
Prevalence of Toxoplasma gondii in Endangered Wild Felines (Felis silvestris and Lynx pardinus) in Spain

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
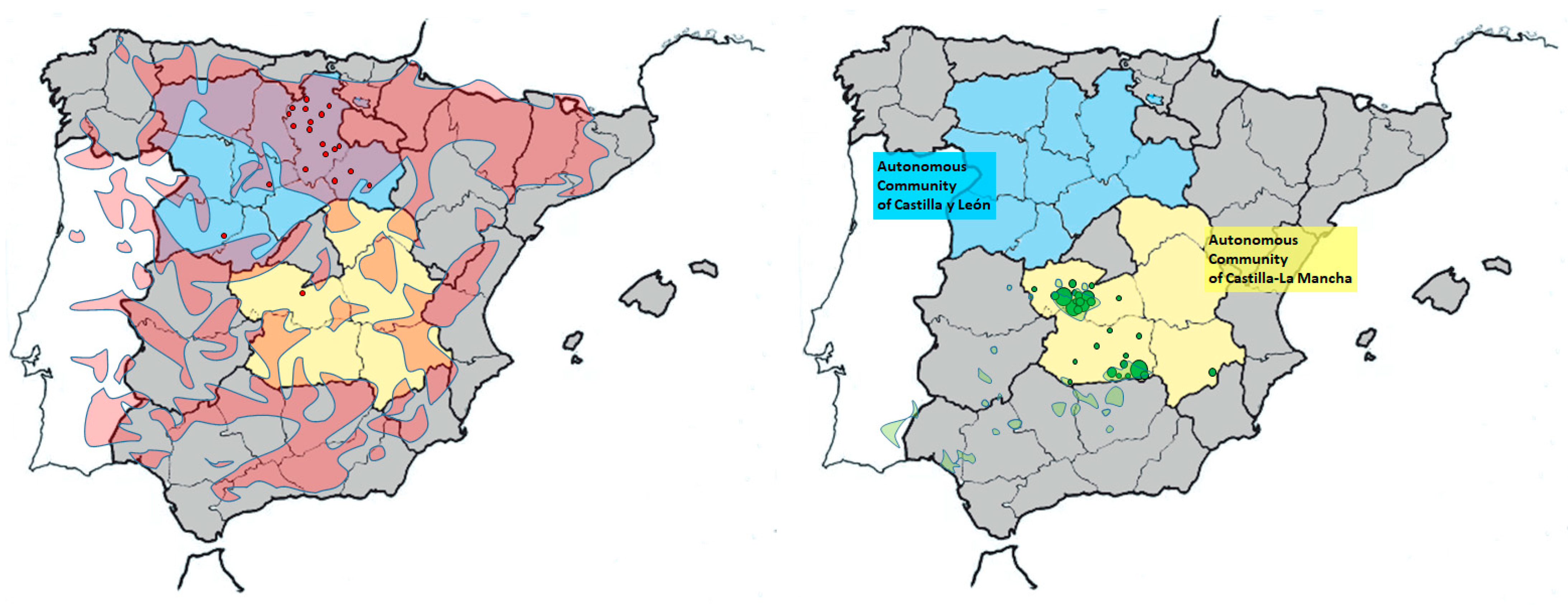
2.1. Study Area and Animals
2.2. Sample Collection and Processing
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 3rd ed.; CRC Press: New York, NY, USA, 2021; p. 564. [Google Scholar] [CrossRef]
- Dardé, M.-L.; Peyron, F. Toxoplasma y Toxoplasmosis. EMC-Pediatr. 2013, 48, 1–12. [Google Scholar] [CrossRef]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.A.; Kwok, O.C.H.; Yang, Y.R.; Su, C. All about toxoplasmosis in cats: The last decade. Vet. Parasitol. 2020, 283, 109145. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Shapiro, K.; Van Wormer, E. Dynamics and epidemiology of Toxoplasma gondii oocyst shedding in domestic and wild felids. Transbound. Emerg. Dis. 2022, 69, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Dubey, J.P. Toxoplasma gondii: Transmission, diagnosis and prevention. Clin. Microbiol. Infect. 2002, 8, 634–640. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.R.; Weis, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Scherrer, P.; Ryser-Degiorgis, M.P.; Marti, I.A.; Borel, S.; Frey, C.F.; Mueller, N.; Ruetten, M.; Basso, W. Exploring the epidemiological role of the Eurasian lynx (Lynx lynx) in the life cycle of Toxoplasma gondii. Int. J. Parasitol. Parasites Wildl. 2023, 21, 1–10. [Google Scholar] [CrossRef]
- Vanwormer, E.; Conrad, P.A.; Miller, M.A.; Melli, A.C.; Carpenter, T.E.; Mazet, J.A. Toxoplasma gondii, source to sea: Higher contribution of domestic felids to terrestrial parasite loading despite lower infection prevalence. Ecohealth 2013, 10, 277–289. [Google Scholar] [CrossRef]
- Teutsch, S.M.; Juranek, D.D.; Sulzer, A.; Dubey, J.P.; Sikes, R.K. Epidemic toxoplasmosis associated with infected cats. N. Engl. J. Med. 1979, 300, 695–699. [Google Scholar] [CrossRef]
- Vollaire, M.R.; Radecki, S.V.; Lappin, M.R. Seroprevalence of Toxoplasma gondii antiboides in clinically ill cats in the United States. Am. J. Vet. Res. 2005, 66, 874–877. [Google Scholar] [CrossRef]
- Opsteegh, M.; Haveman, R.; Swart, A.N.; Mensik-Beerepot, M.E.; Hofhuis, A.; Lagelaar, M.F.M.; van der Giessen, J.W.B. Seroprevalence and risk factors for Toxoplasma gondii infection in domestic catas in The Netherlands. Prev. Vet. Med. 2012, 104, 317–326. [Google Scholar] [CrossRef]
- Ding, H.; Gao, Y.M.; Lamberton, P.H.L.; Lu, D.B. A systematic review and meta-analysis of the seroprevalence of Toxoplasma gondii in catas in mainland China. Parasit. Vectors 2017, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Gauss, C.B.; Almería, S.; Ortuño, A.; Garcia, F.; Dubey, J.P. Seroprevalence of Toxoplasma gondii antibodies in domestic cats from Barcelona, Spain. J. Parasitol. 2003, 89, 1067–1068. [Google Scholar] [CrossRef] [PubMed]
- Montoya Matute, A. La Infección por Toxoplasma gondii en el Gato: Aspectos Epidemiológicos, Diagnóstico y Caracterización de Aislados Autóctonos. PhD. Thesis, Complutense University, Madrid, Spain, 2007. [Google Scholar]
- Millán, J.; Cabezón, O.; Pabón, M.; Dubey, J.P.; Almería, S. Seroprevalence of Toxoplasma gondii and Neospora caninum in feral cats (Felis silvestris catus) in Majorca, Balearic Islands, Spain. Vet. Parasitol. 2009, 165, 323–326. [Google Scholar] [CrossRef]
- Sherry, K.; Miró, G.; Trotta, M.; Miranda, C.; Montoya, A.; Espinosa, C.; Ribas, F.; Furlanello, T.; Solano-Gallego, L. A Serological and Molecular Study of Leishmania infantum Infection in Cats from the Island of Ibiza (Spain). Vector Borne Zoonotic Dis. 2011, 11, 239–245. [Google Scholar] [CrossRef]
- Miró, G.; Ruperez, C.; Checa, R.; Galvez, R.; Hernandez, L.; Garcia, M.; Canorea, I.; Marino, V.; Montoya, A. Current status of L. infantum infection in stray cats in the Madrid region (Spain): Implications for the recent outbreak of human leishmaniosis? Parasit. Vectors 2014, 7, 112. [Google Scholar] [CrossRef]
- Montoya, A.; García, M.; Gálvez, R.; Checa, R.; Marino, V.; Sarquis, J.; Barrera, J.P.; Rupérez, C.; Caballero, L.; Chicharro, C.; et al. Implications of zoonotic and vector-borne parasites to free-roaming cats in central Spain. Vet. Parasitol. 2018, 251, 125–130. [Google Scholar] [CrossRef]
- Candela, M.G.; Fanelli, A.; Carvalho, J.; Serrano, E.; Domenech, G.; Alonso, F.; Martínez-Carrasco, C. Urban landscape and infection risk in free-roaming cats. Zoonoses Public Health 2022, 69, 295–311. [Google Scholar] [CrossRef]
- Villanueva-Saz, S.; Martínez, M.; Giner, J.; González, A.; Tobajas, A.P.; Pérez, M.D.; Lira-Navarrete, E.; González-Ramírez, A.M.; Macías-León, J.; Verde, M.; et al. A cross-sectional serosurvey of SARS-CoV-2 and co-infections in stray cats from the second wave to the sixth wave of COVID-19 outbreaks in Spain. Vet. Res. Commun. 2023, 47, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, R.; Cabezón, O.; Millán, J.; Pabón, M.; Arnal, M.C.; Luco, D.F.; Gortázar, C.; Dubey, J.P.; Almeria, S. Seroprevalence of Toxoplasma gondii antibodies in wild carnivores from Spain. Vet. Parasitol. 2007, 148, 187–192. [Google Scholar] [CrossRef]
- Roelke, M.E.; Johnson, W.E.; Millán, J.; Palomares, F.; Revilla, E.; Rodríguez, A.; Calzada, J.; Ferreras, P.; León-Vizcaíno, L.; Delibes, M.; et al. Exposure to disease agents in the endangered Iberian lynx (Lynx pardinus). Eur. J. Wildl. Res. 2008, 54, 171–178. [Google Scholar] [CrossRef]
- Millán, J.; Candela, M.G.; Palomares, F.; Cubero, M.J.; Rodríguez, A.; Barral, M.; de la Fuente, J.; Almería, S.; León-Vizcaíno, L. Disease threats to the endangered Iberian lynx (Lynx pardinus). Vet. J. 2009, 182, 114–124. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Dubey, J.P.; Martínez, F.; Vargas, A.; Cabezón, O.; Zorrilla, I.; Arenas, A.; Almería, S. Factors affecting seroprevalence of Toxoplasma gondii in the endangered Iberian lynx (Lynx pardinus). Vet. Parasitol. 2010, 167, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Anile, S.; Bizzarri, L.; Lacrimini, M.; Sforzi, A.; Ragni, B.; Devillard, S. Home-Range Size of the European Wildcat (Felis silvestris silvestris): A Report from Two Areas in Central Italy. Mammalia 2017, 82, 1–11. [Google Scholar] [CrossRef]
- Gil-Sanchez, J.M.; Barea-Azcon, J.M.; Jaramillo, J.; Virgos, E.; Herrera-Sanchez, F.J.; Jiménez, J.; Virgós, E. Fragmentation and low density as major conservation challenges for the southernmost populations of the European wildcat. PLoS ONE 2020, 15, e0227708. [Google Scholar] [CrossRef]
- López-Martín, J.M.; García, F.J.; Such, A.; Virgós, E.; Lozano, J.; Duarte, J.; España, A.J. Felis silvestris Schreber, 1777. In Atlas y Libro Rojo de los Mamíferos Terrestres de España; Palomo, L.J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 333–338. [Google Scholar]
- Guzmán, J.N.; García, F.J.; Garrote, G.; Pérez de Ayala, R.; Iglesias, M.C. El lince ibérico (Lynx pardinus) en España y Portugal. Censo-Diagnóstico de sus Poblaciónes; Dirección General para la Biodiversidad, Ministerio de Medio Ambiente: Madrid, Spain, 2004; p. 184. [Google Scholar]
- Anonymous. Censo de Lince Ibérico España y Portugal. 2022. Available online: https://lifelynxconnect.eu/en/census_en/ (accessed on 12 July 2023).
- Klar, N.; Herrmann, M.; Kramer-Schadt, S. Effects and mitigation of road impacts on individual movement behavior of wildcats. J. Wildl. Manag. 2009, 73, 631–638. [Google Scholar] [CrossRef]
- Meli, M.L.; Cattori, V.; Martínez, F.; López, G.; Vargas, A.; Simón, M.A.; Zorrilla, I.; Muñoz, A.; Palomares, F.; López-Bao, J.V.; et al. Feline leukemia virus and other pathogens as important threats to the survival of the critically endangered Iberian lynx (Lynx pardinus). PLoS ONE 2009, 4, e4744. [Google Scholar] [CrossRef]
- Lozano, J.; Malo, A.F. Conservation of the European wildcat (Felis silvestris) in Mediterranean environments: A reassessment of current threats. In Mediterranean Ecosystems: Dynamics, Management and Conservation; Williams, G.S., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 1–31. [Google Scholar]
- Anile, S.; Devillard, S.; Ragni, B.; Rovero, F.; Mattucci, F.; Valvo, M.L. Habitat fragmentation and anthropogenic factors affect wildcat Felis silvestris silvestris occupancy and detectability on Mt Etna. Wildl. Biol. 2019, 1, 1–13. [Google Scholar] [CrossRef]
- Candela, M.G.; Pardavila, X.; Ortega, N.; Lamosa, A.; Mangas, J.G.; Martínez-Carrasco, C. Canine distemper virus may affect European wild cat populations in Central Spain. Mamm. Biol. 2019, 97, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Girling, S.J.; Pizzi, R.; Naylor, A.D.; Richardson, D.; Richardson, U.; Harley, J.; Cole, G.; Brown, D.; Fraser, M.; Tillman, E.; et al. Use of clindamycin in Pallas’ cats (Otocolobus (Felis) manul) to reduce juvenile toxoplasmosis-associated mortality rates. J. Zoo Wildl. Med. 2020, 51, 39–45. [Google Scholar] [CrossRef]
- Pas, A.; Dubey, J.P. Fatal toxoplasmosis in sand cats (Felis margarita). J. Zoo Wildl. Med. 2008, 39, 362–369. [Google Scholar] [CrossRef]
- Dubey, J.P.; Quinn, W.J.; Weinandy, D. Fatal neonatal toxoplasmosis in a bobcat (Lynx rufus). J. Wildl. Dis. 1987, 23, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Lozano Mendoza, J. Ecología del Gato Montés (Felis silvestris) y su Relación con el Conejo de Monte (Oryctolagus cuniculus). Ph.D. Thesis, Complutense University, Madrid, Spain, 2008. [Google Scholar]
- Gerngross, P.; Ambarli, H.; Angelici, F.M.; Anile, S.; Campbell, R.; Ferreras de Andres, P.; Gil-Sanchez, J.M.; Götz, M.; Jerosch, S.; Mengüllüoglu, D.; et al. Felis silvestris. The ICUM Red List of Threatened Species 2022: e.T181049859A181050999. Available online: https://www.iucnredlist.org/species/181049859/181050999 (accessed on 28 July 2023).
- Verhofstede, C.; Sabbe, L.; Van Renterghem, L. Ability of enzyme-linked immunosorbent assays to detect early immunoglobulin G antibodies to Toxoplasma gondii. Eur. J. Clin. Microbiol. 1987, 6, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Thienpont, D.; Rochette, F.; Vanparijs, F.J. Diagnostic de Verminose par Examen Coprologique; Janssen Research Foundation: Beerse, Belgium, 1979; pp. 35–36. [Google Scholar]
- European Scientific Counsel Companion Animal Parasites (ESCCAP). Parasitological Diagnosis in Cats, Dogs and Equines. In ESCCAP Guideline 04 First Edition; ESCCAP: Worcestershire, UK, 2022; p. 19. [Google Scholar]
- Fuentes, I.; Rodríguez, M.; Domingo, C.D.; del Castillo, F.; Juncosa, T.; Alvar, J. Urine sample used for congenital toxoplasmosis diagnosis by PCR. J. Clin. Microbiol. 1996, 34, 2368–2371. [Google Scholar] [CrossRef] [PubMed]
- Homan, W.L.; Vercammen, M.; De Braekeleer, J.; Verschueren, H. Identification of a 200- to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. Int. J. Parasitol. 2000, 30, 69–75. [Google Scholar] [CrossRef]
- Kim, J.H.; Sohn, H.J.; Hwang, W.S.; Hwang, E.K.; Jean, Y.H.; Yamane, I.; Kim, D.Y. In vitro isolation and characterization of bovine Neospora caninum in Korea. Vet. Parasitol. 2000, 90, 147–154. [Google Scholar] [CrossRef]
- Dubey, J.P. Duration of immunity to shedding of Toxoplasma gondii oocysts by cats. J. Parasitol. 1995, 81, 410–415. [Google Scholar] [CrossRef]
- Bolais, P.F.; Galal, L.; Cronemberger, C.M.; Pereira, F.A.; Barbosa, A.D.S.; Dib, L.V.; Amendoeira, M.R.R.; Dardé, M.L.; Mercier, A. Toxoplasma gondii in the faeces of wild felids from the Atlantic Forest, Brazil. Mem. Inst. Oswaldo Cruz 2022, 117, e210302. [Google Scholar] [CrossRef]
- Diakou, A.; Migli, D.; Dimzas, D.; Morelli, S.; Di Cesare, A.; Youlatos, D.; Lymberakis, P.; Traversa, D. Endoparasites of European Wildcats (Felis silvestris) in Greece. Pathogens 2021, 10, 594. [Google Scholar] [CrossRef]
- Dubey, J.P. The evolution of the knowledge of cat and dog coccidia. Parasitology 2009, 136, 1469–1475. [Google Scholar] [CrossRef]
- Dubey, J.P.; Sreekumar, C. Redescription of Hammondia hammondi and its differentiation from Toxoplasma gondii. Int. J. Parasitol. 2003, 33, 1437–1453. [Google Scholar] [CrossRef]
- Izadi, L.; Sarvi, S.; Hosseini, S.A.; Amouei, A.; Sharif, M.; Rahimi, M.T.; Nayeri, T.; Daryani, A. Copro-molecular diagnosis of the Toxoplasmatinae subfamily in dog and cat populations in northern Iran. Epidemiol. Health 2020, 42, e2020074. [Google Scholar] [CrossRef]
- Dabritz, H.A.; Miller, M.A.; Atwill, E.R.; Gardner, I.A.; Leutenegger, C.M.; Melli, A.C.; Conrad, P.A. Detection of Toxoplasma gondii-like oocysts in cat feces and estimates of the environmental oocyst burden. J. Am. Vet. Med. Assoc. 2007, 231, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Montazeri, M.; Mikaeili Galeh, T.; Moosazadeh, M.; Sarvi, S.; Dodangeh, S.; Javidnia, J.; Sharif, M.; Daryani, A. The global serological prevalence of Toxoplasma gondii in felids during the last five decades (1967–2017): A systematic review and meta-analysis. Parasit. Vectors 2020, 13, 82. [Google Scholar] [CrossRef] [PubMed]
- Sioutas, G.; Symeonidou, I.; Gelasakis, A.I.; Tzirinis, C.; Papadopoulos, E. Feline toxoplasmosis in Greece: A countrywide seroprevalence study and associated risk factors. Pathogens 2022, 11, 1511. [Google Scholar] [CrossRef]
- Sharma, R.; Parker, S.; Al-Adhami, B.; Bachand, N.; Jenkins, E. Comparison of tissues (heart vs. brain) and serological tests (MAT, ELISA and IFAT) for detection of Toxoplasma gondii in naturally infected wolverines (Gulo gulo) from the Yukon, Canada. Food Waterborne Parasitol. 2019, 15, e00046. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.D.; Frenkel, J.K. Prevalence of antibodies to Toxoplasma gondii in wild mammals of Missouri and east central Kansas: Biologic and ecologic considerations of transmission. J. Wildl. Dis. 1995, 31, 15–21. [Google Scholar] [CrossRef]
- Dubey, J.P. Comparative infectivity of oocysts and bradyzoites of Toxoplasma gondii for intermediate (mice) and definitive (cats) hosts. Vet. Parasitol. 2006, 140, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Calzada, J.; Guzmán, J.N.; Rodríguez, A. Lynx Pardinus (Temminck, 1827). In Atlas y Libro Rojo de Mamíferos Terrestres de España; Palomo, L.J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 342–347. [Google Scholar]
- Malo, A.F.; Lozano, J.; Huertas, D.L.; Virgos, E. A change of diet from rodents to rabbits (Oryctolagus cuniculus). Is the wildcat (Felis silvestris) a specialist predator? J. Zool. 2004, 263, 401–407. [Google Scholar] [CrossRef]
- Afonso, E.; Thulliez, P.; Pontier, D.; Gilot-Fromont, E. Toxoplasmosis in prey species and consequences for prevalence in feral cats: Not all prey species are equal. Parasitology 2007, 134, 1963–1971. [Google Scholar] [CrossRef]
- Gustafsson, K.; Uggla, A.; Järplid, B. Toxoplasma gondii infection in the mountain hare (Lepus timidus) and domestic rabbit (Oryctolagus cuniculus). I. Pathology. J. Comp. Pathol. 1997, 117, 351–360. [Google Scholar] [CrossRef]
- Sedlák, K.; Literák, I.; Pavlásek, I.; Benák, J. Susceptibility of common voles to experimental toxoplasmosis. J. Wildl. Dis. 2001, 37, 640–642. [Google Scholar] [CrossRef]
- Galeh, T.M.; Sarvi, S.; Montazeri, M.; Moosazadeh, M.; Nakhaei, M.; Shariatzadeh, S.A.; Daryani, A. Global Status of Toxoplasma gondii Seroprevalence in Rodents: A Systematic Review and Meta-Analysis. Front. Vet. Sci. 2020, 7, 461. [Google Scholar] [CrossRef] [PubMed]
- Castro-Scholten, S.; Cano-Terriza, D.; Jiménez-Ruiz, S.; Almería, S.; Risalde, M.A.; Vicente, J.; Acevedo, P.; Arnal, M.C.; Balseiro, A.; Gómez-Guillamón, F.; et al. Seroepidemiology of Toxoplasma gondii in wild ruminants in Spain. Zoonoses Public Health 2021, 68, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Escobar, M.; Calero-Bernal, R.; Regidor-Ceriillo, J.; Vallejo, R.; Benavides, J.; Collantes-Fernández, E.; Ortega-Mora, L.M. Isolation, genotyping, and mouse virulence characterization of Toxoplasma gondii from free ranging Iberian pigs. Front. Vet. Sci. 2020, 7, 604782. [Google Scholar] [CrossRef] [PubMed]
- Almería, S.; Calvete, C.; Pagés, A.; Gauss, C.; Dubey, J.P. Factors affecting the seroprevalence of Toxoplasma gondii infection in wild rabbits (Oryctolagus cuniculus) from Spain. Vet. Parasitol. 2004, 123, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Kornacka, A.; Cybulska, A.; Bień, J.; Goździk, K.; Moskwa, B. The usefulness of direct agglutination test, enzyme-linked immunosorbent assay and polymerase chain reaction for the detection of Toxoplasma gondii in wild animals. Vet. Parasitol. 2016, 228, 85–89. [Google Scholar] [CrossRef]
- Herrmann, D.C.; Maksimov, P.; Maksimov, A.; Sutor, A.; Schwarz, S.; Jaschke, W.; Schliephake, A.; Denzin, N.; Conraths, F.J.; Schares, G. Toxoplasma gondii in foxes and rodents from the German Federal States of Brandenburg and Saxony-Anhalt: Seroprevalence and genotypes. Vet. Parasitol. 2012, 185, 78–85. [Google Scholar] [CrossRef]
- Juránková, J.; Basso, W.; Neumayerová, H.; Baláž, V.; Jánová, E.; Sidler, X.; Deplazes, P.; Koudela, B. Brain is the predilection site of Toxoplasma gondii in experimentally inoculated pigs as revealed by magnetic capture and real-time PCR. Food Microbiol. 2014, 38, 167–170. [Google Scholar] [CrossRef]
- Koethe, M.; Straubinger, R.K.; Pott, S.; Bangoura, B.; Geuthner, A.C.; Daugschies, A.; Ludewig, M. Quantitative detection of Toxoplasma gondii in tissues of experimentally infected turkeys and in retail turkey products by magnetic-capture PCR. Food Microbiol. 2015, 52, 11–17. [Google Scholar] [CrossRef]
- Gisbert Algaba, I.; Verhaegen, B.; Jennes, M.; Rahman, M.; Coucke, W.; Cox, E.; Dorny, P.; Dierick, K.; De Craeye, S. Pork as a source of transmission of Toxoplasma gondii to humans: A parasite burden study in pig tissues after infection with different strains of Toxoplasma gondii as a function of time and different parasite stages. Int. J. Parasitol. 2018, 48, 555–560. [Google Scholar] [CrossRef]
- Dubey, J.P. Persistence of Toxoplasma gondii in the tissues of chronically infected cats. J. Parasitol. 1997, 63, 156–157. [Google Scholar]
- Rico-Torres, C.P.; del Viento-Camacho, A.; Caballero-Ortega, H.; Besné-Mérida, A.; Luna-Pastén, H.; Correa, D.; Palma-García, J.M. First isolation of Toxoplasma gondii from cats of Colima, Mexico: Tissue distribution and genetic characterization. Vet. Parasitol. 2015, 209, 125–128. [Google Scholar] [CrossRef]
- Hill, D.E.; Sreekumar, C.; Jones, J.; Dubey, J.P. Toxoplasma gondii. In Infectious Disease. Foodborne Diseases; Simjee, V.S., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 337–353. [Google Scholar] [CrossRef]
- Opsteegh, M.; Langelaar, M.; Sprong, H.; den Hartog, L.; De Craeye, S.; Bokken, G.; Ajzenberg, D.; Kijlstra, A.; van der Giessen, J. Direct detection and genotyping of Toxoplasma gondii in meat samples using magnetic capture and PCR. Int. J. Food Microbiol. 2010, 139, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Juránková, J.; Hůrková-Hofmannová, L.; Volf, J.; Baláž, V.; Piálek, J. Efficacy of magnetic capture in comparison with conventional DNA isolation in a survey of Toxoplasma gondii in wild house mice. Eur. J. Protistol. 2014, 50, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, B.; Ravel, A.; Leighton, P.; Stephen, C.; Iqbal, A.; Ndao, M.; Konecsni, K.; Fernando, C.; Jenkins, E. Foxes (Vulpes vulpes) as sentinels for parasitic zoonoses, Toxoplasma gondii and Trichinella nativa, in the northeastern Canadian Arctic. Int. J. Parasitol. Parasites Wildl. 2018, 7, 391–397. [Google Scholar] [CrossRef]


{kind=link}
| IFAT IgG | Felis silvestris n (%; 95 CI 1) | Lynx pardinus n (%; 95% CI) | p-Value |
|---|---|---|---|
| Negative | 3 (15.0; 0.0–30.6) | 38 (55.1; 26.5–49.4) | 0.001 |
| Positive—any titre | 17 (85.0; 69.3–100.0) | 31 (44.9; 33.1–56.6) | 0.002 |
| Positive—titre 1/80 | --- | 4 (5.8; 0.4–11.6) | 0.006 |
| Positive—titre 1/160 | 1 (5.0; 0.0–14.5) | 5 (7.2; 0.9–13.0) | 0.724 |
| Positive—titre 1/320 | 4 (20.0; 0.0–12.6) | 6 (8.7; 2.2–15.7) | 0.158 |
| Positive—titre ≥ 1/640 | 12 (60.0; 38.5–81.5) | 16 (23.2; 13.1–32.9) | 0.001 |
| Host Species | Serology (n = 89) | Coprology (n = 89) | Tissue Analysis (n = 80) | |||||
|---|---|---|---|---|---|---|---|---|
| IFAT IgG | IFAT IgM | Telemann | nPCR | qPCR | nPCR 1 | qPCR 1 | Total PCR+ (Any Method) 2 | |
| Felis silvestris | 17 3 | 1 3 | 1 3 | 1 3 | 1 3 | 8 3 | 6 | 10 |
| Lynx pardinus | 31 | 0 | 0 | 0 | 0 | 9 | 3 | 11 |
| Total (both species) | 48 | 1 | 1 | 1 | 1 | 17 | 9 | 21 |
| Felis silvestris | Lynx pardinus | Combined Data from Both Species | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | n+ | % Positive | p-Value | n+ | % Positive | p-Value | n+ | % Positive | p-Value |
| Sex | 0.347 | 0.864 | 0.795 | ||||||
| Male | 13 | 81.2 (13/16) | 21 | 45.6 (21/46) | 34 | 54.8 (34/62) | |||
| Female | 4 | 100 (4/4) | 10 | 43.5 (10/23) | 14 | 51.9 (14/27) | |||
| Age | n/a | 0.002 | 0.003 | ||||||
| Juvenile | --- | --- | 0 | 0.0(0/7) | 0 | 0.0 (0/7) | |||
| Adult | 17 | 85.0 (17/20) | 27 | 46.5 (27/58) | 44 | 56.4 (44/78) | |||
| Senior | --- | --- | 4 | 100.0 (4/4) | 4 | 100.0 (4/4) | |||
| Capture area | 0.015 | ||||||||
| Castilla-La Mancha | n/a | 0.919 | |||||||
| Albacete | --- | --- | 1 | 50.0 (1/2) | 1 | 50.0 (1/2) | |||
| Ciudad Real | 0 | --- | 10 | 41.6 (10/24) | 10 | 41.7 (10/24) | |||
| Toledo | 0 | 0.0 (0/1) | 20 | 46.5 (20/43) | 21 | 45.5 (21/44) | |||
| Total | 0 | 0.0 (0/1) | 31 | 44.9 (31/69) | 31 | 44.3 (31/70) | |||
| Castilla y León | <0.001 | --- | |||||||
| Burgos | 13 | 92.9 (13/14) | 0 | --- | 13 | 92.9 (13/14) | |||
| Salamanca | 0 | 0.0 (0/1) | 0 | --- | 0 | 0.0 (0/1) | |||
| Soria | 3 | 100.0 (3/3) | 0 | --- | 3 | 100 (3/3) | |||
| Valladolid | 1 | 100.0 (1/1) | 0 | --- | 1 | 100 (1/1) | |||
| Total | 17 | 89.5 (17/19) | 0 | 17 | 89.5 (17/19) | ||||
| Host Species | Region | Animals Examined | Serological Method | % Positive | Reference |
|---|---|---|---|---|---|
| Felis silvestris | Andalusia, Castilla-La Mancha, Cantabrian region | 6 | MAT (modified agglutination text) | 50 | [21] |
| Castilla-La Mancha | 9 | ELISA | 56 | [34] | |
| Castilla-La Mancha, Castilla y León | 20 | IFAT | 85 | Present study | |
| Lynx pardinus | Andalusia, Castilla-La Mancha | 27 | MAT | 82 | [21] |
| Andalusia | 48 | IHA (indirect haemagglutination) | 44 | [22] | |
| Andalusia | 26 | MAT | 81 | [23] | |
| Andalusia | 129 | MAT | 63 | [24] | |
| Castilla-La Mancha | 69 | IFAT | 45 | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matas Méndez, P.; Fuentes Corripio, I.; Montoya Matute, A.; Bailo Barroso, B.; Grande Gómez, R.; Apruzzese Rubio, A.; Ponce Gordo, F.; Mateo Barrientos, M. Prevalence of Toxoplasma gondii in Endangered Wild Felines (Felis silvestris and Lynx pardinus) in Spain. Animals 2023, 13, 2488. https://doi.org/10.3390/ani13152488
Matas Méndez P, Fuentes Corripio I, Montoya Matute A, Bailo Barroso B, Grande Gómez R, Apruzzese Rubio A, Ponce Gordo F, Mateo Barrientos M. Prevalence of Toxoplasma gondii in Endangered Wild Felines (Felis silvestris and Lynx pardinus) in Spain. Animals. 2023; 13(15):2488. https://doi.org/10.3390/ani13152488
Chicago/Turabian StyleMatas Méndez, Pablo, Isabel Fuentes Corripio, Ana Montoya Matute, Begoña Bailo Barroso, Rebeca Grande Gómez, Ariadna Apruzzese Rubio, Francisco Ponce Gordo, and Marta Mateo Barrientos. 2023. "Prevalence of Toxoplasma gondii in Endangered Wild Felines (Felis silvestris and Lynx pardinus) in Spain" Animals 13, no. 15: 2488. https://doi.org/10.3390/ani13152488
APA StyleMatas Méndez, P., Fuentes Corripio, I., Montoya Matute, A., Bailo Barroso, B., Grande Gómez, R., Apruzzese Rubio, A., Ponce Gordo, F., & Mateo Barrientos, M. (2023). Prevalence of Toxoplasma gondii in Endangered Wild Felines (Felis silvestris and Lynx pardinus) in Spain. Animals, 13(15), 2488. https://doi.org/10.3390/ani13152488