
Lectin-Mediated Binding of Engineered Lactococcus lactis to Cancer Cells

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culturing
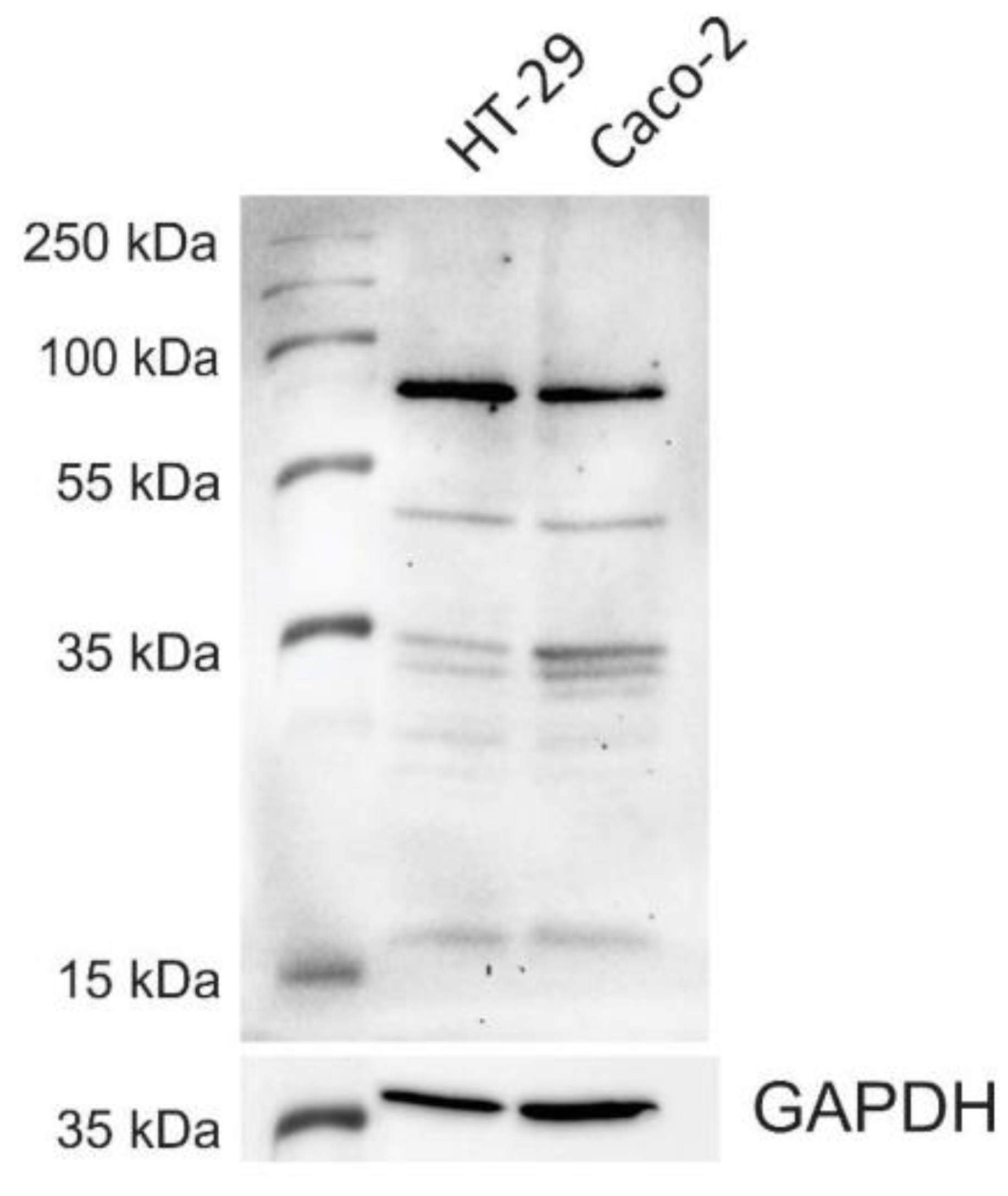
2.2. SDS-PAGE and Western Blotting
2.3. Binding of Recombinant Stx1B by HeLa, HT-29, and Caco-2 Cells
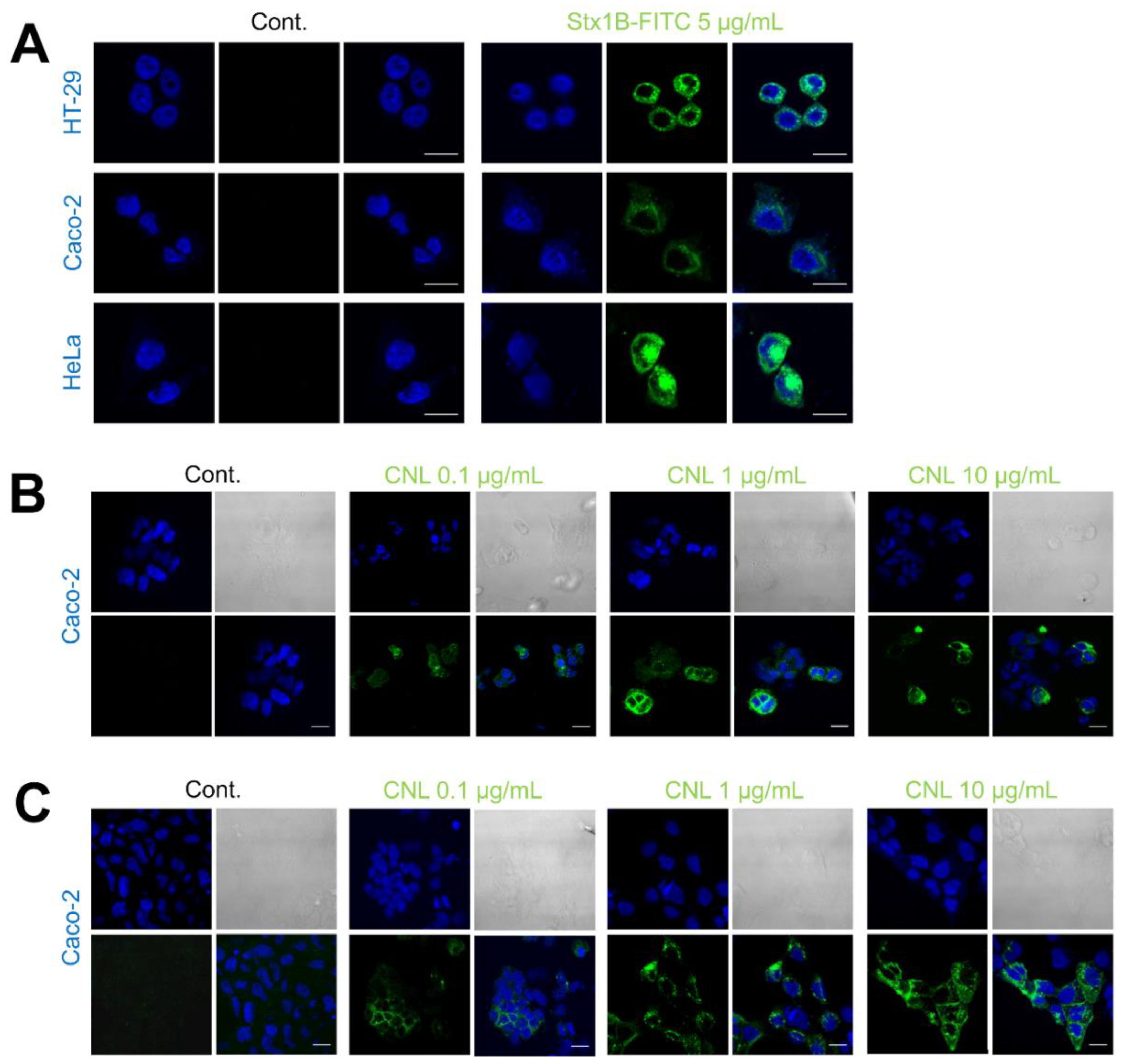
2.4. Immunofluorescence Staining
2.5. Confocal Microscopy
2.6. Cell Viability Assay
2.7. Bacterial Strains and Growth Conditions
2.8. Molecular Cloning
2.9. The Expression of Stx1B and CNL Fusion Proteins in L. lactis
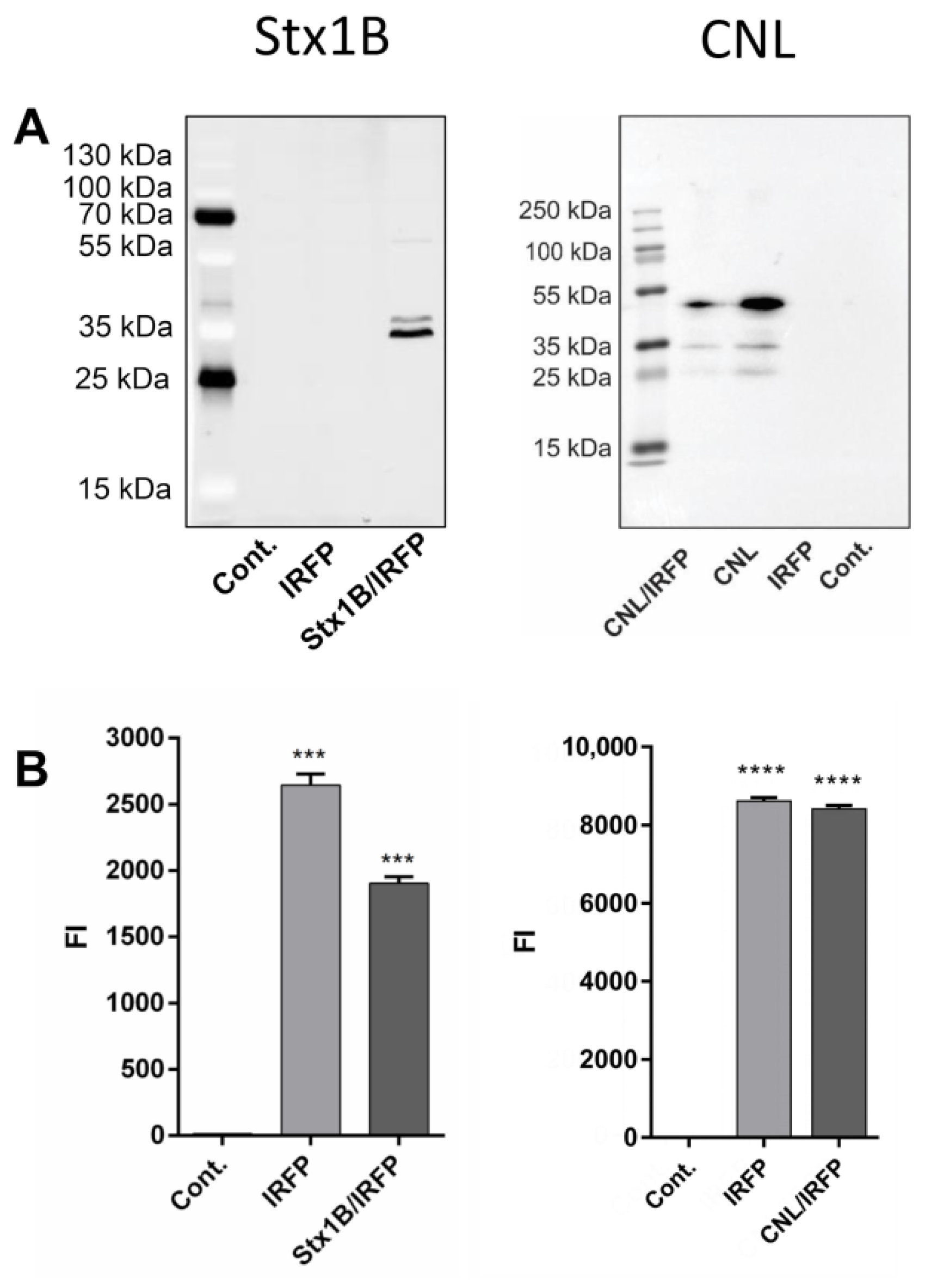
2.10. Fluorescence Measurements of IRFPs
2.11. Flow Cytometry
2.12. L. lactis Cell Adhesion Assay
2.13. Statistical Analyses
3. Results
3.1. CNL-Binding Glycoproteins in HT-29 and Caco-2 Whole-Cell Lysates
3.2. Binding of Recombinant Lectins to HeLa, HT-29, and Caco-2 Cancer Cells
3.3. The Effect of Stx1B and CNL Lectins on Caco-2 Cell Viability
3.4. Genetic Constructs for Stx1B and CNL Lectin Display on L. lactis
3.5. Expression of Lectin Fusion Proteins in L. lactis
3.6. Surface Display of Lectins on L. lactis
3.7. The Adhesion of Lectin-Displaying L. lactis to Cancer Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Žurga, S.; Perišić Nanut, M.; Kos, J.; Sabotič, J. Fungal lectin MpL enables entry of protein drugs into cancer cells and their subcellular targeting. Oncotarget 2017, 8, 26896–26910. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ho, W.L.; Hsu, W.M.; Huang, M.C.; Kadomatsu, K.; Nakagawara, A. Protein glycosylation in cancers and its potential therapeutic applications in neuroblastoma. J. Hematol. Oncol. 2016, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Brandley, B.K.; Schnaar, R.L. Cell-surface carbohydrates in cell recognition and response. J. Leukoc. Biol. 1986, 40, 97–111. [Google Scholar] [CrossRef] [PubMed]
- West, I.; Goldring, O. Lectin affinity chromatography. Methods Mol. Biol. 2004, 244, 159–166. [Google Scholar] [PubMed]
- Duverger, E.; Lamerant-Fayel, N.; Frison, N.; Monsigny, M. Carbohydrate-lectin interactions assayed by SPR. Methods Mol. Biol. 2010, 627, 157–178. [Google Scholar]
- Encarnacao, J.C.; Napolitano, V.; Opassi, G.; Danielson, U.H.; Dubin, G.; Popowicz, G.M.; Munier-Lehmann, H.; Buijs, J.; Andersson, K.; Bjorkelund, H. A real-time cell-binding assay reveals dynamic features of STxB-Gb3 cointernalization and STxB-mediated cargo delivery into cancer cells. FEBS Lett. 2020, 594, 2406–2420. [Google Scholar] [CrossRef]
- Geyer, P.E.; Maak, M.; Nitsche, U.; Perl, M.; Novotny, A.; Slotta-Huspenina, J.; Dransart, E.; Holtorf, A.; Johannes, L.; Janssen, K.P. Gastric adenocarcinomas express the glycosphingolipid Gb3/CD77: Targeting of gastric cancer cells with Shiga toxin B-subunit. Mol. Cancer Ther. 2016, 15, 1008–1017. [Google Scholar] [CrossRef]
- Luginbuehl, V.; Meier, N.; Kovar, K.; Rohrer, J. Intracellular drug delivery: Potential usefulness of engineered Shiga toxin subunit B for targeted cancer therapy. Biotechnol. Adv. 2018, 36, 613–623. [Google Scholar] [CrossRef]
- Bouzari, S.; Oloomi, M.; Azadmanesh, K. Study on induction of apoptosis on HeLa and Vero cells by recombinant shiga toxin and its subunits. Cytotechnology 2009, 60, 105. [Google Scholar] [CrossRef]
- Nakagawa, I.; Nakata, M.; Kawabata, S.; Hamada, S. Regulated expression of the Shiga toxin B gene induces apoptosis in mammalian fibroblastic cells. Mol. Microbiol. 1999, 33, 1190–1199. [Google Scholar] [CrossRef]
- Muro, S. Challenges in design and characterization of ligand-targeted drug delivery systems. J. Control. Release 2012, 164, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Devi, R.V.; Basil-Rose, M.R. Lectins as ligands for directing nanostructured systems. Curr. Drug Deliv. 2018, 15, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Kim, D.Y.; Szardenings, M.; Min, J.H.; Hong, Y.; Choy, H.E.; Min, J.J. RGD peptide cell-surface display enhances the targeting and therapeutic efficacy of attenuated Salmonella-mediated cancer therapy. Theranostics 2016, 6, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, Y.; Liu, X.; Min, J.J.; Tan, W.; Zheng, J.H. Targeted cancer immunotherapy with genetically engineered oncolytic Salmonella typhimurium. Cancer Lett. 2020, 469, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Nizet, V.; Varki, A.; Aebi, M. Microbial Lectins: Hemagglutinins, adhesins, and toxins. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Eds.; Cold Spring Harbor: New York, NY, USA, 2015; pp. 481–491. [Google Scholar] [CrossRef]
- Plavec, T.V.; Berlec, A. Safety aspects of genetically modified lactic acid bacteria. Microorganisms 2020, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Zahirović, A.; Lunder, M. Microbial delivery vehicles for allergens and allergen-derived peptides in immunotherapy of allergic diseases. Front. Microbiol. 2018, 9, 1449. [Google Scholar] [CrossRef] [PubMed]
- Asensi, G.F.; de Sales, N.F.; Dutra, F.F.; Feijo, D.F.; Bozza, M.T.; Ulrich, R.G.; Miyoshi, A.; de Morais, K.; Azevedo, V.A.; Silva, J.T.; et al. Oral immunization with Lactococcus lactis secreting attenuated recombinant staphylococcal enterotoxin B induces a protective immune response in a murine model. Microb. Cell Fact. 2013, 12, 32. [Google Scholar] [CrossRef]
- Lee, M.H.; Roussel, Y.; Wilks, M.; Tabaqchali, S. Expression of Helicobacter pylori urease subunit B gene in Lactococcus lactis MG1363 and its use as a vaccine delivery system against H. pylori infection in mice. Vaccine 2001, 19, 3927–3935. [Google Scholar] [CrossRef]
- Plavec, T.V.; Berlec, A. Engineering of lactic acid bacteria for delivery of therapeutic proteins and peptides. Appl. Microbiol. Biotechnol. 2019, 103, 2053–2066. [Google Scholar] [CrossRef]
- Kosler, S.; Štrukelj, B.; Berlec, A. Lactic acid bacteria with concomitant IL-17, IL-23 and TNF alpha-binding ability for the treatment of inflammatory bowel disease. Curr. Pharm. Biotechnol. 2017, 18, 318–326. [Google Scholar] [CrossRef]
- Škrlec, K.; Pucer Janež, A.; Rogelj, B.; Štrukelj, B.; Berlec, A. Evasin-displaying lactic acid bacteria bind different chemokines and neutralize CXCL8 production in Caco-2 cells. Microb. Biotechnol. 2017, 10, 1732–1743. [Google Scholar] [CrossRef] [PubMed]
- Sreerohini, S.; Balakrishna, K.; Parida, M. Oral immunization of mice with Lactococcus lactis expressing Shiga toxin truncate confers enhanced protection against Shiga toxins of Escherichia coli O157:H7 and Shigella dysenteriae. Apmis 2019, 127, 671–680. [Google Scholar] [CrossRef]
- Zadravec, P.; Mareckova, L.; Petrokova, H.; Hodnik, V.; Perišić Nanut, M.; Anderluh, G.; Štrukelj, B.; Maly, P.; Berlec, A. Development of recombinant Lactococcus lactis displaying albumin-binding domain variants against Shiga toxin 1 B subunit. PLoS ONE 2016, 11, e0162625. [Google Scholar] [CrossRef] [PubMed]
- Engedal, N.; Skotland, T.; Torgersen, M.L.; Sandvig, K. Shiga toxin and its use in targeted cancer therapy and imaging. Microb. Biotechnol. 2011, 4, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, P.B.; Chaturvedi, P.; Fine, R.E.; Ritchie, A.J.; Pober, J.S.; Cleary, T.G.; Newburg, D.S. Tumor necrosis factor alpha increases human cerebral endothelial cell Gb3 and sensitivity to Shiga toxin. Infect. Immun. 2001, 69, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Pohlentz, G.; Schmitz, J.S.; Steil, D.; Humpf, H.U.; Karch, H.; Muthing, J. Shiga toxin glycosphingolipid receptors in human Caco-2 and HCT-8 colon epithelial cell lines. Toxins 2017, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Falguieres, T.; Mallard, F.; Baron, C.; Hanau, D.; Lingwood, C.; Goud, B.; Salamero, J.; Johannes, L. Targeting of Shiga toxin B-subunit to retrograde transport route in association with detergent-resistant membranes. Mol. Biol. Cell 2001, 12, 2453–2468. [Google Scholar] [CrossRef] [PubMed]
- Sabotič, J.; Kos, J. CNL-Clitocybe nebularis lectin-the fungal GalNAcbeta1-4GlcNAc-binding lectin. Molecules 2019, 24, 4204. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Renko, M.; Magister, Š.; Smith, D.F.; Künzler, M.; Štrukelj, B.; Turk, D.; Kos, J.; Sabotič, J. Bivalent carbohydrate binding is required for biological activity of Clitocybe nebularis lectin (CNL), the N,N’-diacetyllactosediamine (GalNAcbeta1-4GlcNAc, LacdiNAc)-specific lectin from basidiomycete C. nebularis. J. Biol. Chem. 2012, 287, 10602–10612. [Google Scholar] [CrossRef]
- Hirano, K.; Matsuda, A.; Shirai, T.; Furukawa, K. Expression of LacdiNAc groups on N-glycans among human tumors is complex. Biomed. Res. Int. 2014, 2014, 981627. [Google Scholar] [CrossRef]
- Peracaula, R.; Tabarés, G.; Royle, L.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M.; de Llorens, R. Altered glycosylation pattern allows the distinction between prostate-specific antigen (PSA) from normal and tumor origins. Glycobiology 2003, 13, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Peracaula, R.; Royle, L.; Tabarés, G.; Mallorquí-Fernández, G.; Barrabés, S.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M.; de Llorens, R. Glycosylation of human pancreatic ribonuclease: Differences between normal and tumor states. Glycobiology 2003, 13, 227–244. [Google Scholar] [CrossRef] [PubMed]
- Machado, E.; Kandzia, S.; Carilho, R.; Altevogt, P.; Conradt, H.S.; Costa, J. N-Glycosylation of total cellular glycoproteins from the human ovarian carcinoma SKOV3 cell line and of recombinantly expressed human erythropoietin. Glycobiology 2011, 21, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liang, J.T.; Huang, H.C.; Shen, T.L.; Chen, H.Y.; Lin, N.Y.; Che, M.I.; Lin, W.C.; Huang, M.C. Beta1,4-N-acetylgalactosaminyltransferase III enhances malignant phenotypes of colon cancer cells. Mol. Cancer Res. 2007, 5, 543–552. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Pohleven, J.; Obermajer, N.; Sabotič, J.; Anzlovar, S.; Sepčić, K.; Kos, J.; Kralj, B.; Štrukelj, B.; Brzin, J. Purification, characterization and cloning of a ricin B-like lectin from mushroom Clitocybe nebularis with antiproliferative activity against human leukemic T cells. Biochim. Biophys. Acta 2009, 1790, 173–181. [Google Scholar] [CrossRef]
- Mierau, I.; Kleerebezem, M. 10 years of the nisin-controlled gene expression system (NICE) in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2005, 68, 705–717. [Google Scholar] [CrossRef]
- De Ruyter, P.G.; Kuipers, O.P.; de Vos, W.M. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol. 1996, 62, 3662–3667. [Google Scholar] [CrossRef]
- Kuipers, O.P.; Beerthuyzen, M.M.; Siezen, R.J.; De Vos, W.M. Characterization of the nisin gene cluster nisABTCIPR of Lactococcus lactis. Requirement of expression of the nisA and nisI genes for development of immunity. Eur. J. Biochem. 1993, 216, 281–291. [Google Scholar] [CrossRef]
- Berlec, A.; Škrlec, K.; Kocjan, J.; Olenic, M.; Štrukelj, B. Single plasmid systems for inducible dual protein expression and for CRISPR-Cas9/CRISPRi gene regulation in lactic acid bacterium Lactococcus lactis. Sci. Rep. 2018, 8, 1009. [Google Scholar] [CrossRef]
- Škrlec, K.; Zadravec, P.; Hlavnickova, M.; Kuchar, M.; Vankova, L.; Petrokova, H.; Krizova, L.; Cerny, J.; Berlec, A.; Maly, P. p19-targeting ILP protein blockers of IL-23/Th-17 pro-inflammatory axis displayed on engineered bacteria of food origin. Int. J. Mol. Sci. 2018, 19, 1933. [Google Scholar] [CrossRef] [PubMed]
- Plavec, T.V.; Kuchar, M.; Benko, A.; Liskova, V.; Cerny, J.; Berlec, A.; Maly, P. Engineered Lactococcus lactis secreting IL-23 receptor-targeted REX protein blockers for modulation of IL-23/Th17-mediated inflammation. Microorganisms 2019, 7, 152. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, Y.; Sato, T.; Niwa, T.; Nakamura, S.; Gotoh, M.; Ikehara, S.K.; Kiyohara, K.; Aoki, C.; Iwai, T.; Nakanishi, H.; et al. Apical golgi localization of N,N’-diacetyllactosediamine synthase, beta4GalNAc-T3, is responsible for LacdiNAc expression on gastric mucosa. Glycobiology 2006, 16, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Ravnikar, M.; Štrukelj, B.; Obermajer, N.; Lunder, M.; Berlec, A. Engineered lactic acid bacterium Lactococcus lactis capable of binding antibodies and tumor necrosis factor alpha. Appl. Environ. Microbiol. 2010, 76, 6928–6932. [Google Scholar] [CrossRef]
- Willysson, A.; Ståhl, A.L.; Gillet, D. Shiga toxin uptake and sequestration in extracellular vesicles is mediated by its B-Subunit. Toxins 2020, 12, 449. [Google Scholar] [CrossRef]
- Keusch, G.T.; Jacewicz, M.; Acheson, D.W.; Donohue-Rolfe, A.; Kane, A.V.; McCluer, R.H. Globotriaosylceramide, Gb3, is an alternative functional receptor for Shiga-like toxin 2e. Infect. Immun. 1995, 63, 1138–1141. [Google Scholar] [CrossRef] [PubMed]
- Falguieres, T.; Maak, M.; von Weyhern, C.; Sarr, M.; Sastre, X.; Poupon, M.F.; Robine, S.; Johannes, L.; Janssen, K.P. Human colorectal tumors and metastases express Gb3 and can be targeted by an intestinal pathogen-based delivery tool. Mol. Cancer Ther. 2008, 7, 2498–2508. [Google Scholar] [CrossRef]
- Kovbasnjuk, O.; Mourtazina, R.; Baibakov, B.; Wang, T.; Elowsky, C.; Choti, M.A.; Kane, A.; Donowitz, M. The glycosphingolipid globotriaosylceramide in the metastatic transformation of colon cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 19087–19092. [Google Scholar] [CrossRef]
- Pirro, M.; Rombouts, Y.; Stella, A.; Neyrolles, O.; Burlet-Schiltz, O.; van Vliet, S.J.; de Ru, A.H.; Mohammed, Y.; Wuhrer, M.; van Veelen, P.A.; et al. Characterization of macrophage galactose-type lectin (MGL) ligands in colorectal cancer cell lines. Biochim. Biophys. Acta 2020, 1864, 129513. [Google Scholar] [CrossRef]
- Che, M.I.; Huang, J.; Hung, J.S.; Lin, Y.C.; Huang, M.J.; Lai, H.S.; Hsu, W.M.; Liang, J.T.; Huang, M.C. β1, 4-N-acetylgalactosaminyltransferase III modulates cancer stemness through EGFR signaling pathway in colon cancer cells. Oncotarget 2014, 5, 3673–3684. [Google Scholar] [CrossRef]
- Bae, Y.H.; Park, K. Targeted drug delivery to tumors: Myths, reality and possibility. J. Control. Release 2011, 153, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Weissenbock, A.; Wirth, M.; Gabor, F. WGA-grafted PLGA-nanospheres: Preparation and association with Caco-2 single cells. J. Control. Release 2004, 99, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Brzin, J.; Vrabec, L.; Leonardi, A.; Cokl, A.; Štrukelj, B.; Kos, J.; Sabotič, J. Basidiomycete Clitocybe nebularis is rich in lectins with insecticidal activities. Appl. Microbiol. Biotechnol. 2011, 91, 1141–1148. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain, Primer, or Plasmid | Relevant Features or Sequence | Reference |
|---|---|---|
| Strains | ||
| E. coli DH5α | endA1 glnV44 thi-1 recA1 relA1 gyrA96 deoR F− Φ80dlacZΔM15 Δ(lacZYA-argF)U169, hsdR17(rK− mK+), λ– | Invitrogen |
| L. lactis NZ9000 | MG1363 nisRK ΔpepN | NIZO |
| Primers | ||
| Stx1B-F-Bam | 5′-AGGATCCAAAAAAACATTATTAATAGCTGCATC-3′ | This work |
| Stx1B-R-Eco | 5′-AGAATTCACGAAAAATAACTTCGCTG-3′ | This work |
| CNL-F-Bam | 5′-TGGATCCTCTATTACACCTGGTACTTATAATATTAC-3′ | This work |
| CNL-R-Eco | 5′-AGAATTCTACGGCAGAGACACTTTC-3′ | This work |
| Genes | ||
| CNL | GGATCCTCTATTACACCTGGTACTTATAATATTACAAATGTTGCTTAT ACAAACAGATTGATTGATTTGACAGGTAGTAATCCTGCTGAAAATA CACTTATTATCGGTCATCATCTTAACAAAACACCTTCAGGTTATGGA AATCAACAGTGGACACTTGTCCAGCGACCACACACAACTATCTATA CTATGCAAGCAGTTAATCCACAATCTTATGTACGAGTTCGTGATGAT AATTTAGTTGACGGAGCAGCACTTGTAGGAAGTCAACAGCCTACAC CTGTCAGTATTGAATCAGCCGGAAATTCAGGTCAATTTCGAATTAAA ATTCCAGATTTAGGTTTAGCTTTAACTTTACCTTCAGACGCAAATAGT ACTCCTATTGTACTTGGAGAAGTTGATGAAACATCTACTAATCAATTG TGGGCATTTGAAAGTGTCTCTGCCGTAGAATTC | This work |
| Plasmids | ||
| pGEM-T Easy | Apr, cloning vector for PCR products | Promega |
| pET28-Stx1B | pET28b containing Stx1B gene | [24] |
| pNZ8148 | pSH71 derivative, PnisA, Cmr, nisin-controlled expression | [38,39,40] |
| pSDBA3b | pNZ8148 containing gene fusion of spUsp45, b-dom, and acmA3b | [22] |
| pSD-Stx1B | pNZ8148 containing gene fusion of spUsp45, flag, stx1B, and acmA3b | This work |
| pSD-CNL | pNZ8148 containing gene fusion of spUsp45, cnl, and acmA3b | This work |
| pNZDual | pNZ8148 containing two multiple cloning sites (MCS1 and MCS2) | [41] |
| pNZD-IRFP | pNZDual with irfp in MCS2 | [41] |
| pNZD-Stx1B | pNZDual with flag-stx1B in MCS1 | This work |
| pNZD-CNL | pNZDual with cnl in MCS1 | This work |
| pNZD-Stx1B-IRFP | pNZDual with flag-stx1B in MCS1 and irfp in MCS2 | This work |
| pNZD-CNL-IRFP | pNZDual with cnl in MCS1 and irfp in MCS2 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plavec, T.V.; Zahirović, A.; Zadravec, P.; Sabotič, J.; Berlec, A. Lectin-Mediated Binding of Engineered Lactococcus lactis to Cancer Cells. Microorganisms 2021, 9, 223. https://doi.org/10.3390/microorganisms9020223
Plavec TV, Zahirović A, Zadravec P, Sabotič J, Berlec A. Lectin-Mediated Binding of Engineered Lactococcus lactis to Cancer Cells. Microorganisms. 2021; 9(2):223. https://doi.org/10.3390/microorganisms9020223
Chicago/Turabian StylePlavec, Tina Vida, Abida Zahirović, Petra Zadravec, Jerica Sabotič, and Aleš Berlec. 2021. "Lectin-Mediated Binding of Engineered Lactococcus lactis to Cancer Cells" Microorganisms 9, no. 2: 223. https://doi.org/10.3390/microorganisms9020223
APA StylePlavec, T. V., Zahirović, A., Zadravec, P., Sabotič, J., & Berlec, A. (2021). Lectin-Mediated Binding of Engineered Lactococcus lactis to Cancer Cells. Microorganisms, 9(2), 223. https://doi.org/10.3390/microorganisms9020223