a Ligand for the Adipocyte Determination Factor PPAR Gamma. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef]
- Capobianco, E.; Jawerbaum, A.; Romanini, M.C.; White, V.; Pustovrh, C.; Higa, R.; Martinez, N.; Mugnaini, M.T.; Soñez, C.; Gonzalez, E. 15-Deoxy-Delta12,14-Prostaglandin J2 and Peroxisome Proliferator-Activated Receptor Gamma (PPARgamma) Levels in Term Placental Tissues from Control and Diabetic Rats: Modulatory Effects of a PPARgamma Agonist on Nitridergic and Lipid Placental Metabolism. Reprod. Fertil. Dev. 2005, 17, 423–433. [Google Scholar] [CrossRef]
- Wei, Y.; Li, W.; Meng, X.; Zhang, L.; Shen, M.; Liu, H. Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary. Animals 2019, 9, 1047. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Musa, H.H.; Sifaldin, A.Z. Prenatal Corticosterone Exposure Programs Growth, Behavior, Reproductive Function and Genes in the Chicken. Asian Pac. J. Reprod. 2016, 5, 271–278. [Google Scholar] [CrossRef]
- Leszczynski, D.E.; Hagan, R.C.; Bitgood, J.J.; Kummerow, F.A. Relationship of Plasma Estradiol and Progesterone Levels to Egg Productivity in Domestic Chicken Hens. Poult. Sci. 1985, 64, 545–549. [Google Scholar] [CrossRef]
- Zhou, S.; Ma, Y.; Yao, J.; Zhao, A.; Xie, C.; Mi, Y.; Zhang, C. TGF-Β1-Induced Collagen Promotes Chicken Ovarian Follicle Development via an Intercellular Cooperative Pattern. Cell Biol. Int. 2021, 45, 1336–1348. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Roles of Dietary Glycine, Proline, and Hydroxyproline in Collagen Synthesis and Animal Growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Kivirikko, K.I.; Pihlajaniemi, T. Collagen Hydroxylases and the Protein Disulfide Isomerase Subunit of Prolyl 4-Hydroxylases. Adv. Enzymol. Relat. Areas Mol. Biol. 1998, 72, 325–398. [Google Scholar] [CrossRef] [PubMed]
- Myllyharju, J. Prolyl 4-Hydroxylases, the Key Enzymes of Collagen Biosynthesis. Matrix Biol. 2003, 22, 15–24. [Google Scholar] [CrossRef]
- Holowatz, L.A.; Thompson, C.S.; Kenney, W.L. L-Arginine Supplementation or Arginase Inhibition Augments Reflex Cutaneous Vasodilatation in Aged Human Skin. J. Physiol. 2006, 574, 573–581. [Google Scholar] [CrossRef]
- Kay, E.J.; Koulouras, G.; Zanivan, S. Regulation of Extracellular Matrix Production in Activated Fibroblasts: Roles of Amino Acid Metabolism in Collagen Synthesis. Front. Oncol. 2021, 11, 719922. [Google Scholar] [CrossRef]
- Yan, J.; Yan, J.-Y.; Wang, Y.-X.; Ling, Y.-N.; Song, X.-D.; Wang, S.-Y.; Liu, H.-Q.; Liu, Q.-C.; Zhang, Y.; Yang, P.-Z.; et al. Spermidine-Enhanced Autophagic Flux Improves Cardiac Dysfunction Following Myocardial Infarction by Targeting the AMPK/mTOR Signalling Pathway. Br. J. Pharmacol. 2019, 176, 3126–3142. [Google Scholar] [CrossRef]
- Rider, J.E.; Hacker, A.; Mackintosh, C.A.; Pegg, A.E.; Woster, P.M.; Casero, R.A. Spermine and Spermidine Mediate Protection against Oxidative Damage Caused by Hydrogen Peroxide. Amino Acids 2007, 33, 231–240. [Google Scholar] [CrossRef]
- Liu, L.; Santora, R.; Rao, J.N.; Guo, X.; Zou, T.; Zhang, H.M.; Turner, D.J.; Wang, J.-Y. Activation of TGF-Beta-Smad Signaling Pathway Following Polyamine Depletion in Intestinal Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1056–G1067. [Google Scholar] [CrossRef]
- Yu, D.; Wan, H.; Tong, C.; Guang, L.; Chen, G.; Su, J.; Zhang, L.; Wang, Y.; Xiao, Z.; Zhai, J.; et al. A Multi-Tissue Metabolome Atlas of Primate Pregnancy. Cell 2024, 187, 764–781.e14. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Zhao, J.; Zhao, Y.; Li, Z.; Yang, S.; Chen, N.; Liu, Y.; Zhang, J.; Zhou, L.; Chen, X. Multi-Omics Reveal the Effects and Regulatory Mechanism of Dietary Magnolol Supplementation on Production Performance of Post-Peak Laying Hens. J. Agric. Food Chem. 2025, 73, 4027–4041. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, S.; Chen, Y.; Deng, K.; Kuang, M.; Gong, Z.; Zhang, K.; Wang, P.; Huang, P.; Zhou, Z.; et al. Exploring Nicotinamide Adenine Dinucleotide Precursors across Biosynthesis Pathways: Unraveling Their Role in the Ovary. FASEB J. 2024, 38, e23804. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Li, S.; Yin, Z. Metabolic Profiles of Serum and Ovarian Tissue in Taihe Black-Boned Silky Fowl During the Early and Peak Laying Periods. Animals 2025, 15, 912. https://doi.org/10.3390/ani15070912
Huang X, Li S, Yin Z. Metabolic Profiles of Serum and Ovarian Tissue in Taihe Black-Boned Silky Fowl During the Early and Peak Laying Periods. Animals. 2025; 15(7):912. https://doi.org/10.3390/ani15070912
Chicago/Turabian StyleHuang, Xuan, Shibao Li, and Zhaozheng Yin. 2025. "Metabolic Profiles of Serum and Ovarian Tissue in Taihe Black-Boned Silky Fowl During the Early and Peak Laying Periods" Animals 15, no. 7: 912. https://doi.org/10.3390/ani15070912
APA StyleHuang, X., Li, S., & Yin, Z. (2025). Metabolic Profiles of Serum and Ovarian Tissue in Taihe Black-Boned Silky Fowl During the Early and Peak Laying Periods. Animals, 15(7), 912. https://doi.org/10.3390/ani15070912