Replacing Forage by Crude Olive Cake in a Dairy Sheep Diet: Effects on Ruminal Fermentation and Microbial Populations in Rusitec Fermenters

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Feeding
2.2. Experimental Diets
2.3. Rusitec Trial
2.4. DNA Isolation and Molecular Biology Analyses
2.5. Analytical Procedures
2.6. Calculations and Statistical Analyses
3. Results and Discussion
3.1. Effect of Crude Olive Cake (COC) Inclusion on Diet Disappearance and Rumen Fermentation Parameters
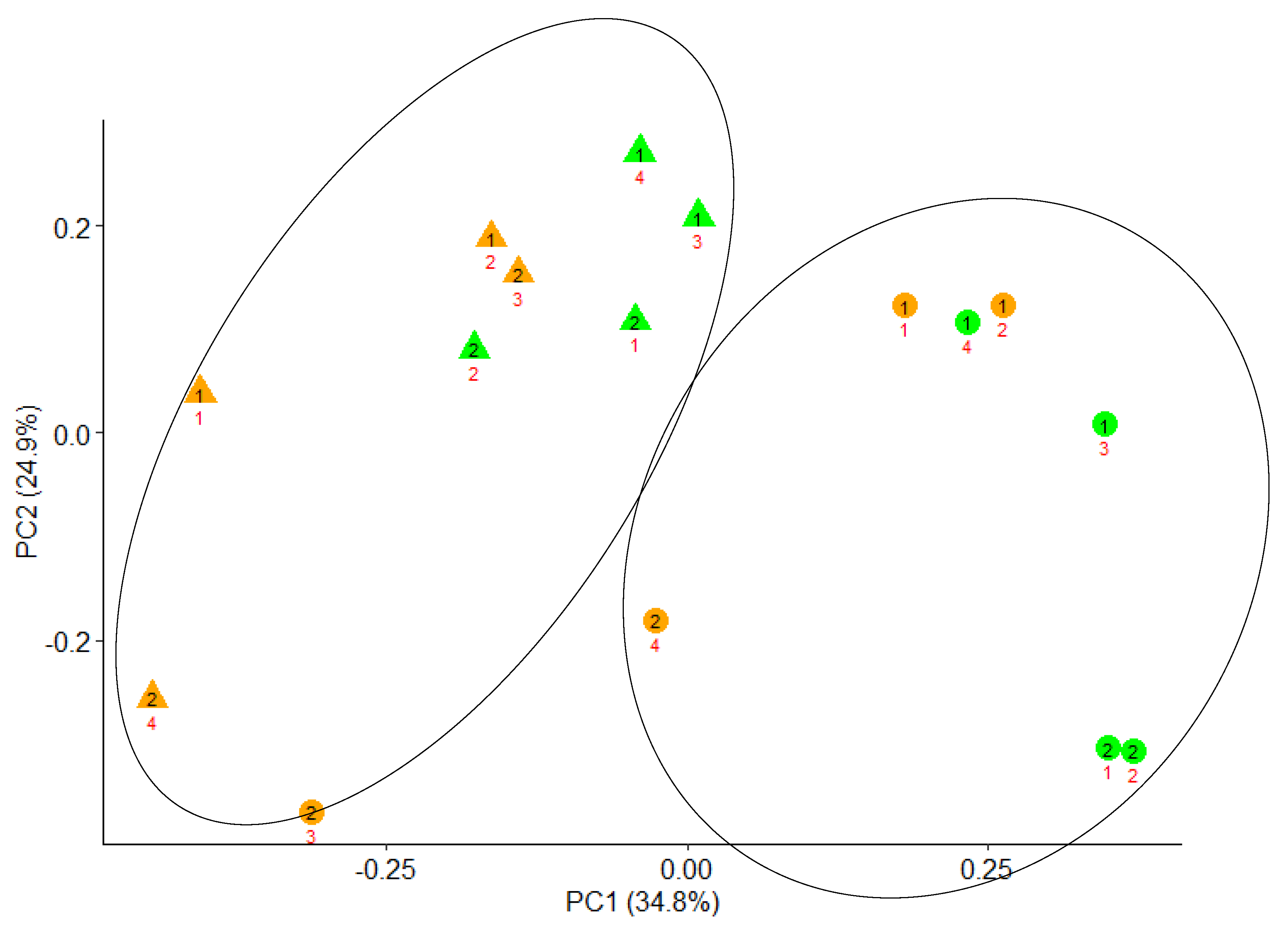
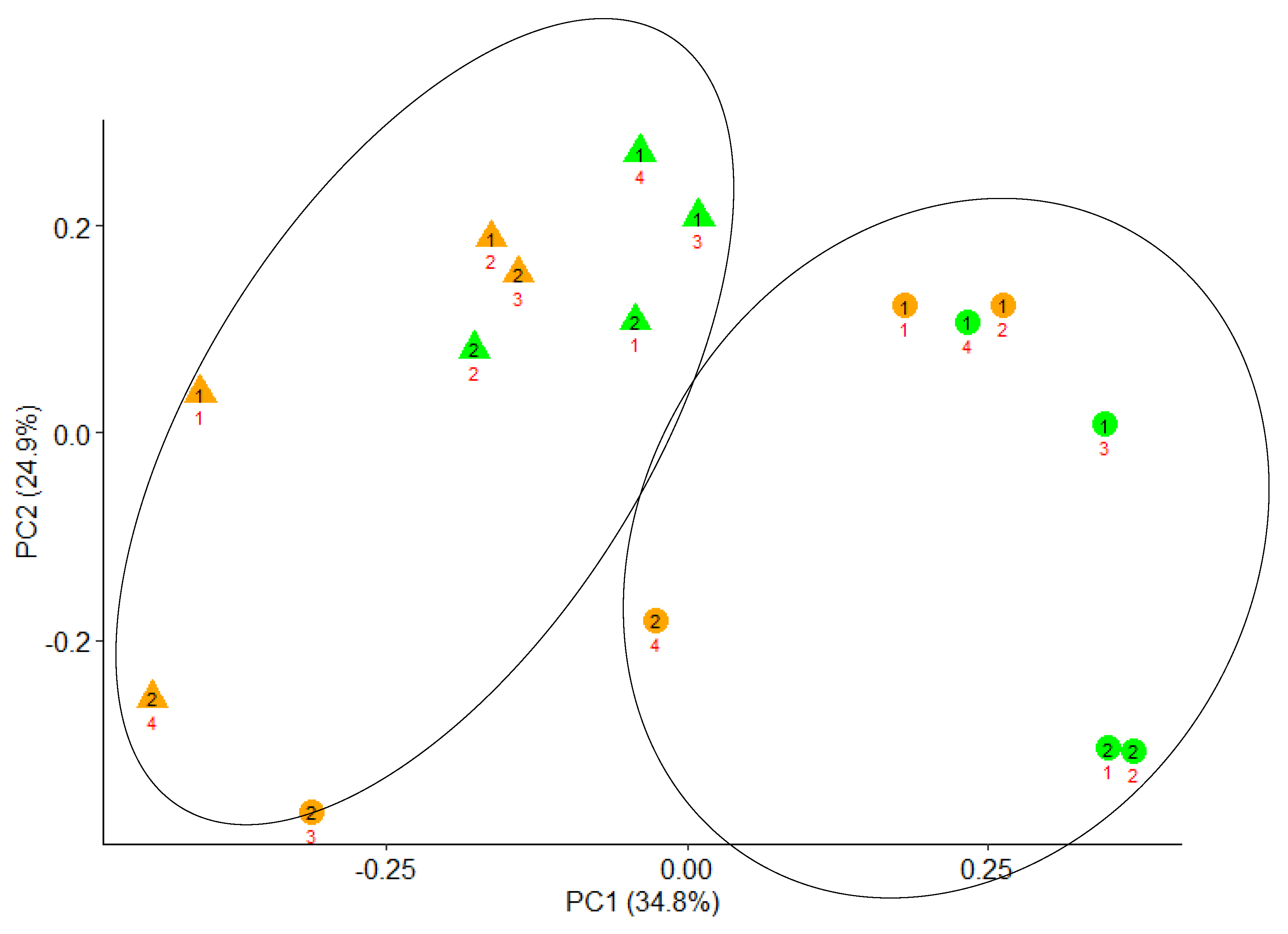
3.2. Effects of Crude Olive Cake (COC) Inclusion on Microbial Protein Synthesis (MPS), Bacterial Diversity, and Microbial Populations
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ministerio de Agricultura, Pesca y Alimentación de España (MAPA). Available online: https://www.mapa.gob.es/estadistica/pags/anuario/2019-Avance/avance/AvAE19.pdf (accessed on 19 May 2020).
- Dermeche, S.; Nadour, M.; Larroche, C.; Moulti-Mati, F.; Michaud, P. Olive mill wastes: Biochemical characterizations and valorization strategies. Process Biochem. 2013, 48, 1532–1552. [Google Scholar] [CrossRef]
- Khosravifar, O.; Maheri-Sis, N.; Aghdam-Shahriar, H.; Salamat-Doost, R.; Baradaran-Hasanzadeh, A.R. Study of the effect of Saccharomyces cerevisiae on nutritional value of exhausted dry olive cake using in vitro gas production technique. J. Anim. Vet. Adv. 2008, 7, 1589–1593. [Google Scholar]
- Amici, A.; Verna, M.; Martillotti, F. Olive by-products in animal feeding: Improvement and utilization. Options Méditerranéennes 1991, 16, 149–152. [Google Scholar]
- FAO. Available online: http://www.fao.org/3/X6545E/X6545E02.htm (accessed on 17 May 2020).
- Molina-Alcaide, E.; Yáñez-Ruiz, D.R. Potential use of olive by-products in ruminant feeding: A review. Anim. Feed Sci. Technol. 2008, 147, 247–264. [Google Scholar] [CrossRef]
- Marcos, C.N.; de Evan, T.; García-Rebollar, P.; de Blas, C.; Carro, M.D. Influence of storage time and processing on chemical composition and in vitro ruminal fermentation of olive cake. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Simitzis, P.E.; Deligeorgis, S.G. Agroindustrial By-Products and Animal Products: A Great Alternative for Improving Food-Quality Characteristics and Preserving Human Health. In Food Quality: Balancing Health and Disease. Volume 13 of Handbook of Food Bioengineering, 1st ed.; Grumezescu, A., Butu, A., Eds.; Academic Press: London, UK, 2018; pp. 253–290. [Google Scholar]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, ruminal fermentation, ciliate protozoal populations, and milk production from dairy cows fed cinnamaldehyde, quebracho condensed tannin, or Yucca schidigera saponin extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef] [PubMed]
- Marcos, C.N.; García-Rebollar, P.; de Blas, C.; Carro, M.D. Variability in the chemical composition and in vitro ruminal fermentation of olive cake by-products. Animals 2019, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Chiofalo, B.; Liotta, L.; Zumbo, A.; Chiofalo, V. Administration of olive cake for ewe feeding: Effect on milk yield and composition. Small Rumin. Res. 2004, 55, 169–176. [Google Scholar] [CrossRef]
- Estaún, J.; Dosil, J.; Al Alami, A.; Gimeno, A.; de Vega, A. Effects of including olive cake in the diet on performance and rumen function of beef cattle. Anim. Prod. Sci. 2014, 54, 1817–1821. [Google Scholar] [CrossRef]
- Farghaly, M.S.; Ebrahem, E.A.E.; Mahmoud, A.E.M. Performance of barki lambs fed on rations containing olive cake with or without polyethylene glycol. Pak. J. Biol. Sci. 2018, 21, 307–313. [Google Scholar]
- Obeidat, B.S. The effects of feeding olive cake and Saccharomyces cerevisiae supplementation on performance, nutrient digestibility and blood metabolites of Awassi lambs. Anim. Feed Sci. Technol. 2017, 231, 131–137. [Google Scholar] [CrossRef]
- Shdaifat, M.M.; Al-Barakah, F.S.; Kanan, A.Q.; Obeidat, B.S. The effect of feeding agricultural by-products on performance of lactating Awassi ewes. Small Rumin. Res. 2013, 113, 11–14. [Google Scholar] [CrossRef]
- Awawdeh, M.S.; Obeidat, B.S. Treated olive cake as a non-forage fiber source for growing awassi lambs: Effects on nutrient intake, rumen and urine pH, performance, and carcass yield. Asian Australas. J. Anim. Sci. 2013, 26, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Owaimer, A.N.; Kraidees, M.S.; Al-Saiady, M.; Zahran, S.; Abouheif, M.A. Effect of feeding olive cake in complete diet on performance and nutrient utilization of lambs. Asian Australas. J. Anim. Sci. 2004, 17, 491–496. [Google Scholar] [CrossRef]
- Abbeddou, S.; Riwahi, S.; Iñiguez, L.; Zaklouta, M.; Hess, H.D.; Kreuzer, M. Ruminal degradability, digestibility, energy content, and influence on nitrogen turnover of various Mediterranean by-products in fat-tailed Awassi sheep. Anim. Feed Sci. Technol. 2011, 163, 99–110. [Google Scholar] [CrossRef]
- Molina-Alcaide, E.; Moumen, A.; Martín-García, I.; Carro, M.D. Comparison of bacterial pellets and microbial markers for the estimation of the microbial nitrogen and amino acids flows from single flow continuous culture fermenters fed diets containing two-stage olive cake. J. Anim. Physiol. Anim. Nutr. 2009, 93, 527–537. [Google Scholar] [CrossRef]
- Pallara, G.; Buccioni, A.; Pastorelli, R.; Minieri, S.; Mele, M.; Rapaccini, S.; Messini, A.; Pauselli, M.; Servili, M.; Giovannetti, L.; et al. Effect of stoned olive pomace on rumen microbial communities and polyunsaturated fatty acid biohydrogenation: An in vitro study. BMC Vet. Res. 2014, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Mannelli, F.; Cappucci, A.; Pini, F.; Pastorelli, R.; Decorosi, F.; Giovannetti, L.; Mele, M.; Minieri, S.; Conte, G.; Pauselli, M.; et al. Effect of different types of olive oil pomace dietary supplementation on the rumen microbial community profile in Comisana ewes. Sci. Rep. 2018, 8, 8455. [Google Scholar] [CrossRef]
- Martínez, M.E.; Ranilla, M.J.; Tejido, M.L.; Ramos, S.; Carro, M.D. Comparison of fermentation of diets of variable composition and microbial populations in the rumen of sheep and Rusitec fermenters. I. Digestibility, fermentation parameters, and microbial growth. J. Dairy Sci. 2010, 93, 3684–3698. [Google Scholar] [CrossRef]
- McDougall, E.I. Studies on ruminant saliva. 1. The composition and output of sheep’s saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Ranilla, M.J.; Carro, M.D.; Valdés, C.; Giráldez, F.J.; López, S. A comparative study of ruminal activity in Churra and Merino sheep offered alfalfa hay. Anim. Sci. 1997, 65, 121–128. [Google Scholar] [CrossRef]
- Ranilla, M.J.; López, S.; Giráldez, F.J.; Valdés, C.; Carro, M.D. Comparative digestibility and digesta flow kinetics in two breeds of sheep. Anim. Sci. 1998, 66, 389–396. [Google Scholar] [CrossRef]
- Carro, M.D.; Miller, E.L. Effect of supplementing a fibre basal diet with different nitrogen forms on ruminal fermentation and microbial growth in an in vitro semi-continuous culture system (RUSITEC). Br. J. Nutr. 1999, 82, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Saro, C.; Ranilla, M.J.; Carro, M.D. Postprandial changes of fiber-degrading microbes in the rumen of sheep fed diets varying in type of forage as monitored by real-time PCR and automated ribosomal intergenic spacer analysis. J. Anim. Sci. 2012, 90, 4487–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.R.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Saro, C.; Ranilla, M.J.; Cifuentes, A.; Rosselló-Mora, R.; Carro, M.D. Technical note: Comparison of automated ribosomal intergenic spacer analysis (ARISA) and denaturing gradient gel electrophoresis (DGGE) to assess bacterial diversity in the rumen of sheep. J. Anim. Sci. 2014, 92, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Luna, G.M.; Dell’Anno, A.; Pietrangeli, B. Comparison of two fingerprinting techniques, terminal restriction fragment length polymorphism and automated ribosomal intergenic spacer analysis, for determination of bacterial diversity in aquatic environments. Appl. Environ. Microbiol. 2006, 72, 5982–5989. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-5; The R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; AOAC International: Gaithersburg, MD, USA, 1999. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Martínez, M.E.; Ranilla, M.J.; Ramos, S.; Tejido, M.L.; Carro, M.D. Effects of dilution rate and retention time of concentrate on efficiency of microbial growth, methane production, and ruminal fermentation in Rusitec fermenters. J. Dairy Sci. 2009, 92, 3930–3938. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, D.I. Quantitative aspects of microbial metabolism in the rumen and hindgut. In Rumen Microbial Metabolism and Ruminant Digestion; Jouany, J.P., Ed.; INRA Editions: Paris, France, 1991; pp. 217–237. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Satter, L.D.; Slyter, L.L. Effect of ammonia concentration on rumen microbial protein production in vitro. Br. J. Nutr. 1974, 32, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, P.; Durmic, Z.; Vadhanabhuti, J.; Vercoe, P.E. Products derived from olive leaves and fruits can alter in vitro ruminal fermentation and methane production. J. Sci. Food Agric. 2016, 97, 1367–1372. [Google Scholar] [CrossRef]
- Kumar, S.; Choudhury, P.K.; Carro, M.D.; Griffith, G.W.; Dagar, S.S.; Puniya, M.; Calabro, S.; Ravella, S.R.; Dhewa, T.; Upadhyay, R.C.; et al. New aspects and strategies for methane mitigation from ruminants. Appl. Microbiol. Biotechnol. 2014, 98, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Rúiz, D.R.; Molina-Alcaide, E. A comparative study of the effect of two-stage olive cake added to alfalfa on digestion and nitrogen losses in sheep and goats. Animal 2007, 1, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Storm, E.; Brown, D.S.; Øskov, E.R. The nutritive value of rumen micro-organisms in ruminants. 3. The digestion of microbial amino and nucleic acids in, and losses of endogenous nitrogen from, the small intestine of sheep. Br. J. Nutr. 1983, 50, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Rúiz, D.R.; Moumen, A.; Martín-García, A.I.; Molina-Alcaide, E. Ruminal fermentation and degradation patterns, protozoa population, and urinary purine derivates excretion in goats and wethers fed diets based on two-stage olive cake: Effect of PEG supply. J. Anim. Sci. 2004, 82, 2023–2032. [Google Scholar] [CrossRef]
- Dehority, B.A. Rumen Microbiology; Nottingham University Press: Nottingham, UK, 2003. [Google Scholar]
- Suárez, M.; Romero, M.P.; Motilva, M.J. Development of a phenol-enriched olive oil with phenolic compounds from olive cake. J. Agric. Food Chem. 2010, 58, 10396–10403. [Google Scholar] [CrossRef]
- Nunes, M.A.; Pimentel, F.B.; Costa, A.S.G.; Alves, R.C.; Oliveira, M.B.P.P. Olive by-products for functional and food applications: Challenging opportunities to face environmental constraints. Innov. Food Sci. Emerg. Technol. 2016, 35, 139–148. [Google Scholar] [CrossRef]
- Patra, A.K.; Min, B.-R.; Saxena, J. Dietary tannins on microbial ecology of the gastrointestinal tract in ruminants. In Dietary Phytochemicals and Microbes; Patra, A.K., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 237–262. [Google Scholar]
- Cappucci, A.; Alves, S.P.; Bessa, R.J.B.; Buccioni, A.; Mannelli, F.; Pauselli, M.; Viti, C.; Pastorelli, R.; Roscini, V.; Serra, A.; et al. Effect of increasing amounts of olive crude phenolic concentrate in the diet of dairy ewes on rumen liquor and milk fatty acid composition. J. Dairy Sci. 2018, 101, 4992–5005. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Item | Diet | |
|---|---|---|
| CON | COC | |
| Ingredients (g/kg DM) | ||
| Alfalfa hay | 200 | 200 |
| Maize silage | 200 | 67 |
| Barley straw | 100 | 67 |
| Olive cake | - | 166 |
| Soybean meal | 147 | 147 |
| Extruded maize | 134 | 134 |
| Barley | 200 | 200 |
| Mineral/vitamin premix 1 | 10 | 10 |
| Calcium soap of fatty acids | 9 | 9 |
| Chemical composition (g/kg DM) | ||
| Organic matter | 943 | 929 |
| Crude protein | 161 | 160 |
| Neutral detergent fiber 2 | 324 | 322 |
| Acid detergent fiber 2 | 163 | 155 |
| Ether extract | 28.2 | 45.0 |
| Item | Diet | SEM | p-Value | |
|---|---|---|---|---|
| CON | COC | |||
| Fermentation parameters | ||||
| pH | 6.07 | 6.22 | 0.088 | 0.02 |
| Ammonia-N (mg/d) | 123 | 142 | 9.7 | 0.24 |
| Total volatile fatty acids (mmol/d) | 94.1 | 91.5 | 2.71 | 0.19 |
| Molar proportions (mol/100 mol) | ||||
| Acetate | 47.8 | 46.9 | 0.50 | 0.01 |
| Propionate | 21.3 | 21.3 | 0.91 | 0.99 |
| Butyrate | 18.0 | 19.4 | 0.53 | 0.001 |
| Isobutyrate | 0.63 | 0.52 | 0.204 | 0.46 |
| Isovalerate | 1.93 | 2.48 | 0.124 | <0.001 |
| Valerate | 5.79 | 5.78 | 0.167 | 0.89 |
| Caproate | 4.5 | 3.73 | 0.185 | <0.001 |
| Acetate/propionate (mol/mol) | 2.35 | 2.22 | 0.103 | 0.22 |
| Methane (mmol/d) | 23.3 | 22.0 | 0.99 | 0.08 |
| Methane/Total VFA (mol/mol) | 0.247 | 0.249 | 0.0060 | 0.67 |
| Diet disappearance (g/g) | ||||
| Dry matter | 0.709 | 0.748 | 0.0085 | <0.001 |
| Organic matter | 0.699 | 0.737 | 0.0087 | <0.001 |
| Neutral Detergent Fiber | 0.259 | 0.403 | 0.0133 | <0.001 |
| Acid Detergent Fiber | 0.182 | 0.282 | 0.0160 | <0.001 |
| Item | Diet | SEM | p-Value | |
|---|---|---|---|---|
| CON | COC | |||
| Microbial protein synthesis (mg N/day) | ||||
| Solid phase | 199 | 171 | 7.2 | 0.05 |
| Liquid phase | 107 | 119 | 2.6 | 0.03 |
| Total | 306 | 290 | 6.3 | 0.15 |
| Efficiency of microbial growth 1 | 32.9 | 32.6 | 0.81 | 0.79 |
| Digesta Phase | Item 1 | Diet | SEM | p-Value | |
|---|---|---|---|---|---|
| CON | COC | ||||
| Solid | Total bacteria 2 | 101 | 193 | 12.2 | 0.01 |
| Total protozoa 2 | 0.0015 | 0.0067 | 0.00106 | 0.06 | |
| Fungi 3 | 46.7 | 36.6 | 19.33 | 0.80 | |
| Archaea 2 | 0.03 | 0.05 | 0.006 | 0.36 | |
| Number of peaks | 21.0 | 22.5 | 0.85 | 0.56 | |
| Shannon index | 3.04 | 3.09 | 0.029 | 0.66 | |
| Liquid | Total bacteria 2 | 1.75 | 2.01 | 0.246 | 0.72 |
| Total protozoa 2 | 0.0002 | 0.0002 | 0.00002 | 0.99 | |
| Fungi 3 | 0.006 | 0.015 | 0.0022 | 0.25 | |
| Archaea 2 | 0.02 | 0.39 | 0.156 | 0.17 | |
| Number of peaks | 29.2 | 30.2 | 1.06 | 0.81 | |
| Shannon index | 3.37 | 3.39 | 0.093 | 0.85 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, J.; Mateos, I.; Saro, C.; González, J.S.; Carro, M.D.; Ranilla, M.J. Replacing Forage by Crude Olive Cake in a Dairy Sheep Diet: Effects on Ruminal Fermentation and Microbial Populations in Rusitec Fermenters. Animals 2020, 10, 2235. https://doi.org/10.3390/ani10122235
García-Rodríguez J, Mateos I, Saro C, González JS, Carro MD, Ranilla MJ. Replacing Forage by Crude Olive Cake in a Dairy Sheep Diet: Effects on Ruminal Fermentation and Microbial Populations in Rusitec Fermenters. Animals. 2020; 10(12):2235. https://doi.org/10.3390/ani10122235
Chicago/Turabian StyleGarcía-Rodríguez, Jairo, Iván Mateos, Cristina Saro, Jesús S. González, María Dolores Carro, and María José Ranilla. 2020. "Replacing Forage by Crude Olive Cake in a Dairy Sheep Diet: Effects on Ruminal Fermentation and Microbial Populations in Rusitec Fermenters" Animals 10, no. 12: 2235. https://doi.org/10.3390/ani10122235
APA StyleGarcía-Rodríguez, J., Mateos, I., Saro, C., González, J. S., Carro, M. D., & Ranilla, M. J. (2020). Replacing Forage by Crude Olive Cake in a Dairy Sheep Diet: Effects on Ruminal Fermentation and Microbial Populations in Rusitec Fermenters. Animals, 10(12), 2235. https://doi.org/10.3390/ani10122235