
Molecular Systematics of the Firefly Genus Luciola (Coleoptera: Lampyridae: Luciolinae) with the Description of a New Species from Singapore

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Field Sampling
2.2. Morphological Analysis
2.3. DNA Extraction, Sequencing and Bioinformatics
2.4. Genetic Data and Phylogenetic Analyses
3. Results
3.1. Morphology
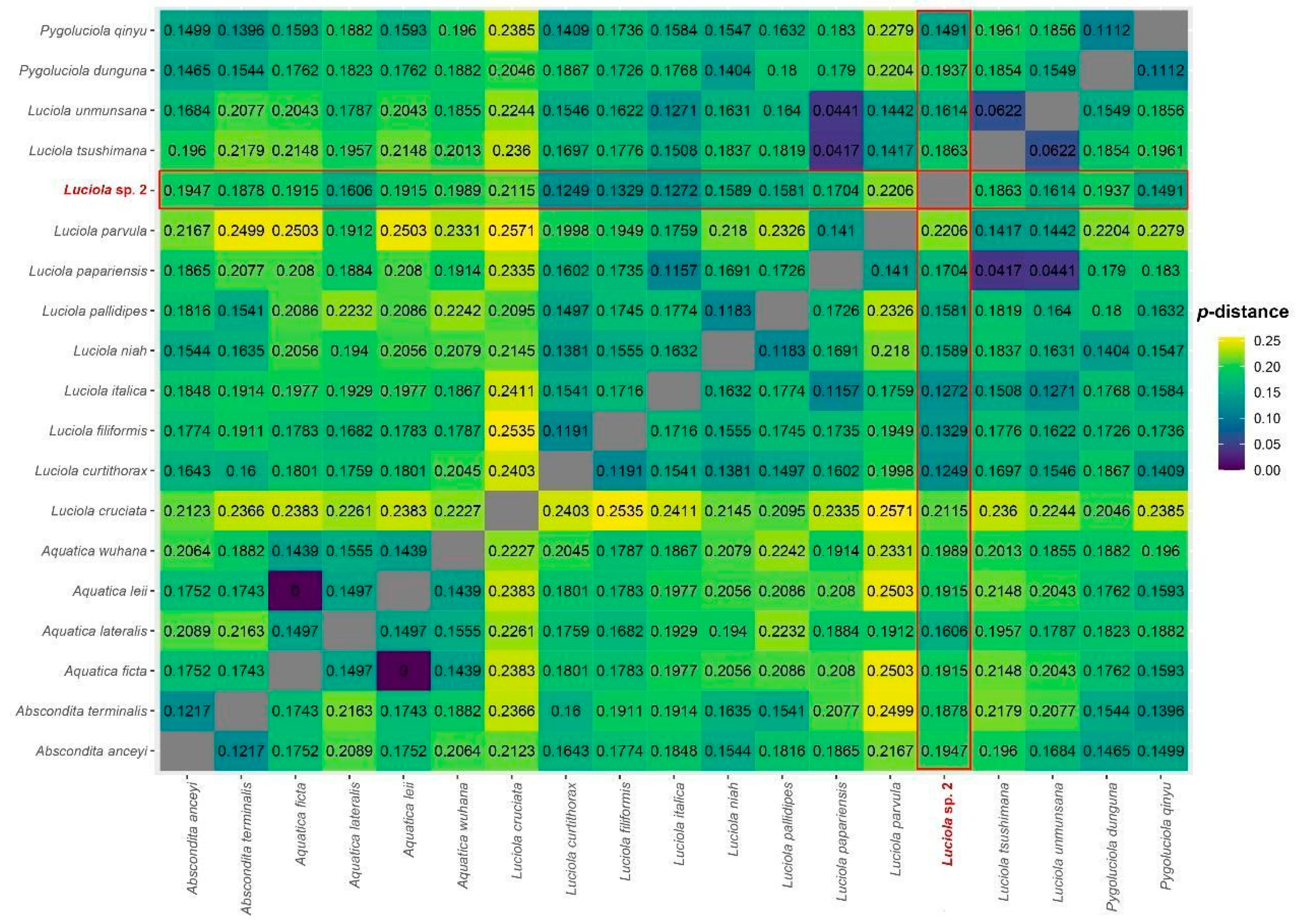
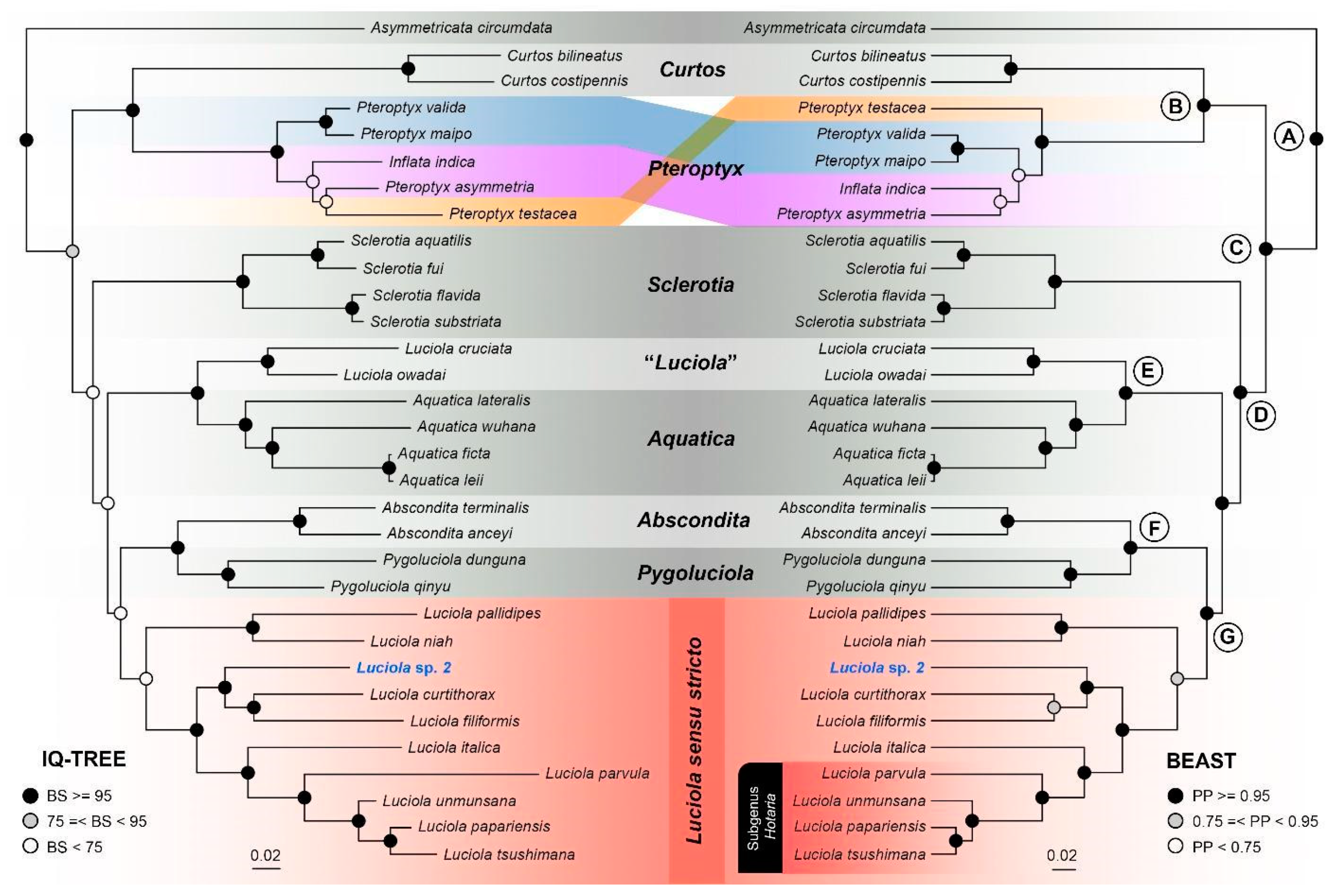
3.2. Genetic Distance, Phylogenetic Analyses, and Divergence Times
3.3. Systematics
- Luciola singapura Jusoh and Ballantyne sp. nov.
- LSID: urn:lsid:zoobank.org:act:A963B320-8889-4FEC-B152-21E3EFAB76F9
- English name: Singapore firefly
- Malay name: Kunang-kunang Singapura
3.3.1. Type Material
3.3.2. Diagnosis
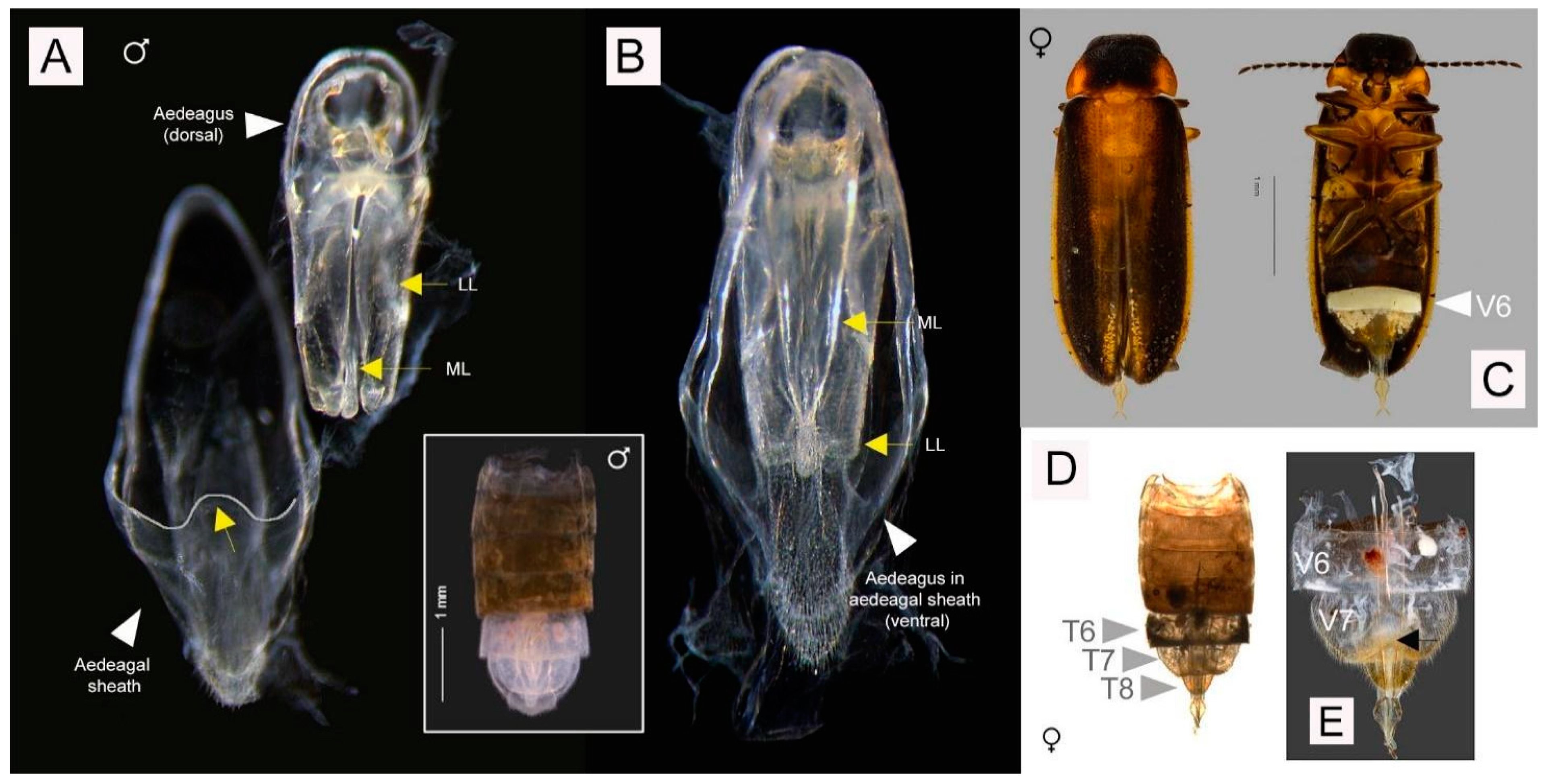
3.3.3. Description
3.3.4. Etymology
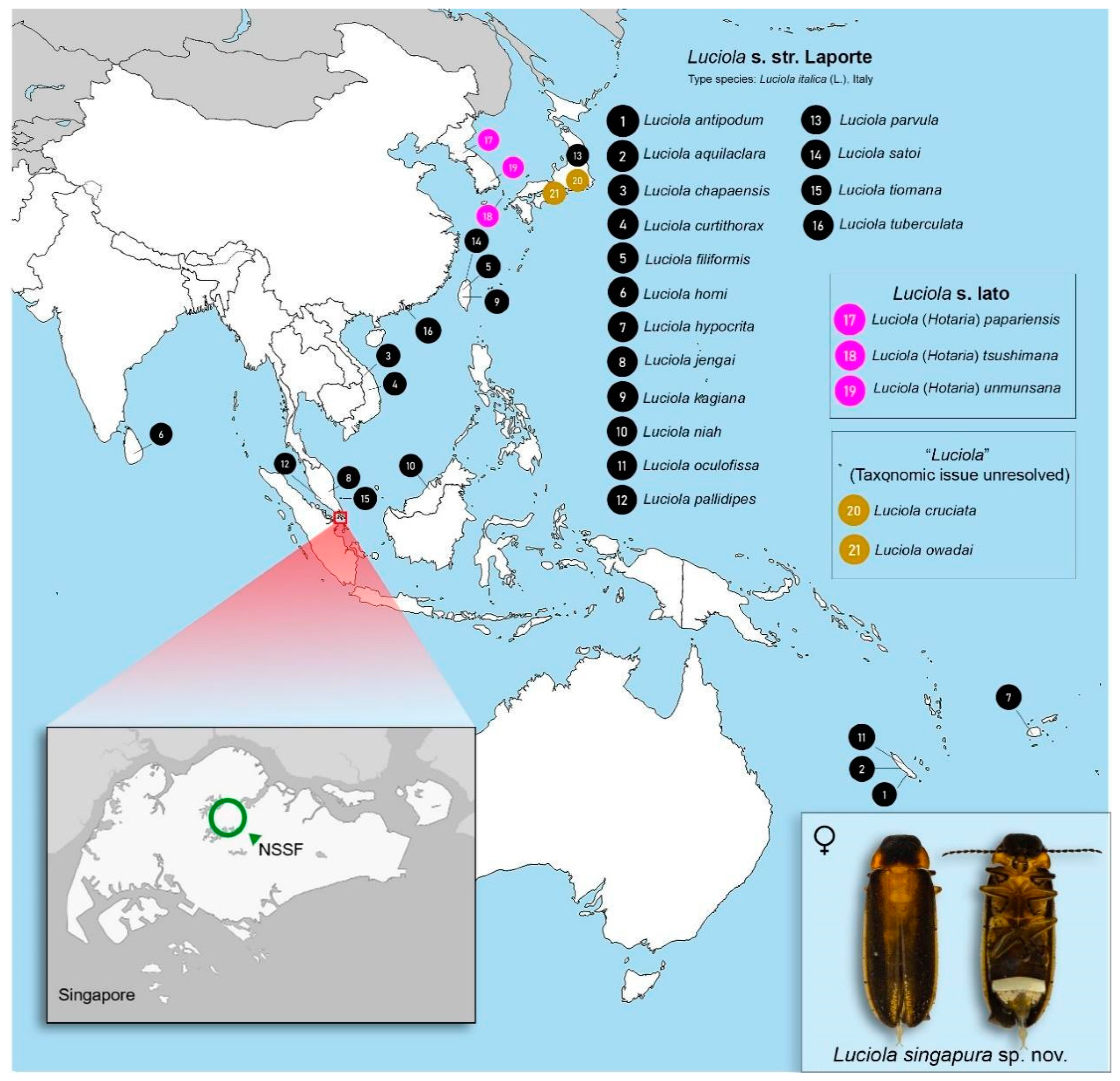
3.3.5. Distribution
3.3.6. Ecology
4. Discussion
4.1. Phylogenetic Relationships and Evolutionary History
4.2. Updated Notes on the Species of Luciola s. str. Laporte throughout the SE Asian and Australopacific Region
- antipodum (Bourgeois, 1884) [46]
- aquilaclara Ballantyne, 2013 in [8]
- chapaensis Pic, 1923 [47]
- curtithorax Pic, 1927 [48]
- filiformis Olivier, 1913 [49]
- horni Bourgeois, 1905 [50]
- hypocrita Olivier, 1888 [51]
- jengai Nada, 2019 in [3] (provisional)
- kagiana Matsumura, 1928 [52]
- niah Jusoh, 2019 in [3]
- oculofissa Ballantyne, 2013 in [8]
- pallidipes Pic, 1928 [53]
- papariensis Doi, 1932 [54] (provisional)
- parvula Kiesenwetter, 1874 [55]
- satoi Jeng and Yang, 2003 in [43]
- singapura Jusoh and Ballantyne sp. nov.
- tiomana Ballantyne, 2019 in [3]
- tsushimana Nakane, 1970 [56] (provisional)
- tuberculata Yiu, 2017 [44]
- unmunsana Doi, 1931 [57] (provisional)
4.3. Status and Conservation of Fireflies in Singapore
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballantyne, L.; Lambkin, C.L.; Boontop, Y.; Jusoh, W.F.A. Revisional studies on the Luciolinae fireflies of Asia (Coleoptera: Lampyridae): 1. The genus Pyrophanes Olivier with two new species. 2. Four new species of Pteroptyx Olivier and 3. A new genus Inflata Boontop, with redescription of Luciola indica (Motsch.) as Inflata indica comb. nov. Zootaxa 2015, 3959, 1–84. [Google Scholar] [CrossRef]
- Ballantyne, L.A.; Lambkin, C.L.; Luan, X.; Boontop, Y.; Nak-Eiam, S.; Pimpasalee, S.; Silalom, S.; Thancharoen, A. Further studies on South Eastern Asian Luciolinae: 1. Sclerotia Ballantyne, a new genus of fireflies with back swimming larvae 2. Triangulara Pimpasalee, a new genus from Thailand (Coleoptera: Lampyridae). Zootaxa 2016, 4170, 201–249. [Google Scholar] [CrossRef]
- Ballantyne, L.A.; Lambkin, C.L.; Ho, J.Z.; Jusoh, W.F.A.; Nada, B.; Nak-Eiam, S.; Thancharoen, A.; Wattanachaiyingcharoen, W.; Yiu, V. The Luciolinae of S. E. Asia and the Australopacific region: A revisionary checklist (Coleoptera: Lampyridae) including description of three new genera and 13 new species. Zootaxa 2019, 4687, 1–174. [Google Scholar] [CrossRef] [PubMed]
- McDermott, F.A. Lampyridae. In Coleopterorum Catalogus Supplementa. Pars 9. Editio Secunda; Steel, W.O., Ed.; W. Junk: S’Gravenhage, The Netherlands, 1966; p. 149. [Google Scholar]
- Ballantyne, L.; Fu, X.; Lambkin, C.; Jeng, M.L.; Faust, L.; Wijekoon, W.M.C.D.; Li, D.; Zhu, T. Studies on South-east Asian fireflies: Abscondita, a new genus with details of life history, flashing patterns and behaviour of Abs. chinensis (L.) and Abs. terminalis (Olivier) (Coleoptera: Lampyridae: Luciolinae. Zootaxa 2013, 3721, 1–48. [Google Scholar] [CrossRef]
- Olivier, E. Coleoptera. Fam. Lampyridae. In Genera Insectorum. Fasc. 53; Wytsman, P., Ed.; Verteneuil & Desmet: Bruxelles, Belgium, 1907; pp. 1–74. [Google Scholar]
- Olivier, E. Pars 9. Lampyridae. In Coleopterorum Catalogus; Schenkling, S., Ed.; Junk, W.: Berlin, Germany, 1910; p. 68. [Google Scholar]
- Ballantyne, L.A.; Lambkin, C.L. Systematics and phylogenetics of Indo-Pacific Luciolinae fireflies (Coleoptera: Lampyridae) and the description of new genera. Zootaxa 2013, 3653, 1–162. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, L.A.; Menayah, R. Redescription of the synchronous firefly, Pteroptyx tener Olivier (Coleoptera: Lampyridae), of Kampung Kuantan, Selangor. Malayan Nat. J. 2000, 54, 323–328. [Google Scholar]
- Ballantyne, L.A.; Lambkin, C. Lampyridae of Australia (Coleoptera: Lampyridae: Luciolinae: Luciolini). Mem. Queensl. Museum 2000, 46, 15–93. [Google Scholar]
- Fu, X.; Ballantyne, L.; Lambkin, C. The external larval morphology of aquatic and terrestrial Luciolinae fireflies (Coleoptera: Lampyridae). Zootaxa 2012, 3405, 1–34. [Google Scholar] [CrossRef]
- Ballantyne, L.A.; Lambkin, C. A phylogenetic reassessment of the rare S. E. Asian firefly genus Pygoluciola Wittmer (Coleoptera: Lampyridae: Luciolinae). Raffles Bull. Zool. 2006, 54, 21–48. [Google Scholar]
- Ballantyne, L.A.; Lambkin, C.L. Systematics of Indo-Pacific fireflies with a redefinition of Australasian Atyphella Olliff, Madagascan Photuroluciola Pic, and description of seven new genera from the Luciolinae (Coleoptera: Lampyridae). Zootaxa 2009, 1997, 1–188. [Google Scholar] [CrossRef]
- Fu, X.H.; Ballantyne, L.A.; Lambkin, C. Aquatica gen. nov. from mainland China with a description of Aquatica wuhana sp. nov. (Coleoptera: Lampyridae: Luciolinae). Zootaxa 2010, 2530, 1–18. [Google Scholar] [CrossRef]
- Jusoh, W.F.A.; Ballantyne, L.; Lambkin, C.L.; Hashim, N.R.; Wahlberg, N. The firefly genus Pteroptyx Olivier revisited (Coleoptera: Lampyridae: Luciolinae). Zootaxa 2018, 4456, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis. Editio Duodecima, Reformata. Tomus I. Pars II; Laur. Salvii: Holmiae, Stockholm, Sweden, 1767. [Google Scholar]
- Ballantyne, L.A. Pygoluciola satoi, a new species of the rare S E Asian firefly genus Pygoluciola Wittmer from the Philippines (Coleoptera: Lampyridae: Luciolinae). Raffles Bull. Zool. 2008, 56, 1–9. [Google Scholar]
- Ballantyne, L.A.; Lambkin, C. A new firefly, Luciola (Pygoluciola) Kinabalua, new species (Coleoptera: Lampyridae), from Malaysia, with observations on a possible copulation clamp. Raffles Bull. Zool. 2001, 49, 363–377. [Google Scholar]
- Fu, X.H.; Ballantyne, L. Taxonomy and behaviour of lucioline fireflies (Coleoptera: Lampyridae: Luciolinae) with redefinition and new species of Pygoluciola Wittmer from mainland China and review of Luciola LaPorte. Zootaxa 2008, 1733, 1–44. [Google Scholar] [CrossRef]
- Choi, Y.S.; Bae, J.S.; Lee, K.S.; Kim, S.R.; Kim, I.; Kim, J.G.; Kim, K.Y.; Kim, S.E.; Suzuki, H.; Lee, S.M.; et al. Genomic structure of the luciferase gene and phylogenetic analysis in the Hotaria-group fireflies. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2003, 134, 199–214. [Google Scholar] [CrossRef]
- Ohba, N. Flash patterns and communication of the two ecotypes in Hotaria parvula. Sci. Rep. Yokosuka City Mus. 2000, 47, 1–22. [Google Scholar]
- Han, T.; Kim, S.H.; Yoon, H.J.; Park, I.G.; Park, H. Evolutionary history of species of the firefly subgenus Hotaria (Coleoptera, Lampyridae, Luciolinae, Luciola) inferred from DNA barcoding data. Contrib. Zool. 2020, 89, 127–145. [Google Scholar] [CrossRef]
- Kawashima, I.; Suzuki, H.; Satô, M. A check-list of Japanese fireflies (Coleoptera, Lampyridae and Rhagophthalmidae). Jpn. J. Syst. Entomol. 2003, 11, 225–262. [Google Scholar]
- Chan, S.H.; Ballantyne, L.A.; Goh, C.A.L. Survey of species diversity, distribution and abundance of fireflies in Singapore. Lampyrid 2012, 2, 113–122. [Google Scholar]
- Jusoh, W.F.A. Coleoptera: A very special firefly and other beetle stories. In Voyageurs, Explorateurs et Scientifiques: The French and Natural History in Singapore; Low, M.E.Y., Pocklington, K., Jusoh, W.F.A., Eds.; Lee Kong Chian Natural History Museum: Singapore, 2019; pp. 212–232. [Google Scholar]
- Ng, P.K.L.; Lim, K.K.P. The conservation status of the Nee Soon freshwater swamp forest of Singapore. Aquat. Conserv. Mar. Freshw. Ecosyst. 1992, 2, 255–266. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012. [Google Scholar] [CrossRef]
- Boisvert, S.; Raymond, F.; Godzaridis, É.; Laviolette, F.; Corbeil, J. Ray Meta: Scalable de novo metagenome assembly and profiling. Genome Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013. [Google Scholar] [CrossRef]
- Borowiec, M.L. AMAS: A fast tool for alignment manipulation and computing of summary statistics. PeerJ 2016, 4, e1660. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S.; Le, S.V. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In 2010 Proceedings of the Gateway Computing Environments Workshop, GCE 2010; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Bouckaert, R.R.; Drummond, A.J. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6. Available online: http://beast.bio.ed.ac.uk/Tracer2014 (accessed on 20 September 2020).
- Jeng, M.-L.; Yang, P.-S.; Lai, J. Notes on the genus Luciola (Coleoptera, Lampyridae, Luciolinae) of Taiwan. Spec. Bull. Jpn. Soc. Coleopterol. 2003, 6, 247–262. [Google Scholar]
- Yiu, V. A study of Rhagophthalmidae and Lampyridae in Hong Kong with descriptions of new species (Coleoptera): Part 2. Lampyrid 2017, 4, 67–119. [Google Scholar]
- Jusoh, W.F.A.; Ballantyne, L.; Chan, K.O. DNA-based species delimitation reveals cryptic and incipient species in synchronous flashing fireflies (Coleoptera: Lampyridae) of Southeast Asia. Biol. J. Linn. Soc. 2020. [Google Scholar] [CrossRef]
- Bourgeois, J. Dascillides & Malacodermes de Nouvelle-Calédonie. Rev. d’entomologie 1884, 3, 278–290. [Google Scholar]
- Pic, M. Deuxième addenda. Faune Entomol. l’Indo-Chine Française 1923, 6, 58–63. [Google Scholar]
- Pic, M. Malacodermes exotiques. L’Échange Rev. Linnéenne 1927, XLIII, 49. [Google Scholar]
- Olivier, E.H. Sauter’s Formosa-Ausbeute: Lampyridae (Col.). Entomol. Mittellungen 1913, II, 271. [Google Scholar]
- Bourgeois, J. Voyage du D.W. Horn à Ceylan. Malacodermes et Lymexylonides. II. Lampyridini. Ann. Société Entomol. Fr. 1905, 74, 127. [Google Scholar]
- Olivier, E. Etudes sur les Lampyrides IV. Descriptions d’espèces nouvelles. Ann. Société Entomol. Fr. 1888, 6, 47–62. [Google Scholar]
- Matsumura, M. The fireflies. In Using Interesting Insects as Teaching Tool; Tokyodô Shoten: Tokyo, Japan, 1928; pp. 39–70. (In Japanese) [Google Scholar]
- Pic, M. Malacodermes exotiques. L’Échange, Rev. Linnéenne 1928, XLIV, 58–63. [Google Scholar]
- Doi, H. A new species of Luciola from Korea. Luciola papariensis sp. nov. J. Chosen Nat. Hist. Soc. 1932, 14, 64. [Google Scholar]
- Kiesenwetter, H. Die Malacodermen Japans nach den Ergebnisse der Sammlungen des Herrn L. Lewis während der Jahre 1869–1871. Dtsch. Entomol. Z. 1874, 18, 241–288. [Google Scholar]
- Nakane, T. On the Cantharoid Coleoptera found in the islands of Tsushima (Insecta). Mem. Natl. Sci. Mus. 1970, 3, 287–288. [Google Scholar]
- Doi, H. A new species of Luciola from Korea. Luciola unmunsana sp. nov. J. Chosen Nat. Hist. Soc. 1931, 12, 54–55. [Google Scholar]
- Tan, J.C.H. Fireflies of Pulau Ubin, Singapore. Nat. Singap. 2018, 11, 127–138. [Google Scholar]
- Davison, G.W.H.; Ng, P.K.L.; Chew, H.H. The Singapore Red Data Book: Threatened Plants and Animals of Singapore, 2nd ed.; Nature Society (Singapore): Singapore, 2008; ISBN 9789810802004. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Definition |
|---|---|
| a | aedeagal dimension, distance from dorsal base of the lateral lobes to the apex of the median lobe (see b); expressed as b/a [15] (p. 25, Figure 25; p.37, Figure 104) |
| A | pronotal dimension measured from above; width across anterior third |
| ASD | distance between antennal sockets |
| ASW | antennal socket greatest diameter |
| b | aedeagal dimension, distance from the dorsal base of the lateral lobes to the apex of the lateral lobes (see a); expressed as b/a [15] (p. 25, Figure 25; p.37, Figure 104) |
| B | pronotal dimension measured from above; width across middle |
| BL | body length measured as median length of pronotum plus length of elytron |
| C | pronotal dimension measured from above; width across posterior third |
| FS | antennal flagellar segments |
| GHW | greatest head width (across eyes, measured parallel to ASD) |
| L | length |
| Legs 1, 2 etc | legs and parts of legs are referred to by their segment number e.g., legs 1 are prothoracic legs; tarsi 2 = mesothoracic tarsi; femora 3 = metathoracic femora. |
| LL | lateral lobes, aedeagus |
| LO | light organ |
| ML | median lobe of the aedeagus |
| MN | mesonotal plates |
| MS | mesoscutellum |
| MPP | median posterior projection ventrite 7 male only |
| PLP | posterolateral projections ventrite 7 male only |
| SIW | smallest interocular width (measured horizontally and may be on the same level as ASD, ASW, or above it if the eyes are closer there) |
| T7, 8 etc. | abdominal tergites |
| V6, 7 etc. | abdominal ventrites, referred to by actual, not visible number |
| W | width |
| W/L | width/length |
| x | times |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jusoh, W.F.A.; Ballantyne, L.; Chan, S.H.; Wong, T.W.; Yeo, D.; Nada, B.; Chan, K.O. Molecular Systematics of the Firefly Genus Luciola (Coleoptera: Lampyridae: Luciolinae) with the Description of a New Species from Singapore. Animals 2021, 11, 687. https://doi.org/10.3390/ani11030687
Jusoh WFA, Ballantyne L, Chan SH, Wong TW, Yeo D, Nada B, Chan KO. Molecular Systematics of the Firefly Genus Luciola (Coleoptera: Lampyridae: Luciolinae) with the Description of a New Species from Singapore. Animals. 2021; 11(3):687. https://doi.org/10.3390/ani11030687
Chicago/Turabian StyleJusoh, Wan F. A., Lesley Ballantyne, Su Hooi Chan, Tuan Wah Wong, Darren Yeo, B. Nada, and Kin Onn Chan. 2021. "Molecular Systematics of the Firefly Genus Luciola (Coleoptera: Lampyridae: Luciolinae) with the Description of a New Species from Singapore" Animals 11, no. 3: 687. https://doi.org/10.3390/ani11030687
APA StyleJusoh, W. F. A., Ballantyne, L., Chan, S. H., Wong, T. W., Yeo, D., Nada, B., & Chan, K. O. (2021). Molecular Systematics of the Firefly Genus Luciola (Coleoptera: Lampyridae: Luciolinae) with the Description of a New Species from Singapore. Animals, 11(3), 687. https://doi.org/10.3390/ani11030687