
Chromium Stress Tolerance of a C4 (Zea mays L.) and C3 (Vigna radiata L.) Plants Primed with UV and Gamma-Treated Bacillus subtilis


Abstract
:
1. Introduction
2. Materials and Methods
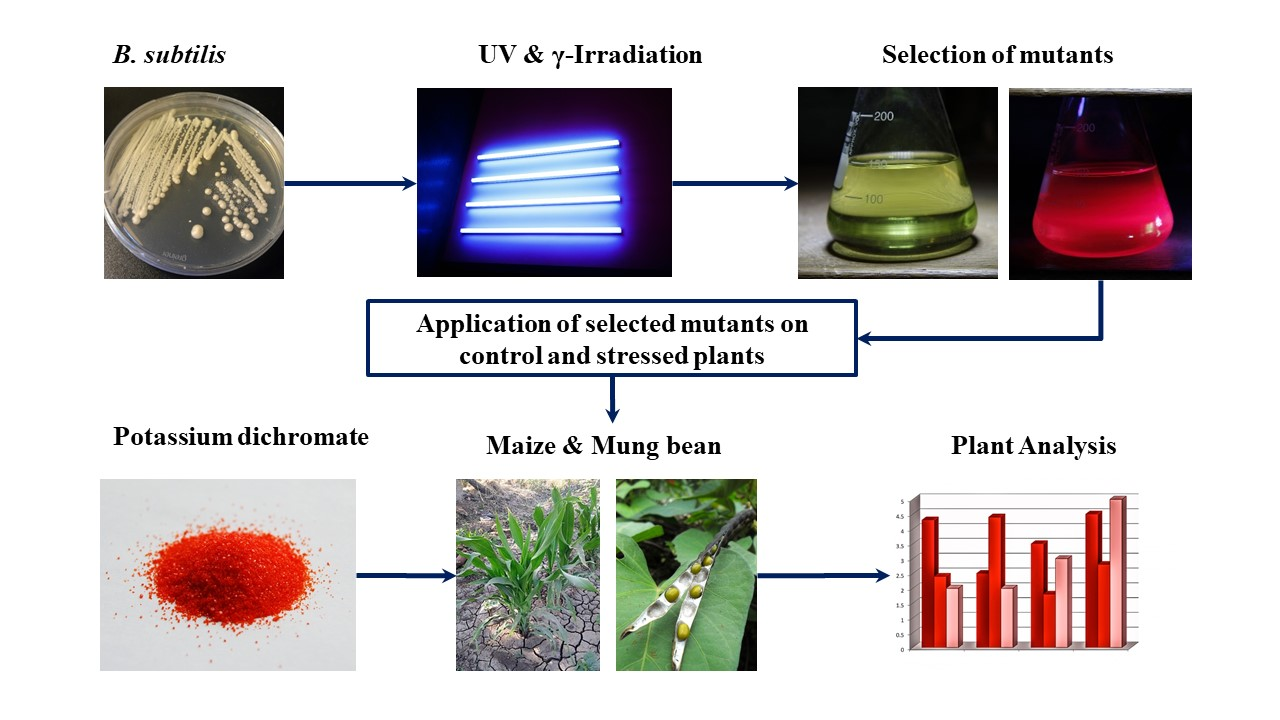
2.1. Mutagenic Treatments and Selection of Mutants
2.2. Minimum Inhibitory Concentration
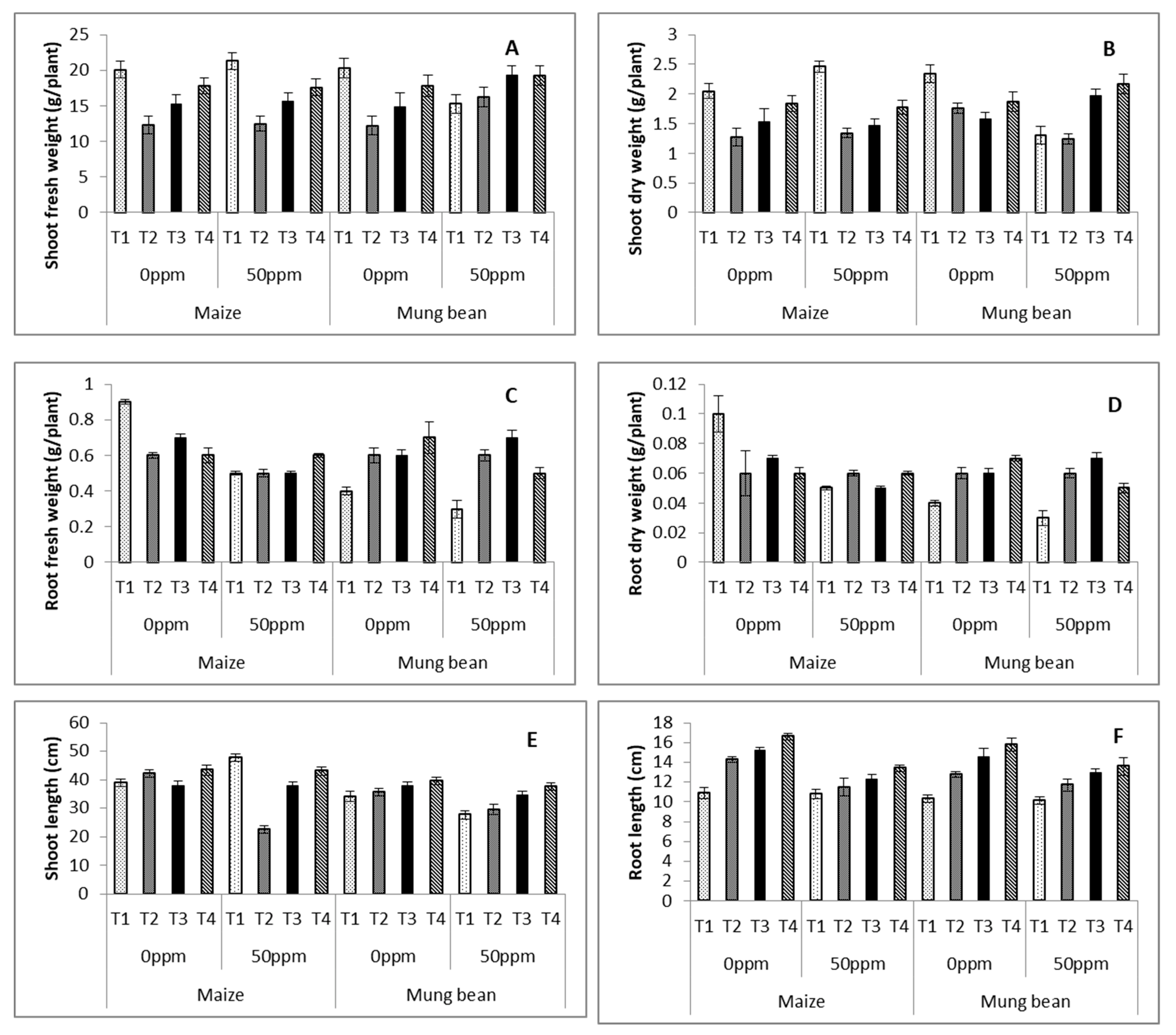
2.3. Field Experiment
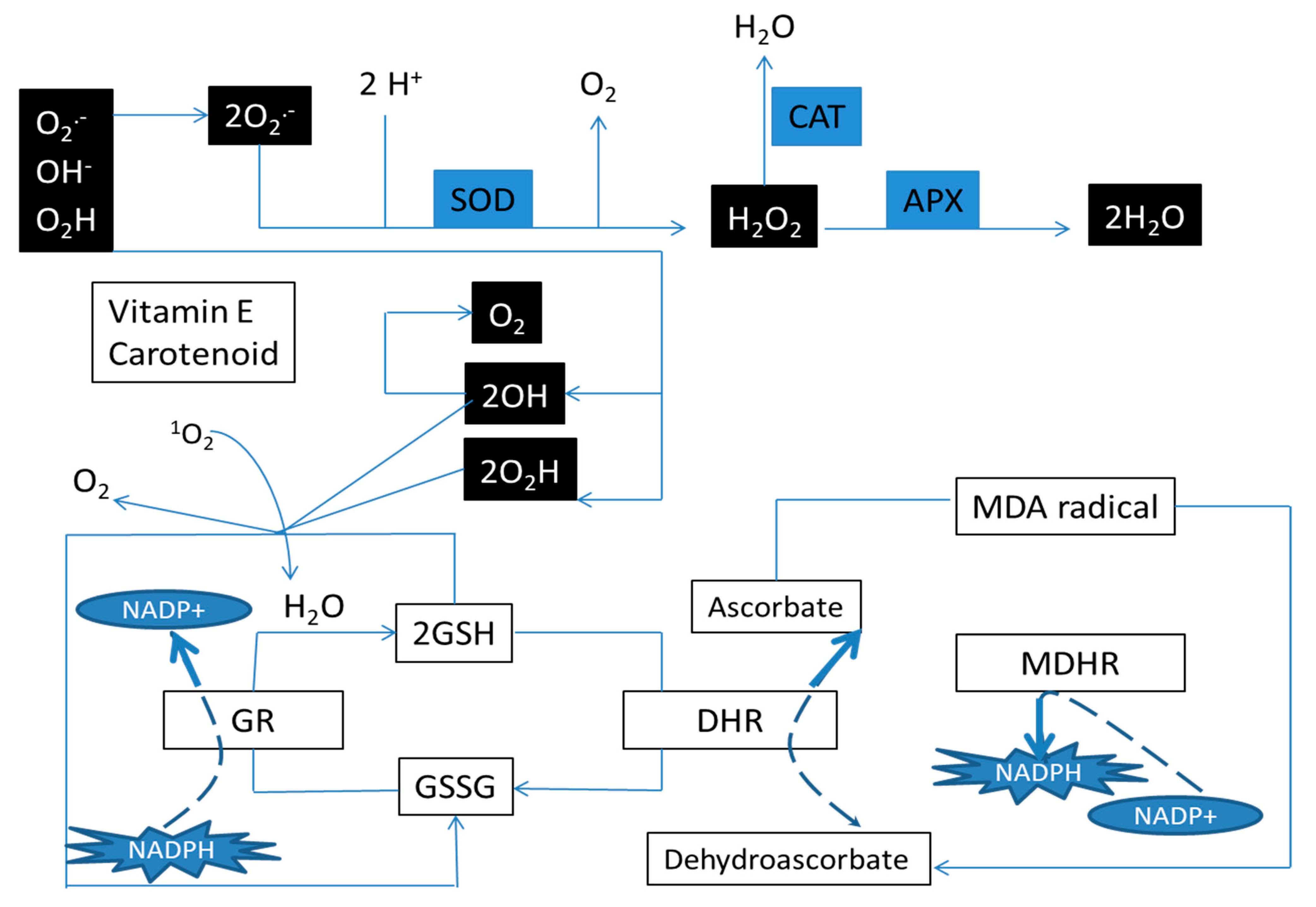
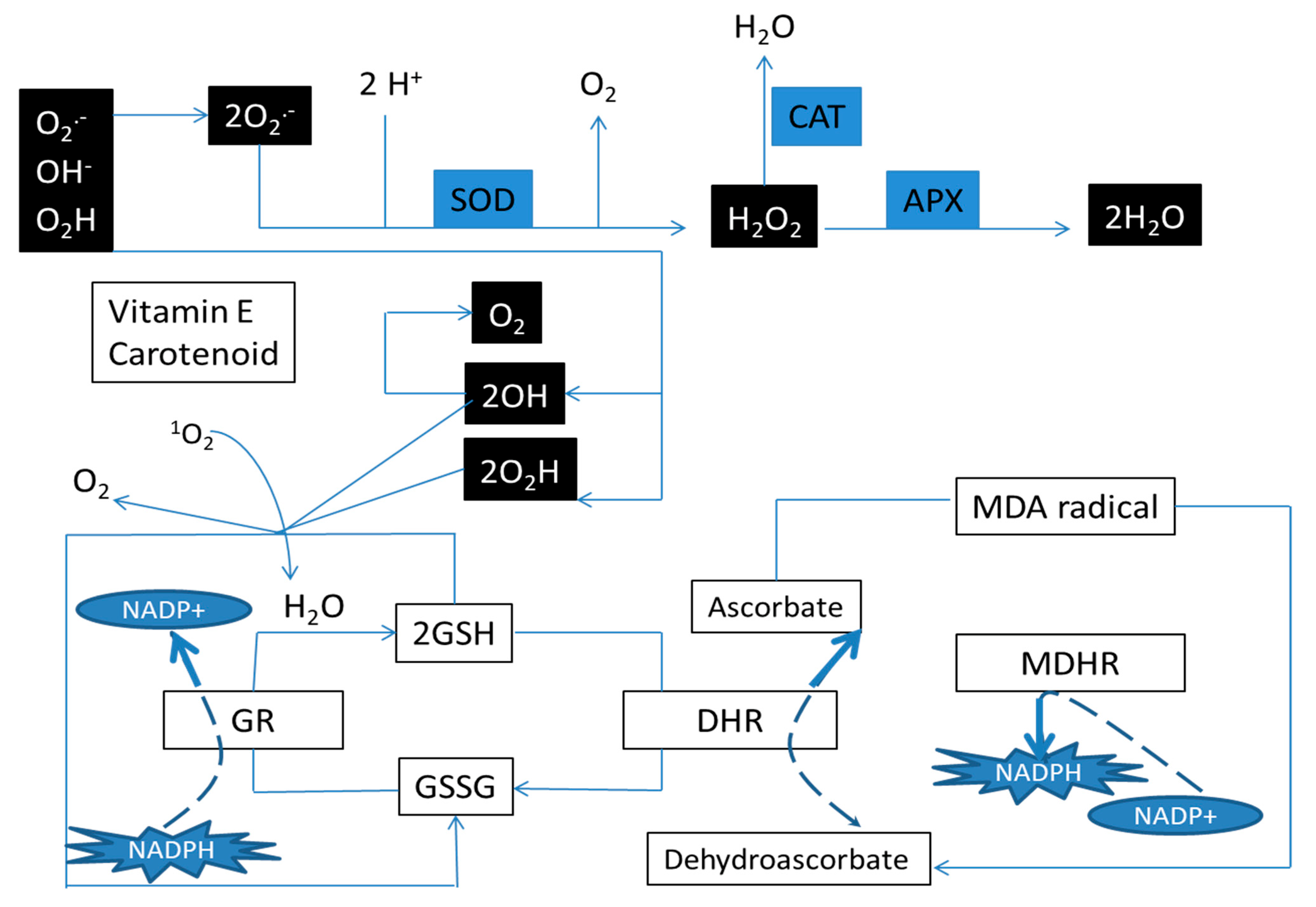
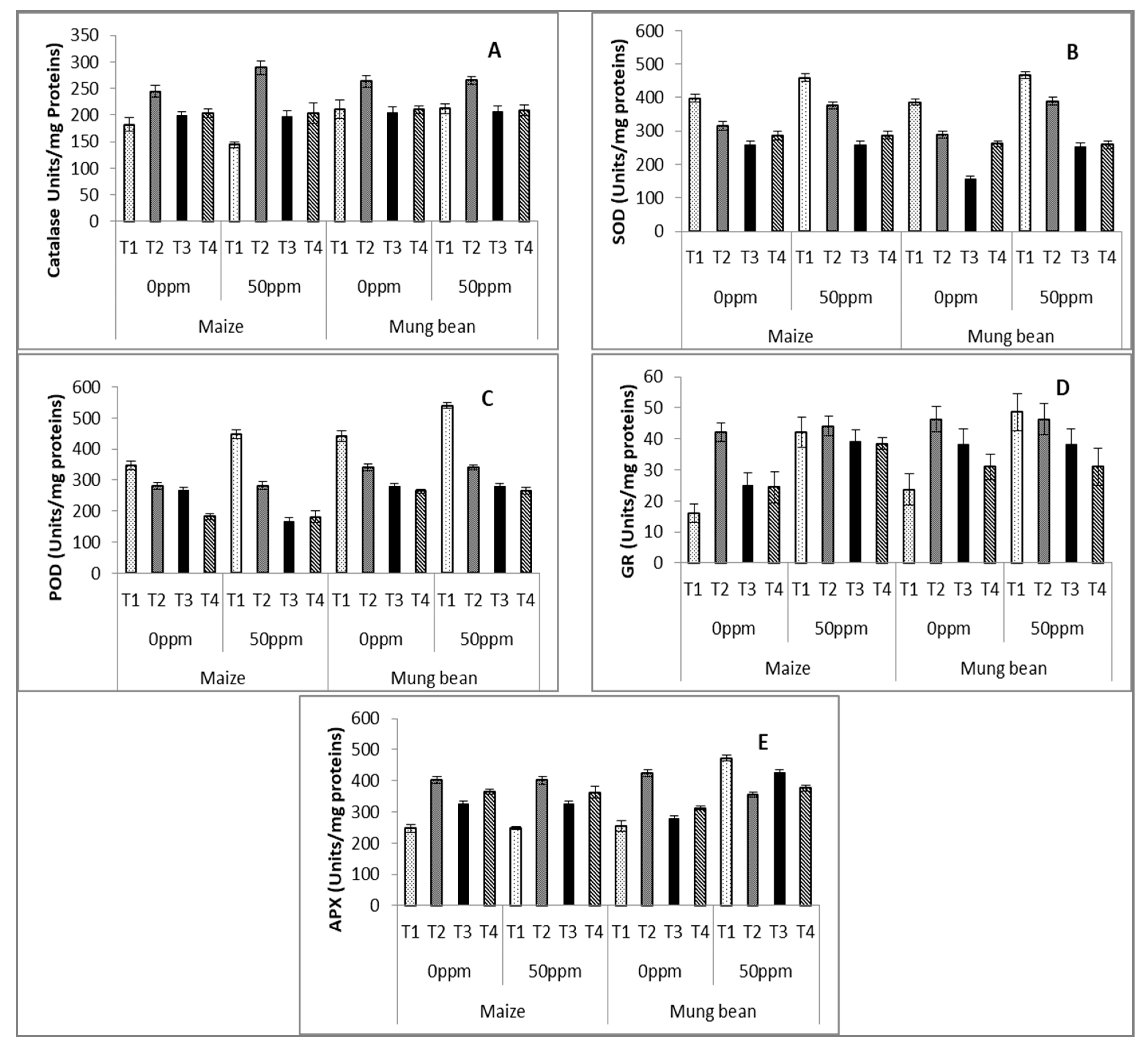
2.4. Antioxidant Enzymes
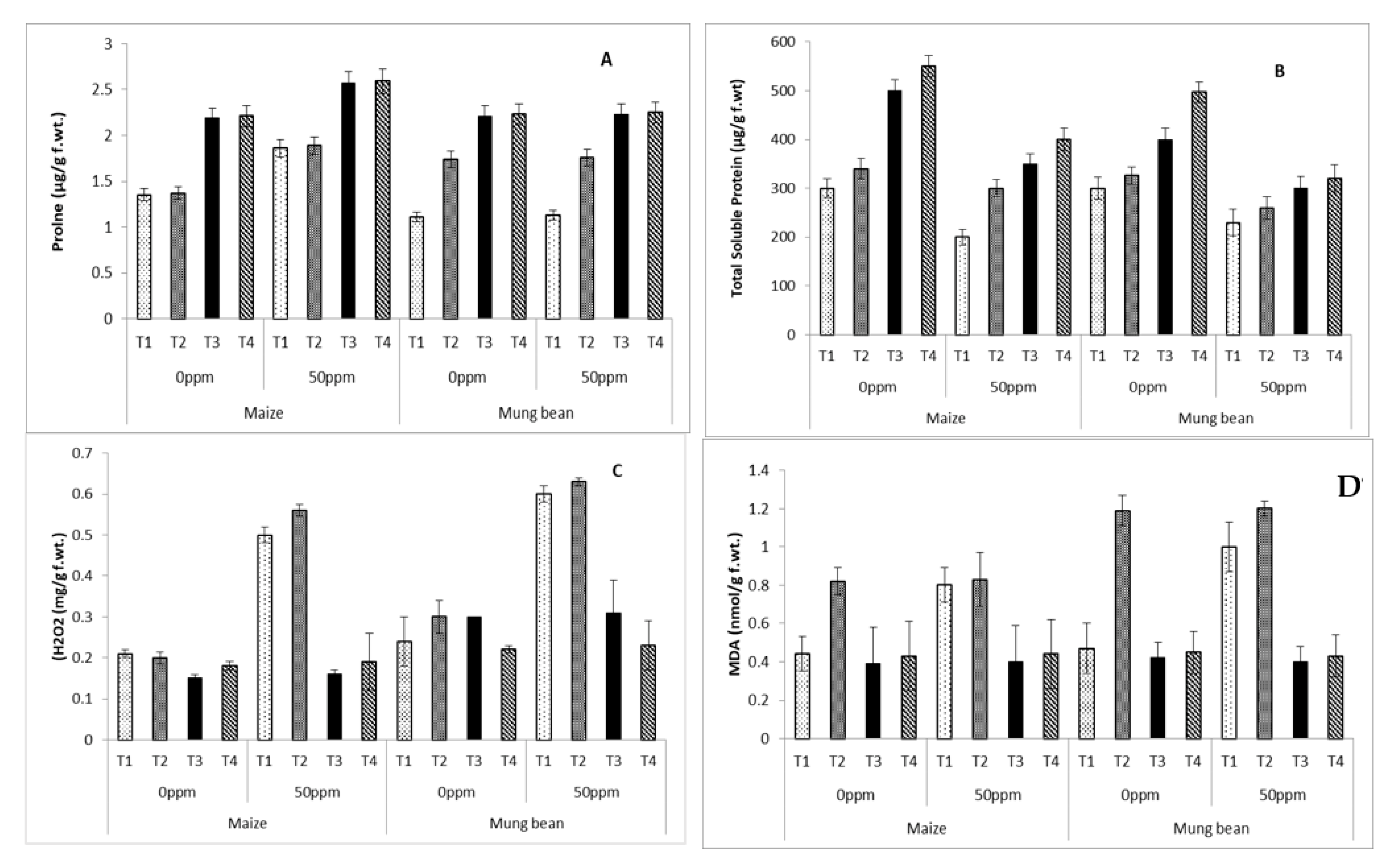
2.5. Lipid Peroxidation
2.6. Osmolyte and Protein Content
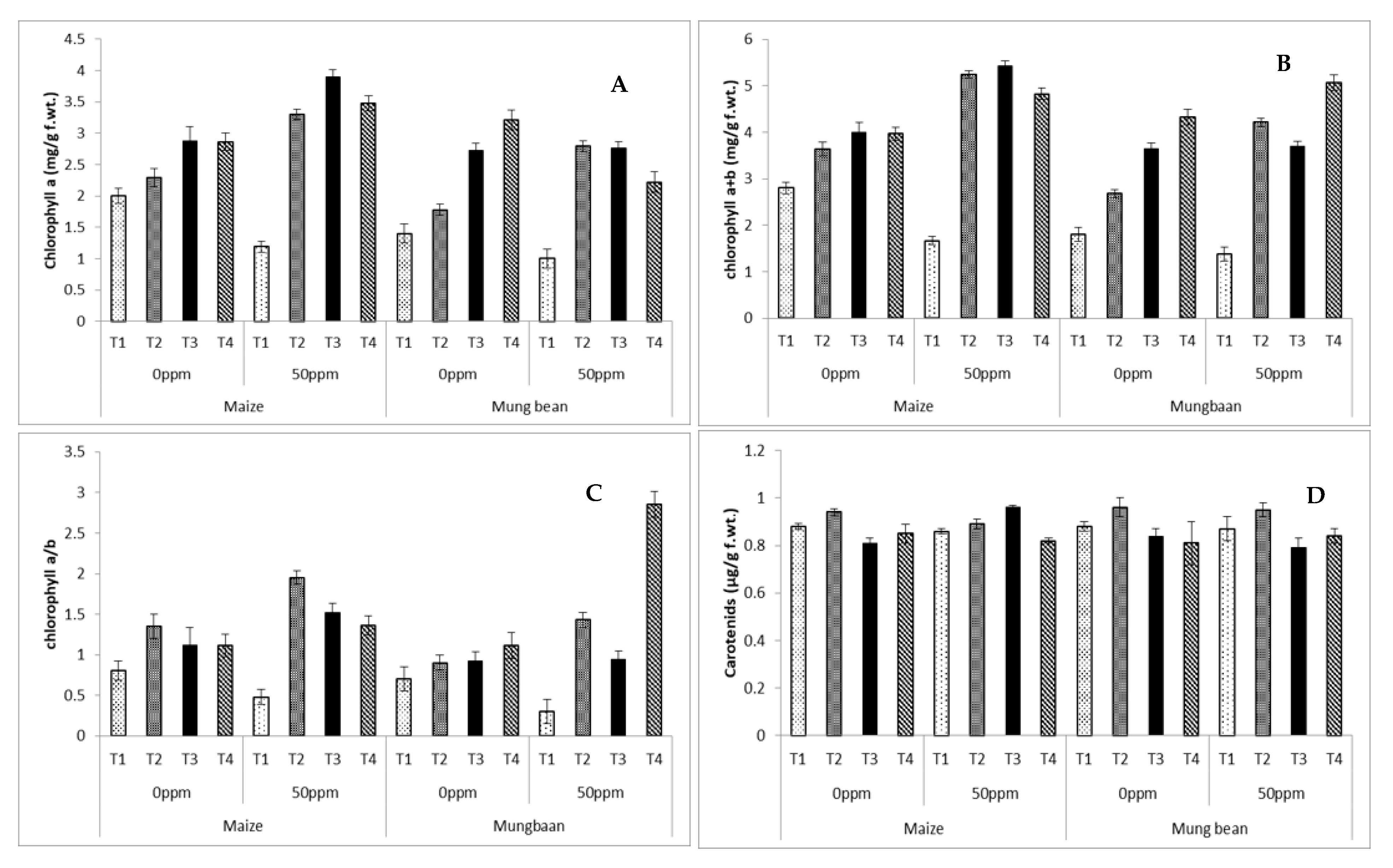
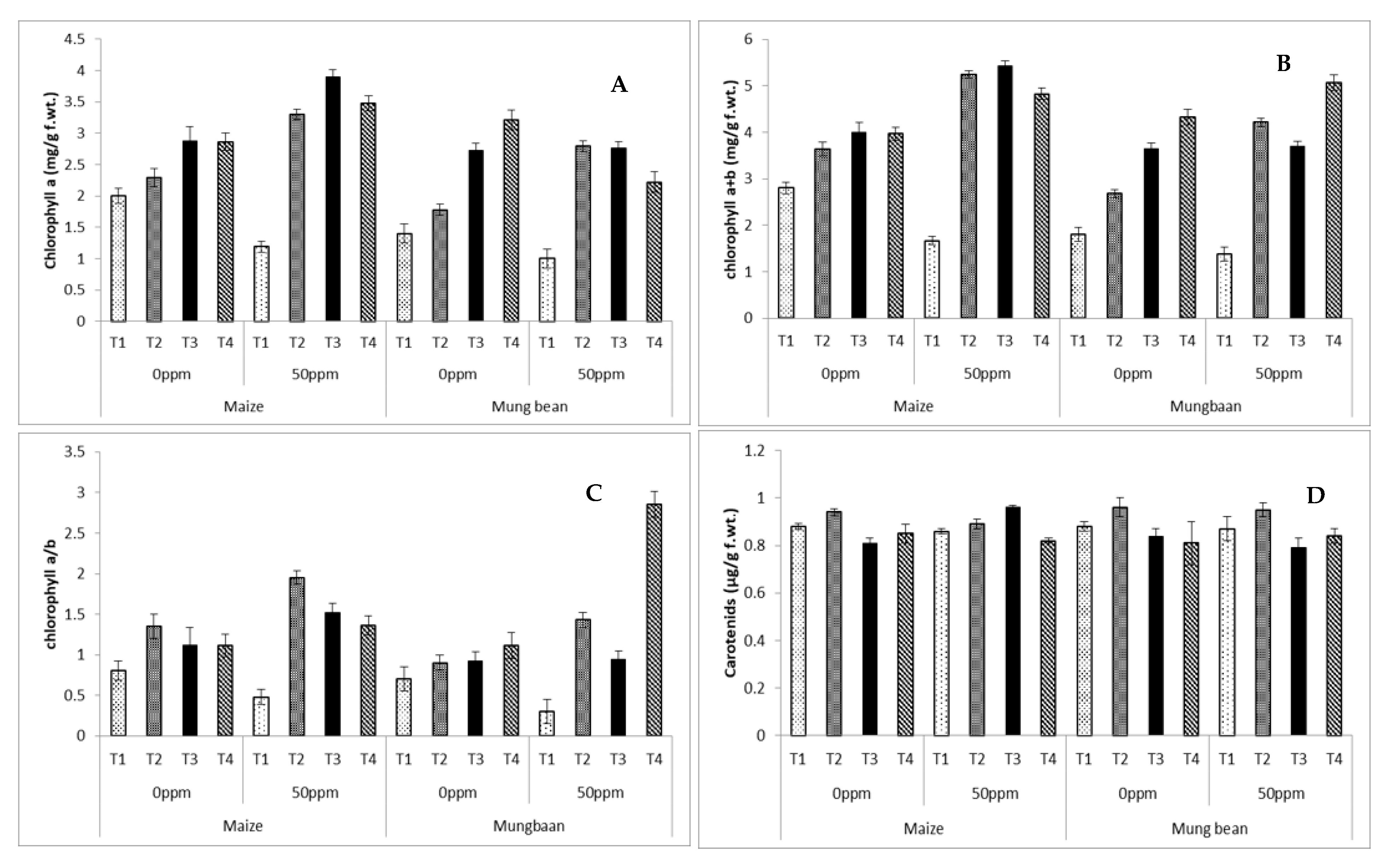
2.7. Photosynthetic Pigments
2.8. Statistical Analysis
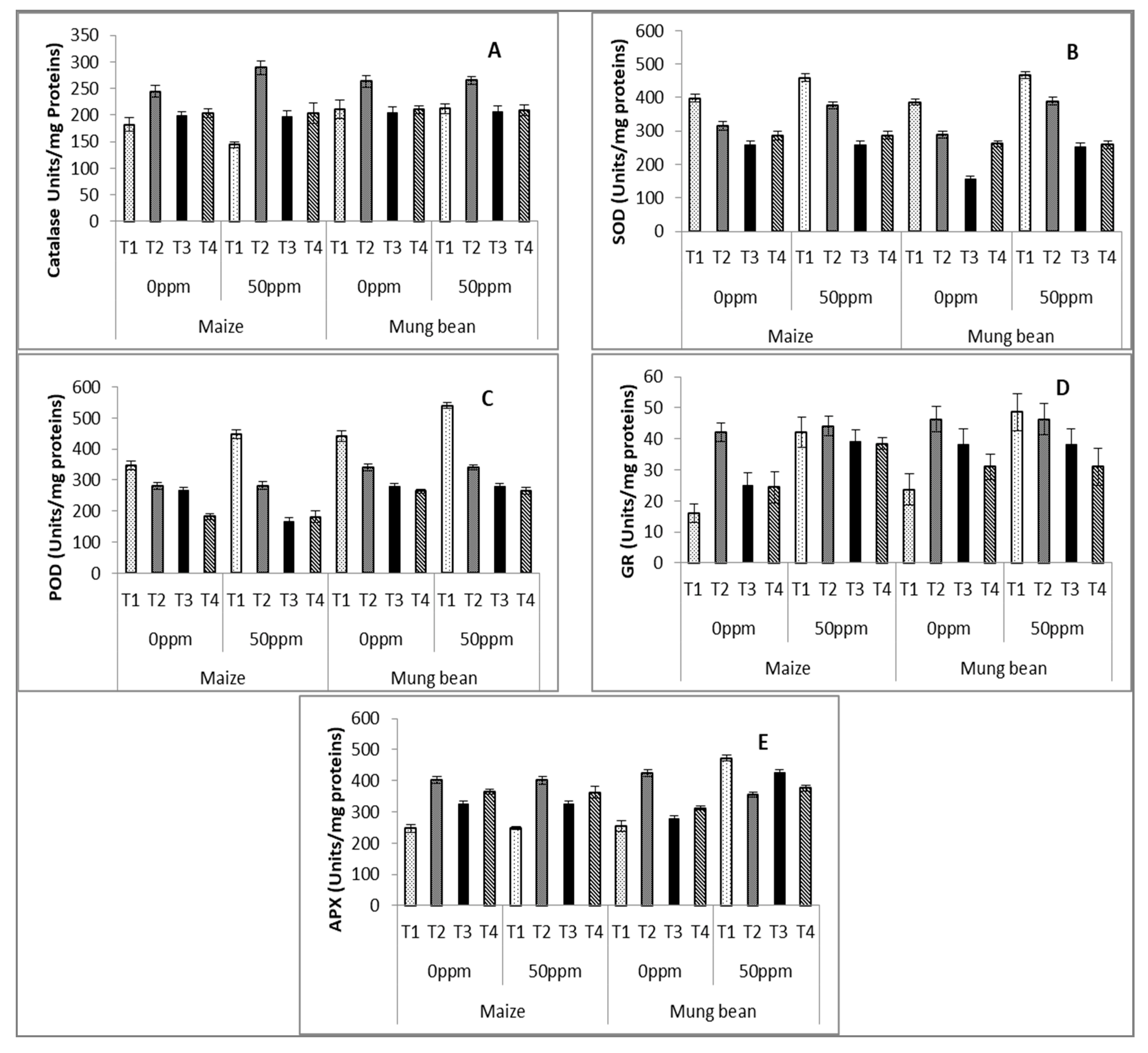
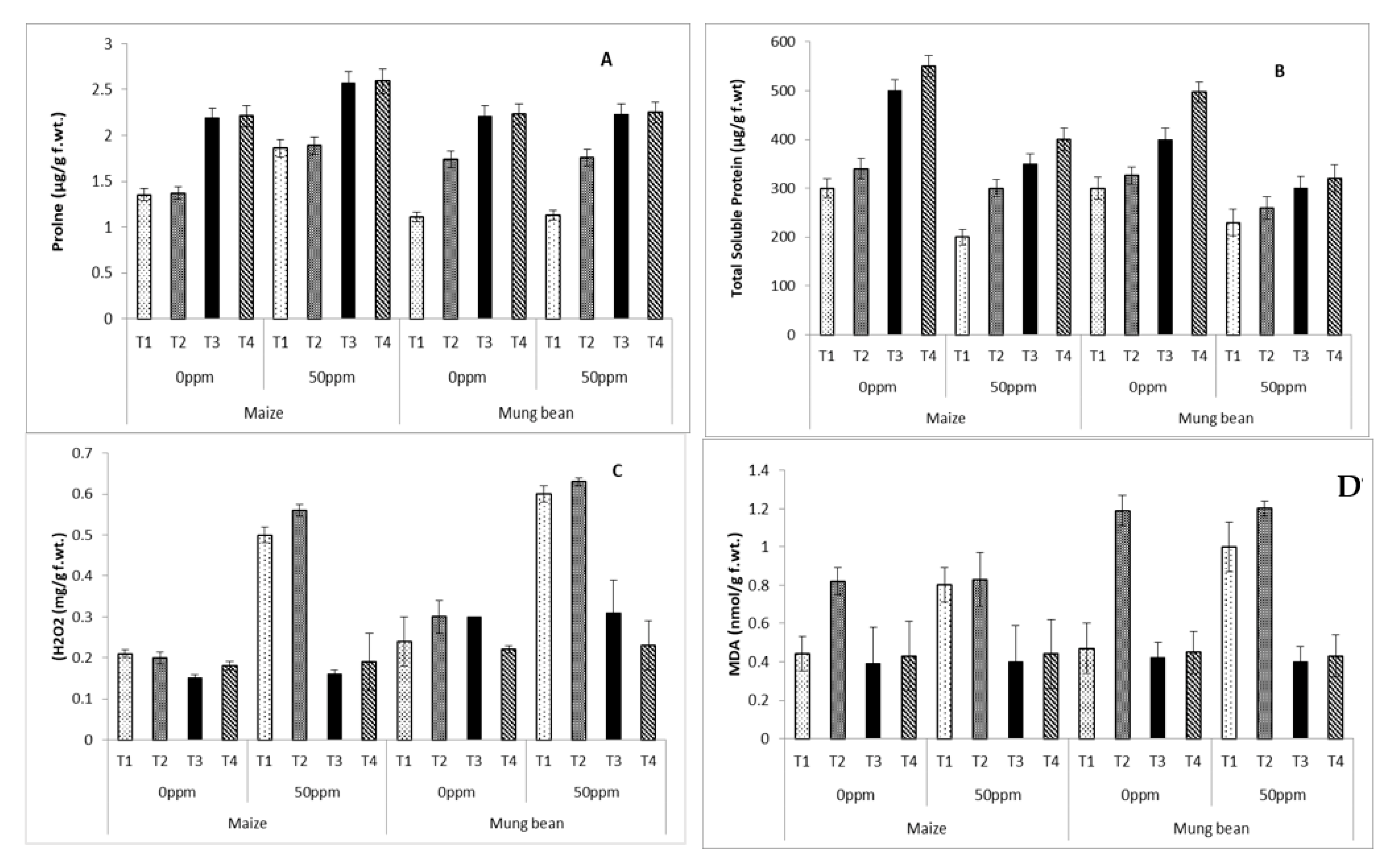
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Wang, R.; Gao, F.; Guo, B.-Q.; Huang, J.-C.; Wang, L.; Zhou, Y.-J. Short-term chromium-stress-induced alterations in the maize leaf proteome. Int. J. Mol. Sci. 2013, 14, 11125–11144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farid, M.; Shakoor, M.B.; Ehsan, S.; Ali, S.; Zubair, M.; Hanif, M. Morphological, physiological and biochemical responses of different plant species to Cd stress. Int. J. Chem. Biochem. Sci. 2013, 3, 53–60. [Google Scholar]
- Dey, S.; Mukhopadhyay, T.; Naskar, S.; Dey, T.; Chalak, H.; Adhikari, S. Probabilistic characterisation for dynamics and stability of laminated soft core sandwich plates. J. Sandw. Struct. Mater. 2019, 21, 366–397. [Google Scholar] [CrossRef]
- Ranieri, E.; Gikas, P. Effects of plants for reduction and removal of hexavalent chromium from a contaminated soil. Water Air Soil Pollut. 2014, 225, 1981. [Google Scholar] [CrossRef]
- Babula, P.; Adam, V.; Opatrilova, R.; Zehnalek, J.; Havel, L.; Kizek, R. Uncommon heavy metals, metalloids and their plant toxicity: A review. In Organic Farming, Pest Control and Remediation of Soil Pollutant; Springer: Cham, Switzerland, 2009; pp. 275–317. [Google Scholar]
- Rodriguez, E.; Santos, C.; Azevedo, R.; Moutinho-Pereira, J.; Correia, C.; Dias, M.C. Chromium (VI) induces toxicity at different photosynthetic levels in pea. Plant Physiol. Biochem. 2012, 53, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Dixit, V.; Pandey, V.; Shyam, R. Chromium ions inactivate electron transport and enhance superoxide generation in vivo in pea (Pisum sativum L. cv. Azad) root mitochondria. Plant Cell Environ. 2002, 25, 687–693. [Google Scholar] [CrossRef]
- Choudhury, S.; Panda, S.K. Toxic effects, oxidative stress and ultrastructural changes in moss Taxithelium nepalense (Schwaegr.) Broth. under chromium and lead phytotoxicity. Water Air Soil Pollut. 2005, 167, 73–90. [Google Scholar] [CrossRef]
- Brunetti, G.; Farrag, K.; Rovira, P.S.; Nigro, F.; Senesi, N. Greenhouse and field studies on Cr, Cu, Pb and Zn phytoextraction by Brassica napus from contaminated soils in the Apulia region, Southern Italy. Geoderma 2011, 160, 517–523. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Ashraf, A.; Sarfraz, R.A.; Mahmood, A.; ud Din, M. Chemical composition and in vitro antioxidant and antitumor activities of Eucalyptus camaldulensis Dehn. leaves. Ind. Crop. Prod. 2015, 74, 241–248. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.-Z.; Peng, T.; Huang, X.-S.; Fan, Q.-J.; Liu, J.-H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Freitas, H. Effects of inoculation of plant-growth promoting bacteria on Ni uptake by Indian mustard. Bioresour. Technol. 2008, 99, 3491–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Xu, T.; Chen, L.; Chen, J.; Rao, C.; Xiao, X.; Wan, Y.; Zeng, G.; Long, F.; Liu, C. Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. SLS18. Appl. Microbiol. Biotechnol. 2012, 93, 1745–1753. [Google Scholar] [CrossRef]
- Idris, R.; Trifonova, R.; Puschenreiter, M.; Wenzel, W.W.; Sessitsch, A. Bacterial communities associated with flowering plants of the Ni hyperaccumulator Thlaspi goesingense. Appl. Environ. Microbiol. 2004, 70, 2667–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Shanab, R.; Angle, J.; Chaney, R. Bacterial inoculants affecting nickel uptake by Alyssum murale from low, moderate and high Ni soils. Soil Biol. Biochem. 2006, 38, 2882–2889. [Google Scholar] [CrossRef]
- Weyens, N.; Van der Lelie, D.; Taghavi, S.; Newman, L.; Vangronsveld, J. Exploiting plant–microbe partnerships to improve biomass production and remediation. Trends Biotechnol. 2009, 27, 591–598. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.K.; Tissue, D.T.; Thomas, R.B.; Strain, B.R. Comparative responses of model C3 and C4 plants to drought in low and elevated CO2. Glob. Chang. Biol. 1999, 5, 857–867. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Sheng, X.-F.; Xia, J.-J.; Jiang, C.-Y.; He, L.-Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Wuana, R.; Okieimen, F. Phytoremediation potential of maize (Zea mays L.). A review. Afr. J. Gen. Agric. 2010, 6, 275–287. [Google Scholar]
- Javed, S.; Meraj, M.; Bukhari, S.A.; Irfan, R.; Mahmood, S. Hyper-production of alkaline protease by mutagenic treatment of Bacillus subtilis M-9 using agroindustrial wastes in submerged fermentation. J. Microb. Biochem. Technol. 2013, 5, 74–80. [Google Scholar]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Wang, S.Y.; Jiao, H.J.; Faust, M. Changes in ascorbate, glutathione, and related enzyme activities during thidiazuron-induced bud break of apple. Physiol. Plant. 1991, 82, 231–236. [Google Scholar] [CrossRef]
- Ghanti, K.; Rao, S.; Venugopal, R.; Rao, S.; Kaviraj, C.; Barad, A. Somatic embryogenesis and Agrobacterium-mediated transformation in chickpea (Cicer arietinum L.). Plant Cell Biotechnol. Mol. Biol. 2002, 3, 51–58. [Google Scholar]
- Donahue, J.L.; Okpodu, C.M.; Cramer, C.L.; Grabau, E.A.; Alscher, R.G. Responses of antioxidants to paraquat in pea leaves (relationships to resistance). Plant Physiol. 1997, 113, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Peterson, G.L. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- Palta, J.P. Leaf chlorophyll content. Remote Sens. Rev. 1990, 5, 207–213. [Google Scholar] [CrossRef]
- Davis, D.; Gross, E. Protein-protein interactions of the light-harvesting chlorophyll ab protein. II. Evidence for two stages of cation independent association. Biochim. Biophys. Acta (BBA) Bioenerg. 1976, 449, 554–564. [Google Scholar] [CrossRef]
- Javed, S.; Yasin Ashraf, M.; Meraj, M.; Anwer Bukhari, S.; Zovia, I. Salinity and drought induced antioxidant responses in different cultivars of safflower (Carthamus tinctorius L.). Curr. Pharm. Biotechnol. 2013, 14, 814–819. [Google Scholar] [CrossRef]
- Prazeres, C.S.; Coelho, C.M. Osmolyte accumulation and antioxidant metabolism during germination of vigorous maize seeds subjected to water deficit. Acta Sci. Agron. 2020, 42, e42476. [Google Scholar] [CrossRef]
- Mahmood, S.; Afzal, B.; Perveen, S.; Wahid, A.; Azeem, M.; Iqbal, N. He-Ne Laser Seed Treatment Improves the Nutraceutical Metabolic Pool of Sunflowers and Provides Better Tolerance against Water Deficit. Front. Plant Sci. 2021, 17, 804. [Google Scholar]
- Farooq, A.; Bukhari, S.A.; Akram, N.A.; Ashraf, M.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Exogenously applied ascorbic acid-mediated changes in osmoprotection and oxidative defense system enhanced water stress tolerance in different cultivars of safflower (Carthamus tinctorious L.). Plants 2020, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Reddy, C.A.; Saravanan, R.S. Polymicrobial multi-functional approach for enhancement of crop productivity. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 82, pp. 53–113. [Google Scholar]
- Anjum, S.A.; Ashraf, U.; Imran, K.; Tanveer, M.; Shahid, M.; Shakoor, A.; Longchang, W. Phyto-toxicity of chromium in maize: Oxidative damage, osmolyte accumulation, anti-oxidative defense and chromium uptake. Pedosphere 2017, 27, 262–273. [Google Scholar] [CrossRef]
- Ganesh, K.S.; Baskaran, L.; Rajasekaran, S.; Sumathi, K.; Chidambaram, A.; Sundaramoorthy, P. Chromium stress induced alterations in biochemical and enzyme metabolism in aquatic and terrestrial plants. Colloids Surf. B Biointerfaces 2008, 63, 159–163. [Google Scholar] [CrossRef]
- Ovando-Martinez, M.; Sáyago-Ayerdi, S.; Agama-Acevedo, E.; Goñi, I.; Bello-Pérez, L.A. Unripe banana flour as an ingredient to increase the undigestible carbohydrates of pasta. Food Chem. 2009, 113, 121–126. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çiçek, N.; Çakirlar, H. The effect of salinity on some physiological parameters in two maize cultivars. Bulg. J. Plant Physiol. 2002, 28, 66–74. [Google Scholar]
- Singh, V.; Praveen, V.; Khan, F.; Tripathi, C.K.M. Phylogenetics of an antibiotic producing Streptomyces strain isolated from soil. Bioinformation 2009, 4, 5334. [Google Scholar] [CrossRef] [Green Version]
- Augustine, A.; Onwuka, J.; Albert, C. Determination of heavy metal concentration in Neem (Azadirachta indica) leaves, bark and soil along some major roads in Lafia, Nasarawa State Nigeria. J. Environ. Chem. Ecotoxicol. 2016, 8, 38–43. [Google Scholar]
- Shah, A.A.; Yasin, N.A.; Akram, K.; Ahmad, A.; Khan, W.U.; Akram, W.; Akbar, M. Ameliorative role of Bacillus subtilis FBL-10 and silicon against lead induced stress in Solanum melongena. Plant Physiol. Biochem. 2021, 158, 486–496. [Google Scholar] [CrossRef]
- Parida, B.; Chhibba, I.; Nayyar, V. Influence of nickel-contaminated soils on fenugreek (Trigonella corniculata L.) growth and mineral composition. Sci. Hortic. 2003, 98, 113–119. [Google Scholar] [CrossRef]
- Souza, L.R.R.; Bernardes, L.E.; Barbetta, M.F.S.; Da Veiga, M.A.M.S. Iron oxide nanoparticle phytotoxicity to the aquatic plant Lemna minor: Effect on reactive oxygen species (ROS) production and chlorophyll a/chlorophyll b ratio. Environ. Sci. Pollut. Res. 2019, 26, 24121–24131. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Growth on Selective Marker |
|---|---|
| BSU-1 | -ve |
| BSU-2 | -ve |
| BSU-3 | -ve |
| BSU-4 | +ve |
| BSU-5 | -ve |
| BSU-6 | -ve |
| BSU-7 | +ve |
| BSγ-1 | -ve |
| BSγ-2 | -ve |
| BSγ-3 | -ve |
| BSγ-4 | +ve |
| Strains | Cr Concentration (ppm) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | 120 | 150 | 180 | 210 | |
| BSU-1 | +ve | +ve | -ve | -ve | -ve | -ve | -ve | -ve |
| BSU-2 | +ve | +ve | +ve | -ve | -ve | -ve | -ve | -ve |
| BSU-3 | +ve | +ve | -ve | -ve | -ve | -ve | -ve | -ve |
| BSU-4 | +ve | +ve | +ve | -ve | -ve | -ve | -ve | -ve |
| BSU-5 | +ve | +ve | -ve | +ve | -ve | -ve | -ve | -ve |
| BSU-6 | +ve | +ve | -ve | -ve | -ve | -ve | -ve | -ve |
| BSU-7 | +ve | +ve | +ve | +ve | +ve | +ve | +ve | -ve |
| BSγ-1 | +ve | +ve | +ve | -ve | -ve | -ve | -ve | -ve |
| BSγ-2 | +ve | +ve | -ve | -ve | -ve | -ve | -ve | -ve |
| BSγ-3 | +ve | +ve | -ve | -ve | -ve | -ve | -ve | -ve |
| BSγ-4 | +ve | +ve | +ve | +ve | +ve | +ve | +ve | -ve |
| Source of Variation | df | Shoot FW | Shoot DW | Root FW | Root DW | Shoot Length |
|---|---|---|---|---|---|---|
| Stress (S) | 2 | 0.9325 *** | 0.1386 *** | 0.0902 *** | 0.0856 *** | 0.680 *** |
| Treatment (T) | 3 | 113.212 *** | 1.282 *** | 29.10 *** | 1.064 *** | 270.12 *** |
| S × T | 6 | 0.346 *** | 0.720 *** | 0.234 *** | 0.0119 *** | 0.3113 *** |
| Root Length | Proline | MDA | H2O2 | Protein | ||
| Stress (S) | 2 | 0.285 *** | 0.0077 *** | 3.083 *** | 0.0013 *** | 2.586 *** |
| Treatment (T) | 3 | 1150.88 *** | 0.499 *** | 0.3531 *** | 0.3165 *** | 0.105 *** |
| S × T | 6 | 0.547 *** | 1.409 ns | 0.0010 *** | 1.57 ns | 3.861 ns |
| Catalase | APX | GR | POD | SOD | ||
| Stress (S) | 2 | 8.694 ns | 2.194 *** | 0.0017 * | 8.0833 *** | 11.083 *** |
| Treatment (T) | 3 | 193,581.7 *** | 67,589.06 *** | 1078.4 *** | 202,487.74 *** | 98,622.4 *** |
| S × T | 6 | 15.95 ns | 1.3425 *** | 3.416 ns | 0.0555 *** | 0.0833 ns |
| Chl a | Chl a + b | Car | Chla/b | |||
| Stress (S) | 2 | 0.0010 ns | 2.612 *** | 0.0016 *** | 2.61 *** | |
| Treatment (T) | 3 | 68.135 *** | 7.59 *** | 0.0120 *** | 7.62 *** | |
| S × T | 6 | 0.0029 ns | 2.41 *** | 0.0084 *** | 2.391 *** |
| Source of Variation | df | Shoot FW | Shoot DW | Root FW | Root DW | Shoot Length |
|---|---|---|---|---|---|---|
| Stress (S) | 2 | 9.284 *** | 1.425 *** | 0.0033 *** | 0.0350 *** | 0.0099 *** |
| Treatment (T) | 3 | 0.0096 *** | 2.469 *** | 0.0029 *** | 1.0230 *** | 253.78 *** |
| S × T | 6 | 0.0011 *** | 3.491 *** | 0.0012 *** | 0.0092 *** | 0.1922 *** |
| Root Length | Proline | MDA | H2O2 | Protein | ||
| Stress (S) | 2 | 0.4967 *** | 0.0052 *** | 2.860 *** | 0.0015 *** | 5.361 *** |
| Treatment (T) | 3 | 133.16 *** | 1.1732 *** | 1.281 *** | 0.313 *** | 0.141 *** |
| S × T | 6 | 0.2990 *** | 1.111 ns | 3.675 *** | 1.38 ns | 1.484 ns |
| Catalase | APX | GR | POD | SOD | ||
| Stress (S) | 2 | 6.583 *** | 3166.5 *** | 7.583 *** | 6.0277 *** | 6.194 *** |
| Treatment (T) | 3 | 284,498.0 *** | 8134.02 *** | 1411.4 *** | 178,010.19 *** | 178,701.3 *** |
| S × T | 6 | 0.879 *** | 25,551.1 *** | 2.5 ns | 9.435 *** | 2.083 *** |
| Chl a | Chl a + b | Car | Chl a/b | |||
| Stress (S) | 2 | 0.0015 ns | 2.6 *** | 6.194 ** | 2.61 *** | |
| Treatment (T) | 3 | 60.102 *** | 7.63 *** | 0.0350 *** | 7.62 *** | |
| S × T | 6 | 2.0833 ns | 2.4 *** | 0.0025 *** | 2.391 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahzad, Q.; Mahmood, S.; Javed, S.; Mushtaq, T. Chromium Stress Tolerance of a C4 (Zea mays L.) and C3 (Vigna radiata L.) Plants Primed with UV and Gamma-Treated Bacillus subtilis. Microorganisms 2021, 9, 2313. https://doi.org/10.3390/microorganisms9112313
Shahzad Q, Mahmood S, Javed S, Mushtaq T. Chromium Stress Tolerance of a C4 (Zea mays L.) and C3 (Vigna radiata L.) Plants Primed with UV and Gamma-Treated Bacillus subtilis. Microorganisms. 2021; 9(11):2313. https://doi.org/10.3390/microorganisms9112313
Chicago/Turabian StyleShahzad, Qasim, Saqib Mahmood, Sadia Javed, and Tariq Mushtaq. 2021. "Chromium Stress Tolerance of a C4 (Zea mays L.) and C3 (Vigna radiata L.) Plants Primed with UV and Gamma-Treated Bacillus subtilis" Microorganisms 9, no. 11: 2313. https://doi.org/10.3390/microorganisms9112313
APA StyleShahzad, Q., Mahmood, S., Javed, S., & Mushtaq, T. (2021). Chromium Stress Tolerance of a C4 (Zea mays L.) and C3 (Vigna radiata L.) Plants Primed with UV and Gamma-Treated Bacillus subtilis. Microorganisms, 9(11), 2313. https://doi.org/10.3390/microorganisms9112313