
Study of Zoonotic Pathogens in Alien Population of Veiled Chameleons (Chamaeleo calyptratus) in the Canary Islands (Spain)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Isolation of Bacteria from the Samples
2.3. Molecular Identification
2.3.1. DNA Extraction
2.3.2. PCR Identification
2.4. Co-Infection Index (Ic)
2.5. Statistical Analysis
3. Results
3.1. Campylobacter spp.
3.2. Escherichia coli Virulence Genes (stx1, stx2, and eae)
3.3. Listeria monocytogenes
3.4. Mycobacterium spp.
3.5. Pseudomonas spp.
3.6. Salmonella spp.
3.7. Staphylococcus spp.
3.8. Vibrio sp.
3.9. Yersinia enterocolitica
3.10. Co-Infection and Co-Infection Index (Ic)
4. Discussion
4.1. Campylobacter spp.
4.2. Escherichia coli Virulence Genes (stx1, stx2, and eae)
4.3. Listeria monocytogenes
4.4. Mycobacterium spp.
4.5. Pseudomonas spp.
4.6. Salmonella spp.
4.7. Staphylococcus spp.
4.8. Vibrio spp.
4.9. Yersinia enterocolitica
4.10. Summary
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, M.C.; Adams, H.; Hope, B.; Powell, M. Risk Assessment for Invasive Species. Risk Anal. 2004, 24, 787–793. [Google Scholar] [CrossRef]
- Najberek, K.; Olszańska, A.; Tokarska-Guzik, B.; Mazurska, K.; Dajdok, Z.; Solarz, W. Invasive alien species as reservoirs for pathogens. Ecol. Indic. 2022, 139, 108879. [Google Scholar] [CrossRef]
- Ciencia Canaria—Invasoras: Las Especies Que Acechan en Las Islas. Available online: https://www.cienciacanaria.es/secciones/a-fondo/1174-invasoras-las-especies-que-acechan-en-las-islas (accessed on 21 April 2022).
- Especies Exóticas Invasoras. Available online: https://www.gobiernodecanarias.org/medioambiente/materias/biodiversidad/especies-exoticas-invasoras/efectos-sopbre-la-biodiversidad-local/efectos-potenciales-introduccion/ (accessed on 19 May 2022).
- Efectos. Available online: https://www.gobiernodecanarias.org/medioambiente/materias/biodiversidad/especies-exoticas-invasoras/efectos-sopbre-la-biodiversidad-local/ (accessed on 21 April 2022).
- La Serpiente Invasora de Gran Canaria—#STOPCULEBRAREAL. Available online: https://www.stopculebrareal.com/info/ (accessed on 21 April 2022).
- Exos. Available online: https://www.biodiversidadcanarias.es/exos/especie/E09198 (accessed on 21 April 2022).
- Gillette, C.R.; Krysko, K.L. New County Record for The Veiled Chameleon, Chamaeleo calyptratus Duméril and Bibron 1851 (Sauria: Chamaeleonidae), in Florida. Reptiles Amphib. 2012, 19, 130–131. [Google Scholar] [CrossRef]
- López, C.; Clemente, S.; Almeida, C.; Brito, A.; Hernández, M. A genetic approach to the origin of Millepora sp. in the eastern Atlantic. Coral Reefs 2015, 34, 631–638. [Google Scholar] [CrossRef]
- Wang, G.; Clark, C.G.; Taylor, T.M.; Pucknell, C.; Barton, C.; Price, L.; Woodward, D.L.; Rodgers, F.G. Colony Multiplex PCR Assay for Identification and Differentiation of Campylobacter jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus. J. Clin. Microbiol. 2002, 40, 4744–4747. [Google Scholar] [CrossRef]
- Blanco, M.; Blanco, J.E.; Mora, A.; Dahbi, G.; Alonso, M.P.; González, E.A.; Bernárdez, M.I.; Blanco, J. Serotypes, virulence genes, and intimin types of shiga toxin (verotoxin)-producing Escherichia coli isolates from cattle in Spain and identification of a new intimin variant gene (eae-ξ). J. Clin. Microbiol. 2004, 42, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Jaton, K.; Sahli, R.; Bille, J. Development of Polymerase Chain Reaction assays for detection of Listeria monocytogenes in clinical cerebrospinal fluid samples. J. Clin. Microbiol. 1992, 30, 1931–1936. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Choi, Y.; Jeon, B.Y.; Jin, H.; Cho, S.N.; Lee, H. A simple and efficient Multiplex PCR assay for the identification of Mycobacterium genus and Mycobacterium tuberculosis complex to the species level. Yonsei Med. J. 2013, 54, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- De Vos, D.; Lim, A., Jr.; Pirnay, J.P.; Struelens, M.; Vandenvelde, C.; Duinslaeger, L.; Vanderkelen, A.; Cornelis, P. Direct detection and identification of Pseudomonas aeruginosa in clinical samples such as skin biopsy specimens and expectorations by Multiplex PCR Based on two outer membrane lipoprotein genes, oprI and oprL. J. Clin. Microbiol. 1997, 35, 1295–1299. [Google Scholar] [CrossRef]
- Guimarães de Freitas, C.; Patrícia Santana, A.; da Silva, P.H.C.; Salvador Picão Gonçalves, V.; Ferreira Barros, M.A.; Gonçalves Torres, F.A.; Murata, L.S.; Perecmanis, S. PCR multiplex for detection of Salmonella Enteritidis, Typhi and Typhimurium and occurrence in poultry meat. Int. J. Food Microbiol. 2010, 139, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Campos-Peña, E.; Martín-Nuñez, E.; Pulido-Reyes, G.; Martín-Padrón, J.; Caro-Carrillo, E.; Donate-Correa, J.; Lorenzo-Castrillejo, I.; Alcoba-Flórez, J.; Machín, F.; Méndez-Alvarez, S. Multiplex PCR assay for identification of six different Staphylococcus spp. and simultaneous detection of methicillin and mupirocin resistance. J. Clin. Microbiol. 2014, 52, 2698–2701. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, G.; Wang, H.; Chen, J.; Shi, X.; Zou, G.; Wei, Q.; Sun, X. Design of Vibrio 16S rRNA gene specific primers and their application in the analysis of seawater Vibrio community. J. Ocean Univ. China 2006, 5, 157–164. [Google Scholar]
- Neogi, S.B.; Chowdhury, N.; Asakura, M.; Hinenoya, A.; Haldar, S.; Saidi, S.M.; Kogure, K.; Lara, R.J.; Yamasaki, S. A highly sensitive and specific multiplex PCR assay for simultaneous detection of Vibrio cholerae, Vibrio parahaemolyticus and Vibrio vulnificus. Lett. Appl. Microbiol. 2010, 51, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Wannet, W.J.B.; Reessink, M.; Brunings, H.A.; Maas, H.M.E. Detection of pathogenic Yersinia enterocolitica by rapid and sensitive duplex PCR assay. J. Clin. Microbiol. 2001, 39, 4483–4486. [Google Scholar] [CrossRef]
- Ginsberg, H.S. Potential Effects of Mixed Infections in Ticks on Transmission Dynamics of Pathogens: Comparative Analysis of Published Records. Exp. Appl. Acarol. 2008, 46, 29–41. [Google Scholar] [CrossRef]
- Masila, N.M.; Ross, K.E.; Gardner, M.G.; Whiley, H. Zoonotic and Public Health Implications of Campylobacter Species and Squamates (Lizards, Snakes and Amphisbaenians). Pathogens 2020, 9, 799. [Google Scholar] [CrossRef]
- Fitzgerald, C. Campylobacter. Clin. Lab. Med. 2015, 35, 289–298. [Google Scholar] [CrossRef]
- De Luca, C.; Iraola, G.; Apostolakos, I.; Boetto, E.; Piccirillo, A. Occurrence and diversity of Campylobacter species in captive chelonians. Vet. Microbiol. 2020, 241, 108567. [Google Scholar] [CrossRef]
- Huang, H.; Brooks, B.W.; Lowman, R.; Carrillo, C.D. Campylobacter species in animal, food, and environmental sources, and relevant testing programs in Canada. Can. J. Microbiol. 2015, 61, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.J.; Kik, M.; Miller, W.G.; Duim, B.; Wagenaar, J.A. Campylobacter iguaniorum sp. nov., isolated from reptiles. Int. J. Syst. Evol. Microbiol. 2015, 65, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, A.; Niero, G.; Calleros, L.; Pérez, R.; Naya, H.; Iraola, G. Campylobacter geochelonis sp. nov. isolated from the western Hermann’s tortoise (Testudo hermanni hermanni). Int. J. Syst. Evol. Microbiol. 2016, 66, 3468–3476. [Google Scholar] [CrossRef] [PubMed]
- Gourmelon, M.; Boukerb, A.M.; Nabi, N.; Banerji, S.; Joensen, K.G.; Serghine, J.; Cormier, A.; Megraud, F.; Lehours, P.; Alter, T.; et al. Genomic Diversity of Campylobacter lari group isolates from Europe and Australia in a One Health context. Appl. Environ. Microbiol. 2022, 88, e0136822. [Google Scholar] [CrossRef]
- Categoría:Aves—CanariWiki. Available online: https://www3.gobiernodecanarias.org/medusa/wiki/index.php?title=Categor%C3%ADa:Aves (accessed on 31 May 2023).
- Martinot, M.; Jaulhac, B.; Moog, F.; De Martino, S.; Kehrli, P.; Monteil, H.; Piemont, Y. Campylobacter lari bacteremia. Clin. Microbiol. Infect. 2001, 7, 96–97. [Google Scholar] [CrossRef]
- Krause, R.; Ramschak-Schwarzer, S.; Gorkiewicz, G.; Schnedl, W.J.; Feierl, G.; Wenisch, C.; Reisinger, E.C. Recurrent septicemia due to Campylobacter fetus and Campylobacter lari in an immunocompetent patient. Infection 2002, 30, 171–174. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications—A review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef]
- Kolenda, R.; Burdukiewicz, M.; Schierack, P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. coli. Front. Cell. Infect. Microbiol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Guragain, M.; Schmidt, J.W.; Dickey, A.M.; Bosilevac, J.M. Distribution of extremely heat-resistant Escherichia coli in the beef production and processing continuum. J. Food. Prot. 2023, 86, 100031. [Google Scholar] [CrossRef]
- Karmali, M.A.; Gannon, V.; Jan, M.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Stepien-Pysniak, D.; Szczepaniak, K.; Turchi, B.; Urban-Chmiel, R. Virulence profiles and antibiotic susceptibility of Escherichia coli strains from pet reptiles. Pathogens 2022, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Gopee, N.V.; Adesiyun, A.A.; Caesar, K. A longitudinal study of Escherichia coli strains isolated from captive mammals, birds, and reptiles in Trinidad. J. Zoo Wildl. Med. 2000, 31, 353–360. [Google Scholar]
- Bautista-Trujillo, G.U.; Gutiérrez-Miceli, F.A.; Mandujano-García, L.; Oliva-Llaven, M.A.; Ibarra-Martínez, C.; Mendoza-Nazar, P.; Ruiz-Sesma, B.; Tejeda-Cruz, C.; Pérez-Vázquez, L.C.; Pérez-Batrez, J.E.; et al. Captive green iguana carries diarrheagenic Escherichia coli pathotypes. Front. Vet. Sci. 2020, 7, 99. [Google Scholar] [CrossRef]
- Martínez, R.; Sánchez, S.; Alonso, J.M.; Herrera-León, S.; Rey, J.; Echeita, M.A.; Morán, J.M.; García-Sánchez, A. Salmonella spp. and Shiga toxin-producing Escherichia coli prevalence in an ocellated lizard (Timon lepidus) research center in Spain. Foodborne Pathog. Dis. 2011, 8, 1309–1311. [Google Scholar] [CrossRef]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga Toxin Producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef]
- Vieira, M.A.; Andrade, J.R.; Trabulsi, L.R.; Rosa, A.C.; Días, A.M.; Ramos, S.R.; Frankel, G.; Gomes, T.A. Phenotypic and genotypic characteristics of Escherichia coli strains of non-enteropathogenic E. coli (EPEC) serogroups that carry eae and lack the EPEC adherence factor and Shiga toxin DNA probe sequences. J. Infect. Dis. 2001, 183, 762–772. [Google Scholar] [CrossRef]
- Balière, C.; Rincé, A.; Delannoy, S.; Fach, P.; Gourmelon, M. Molecular profiling of Shiga toxin-producing Escherichia coli and enteropathogenic E. coli strains isolated from French coastal environments. Appl. Environ. Microbiol. 2016, 82, 3913–3927. [Google Scholar] [CrossRef] [PubMed]
- Disson, O.; Moura, A.; Lecuit, M. Making sense of the biodiversity and virulence of Listeria monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Guldimann, C.; Märtlbauer, E. Asymptomatic carriage of Listeria monocytogenes by animals and humans and its impact on the food chain. Foods 2022, 11, 3472. [Google Scholar] [CrossRef] [PubMed]
- Schönberg, A.; Gerigk, K. Listeria in effluents from the food-processing industry. Rev. Sci. Tech. 1991, 10, 787–797. [Google Scholar] [CrossRef]
- Weber, A.; Prell, A.; Potel, J.; Schäfer, R. Vorkommen von Listeria monocytogenes bei Schlangen, Schildkröten, Echsen und Amphibien in der Heimtierhaltung [Occurrence of Listeria monocytogenes in snakes, tortoises, lizards and amphibians raised as pets]. Berl. Munch. Tierarztl. Wochenschr. 1993, 106, 293–295. [Google Scholar] [PubMed]
- Weber, A.; Potel, J.; Schäfer-Schmidt, R.; Prell, A.; Datzmann, C. Untersuchungen zum Vorkommen von Listeria monocytogenes in Kotproben von Haus- und Heimtieren [Studies on the occurrence of Listeria monocytogenes in fecal samples of domestic and companion animals]. Zentralblatt Hyg. Umweltmed. 1995, 198, 117–123. [Google Scholar]
- Chen, T.; Orsi, R.H.; Chen, R.; Gunderson, M.; Roof, S.; Wiedmann, M.; Childs-Sanford, S.E.; Cummings, K.J. Characterization of Listeria monocytogenes isolated from wildlife in central New York. Vet. Med. Sci. 2022, 8, 1319–1329. [Google Scholar] [CrossRef]
- Nowakiewicz, A.; Ziółkowska, G.; Zięba, P.; Dziedzic, B.M.; Gnat, S.; Wójcik, M.; Dziedzic, R.; Kostruba, A. Aerobic bacterial microbiota isolated from the cloaca of the European pond turtle (Emys orbicularis) in Poland. J. Wildl. Dis. 2015, 51, 255–259. [Google Scholar] [CrossRef]
- Matt, C.L.; Ramachandran, A.; Allison, R.W.; Wall, C.R.; Dieterly, A.M.; Brandão, J. Listeria monocytogenes in an inland bearded dragon (Pogona vitticeps). J. Exot. Pet Med. 2019, 30, 76–81. [Google Scholar] [CrossRef]
- Girling, S.J.; Fraser, M.A. Listeria monocytogenes septicaemia in an inland bearded dragon, Pogona vitticeps. J. Herpetol. Med. Surg. 2004, 14, 6–9. [Google Scholar] [CrossRef]
- Di Renzo, L.; De Angelis, M.E.; Torresi, M.; Di Lollo, V.; Di Teodoro, G.; Averaimo, D.; Defourny, S.V.P.; Di Giacinto, F.; Profico, C.; Olivieri, V.; et al. First Report of Septicaemic Listeriosis in a loggerhead sea turtle (Caretta caretta) stranded along the adriatic coast: Strain detection and sequencing. Animals 2022, 12, 2364. [Google Scholar] [CrossRef]
- Mitchell, M.A. Mycobacterial infections in reptiles. Vet. Clin. North. Am. Exot. Anim. Pract. 2012, 15, 101–111. [Google Scholar]
- Tortoli, E. Microbiological features and clinical relevance of new species of the genus Mycobacterium. Clin. Microbiol. Rev. 2014, 27, 727–752. [Google Scholar] [CrossRef] [PubMed]
- Ebani, V.V. Domestic reptiles as source of zoonotic bacteria: A mini review. Asian Pac. J. Trop. Med. 2017, 10, 723–728. [Google Scholar] [CrossRef]
- Soldati, G.; Lu, Z.H.; Vaughan, L.; Polkinghorne, A.; Zimmermann, D.R.; Huder, J.B.; Pospischil, A. Detection of mycobacteria and chlamydiae in granulomatous inflammation of reptiles: A retrospective study. Vet. Pathol. 2004, 41, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, L.S.; das Neves Dias-Neto, R.; Cagnini, D.Q.; Yamatogi, R.S.; Oliveira-Filho, J.P.; Nemer, V.; Teixeira, R.H.; Biondo, A.W.; Araújo, J.P., Jr. Mycobacterium genavense infection in two species of captive snakes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 18, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Reavill, D.R.; Schmidt, R.E. Mycobacterial lesions in fish, amphibians, reptiles, rodents, lagomorphs, and ferrets with reference to animal models. Vet. Clin. N. Am. Exot. Anim. Pract. 2012, 15, 25–40. [Google Scholar] [CrossRef]
- Ebani, V.V.; Fratini, F.; Bertelloni, F.; Cerri, D.; Tortoli, E. Isolation and identification of mycobacteria from captive reptiles. Res. Vet. Sci. 2012, 93, 1136–1138. [Google Scholar] [CrossRef]
- Maluta, A.; Zając, M.; Krajewska-Wędzina, M.; Wasyl, D.; Heckers, K.; Didkowska, A.; Anusz, K. Mixed infection of Mycobacterium szulgai, M. lentiflavum, and Gram-negative bacteria as a cause of death in a brown caiman Caiman crocodylus: A case report. Vet. Sci. 2022, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Bouricha, M.; Castan, B.; Duchene-Parisi, E.; Drancourt, M. Mycobacterium marinum infection following contact with reptiles: Vivarium granuloma. Int. J. Infect. Dis. 2014, 21, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Thoen, C.; Lobue, P.; de Kantor, I. The importance of Mycobacterium bovis as a zoonosis. Vet. Microbiol. 2006, 112, 339–345. [Google Scholar] [CrossRef]
- Une, Y.; Mori, T. Tuberculosis as a zoonosis from a veterinary perspective. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 415–425. [Google Scholar] [CrossRef]
- Mena, K.D.; Gerba, C.P. Risk assessment of Pseudomonas aeruginosa in water. Rev. Environ. Contam. Toxicol. 2009, 201, 71–115. [Google Scholar]
- Ebani, V.V.; Fratini, F.; Ampola, M.; Rizzo, E.; Cerri, D.; Andreani, E. Pseudomonas and Aeromonas isolates from domestic reptiles and study of their antimicrobial in vitro sensitivity. Vet. Res. Commun. 2008, 32, 195–198. [Google Scholar] [CrossRef]
- Muñoz-Ibarra, E.; Molina-López, R.A.; Durán, I.; Garcias, B.; Martín, M.; Darwich, L. Antimicrobial resistance in bacteria isolated from exotic pets: The situation in the Iberian Peninsula. Animals 2022, 12, 1912. [Google Scholar] [CrossRef]
- Ladyman, J.M.; Kuchling, G.; Burford, D.; Boardman, W.; Raidal, S.R. Skin disease affecting the conservation of the western swamp tortoise (Pseudemydura umbrina). Aust. Vet. J. 1998, 76, 743–745. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wu, Q.; Qin, X.; Yang, C.; Luo, S.; He, J.; Cheng, Q.; Wu, Z. Identification of Pseudomonas aeruginosa from the skin ulcer disease of crocodile lizards (Shinisaurus crocodilurus) and probiotics as the control measure. Front. Vet. Sci. 2022, 9, 850684. [Google Scholar] [CrossRef] [PubMed]
- Seixas, R.; Pissarra, H.; Santos, J.; Bernardino, R.; Fernandes, T.; Correia, J.; Vilela, C.L.; Oliveira, M. Severe fibrinonecrotic enteritis caused by Pseudomonas aeruginosa in a captive monitor lizard (Varanus niloticus). J. Zoo Wildl. Med. 2014, 45, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Cristina, R.T.; Kocsis, R.; Dégi, J.; Muselin, F.; Dumitrescu, E.; Tirziu, E.; Herman, V.; Darău, A.P.; Oprescu, I. Pathology and Prevalence of Antibiotic-Resistant Bacteria: A Study of 398 Pet Reptiles. Animals 2022, 12, 1279. [Google Scholar] [CrossRef]
- Sala, A.; Di Ianni, F.; Pelizzone, I.; Bertocchi, M.; Santospirito, D.; Rogato, F.; Flisi, S.; Spadini, C.; Iemmi, T.; Moggia, E.; et al. The prevalence of Pseudomonas aeruginosa and multidrug resistant Pseudomonas aeruginosa in healthy captive ophidian. PeerJ 2019, 7, e6706. [Google Scholar] [CrossRef]
- Colinon, C.; Jocktane, D.; Brothier, E.; Rossolini, G.M.; Cournoyer, B.; Nazaret, S. Genetic analyses of Pseudomonas aeruginosa isolated from healthy captive snakes: Evidence of high inter- and intrasite dissemination and occurrence of antibiotic resistance genes. Environ. Microbiol. 2010, 12, 716–729. [Google Scholar] [CrossRef]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella serovars isolated from reptiles in Norwegian zoos. Acta Vet. Scand. 2020, 62, 3. [Google Scholar] [CrossRef]
- McWhorter, A.; Owens, J.; Valcanis, M.; Olds, L.; Myers, C.; Smith, I.; Trott, D.; McLelland, D. In vitro invasiveness and antimicrobial resistance of Salmonella enterica subspecies isolated from wild and captive reptiles. Zoonoses Public Health 2021, 68, 402–412. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: A review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef]
- Cota, J.B.; Carvalho, A.C.; Dias, I.; Reisinho, A.; Bernardo, F.; Oliveira, M. Salmonella spp. in pet reptiles in Portugal: Prevalence and chlorhexidine gluconate antimicrobial efficacy. Antibiotics 2021, 10, 324. [Google Scholar] [CrossRef]
- Lukac, M.; Pedersen, K.; Prukner-Radovcic, E. Prevalence of Salmonella in captive reptiles from Croatia. J. Zoo Wildl. Med. 2015, 46, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Merkevičienė, L.; Butrimaitė-Ambrozevičienė, Č.; Paškevičius, G.; Pikūnienė, A.; Virgailis, M.; Dailidavičienė, J.; Daukšienė, A.; Šiugždinienė, R.; Ruzauskas, M. Serological variety and antimicrobial resistance in Salmonella isolated from reptiles. Biology 2022, 11, 836. [Google Scholar] [CrossRef] [PubMed]
- Hydeskov, H.B.; Guardabassi, L.; Aalbaek, B.; Olsen, K.E.; Nielsen, S.S.; Bertelsen, M.F. Salmonella prevalence among reptiles in a zoo education setting. Zoonoses Public Health 2013, 60, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Corrente, M.; Madio, A.; Friedrich, K.G.; Greco, G.; Desario, C.; Tagliabue, S.; D’Incau, M.; Campolo, M.; Buonavoglia, C. Isolation of Salmonella strains from reptile faeces and comparison of different culture media. J. Appl. Microbiol. 2004, 96, 709–715. [Google Scholar] [CrossRef]
- Monzón Moreno, C.; Ojeda Vargas, M.M.; Echeita, A.; Usera, M.A. Occurrence of Salmonella in cold-blooded animals in Gran Canaria, Canary Islands, Spain. Antonie Van Leeuwenhoek 1995, 68, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Santana-Hernández, K.M.; Rodríguez-Ponce, E.; Medina, I.R.; Acosta-Hernández, B.; Priestnall, S.L.; Vega, S.; Marin, C.; Cerdà-Cuéllar, M.; Marco-Fuertes, A.; Ayats, T.; et al. One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island. Animals 2023, 13, 1790. [Google Scholar] [CrossRef]
- Cristina, R.T.; Janos, D. Multiresistant Staphylococcus intermedius isolated from otitis externa in dogs and them human owners—A practical approach. Afr. J. Pharm. Pharmacol. 2013, 7, 1351–1356. [Google Scholar] [CrossRef]
- Graveland, H.; Duim, B.; van Duijkeren, E.; Heederik, D.; Wagenaar, J.A. Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int. J. Med. Microbiol. 2011, 301, 630–634. [Google Scholar] [CrossRef]
- Fitzgerald, J.R. Evolution of Staphylococcus aureus during human colonization and infection. Infect. Genet. Evol. 2014, 21, 542–547. [Google Scholar] [CrossRef]
- Espinosa-Gongora, C.; Chrobak, D.; Moodley, A.; Bertelsen, M.F.; Guardabassi, L. Occurrence and distribution of Staphylococcus aureus lineages among zoo animals. Vet. Microbiol. 2012, 158, 228–231. [Google Scholar] [CrossRef]
- Severn, M.M.; Williams, M.R.; Shahbandi, A.; Bunch, Z.L.; Lyon, L.M.; Nguyen, A.; Zaramela, L.S.; Todd, D.A.; Zengler, K.; Cech, N.B.; et al. The ubiquitous human skin commensal Staphylococcus hominis protects against opportunistic pathogens. mBio 2022, 13, e0093022. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef]
- Uddin, O.; Hurst, J.; Alkayali, T.; Schmalzle, S.A. Staphylococcus hominis cellulitis and bacteremia associated with surgical clips. IDCases 2022, 27, e01436. [Google Scholar] [CrossRef]
- Aykut, A.; Sevik, M.O.; Şan, B.; Şahin, Ö. Staphylococcus hominis: A rare cause of endophthalmitis. Arq. Bras. Oftalmol. 2022, 86, 281–283. [Google Scholar] [CrossRef] [PubMed]
- El-Jakee, J.K.; Aref, N.E.; Gomaa, A.; El-Hariri, M.D.; Galal, H.M.; Omar, S.A.; Samir, A. Emerging of coagulase negative staphylococci as a cause of mastitis in dairy animals: An environmental hazard International. J. Vet. Med. Sci. 2013, 1, 74–78. [Google Scholar] [CrossRef]
- Loncaric, I.; Tichy, A.; Handler, S.; Szostak, M.P.; Tickert, M.; Diab-Elschahawi, M.; Spergser, J.; Künzel, F. Prevalence of methicillin-resistant Staphylococcus sp. (MRS) in different companion animals and determination of risk factors for colonization with MRS. Antibiotics 2019, 8, 36. [Google Scholar] [CrossRef]
- Ocloo, R.; Nyasinga, J.; Munshi, Z.; Hamdy, A.; Marciniak, T.; Soundararajan, M.; Newton-Foot, M.; Ziebuhr, W.; Shittu, A.; Revathi, G.; et al. Epidemiology and antimicrobial resistance of staphylococci other than Staphylococcus aureus from domestic animals and livestock in Africa: A systematic review. Front. Vet. Sci. 2022, 9, 1059054. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D.; Pruzzo, C.; Vezzulli, L.; Kaper, J.B. Vibrio Species. In Food Microbiology: Fundamentals and Frontiers, 4th ed.; Doyle, M.P., Buchanan, R.L., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 401–439. [Google Scholar]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental occurrence and clinical impact of Vibrio vulnificus and Vibrio parahaemolyticus: A European perspective. Environ. Microbiol. Rep. 2010, 2, 7–18. [Google Scholar] [CrossRef]
- Fernández, F.S.; Alonso, G. Cólera y Vibrio cholerae [Cholera and Vibrio cholerae]. INHRR 2009, 40, 50–69. [Google Scholar]
- Magnino, S.; Colin, P.; Dei-Cas, E.; Madsen, M.; McLauchlin, J.; Nöckler, K.; Maradona, M.P.; Tsigarida, E.; Vanopdenbosch, E.; Van Peteghem, C. Biological risks associated with consumption of reptile products. Int. J. Food Microbiol. 2009, 134, 163–175. [Google Scholar] [CrossRef]
- Lopardo, A.H. Microbiología de las infecciones posteriores a mordeduras [Microbiology of post-bite infections. Med. Infant. 2018, 24, 38–45. [Google Scholar]
- Wang, J.; Yan, M.; Gao, H.; Lu, X.; Kan, B. Vibrio cholerae colonization of soft-shelled turtles. Appl. Environ. Microbiol. 2017, 83, 713–717. [Google Scholar] [CrossRef]
- Abreu-Acosta, N.; Pino-Vera, R.; Izquierdo-Rodríguez, E.; Afonso, O.; Foronda, P. Zoonotic Bacteria in Anolis sp., an Invasive Species Introduced to the Canary Islands (Spain). Animals 2023, 13, 414. [Google Scholar] [CrossRef] [PubMed]
- Leon-Velarde, C.G.; Jun, J.W.; Skurnik, M. Yersinia phages and food safety. Viruses 2019, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- Terech-Majewska, E.; Pajdak, J.; Platt-Samoraj, A.; Szczerba-Turek, A.; Bancerz-Kisiel, A.; Grabowska, K. Characterization of Yersinia enterocolitica strains potentially virulent for humans and animals in river water. J. Appl. Microbiol. 2016, 121, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Shayegani, M.; Stone, W.B.; DeForge, I.; Root, T.; Parsons, L.M.; Maupin, P. Yersinia enterocolitica and related species isolated from wildlife in New York State. Appl. Environ. Microbiol. 1986, 52, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Syczyło, K.; Platt-Samoraj, A.; Bancerz-Kisiel, A.; Szczerba-Turek, A.; Lipczyńska, K.; Jabłoński, A.; Procajło, Z.; Szweda, W. Monitoring of Yersinia enterocolitica strains from free-living animals using different methods. Pol. J. Vet. Sci. 2016, 19, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Viney, M.E.; Graham, A.L. Patterns and processes in parasite co-infection. Adv. Parasitol. 2013, 82, 321–369. [Google Scholar] [PubMed]





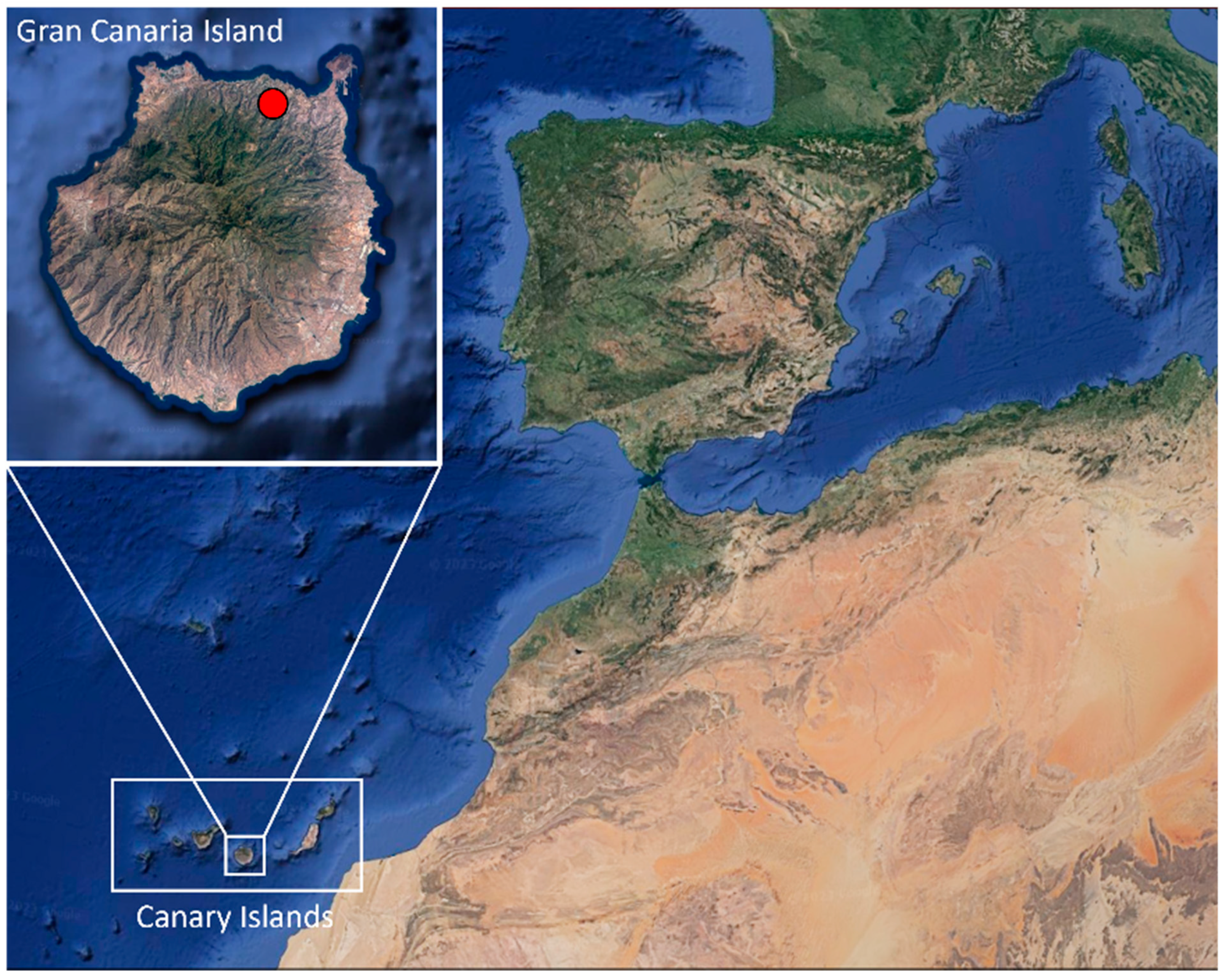{kind=link}
{kind=link}
{kind=link}
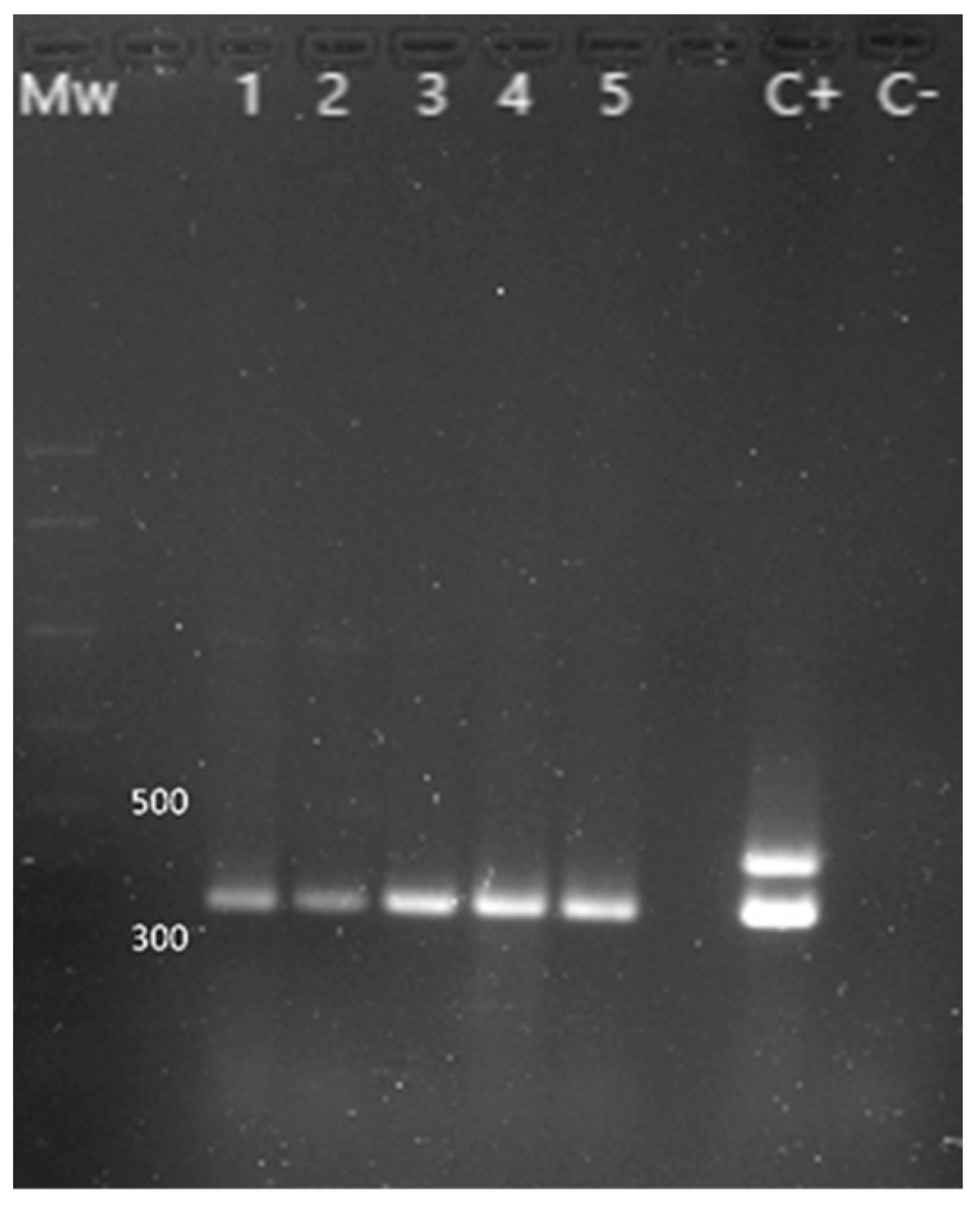{kind=link}
| Bacteria | No. of Positive Isolates (n) | Prevalence (%) |
|---|---|---|
| Campylobacter spp. | 2 (40) | 5.0 |
| Escherichia coli (eae, stx1, and stx2 genes) | 3 (40) | 7.5 |
| Listeria monocytogenes | 2 (40) | 5.0 |
| Mycobacterium spp. | 5 (40) | 12.5 |
| Pseudomonas spp. | 13 (40) | 32.5 |
| Salmonella spp. | 16 (40) | 40.0 |
| Staphylococcus spp. | 7 (40) | 17.5 |
| Vibrio spp. | 1 (40) | 2.5 |
| Yersinia enterocolitica | 11 (21) | 52.4 |
| Bacteria | Positive Males (Prevalence) n = 11 | Positive Females (Prevalence) n = 21 | Positive Juveniles (Prevalence) n = 4 | Total Positive Individuals (Prevalence) N = 40 |
|---|---|---|---|---|
| Pseudomonas spp. | 5 (45.5%) | 7 (33.3%) | 1 (25.0%) | 13 (32.5%) |
| Pseudomonas aeruginosa | 3 (27.3%) | 5 (23.8%) | 0 | 8 (20%) |
| Bacteria | Positive Males (Prevalence) n = 11 | Positive Females (Prevalence) n = 21 | Indetermined Sex (Prevalence) n = 4 | Positive Juveniles (Prevalence) n = 4 | Total Positive Individuals (Prevalence) N = 40 |
|---|---|---|---|---|---|
| Salmonella spp. | 4 (36.4%) | 9 (42.9%) | 1 (25.0%) | 2 (50.0%) | 15 (37.5%) |
| S. Typhi | 0 | 3 (14.3%) | 0 | 0 | 3 (7.5%) |
| S. Typhimurium | 1 (9.1%) | 2 (9.52%) | 0 | 0 | 3 (7.5%) |
| S. Enteritidis | 0 | 0 | 0 | 0 | 0 |
| Bacteria | Positive Males (Prevalence) n = 11 | Positive Females (Prevalence) n = 21 | Positive Juveniles (Prevalence) n = 4 | Total Positive Individuals (Prevalence) N = 40 |
|---|---|---|---|---|
| Staphylococcus spp. | 1 (9.1%) | 5 (23.8%) | 1 (25.0%) | 7 (17.5%) |
| S. hominis | 0 | 1 (4.8%) | 0 | 1 (2.5%) |
| Methicillin resistance gene | 1 (9.1%) | 2 (9.5%) | 1 (25.0%) | 4 (10.0%) |
| Mupirocin resistance gene | 0 | 2 (9.5%) | 1 (25.0%) | 3 (7.5%) |
| No. of Animals with Co-Infections | Identified Bacteria/Virulence Genes | Percentage% of Co-Infections (n = 36) |
|---|---|---|
| 7 | Pseudomonas spp. + Salmonella spp. | 19.4 |
| 5 | Salmonella spp. + Staphylococcus spp. | 13.9 |
| 3 | Mycobacterium spp. + Pseudomonas spp. | 8.3 |
| 3 | Mycobacterium spp. + Salmonella spp. | 8.3 |
| 3 | Pseudomonas spp. + Staphylococcus spp. | 8.3 |
| 2 | L. monocytogenes + Staphylococcus spp. | 5.6 |
| 2 | S. Typhi + stx1 gene | 5.6 |
| 1 | C. lari + Mycobacterium spp. | 2.8 |
| 1 | eae gene + Mycobacterium spp. | 2.8 |
| 1 | eae gene + P. aeruginosa | 2.8 |
| 1 | eae gene + S. Typhimurium | 2.8 |
| 1 | L. monocytogenes + P. aeruginosa | 2.8 |
| 1 | L. monocytogenes + Vibrio spp. | 2.8 |
| 1 | Mycobacterium spp. + methicillin-resistant Staphylococcus sp. | 2.8 |
| 1 | P. aeruginosa + Vibrio spp. | 2.8 |
| 1 | S. hominis + P. aeruginosa | 2.8 |
| 1 | methicillin-resistant Staphylococcus sp. + stx1 gene | 2.8 |
| Identified Bacteria/Virulence Genes | Co-Infection Index (Ic) | p-Value |
|---|---|---|
| Pseudomonas spp. + Salmonella spp. | 8.2 | 0.215 |
| Salmonella spp + Staphylococcus spp. | 17.8 | 0.094 |
| Mycobacterium spp. + Pseudomonas spp. | 9.2 | 0.307 |
| Mycobacterium spp. + Salmonella spp. | 5.6 | 0.373 |
| Pseudomonas spp. + Staphylococcus spp. | 4.3 | 0.662 |
| L. monocytogenes + Staphylococcus spp. | 21.4 | 0.027 |
| S. Typhi + stx1 gene | 61.7 | 0.004 |
| C. lari + Mycobacterium spp. | 17.5 | 0.125 |
| eae gene + Mycobacterium spp. | 17.5 | 0.125 |
| eae gene + P. aeruginosa | 10.0 | 0.200 |
| eae gene + S. Typhimurium | 30.8 | 0.075 |
| L.monocytogenes + P. aeruginosa | 6.7 | 0.364 |
| L.monocytogenes + Vibrio spp. | 47.5 | 0.050 |
| Mycobacterium spp. + methicillin-resistant Staphylococcus sp. | 6.3 | 0.427 |
| P. aeruginosa + Vibrio spp. | 10.0 | 0.200 |
| S. hominis + P. aeruginosa | 10.0 | 0.200 |
| Methicillin-resistant Staphylococcus sp. + stx1 gene | 16.0 | 0.192 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pino-Vera, R.; Abreu-Acosta, N.; Foronda, P. Study of Zoonotic Pathogens in Alien Population of Veiled Chameleons (Chamaeleo calyptratus) in the Canary Islands (Spain). Animals 2023, 13, 2288. https://doi.org/10.3390/ani13142288
Pino-Vera R, Abreu-Acosta N, Foronda P. Study of Zoonotic Pathogens in Alien Population of Veiled Chameleons (Chamaeleo calyptratus) in the Canary Islands (Spain). Animals. 2023; 13(14):2288. https://doi.org/10.3390/ani13142288
Chicago/Turabian StylePino-Vera, Román, Néstor Abreu-Acosta, and Pilar Foronda. 2023. "Study of Zoonotic Pathogens in Alien Population of Veiled Chameleons (Chamaeleo calyptratus) in the Canary Islands (Spain)" Animals 13, no. 14: 2288. https://doi.org/10.3390/ani13142288
APA StylePino-Vera, R., Abreu-Acosta, N., & Foronda, P. (2023). Study of Zoonotic Pathogens in Alien Population of Veiled Chameleons (Chamaeleo calyptratus) in the Canary Islands (Spain). Animals, 13(14), 2288. https://doi.org/10.3390/ani13142288