QTLs and Candidate Genes Associated with Semen Traits in Merino Sheep

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Phenotype Data
2.2. Genotype Data and Quality Control
2.3. Model for Analysis
2.3.1. Statistical Model
2.3.2. Identification of SNPs and Associated Candidate Genes
2.4. Heritability and Variance Component Estimation of Semen Phenotypes
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amiridis, G.S.; Fthenakis, G.C. Reproductive health management of sheep and goats. Preface. Anim. Reprod. Sci. 2012, 130, 125. [Google Scholar] [CrossRef] [PubMed]
- De Vries, A. Reproduction: Economics of reproductive efficiency. Rev. Colomb. Cienc. Pecu. 2017, 30, 218–221. [Google Scholar]
- Sell-Kubiak, E.B.; Dobrzanski, J.; Derks, M.F.L.; Lopes, M.; Szwaczkowski, T. Meta-analysis of SNPs determining litter traits in pigs. Genes 2022, 13, 1730. [Google Scholar] [CrossRef]
- Gebreselassie, G.; Berihulay, H.; Jiang, L.; Ma, Y. Review on Genomic Regions and Candidate Genes Associated with Economically Important Production and Reproduction Traits in Sheep (Ovies aries). Animals 2020, 10, 33. [Google Scholar] [CrossRef]
- Ma, L.; Cole, J.B.; Da, Y.; VanRaden, P.M. Symposium review: Genetics, genome-wide association study, and genetic improvement of dairy fertility traits. J. Dairy Sci. 2019, 102, 3735–3743. [Google Scholar] [CrossRef]
- Serrano, M.; Ramón, M.; Calvo, J.H.; Jiménez, M.Á.; Freire, F.; Vázquez, J.M.; Arranz, J.J. Genome-wide association studies for sperm traits in Assaf sheep breed. Animal 2021, 15, 100065. [Google Scholar] [CrossRef] [PubMed]
- Evans, G. Salamon’s Artificial Insemination of Sheep and Goats; Butterworths: Sydney, Australia, 1987. [Google Scholar]
- Sheep Genetics, Meat and Livestock Australia. Sheep Genetics. Available online: https://search.sheepgenetics.org.au/search/dashboard (accessed on 2 July 2023).
- Holler, T.L.; Dean, M.; Taylor, T.; Poole, D.H.; Thonney, M.L.; Thomas, D.L.; Pate, J.L.; Whitley, N.; Dailey, R.A.; Inskeep, E.K. Effects of service sire on prenatal mortality and prolificacy in ewes. J. Anim. Sci. 2014, 92, 3108–3115. [Google Scholar] [CrossRef]
- Aibazov, M.; Trukhachev, V.; Selionova, M.; Malorodov, V. Seasonal changes in testis size, testosterone levels and sperm production quality in meat rams. Reprod. Domest. Anim. 2022, 57, 1125–1135. [Google Scholar] [CrossRef]
- Zaher, H.A.; Alawaash, S.A.; Swelum, A.A. Effects of season and breed on the reproductive performance of sheep. J. Anim. Reprod. Biotechnol. 2020, 35, 149–154. [Google Scholar] [CrossRef]
- Guan, Y.; Malecki, I.A.; Hawken, P.A.R.; Linden, M.D.; Martin, G.B. Under-nutrition reduces spermatogenic efficiency and sperm velocity, and increases sperm DNA damage in sexually mature male sheep. Anim. Reprod. Sci. 2014, 149, 163–172. [Google Scholar] [CrossRef]
- Jiménez-Rabadán, P.; Soler, A.J.; Ramón, M.; García-Álvarez, O.; Maroto-Morales, A.; Iniesta-Cuerda, M.; Fernández-Santos, M.R.; Montoro, V.; Pérez-Guzmán, M.D.; Garde, J.J. Influence of semen collection method on sperm cryoresistance in small ruminants. Anim. Reprod. Sci. 2016, 167, 103–108. [Google Scholar] [CrossRef] [PubMed]
- van Tilburg, M.; Sousa, S.; Lobo, M.D.P.; Monteiro-Azevedo, A.C.O.M.; Azevedo, R.A.; Araújo, A.A.; Moura, A.A. Mapping the major proteome of reproductive fluids and sperm membranes of rams: From the cauda epididymis to ejaculation. Theriogenology 2021, 159, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Rickard, J.P.; Leahy, T.; Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Lynch, G.W.; Druart, X.; de Graaf, S.P. The identification of proteomic markers of sperm freezing resilience in ram seminal plasma. J. Proteom. 2015, 126, 303–311. [Google Scholar] [CrossRef]
- Akalın, P.P.; Bülbül, B.; Çoyan, K.; Başpınar, N.; Kırbaş, M.; Bucak, M.N.; Güngör, Ş.; Öztürk, C. Relationship of blood and seminal plasma ceruloplasmin, copper, iron and cadmium concentrations with sperm quality in Merino rams. Small Rumin. Res. 2015, 133, 135–139. [Google Scholar] [CrossRef]
- Hodge, M.J.; de Las Heras-Saldana, S.; Rindfleish, S.J.; Stephen, C.P.; Pant, S.D. Characterization of Breed Specific Differences in Spermatozoal Transcriptomes of Sheep in Australia. Genes 2021, 12, 203. [Google Scholar] [CrossRef]
- David, I.; Kohnke, P.; Lagriffoul, G.; Praud, O.; Plouarboué, F.; Degond, P.; Druart, X. Mass sperm motility is associated with fertility in sheep. Anim. Reprod. Sci. 2015, 161, 75–81. [Google Scholar] [CrossRef]
- Eppleston, J.; Salamon, S.; Moore, N.W.; Evans, G. The depth of cervical insemination and site of intrauterine insemination and their relationship to the fertility of frozen-thawed ram semen. Anim. Reprod. Sci. 1994, 36, 211–225. [Google Scholar] [CrossRef]
- Morris, L.H.; Johnson, W.H.; Leibo, S.P.; Buckrell, B.C. Relationship between the characteristics of frozen-thawed ram spermatozoa and in vitro embryo production. Reprod. Fertil. Dev. 2001, 13, 193–201. [Google Scholar] [CrossRef]
- Pelayo, R.; Ramón, M.; Granado-tajada, I.; Ugarte, E.; Serrano, M.; Gutiérrez-gil, B.; Arranz, J.-J. Estimation of the genetic parameters for semen traits in Spanish dairy sheep. Animals 2019, 9, 1147. [Google Scholar] [CrossRef] [PubMed]
- Hodge, M.J.; Rindfleish, S.J.; Heras-Saldana, S.D.L.; Stephen, C.P.; Pant, S.D. Heritability and Genetic Parameters for Semen Traits in Australian Sheep. Animals 2022, 12, 2946. [Google Scholar] [CrossRef]
- David, I.; Druart, X.; Lagriffoul, G.; Manfredi, E.; Robert-Granié, C.; Bodin, L. Genetic and environmental effects on semen traits in Lacaune and Manech tête rousse AI rams. Genet. Sel. Evol. 2007, 39, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Rege, J.E.O.; Toe, F.; Mukasa-Mugerwa, E.; Tembely, S.; Anindo, D.; Baker, R.L.; Lahlou-Kassi, A. Reproductive characteristics of Ethiopian highland sheep: II. Genetic parameters of semen characteristics and their relationships with testicular measurements in ram lambs. Small Rumin. Res. 2000, 37, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Talouarn, E.; Bardou, P.; Palhière, I.; Oget, C.; Clément, V.; Tosser-Klopp, G.; Rupp, R.; Robert-Granié, C. Genome wide association analysis on semen volume and milk yield using different strategies of imputation to whole genome sequence in French dairy goats. BMC Genet. 2020, 21, 19. [Google Scholar] [CrossRef]
- Nagai, R.; Kinukawa, M.; Watanabe, T.; Ogino, A.; Kurogi, K.; Adachi, K.; Satoh, M.; Uemoto, Y. Genome-wide detection of non-additive quantitative trait loci for semen production traits in beef and dairy bulls. Animal 2022, 16, 100472. [Google Scholar] [CrossRef]
- Butler, M.L.; Hartman, A.R.; Bormann, J.M.; Weaber, R.L.; Grieger, D.M.; Rolf, M.M. Genome-wide association study of beef bull semen attributes. BMC Genom. 2022, 23, 74. [Google Scholar] [CrossRef] [PubMed]
- Sweett, H.; Fonseca, P.A.S.; Suárez-Vega, A.; Livernois, A.; Miglior, F.; Cánovas, A. Genome-wide association study to identify genomic regions and positional candidate genes associated with male fertility in beef cattle. Sci. Rep. 2020, 10, 20102. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Diaz, J.; Cenadelli, S.; Bornaghi, V.; Bongioni, G.; Montedoro, S.M.; Achilli, A.; Capelli, C.; Rincon, J.C.; Milanesi, M.; Passamonti, M.M.; et al. Identification of genomic regions associated with total and progressive sperm motility in Italian Holstein bulls. J. Dairy Sci. 2023, 106, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Peñagaricano, F. Unravelling the genomic architecture of bull fertility in Holstein cattle. BMC Genet. 2016, 17, 143. [Google Scholar] [CrossRef]
- Hiltpold, M.; Kadri, N.K.; Janett, F.; Witschi, U.; Schmitz-Hsu, F.; Pausch, H. Autosomal recessive loci contribute significantly to quantitative variation of male fertility in a dairy cattle population. BMC Genom. 2021, 22, 225. [Google Scholar] [CrossRef]
- Diniz, D.B.; Lopes, M.S.; Broekhuijse, M.L.W.J.; Lopes, P.S.; Harlizius, B.; Guimarães, S.E.F.; Duijvesteijn, N.; Knol, E.F.; Silva, F.F. A genome-wide association study reveals a novel candidate gene for sperm motility in pigs. Anim. Reprod. Sci. 2014, 151, 201–207. [Google Scholar] [CrossRef]
- Gao, N.; Chen, Y.; Liu, X.; Zhao, Y.; Zhu, L.; Liu, A.; Jiang, W.; Peng, X.; Zhang, C.; Tang, Z.; et al. Weighted single-step GWAS identified candidate genes associated with semen traits in a Duroc boar population. BMC Genom. 2019, 20, 797. [Google Scholar] [CrossRef] [PubMed]
- Gòdia, M.; Reverter, A.; González-Prendes, R.; Ramayo-Caldas, Y.; Castelló, A.; Roíguez-Gil, J.E.; Sánchez, A.; Clop, A. A systems biology framework integrating GWAS and RNA-seq to shed light on the molecular basis of sperm quality in swine. Genet. Sel. Evol. 2020, 52, 72. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.B.D.; Bastiaansen, J.W.M.; Broekhuijse, M.L.W.J.; Lopes, M.S.; Knol, E.F.; Harlizius, B.; Guimarães, S.E.F.; Silva, F.F.; Lopes, P.S. Weighted single-step GWAS and gene network analysis reveal new candidate genes for semen traits in pigs. Genet. Sel. Evol. 2018, 50, 40. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.B.D.; Lopes, M.S.; Broekhuijse, M.L.W.J.; Guimarães, S.E.F.; Kno, E.F.; Bastiaansen, J.W.M.; Silva, F.; Lopes, P.S. Genetic parameters for semen quality and quantity traits in five pig lines. J. Anim. Sci. 2017, 95, 4251–4259. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, Q.; Liao, W.; Zhang, W.; Li, T.; Li, J.; Zhang, Z.; Huang, X.; Zhang, H. Identification of New Candidate Genes Related to Semen Traits in Duroc Pigs through Weighted Single-Step GWAS. Animals 2023, 13, 365. [Google Scholar] [CrossRef]
- Hering, D.M.; Oleński, K.; Ruść, A.; Kaminski, S. Genome-wide association study for semen volume and total number of sperm in Holstein-Friesian bulls. Anim. Reprod. Sci. 2014, 151, 126–130. [Google Scholar] [CrossRef]
- Mańkowska, A.; Brym, P.; Paukszto, Ł.; Jastrzębski, J.P.; Fraser, L. Gene Polymorphisms in Boar Spermatozoa and Their Associations with Post-Thaw Semen Quality. Int. J. Mol. Sci. 2020, 21, 1902. [Google Scholar] [CrossRef]
- Torabi, F.; Bogle, O.A.; Estanyol, J.M.; Oliva, R.; Miller, D. Zona pellucida-binding protein 2 (ZPBP2) and several proteins containing BX7B motifs in human sperm may have hyaluronic acid binding or recognition properties. Mol. Human Reprod. 2017, 23, 803–816. [Google Scholar] [CrossRef]
- Martinez, C.A.; Roca, J.; Alvarez-Rodriguez, M.; Rodriguez-Martinez, H. miRNA-Profiling in Ejaculated and Epididymal Pig Spermatozoa and Their Relation to Fertility after Artificial Insemination. Biology 2022, 11, 236. [Google Scholar] [CrossRef]
- Nogueira, E.; Tirpák, F.; Hamilton, L.E.; Zigo, M.; Kerns, K.; Sutovsky, M.; Kim, J.; Volkmann, D.; Jovine, L.; Taylor, J.F.; et al. A Non-Synonymous Point Mutation in a WD-40 Domain Repeat of EML5 Leads to Decreased Bovine Sperm Quality and Fertility. Front. Cell Dev. Biol. 2022, 10, 872740. [Google Scholar] [CrossRef]
- Kynetec. MLA and AWI Wool and Sheepmeat Survey Report—Sheepmeat; Meat & Livestock Australia (MLA): Sydney, Australia; Australian Wool Innovation (AWI): Sydney, Australia, 2022. [Google Scholar]
- Moghaddar, N.; Gore, K.P.; Daetwyler, H.D.; Hayes, B.J.; van der Werf, J.H.J. Accuracy of genotype imputation based on random and selected reference sets in purebred and crossbred sheep populations and its effect on accuracy of genomic prediction. Genet. Sel. Evol. 2015, 47, 97. [Google Scholar] [CrossRef]
- Browning, B.L.; Yu, Z. Simultaneous Genotype Calling and Haplotype Phasing Improves Genotype Accuracy and Reduces False-Positive Associations for Genome-wide Association Studies. Am. J. Hum. Genet. 2009, 85, 847–861. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.2.; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Rönnegård, L.; McFarlane, S.E.; Husby, A.; Kawakami, T.; Ellegren, H.; Qvarnström, A. Increasing the power of genome wide association studies in natural populations using repeated measures–evaluation and implementation. Methods Ecol. Evol. 2016, 7, 792–799. [Google Scholar] [CrossRef]
- Santana, M.; Kadarmideen, H.; Pant, S.; Alexandre, P.; Junior, G.O.; Gomes, R.; Utsunomiya, H.H.F.; Garcia, J.F.; Fukumasu, H.; Ferraz, J.B.S. Systems genetics investigations for feed intake, feed efficiency and performance in Nellore (Bos indicus) Cattle. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014. [Google Scholar]
- Rangwala, S.H.; Kuznetsov, A.; Ananiev, V.; Asztalos, A.; Borodin, E.; Evgeniev, V.; Joukov, V.; Lotov, V.; Pannu, R.; Rudnev, D.; et al. Accessing NCBI data using the NCBI sequence viewer and genome data viewer (GDV). Genom. Res. 2021, 31, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman: Harlow, UK, 1996. [Google Scholar]
- Schürmann, A.; Axer, H.; Scheepers, A.; Doege, H.; Joost, H.-G. glucose transport facilitator GLUT8 is predominantly associated with the acrosomal region of mature spermatozoa. Cell Tissue Res. 2002, 307, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Gawlik, V.; Schmidt, S.; Scheepers, A.; Wennemuth, G.; Augustin, R.; Aumüller, G.; Moser, M.; Al-Hasani, H.; Kluge, R.; Joost, H.-G.; et al. Targeted disruption of Slc2a8 (GLUT8) reduces motility and mitochondrial potential of spermatozoa. Mol. Membr. Biol. 2008, 25, 224–235. [Google Scholar] [CrossRef]
- Adastra, K.L.; Frolova, A.I.; Chi, M.M.; Cusumano, D.; Bade, M.; Carayannopoulos, M.O.; Moley, K.H. Slc2a8 deficiency in mice results in reproductive and growth impairments. Biol. Reprod. 2012, 87, 49. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Wei, L.L.; Zhang, X.H.; Jin, H.J.; Chen, S.R. Loss of perinuclear theca ACTRT1 causes acrosome detachment and severe male subfertility in mice. Development 2022, 149, dev200489. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, Y.; Song, N.; Fan, Y.; Li, K.; Teng, X.; Guo, Q.; Ding, Z. Proteomic pattern changes associated with obesity-induced asthenozoospermia. Andrology 2015, 3, 247–259. [Google Scholar] [CrossRef]
- Lv, C.; Larbi, A.; Memon, S.; Liang, J.; Zhao, X.; Shao, Q.; Wu, G.; Quan, G. The proteomic characterization of ram sperm during cryopreservation analyzed by the two-dimensional electrophoresis coupled with mass spectrometry. Cryobiology 2020, 97, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hidalgo, D.; Macías-García, B.; García-Marín, L.J.; Bragado, M.J.; González-Fernández, L. Boar spermatozoa proteomic profile varies in sperm collected during the summer and winter. Anim. Reprod. Sci. 2020, 219, 106513. [Google Scholar] [CrossRef] [PubMed]
- Murase, T.; Imaeda, N.; Yamada, H.; Miyazawa, K. Seasonal changes in semen characteristics, composition of seminal plasma and frequency of acrosome reaction induced by calcium and calcium ionophore A23187 in large white boars. J. Reprod. Dev. 2007, 53, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, B.; Wang, X.; Liu, X.; Zhang, Q.; Chen, Y. Estimation of genetic parameters and season effects for semen traits in three pig breeds of South China. J. Anim. Breed. Genet. 2019, 136, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zasiadczyk, L.; Fraser, L.; Kordan, W.; Wasilewska, K. Individual and seasonal variations in the quality of fractionated boar ejaculates. Theriogenology 2015, 83, 1287–1303. [Google Scholar] [CrossRef]
- Talluri, T.R.; Kumaresan, A.; Sinha, M.K.; Paul, N.; Ebenezer Samuel King, J.P.; Datta, T.K. Integrated multi-omics analyses reveals molecules governing sperm metabolism potentially influence bull fertility. Sci. Rep. 2022, 12, 10692. [Google Scholar] [CrossRef]
- Zhao, G.; Zhao, X.; Bai, J.; Dilixiati, A.; Song, Y.; Haire, A.; Zhao, S.; Aihemaiti, A.; Fu, X.; Wusiman, A. Metabolomic and Transcriptomic Changes Underlying the Effects of L-Citrulline Supplementation on Ram Semen Quality. Animals 2023, 13, 217. [Google Scholar] [CrossRef]
- Cornwall, G.A.; von Horsten, H.H. Sperm Maturation in the Epididymis: Role of Segment-Specific Micro Environments; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Słowińska, M.; Paukszto, Ł.; Paweł Jastrzębski, J.; Bukowska, J.; Kozłowski, K.; Jankowski, J.; Ciereszko, A. Transcriptome analysis of turkey (Meleagris gallopavo) reproductive tract revealed key pathways regulating spermatogenesis and post-testicular sperm maturation. Poult. Sci. 2020, 99, 6094–6118. [Google Scholar] [CrossRef]
- Chapman, J.C.; Michael, S.D. Proposed mechanism for sperm chromatin condensation/decondensation in the male rat. Reprod. Biol. Endocrinol. 2003, 1, 20. [Google Scholar] [CrossRef]
- Llavanera, M.; Ribas-Maynou, J.; Delgado-Bermúdez, A.; Recuero, S.; Muiño, R.; Hidalgo, C.O.; Tamargo, C.; Bonet, S.; Mateo-Otero, Y.; Yeste, M. Sperm chromatin condensation as an in vivo fertility biomarker in bulls: A flow cytometry approach. J. Anim. Sci. Biotechnol. 2021, 12, 115. [Google Scholar] [CrossRef]
- Milardi, D.M.D.P.D.; Grande, G.M.D.; Vincenzoni, F.B.D.; Messana, I.B.D.; Pontecorvi, A.M.D.; De Marinis, L.M.D.; Castagnola, M.B.D.; Marana, R.M.D. Proteomic approach in the identification of fertility pattern in seminal plasma of fertile men. Fertil. Steril. 2012, 97, 67–73.e61. [Google Scholar] [CrossRef] [PubMed]
- Ghaoui, R.E.H.; Gillan, L.; Thomson, P.C.; Evans, G.; Maxwell, W.C. Effect of seminal plasma fractions from entire and vasectomized rams on the motility characteristics, membrane status, and in vitro fertility of ram spermatozoa. J. Androl. 2007, 28, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Ghanayem, B.I.; Bai, R.; Kissling, G.E.; Travlos, G.; Hoffler, U. Diet-Induced Obesity in Male Mice Is Associated with Reduced Fertility and Potentiation of Acrylamide-Induced Reproductive Toxicity. Biol. Reprod. 2010, 82, 96–104. [Google Scholar] [CrossRef]
- Freitas, M.J.; Vijayaraghavan, S.; Fardilha, M. Signaling mechanisms in mammalian sperm motility. Biol. Reprod. 2017, 96, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S.; Ho, H.C. Hyperactivated motility in sperm. Reprod. Domest. Anim. 2003, 38, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Schenkel, F.S.; Melo, A.L.P.; Oliveira, H.R.; Pedrosa, V.B.; Araujo, A.C.; Melka, M.G.; Brito, L.F. Identifying pleiotropic variants and candidate genes for fertility and reproduction traits in Holstein cattle via association studies based on imputed whole-genome sequence genotypes. BMC Genom. 2022, 23, 331. [Google Scholar] [CrossRef]
- Yao, X.; Yang, H.; Zhang, Y.; Ren, C.; Nie, H.; Fan, Y.; Zhou, W.; Wang, S.; Feng, X.; Wang, F. Characterization of GALNTL5 gene sequence and expression in ovine testes and sperm. Theriogenology 2017, 95, 54–61. [Google Scholar] [CrossRef]
- Hitit, M.; Özbek, M.; Ayaz-Guner, S.; Guner, H.; Oztug, M.; Bodu, M.; Kirbas, M.; Bulbul, B.; Bucak, M.N.; Ataman, M.B.; et al. Proteomic fertility markers in ram sperm. Anim. Reprod. Sci. 2021, 235, 106882. [Google Scholar] [CrossRef]
- Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.L.; Dacheux, J.L.; Guérin, Y.; Gatti, J.L.; de Graaf, S.P.; Druart, X. Ram seminal plasma proteome and its impact on liquid preservation of spermatozoa. J. Proteom. 2014, 109, 245–260. [Google Scholar] [CrossRef]
- Westfalewicz, B.; Dietrich, M.A.; Mostek, A.; Partyka, A.; Bielas, W.; Niżański, W.; Ciereszko, A. Analysis of bull (Bos taurus) seminal vesicle fluid proteome in relation to seminal plasma proteome. J. Dairy Sci. 2017, 100, 2282–2298. [Google Scholar] [CrossRef]
- Huang, Y.-L.; Fu, Q.; Pan, H.; Chen, F.-M.; Zhao, X.-L.; Wang, H.-J.; Zhang, P.-F.; Huang, F.-L.; Lu, Y.-Q.; Zhang, M. Spermatogenesis-associated proteins at different developmental stages of buffalo testicular seminiferous tubules identified by comparative proteomic analysis. Proteomics 2016, 16, 2005–2018. [Google Scholar] [CrossRef]
- Galan, C.; Serra, R.W.; Sun, F.; Rinaldi, V.D.; Conine, C.C.; Rando, O.J. Stability of the cytosine methylome during post-testicular sperm maturation in mouse. PLoS Genet. 2021, 17, e1009416. [Google Scholar] [CrossRef]
- Cornwall, G.A. New insights into epididymal biology and function. Hum. Reprod. Update 2009, 15, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Hafez, B.H.E.S.E. Reproduction in Farm Animals, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Li, T.; Wang, H.; Luo, R.; Shi, H.; Su, M.; Wu, Y.; Li, Q.; Ma, K.; Zhang, Y.; Ma, Y. Identification and Functional Assignment of Genes Implicated in Sperm Maturation of Tibetan Sheep. Animals 2023, 13, 1553. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, M.G.M.G.; Visconti, P.E.P.E. Molecular changes and signaling events occurring in sperm during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef] [PubMed]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef]
- Balogh Eszter, E. Genome Wide Association Study and Complex Andrological Examination of Hungarian Swine Breeds in Order to Improve Reproductive Parameters. Ph.D. Thesis, University of Debrecen, Debrecen, Hungary, 2020. [Google Scholar]
- Lazaros, L.; Xita, N.; Kaponis, A.; Hatzi, E.; Plachouras, N.; Sofikitis, N.; Zikopoulos, K.; Georgiou, I. The association of aromatase (CYP19) gene variants with sperm concentration and motility. Asian J. Androl. 2011, 13, 292–297. [Google Scholar] [CrossRef]
- Carreau, S.; de Vienne, C.; Galeraud-Denis, I. Aromatase and estrogens in man reproduction: A review and latest advances. Adv. Med. Sci. 2008, 53, 139–144. [Google Scholar] [CrossRef]
- Simpson, E.R.; Clyne, C.; Rubin, G.; Boon, W.C.; Robertson, K.; Britt, K.; Speed, C.; Jones, M. Aromatase—A brief overview. Annu. Rev. Physiol. 2002, 64, 93–127. [Google Scholar] [CrossRef]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Dodson, M.M.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar]
- Carreau, S.; Silandre, D.; Bourguiba, S.; Hamden, K.; Said, L.; Lambard, S.; Galeraud-Denis, I.; Delalande, C. Estrogens and male reproduction: A new concept. Braz. J. Med. Biol. Res. 2007, 40, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Aquila, S.; Sisci, D.; Gentile, M.; Middea, E.; Siciliano, L.; Andò, S. Human Ejaculated Spermatozoa Contain Active P450 Aromatase. J. Clin. Endocrinol. Metab. 2002, 87, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Lambard, S.; Galeraud-Denis, I.; Saunders, P.T.; Carreau, S. Human immature germ cells and ejaculated spermatozoa contain aromatase and oestrogen receptors. J. Mol. Endocrinol. 2004, 32, 279–289. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Mulligan, B.P.; Kim, H.-M.; Yang, B.-C.; Lee, C.-K. Quantitative analysis of sperm mRNA in the pig: Relationship with early embryo development and capacitation. Reprod. Fertil. Dev. 2013, 25, 807–817. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.; Jiang, Y.; Li, J.; Chen, M.; Wang, R.; Zhao, J.; Peng, Z.; Huang, H.; Liu, R. Whole Exome Sequencing Identifies Genes Associated with Non-Obstructive Azoospermia. Front. Genet. 2022, 13, 872179. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Enatsu, N.; Fujisawa, M. Management of non-obstructive azoospermia. Reprod. Med. Biol. 2016, 15, 165–173. [Google Scholar] [CrossRef]
- Cochran, S.D.; Cole, J.B.; Null, D.J.; Hansen, P.J. Discovery of single nucleotide polymorphisms in candidate genes associated with fertility and production traits in Holstein cattle. BMC Genet. 2013, 14, 49. [Google Scholar] [CrossRef] [PubMed]
- Cochran, S.D.; Cole, J.B.; Null, D.J.; Hansen, P.J. Single Nucleotide Polymorphisms in Candidate Genes Associated with Fertilizing Ability of Sperm and Subsequent Embryonic Development in Cattle. Biol. Reprod. 2013, 89, 69. [Google Scholar] [CrossRef]
- Singh, R.; Junghare, V.; Hazra, S.; Singh, U.; Sengar, G.S.; Raja, T.V.; Kumar, S.; Tyagi, S.; Das, A.K.; Kumar, A.; et al. Database on spermatozoa transcriptogram of catagorised Frieswal crossbred (Holstein Friesian X Sahiwal) bulls. Theriogenology 2019, 129, 130–145. [Google Scholar] [CrossRef]
- Garde, J.; Roldan, E.R.S. rab3-Peptide stimulates exocytosis of the ram sperm acrosome via interaction with cyclic AMP and phospholipase A2 metabolites. FEBS Lett. 1996, 391, 263–268. [Google Scholar] [CrossRef]
- Breitbart, H.; Shabtay, O. Sperm Acrosome Reaction. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Academic Press: Oxford, MS, USA, 2018; pp. 284–288. [Google Scholar]
- Wang, X.; Yang, C.; Guo, F.; Zhang, Y.; Ju, Z.; Jiang, Q.; Zhao, X.; Liu, Y.; Zhao, H.; Wang, J.; et al. Integrated analysis of mRNAs and long noncoding RNAs in the semen from Holstein bulls with high and low sperm motility. Sci. Rep. 2019, 9, 2092. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.K.; Gupta, N.; Sankhwar, S.N.; Rajender, S. Differential Genes Expression between Fertile and Infertile Spermatozoa Revealed by Transcriptome Analysis. PLoS ONE 2015, 10, e0127007. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Aghajanian, H.K.; Haig-Ladewig, L.A.; Gerton, G.L. Sorbitol Can Fuel Mouse Sperm Motility and Protein Tyrosine Phosphorylation via Sorbitol Dehydrogenase. Biol. Reprod. 2009, 80, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Starmer, J.; Martin, E.R. A multiple testing correction method for genetic association studies using correlated single nucleotide polymorphisms. Genet. Epidemiol. 2008, 32, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yin, H.; Zhang, X.; Sun, D.; Zhang, Q.; Liu, J.; Ding, X.; Zhang, Y.; Zhang, S. Genome-wide association study for semen traits of the bulls in Chinese Holstein. Anim. Genet. 2016, 48, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Fortes, M.R.S.; Reverter, A.; Kelly, M.; McCulloch, R.; Lehnert, S.A. Genome-wide association study for inhibin, luteinizing hormone, insulin-like growth factor 1, testicular size and semen traits in bovine species. Andrology 2013, 1, 644–650. [Google Scholar] [CrossRef]
- Liu, S.; Yin, H.; Li, C.; Qin, C.; Cai, W.; Cao, M.; Zhang, S. Genetic effects of PDGFRB and MARCH1 identified in GWAS revealing strong associations with semen production traits in Chinese Holstein bulls. BMC Genet. 2017, 18, 63. [Google Scholar] [CrossRef]
- Mei, Q.; Fu, C.; Sahana, G.; Chen, Y.; Yin, L.; Miao, Y.; Zhao, S.; Xiang, T. Identification of new semen trait-related candidate genes in Duroc boars through genome-wide association and weighted gene co-expression network analyses. J. Anim. Sci. 2021, 99, skab188. [Google Scholar] [CrossRef]
- Gredler, B.; Fuerst, C.; Fuerst-Waltl, B.; Schwarzenbacher, H.; Sölkner, J. Genetic Parameters for Semen Production Traits in Austrian Dual-Purpose Simmental Bulls. Reprod. Domest. Anim. 2007, 42, 326–328. [Google Scholar] [CrossRef]
- Berry, D.P.; Eivers, B.; Dunne, G.; McParland, S. Genetics of bull semen characteristics in a multi-breed cattle population. Theriogenology 2019, 123, 202–208. [Google Scholar] [CrossRef]
- Furstoss, V.; David, I.; Leboeuf, B.; Guillouet, P.; Boué, P.; Bodin, L. Genetic and non-genetic parameters of several characteristics of production and semen quality in young bucks. Anim. Reprod. Sci. 2009, 110, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J. Heritabilities and genetic correlations for litter size and semen traits in Czech Large White and Landrace pigs. J. Anim. Sci. 2010, 88, 2893–2903. [Google Scholar] [CrossRef] [PubMed]
- Smital, J.; Wolf, J.; Sousa, L.L.D. Estimation of genetic parameters of semen characteristics and reproductive traits in AI boars. Anim. Reprod. Sci. 2005, 86, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.; Eustache, F.; Ducot, B.; Blandin, T.; Daudin, M.; Diaz, I.; Matribi, S.E.; Gony, B.; Keskes, L.; Kolbezen, M.; et al. Intra- and inter-individual variability in human sperm concentration, motility and vitality assessment during a workshop involving ten laboratories. Hum. Reprod. 2000, 15, 2360–2368. [Google Scholar] [CrossRef]


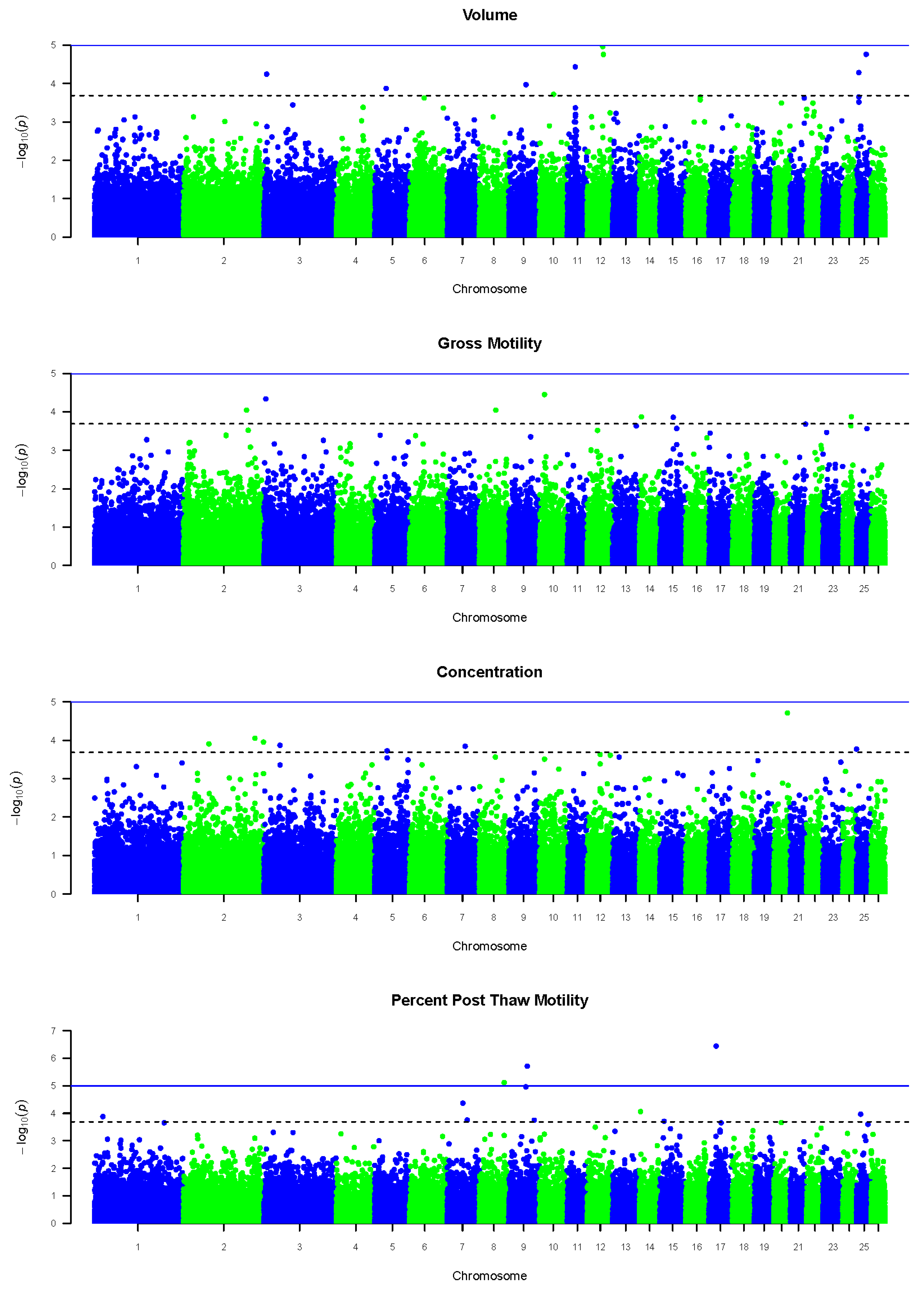{kind=link}
| Volume | Gross Motility | Concentration | Percent Post-Thaw Motility | |
|---|---|---|---|---|
| Number of Rams | 246 | 246 | 245 | 242 |
| Number of Records | 4506 | 4486 | 3758 | 3100 |
| Mean (±Standard Deviation) | 1.24 (±0.48) | 4.35 (±1.15) | 4319.75 (±859.41) | 55.25 (±15.34) |
| Trait | OAR | Location (Mb) | Start Window (Mb) | End Window (Mb) | p-Value | |
|---|---|---|---|---|---|---|
| Volume | 3 | 8.91 | 3.14 | 8.41 | 9.41 | 5.70 × 105 |
| 5 | 35.60 | 6.90 | 35.10 | 36.10 | 1.34 × 104 | |
| 9 | 53.65 | 0.72 | 53.15 | 54.15 | 1.11 × 104 | |
| 10 | 44.48 | 0.07 | 43.98 | 44.98 | 1.92 × 104 | |
| 11 | 25.38 | 4.61 | 24.88 | 25.88 | 3.68 × 105 | |
| 12 | 48.41 | 0.47 | 47.91 | 48.91 | 1.09 × 105 | |
| 12 | 50.58 | 0.65 | 50.08 | 51.08 | 1.75 × 105 | |
| 25 | 7.62 | 0.48 | 7.12 | 8.12 | 5.19 × 105 | |
| 25 | 30.55 | 0.04 | 30.05 | 31.05 | 1.74 × 105 | |
| Gross Motility | 2 | 195.43 | 0.41 | 194.93 | 195.93 | 9.07 × 105 |
| 3 | 6.18 | 0.21 | 5.68 | 6.68 | 4.64 × 105 | |
| 8 | 50.82 | 2.30 | 50.32 | 51.32 | 9.03 × 105 | |
| 10 | 16.62 | 2.03 × 103 | 16.12 | 17.12 | 3.56 × 105 | |
| 14 | 6.22 | 1.67 | 5.72 | 6.72 | 1.36 × 104 | |
| 15 | 42.21 | 1.79 | 41.71 | 42.71 | 1.39 × 104 | |
| 24 | 26.44 | 1.21 | 25.94 | 26.94 | 1.35 × 104 | |
| Concentration | 2 | 78.88 | 0.98 | 78.38 | 79.38 | 1.25 × 104 |
| 2 | 221.22 | 0.16 | 220.72 | 221.72 | 8.82 × 105 | |
| 2 | 247.87 | 0.02 | 247.37 | 248.37 | 1.11 × 104 | |
| 3 | 49.95 | 0.41 | 49.45 | 50.45 | 1.35 × 104 | |
| 5 | 38.41 | 0.02 | 37.91 | 38.91 | 1.88 × 104 | |
| 7 | 55.95 | 1.29 | 55.45 | 56.45 | 1.43 × 104 | |
| 20 | 42.51 | 1.70 | 42.01 | 43.01 | 1.94 × 105 | |
| 25 | 0.74 | 0.09 | 0.24 | 1.24 | 1.69 × 104 | |
| Percent Post-Thaw Motility | 1 | 25.78 | 0.22 | 25.28 | 26.28 | 1.32 × 104 |
| 7 | 49.04 | 0.84 | 48.54 | 49.54 | 4.31 × 105 | |
| 7 | 62.09 | 1.67 × 104 | 61.59 | 62.59 | 1.76 × 104 | |
| 8 | 76.70 | 0.39 | 76.20 | 77.20 | 7.72 × 106 | |
| 9 | 53.42 | 8.36 | 52.92 | 53.92 | 1.10 × 105 | |
| 9 | 57.73 | 4.80 × 103 | 57.23 | 58.23 | 1.95 × 106 | |
| 9 | 79.08 | 0.16 | 78.58 | 79.58 | 1.79 × 104 | |
| 14 | 3.61 | 13.70 | 3.12 | 4.12 | 8.71 × 105 | |
| 15 | 13.90 | 0.48 | 13.40 | 14.40 | 1.96 × 104 | |
| 17 | 22.67 | 1.47 | 22.17 | 23.17 | 3.62 × 107 | |
| 25 | 13.35 | 2.47 | 12.85 | 13.85 | 1.09 × 104 |
| Trait | QTL Region (OAR:Mb) | Candidate Gene Symbol |
|---|---|---|
| Volume | 3:8.41–9.41 | TTC16 A, PTRH1, C3H9orf117, STXBP1 A, FAM129B A, LRSAM1 A, RPL12 A, ZNF79, SLC2A8 A, GARNL3, RALGPS1 A, ANGPTL2 A, ZBTB34 A, ZBTB43 A, and LMX1B A |
| 5:35.1–36.1 | COMMD10, ARL10, NOP16 A, HIGD2A A, CLTB A, FAF2, RNF44 A, CDHR2 A, GPRIN1, SNCB, EIF4E1B, TSPAN17, UNC5A, HK3 A, UIMC1, ZNF346, FGFR4 A, and NSD1 A | |
| 9:53.15–54.15 | - | |
| 10:43.98–44.98 | KLHL1 A | |
| 11:24.88–25.88 | KIAA0753, PITPNM3, TRNAK-UUU A, FAM64A, AIPL1, WSCD1, TRNAW-CCA A, NLRP1 A, TRNAC-GCA A, MIS12 A, DERL2 A, DHX33, C1QBP A, RPAIN A, and NUP88 A, RABEP1 A | |
| 12:47.91–48.91 | PRDM16 A, ACTRT2 A, TRNAC-GCA A, TTC34, MMEL1 A, FAM213B, HES5 A, PANK4, PLCH2 A, PEX10 A, RER1, MORN1, SKI, C12H1orf86, PRKCZ A, GABRD A, CFAP74, TMEM52 A, and CALML6 | |
| 12:50.08–51.08 | CASP9 A, EFHD2 A, FHAD1, and TMEM51 A | |
| 25:7.12–8.12 | SLC35F3 A, TARBP1 A, IRF2BP2 A, TRNAR-CCU, TOMM20 A, RBM34 A, ARID4B A, and GGPS1 A | |
| 25:30.05–31.05 | ADK, KAT6B A, DUPD1, DUSP13 A, SAMD8 A, VDAC2 A, COMTD1, ZNF503 A, and C25H10orf11 | |
| Gross Motility | 2:194.93–195.93 | - |
| 3:5.68–6.68 | FIBCD1 A, QRFP A, ABL1 A, EXOSC2 A, PRDM12, FUBP3, ASS1 A, HMCN2, NCS1, GPR107, FNBP1, USP20, C3H9orf78, TOR1A, and TOR1B | |
| 8:50.32–51.32 | SYNCRIP, SNX14, and NT5E A | |
| 10:16.12–17.12 | SPERT A, SIAH3, ZC3H13 A, CPB2, LCP1 A, LRRC63, KIAA0226L, LRCH1, ESD, and TRNAG-CCC A | |
| 14:5.72–6.72 | MAF | |
| 15:41.71–42.71 | TRNAW-CCA A, RNF141 A, AMPD3, ADM, SBF2, SWAP70 A, WEE1 A, and ZNF143 | |
| 24:25.94–26.94 | ATP2A1, SH2B1 A, TUFM A, ATXN2L A, CLN3, APOBR A, IL27 A, NUPR1 A, CCDC101 A, SLX1A, BOLA2B, CORO1A, MAPK3 A, GDPD3, YPEL3 A, TBX6, PPP4C A, ALDOA A, FAM57B, C24H16orf92, TRNAG-CCC A, DOC2A, INO80E, HIRIP3, TAOK2 A, TMEM219, KCTD13 A, ASPHD1, SEZ6L2, CDIPT A, MVP, PAGR1, PRRT2, MAZ, KIF22 A, ZG16 A, C24H16orf54, QPRT, SPN, CD2BP2, TBC1D10B A, MYLPF, ZNF48 A, ZNF771 A, DCTPP1, SEPHS2 A, ITGAL A, ZNF768, ZNF688, ZNF689 A, PRR14, FBRS A, SRCAP A, TMEM265, TRNAR-UCG, TRNAR-UCG, PHKG2 A, C24H16orf93, RNF40 A, and ZNF629 | |
| Concentration | 2:78.38–79.38 | - |
| 2:220.72–221.72 | - | |
| 2:247.37–248.37 | ACTL8 A, ARHGEF10L A, RCC2 A, PADI6 A, PADI3 A, PADI1 A, and PADI2 A | |
| 3:49.45–50.45 | - | |
| 5:37.91–38.91 | TRIM52, ZNF496 A, NLRP3 A, GCSAML A, TRIM58 | |
| 7:55.45–56.45 | TMOD3 A, TMOD2, LYSMD2, SCG3 A, DMXL2 A, GLDN, CYP19 A, TNFAIP8L3 A, and AP4E1 A | |
| 20:42.01–43.01 | SIRT5, GFOD1, TBC1D7, and PHACTR1 A | |
| 25:0.24–1.24 | RAB4A A, SPHAR, CCSAP A, ACTA1 A, NUP133 A, ABCB10 A, TAF5L A, and URB2 A | |
| Percent Post-Thaw Motility | 1:25.28–26.28 | C1H1orf185, RNF11 A, TRNAY-GUA, TTC39A, EPS15 A, OSBPL9 A, NRD1 A, RAB3B A, TXNDC12 A, KTI12, and BTF3L4 A |
| 7:48.54–49.54 | ADAM10, LIPC, AQP9 A, and ALDH1A2 A | |
| 7:61.59–62.59 | SQRDL A, BLOC1S6 A, SLC30A4 A, C7H15orf48, SPATA5L1, GATM A, SLC28A2 A, TRNAH-GUG, SHF A, DUOX1 A, DUOXA1 A, DUOXA2 A, DUOX2 A, SORD A, and FERMT2 A | |
| 8:76.2–77.2 | SYNE1 A, MYCT1 A, VIP A, FBXO5 A, MTRF1L A, and RGS17 A | |
| 9:52.92–53.92 | PEX2 A | |
| 9:57.23–58.23 | FABP5 A, PMP2 A, FABP9 A, FABP4 A, FABP12 A, NOV, MAL2, COLEC10, and TNFRSF11B A | |
| 9:78.58–79.58 | NIPAL2, POP1, HRSP12 A, ERICH5, RPL30 A, MATN2 A, LAPTM4B A, MTDH A, and TSPYL5 A | |
| 14:3.12–4.12 | MON1B A, SYCE1L A, and ADAMTS18 A | |
| 15:13.4–14.4 | MAML2, MTMR2 A, CEP57 A, and FAM76B A | |
| 17:22.17–23.17 | - | |
| 25:12.85–13.85 | ZNF25 A, ZNF33B A, ZNF248, BMS1, TRNAE-UUC, RET, CSGALNACT2 A, RASGEF1A A, FXYD4 A, HNRNPF A, and BICC1 |
| Trait | r (± SE) | |||||
|---|---|---|---|---|---|---|
| Volume | 0.005 | 0.006 | 0.010 | 0.027 | 0.104 | 0.436 (±0.170) |
| Gross Motility | 0.011 | 0.013 | 0.011 | 0.044 | 0.139 | 0.443 (±0.161) |
| Concentration | 3.610 | 4.230 | 10.981 | 31.692 | 0.071 | 0.373 (±0.186) |
| Percent Post-Thaw Motility | 0.106 | 0.109 | 0.224 | 0.707 | 0.092 | 0.383 (±0.182) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodge, M.J.; de las Heras-Saldana, S.; Rindfleish, S.J.; Stephen, C.P.; Pant, S.D. QTLs and Candidate Genes Associated with Semen Traits in Merino Sheep. Animals 2023, 13, 2286. https://doi.org/10.3390/ani13142286
Hodge MJ, de las Heras-Saldana S, Rindfleish SJ, Stephen CP, Pant SD. QTLs and Candidate Genes Associated with Semen Traits in Merino Sheep. Animals. 2023; 13(14):2286. https://doi.org/10.3390/ani13142286
Chicago/Turabian StyleHodge, Marnie J., Sara de las Heras-Saldana, Sally J. Rindfleish, Cyril P. Stephen, and Sameer D. Pant. 2023. "QTLs and Candidate Genes Associated with Semen Traits in Merino Sheep" Animals 13, no. 14: 2286. https://doi.org/10.3390/ani13142286
APA StyleHodge, M. J., de las Heras-Saldana, S., Rindfleish, S. J., Stephen, C. P., & Pant, S. D. (2023). QTLs and Candidate Genes Associated with Semen Traits in Merino Sheep. Animals, 13(14), 2286. https://doi.org/10.3390/ani13142286