
Impact of Nutrients on Protozoa Community Diversity and Structure in Litter of Two Natural Grass Species in a Copper Tailings Dam, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Litter Sampling
2.2. Litter Sample Collection
2.3. Chemical Properties and Enzyme Activities in Litter
2.4. DNA Extraction Using High-Throughput Sequencing
2.5. Sequence Processing and Taxonomic Classification
2.6. Statistical Analysis
3. Results
3.1. Physical and Chemical Properties of Natural Grass Litter
3.2. Community Diversity of Litter Protozoa
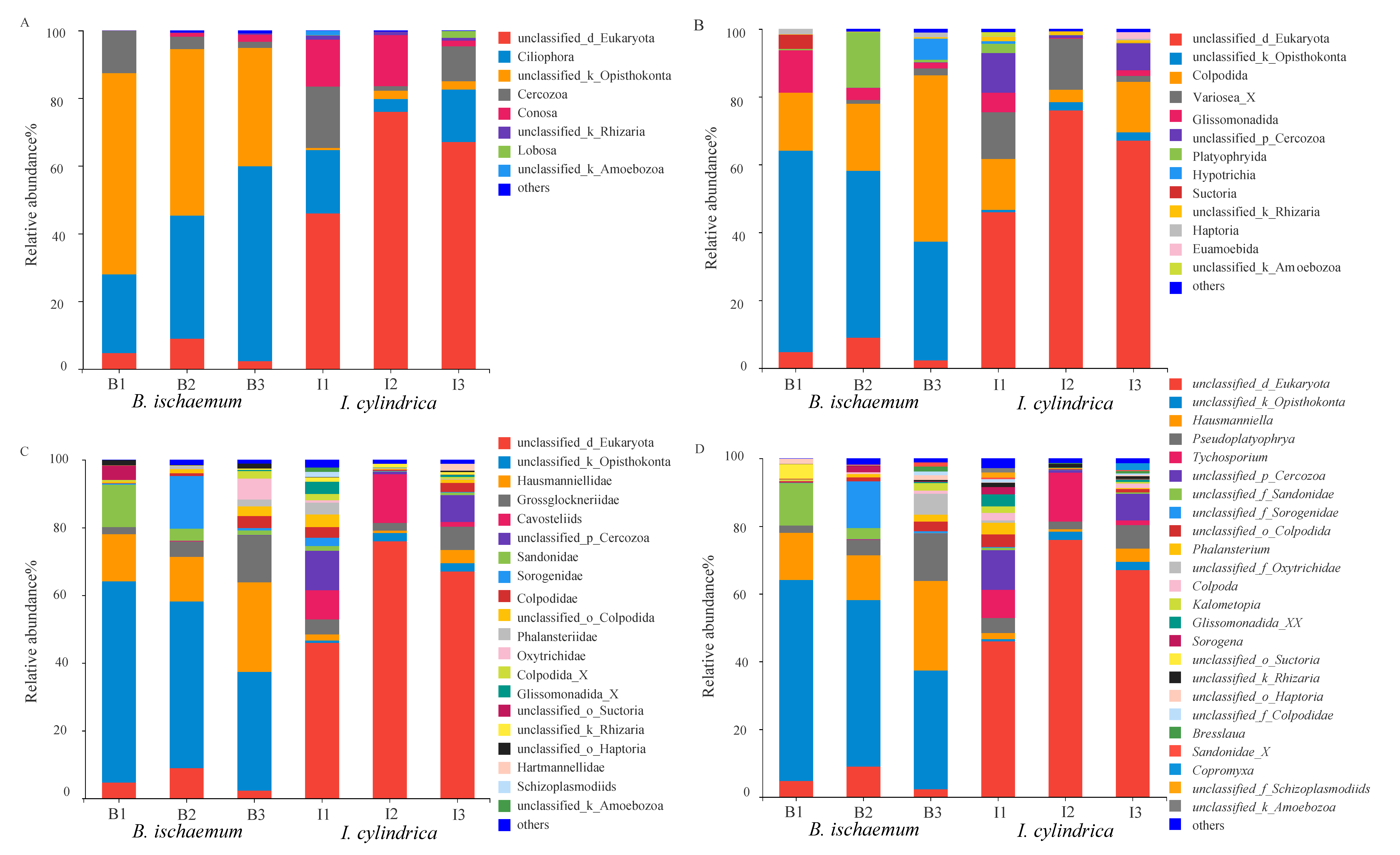
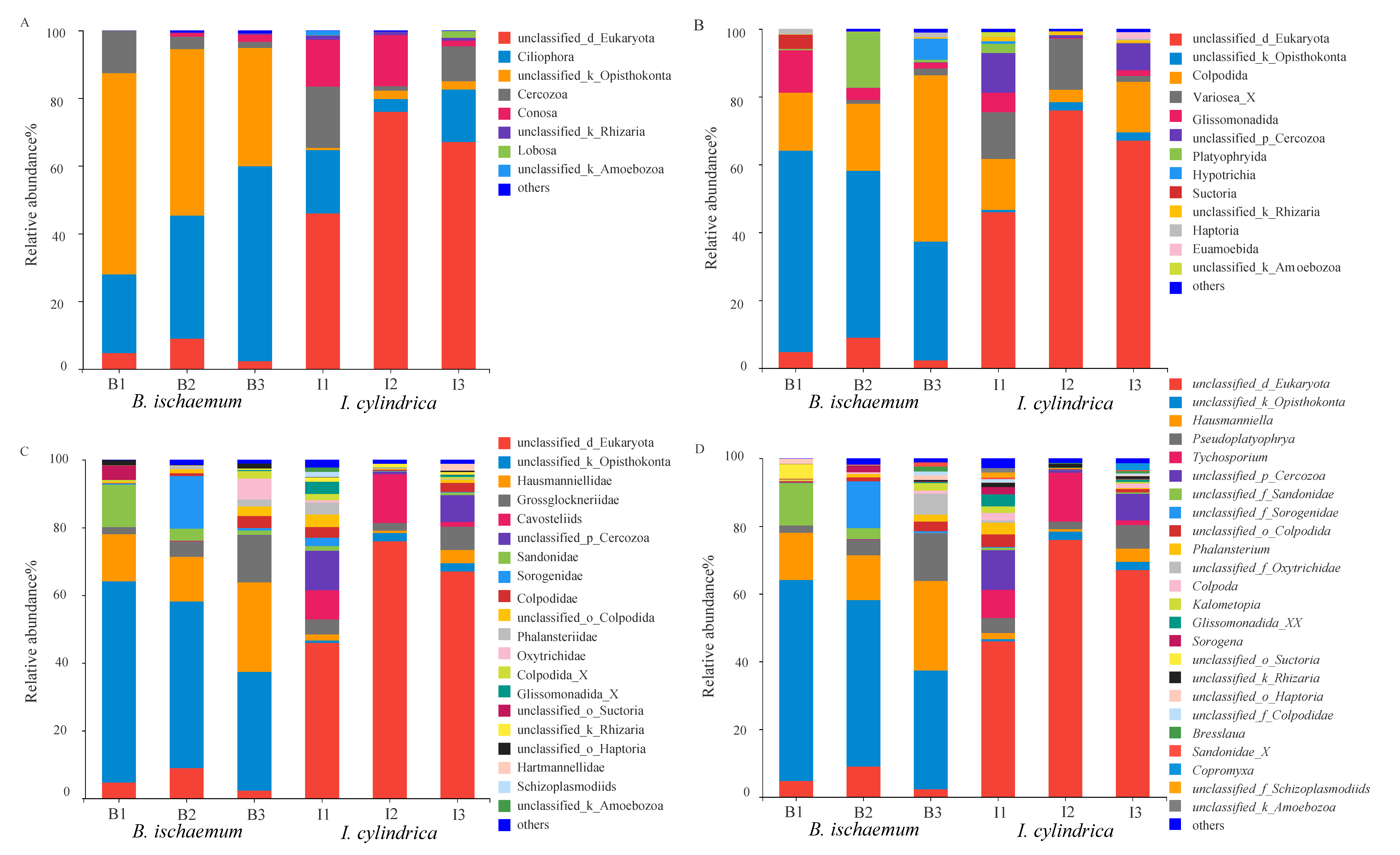
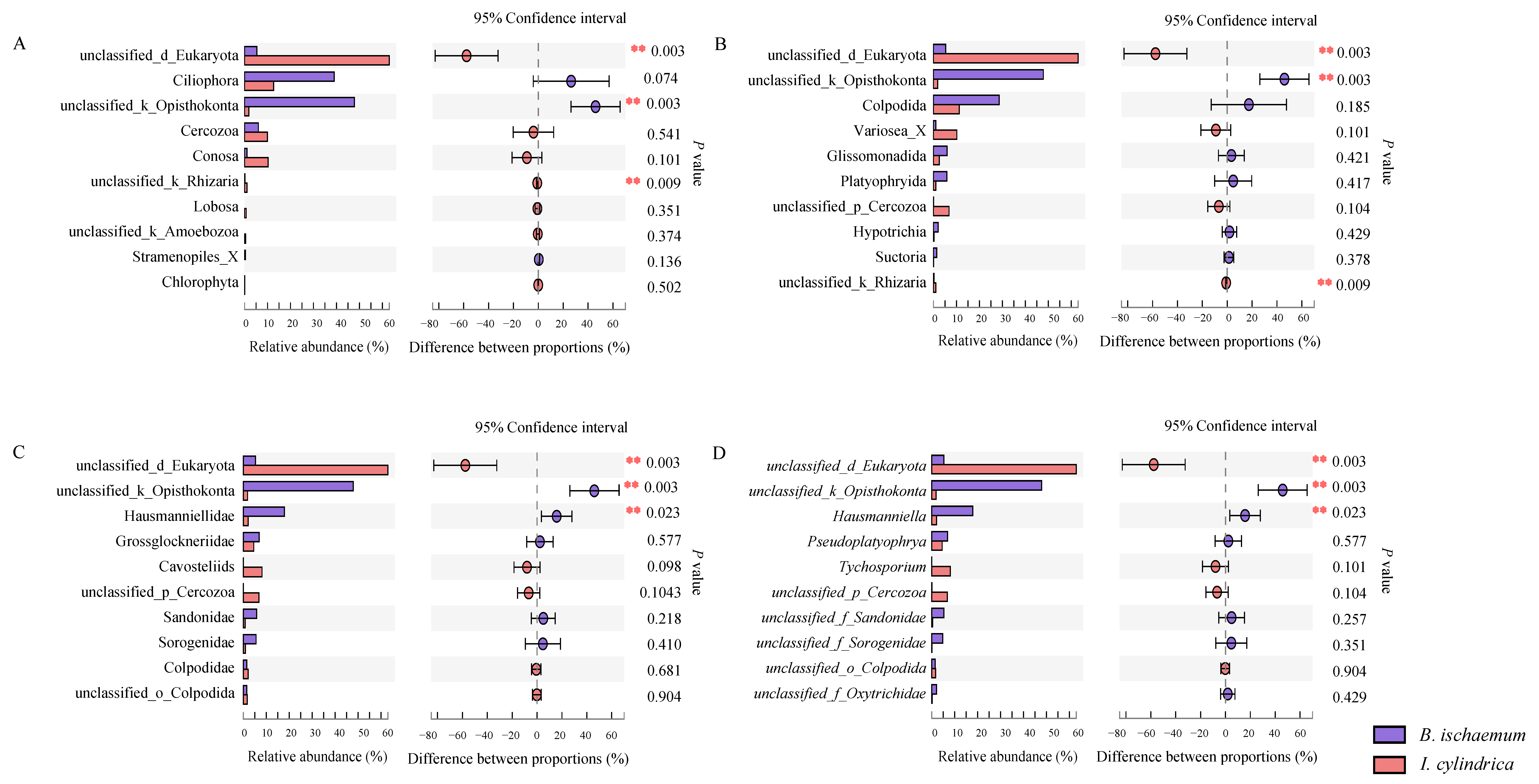
3.3. Community Composition of Litter Protozoa
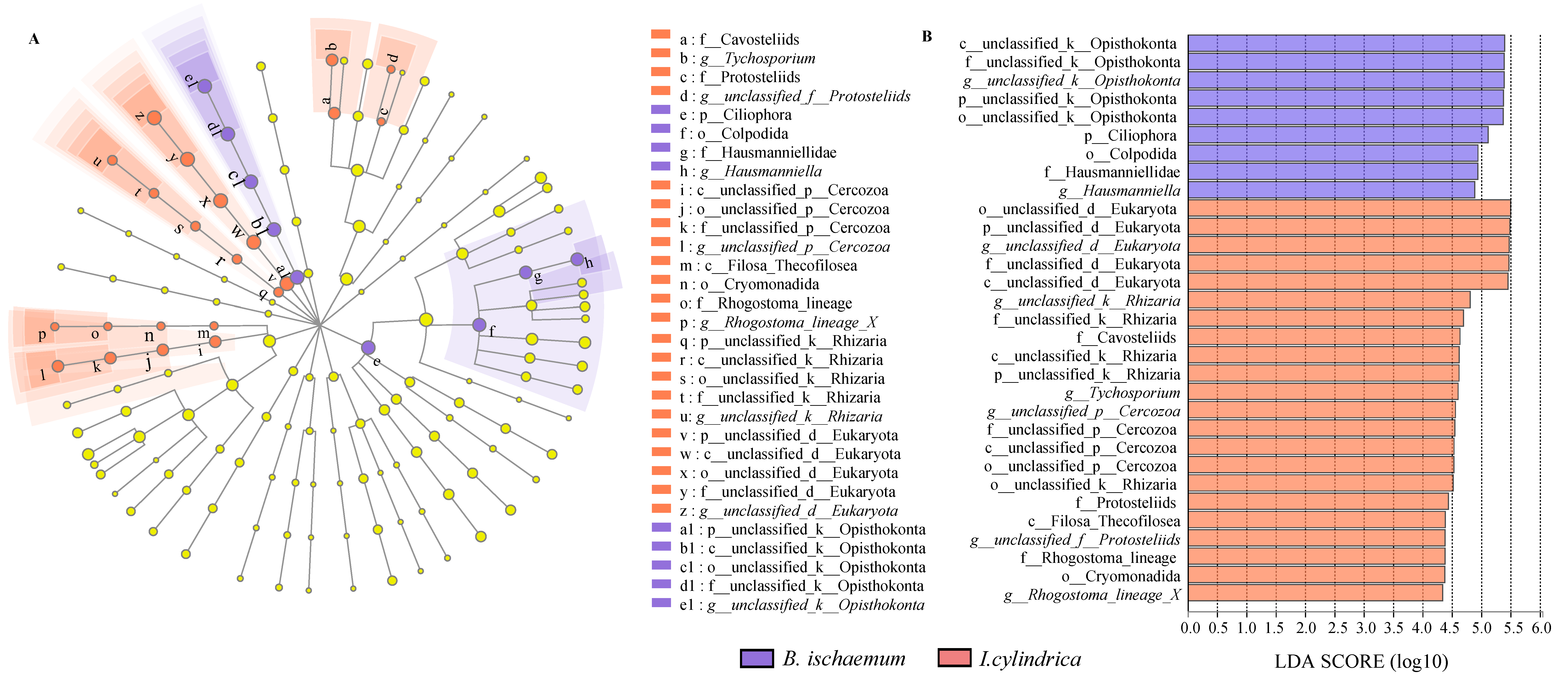
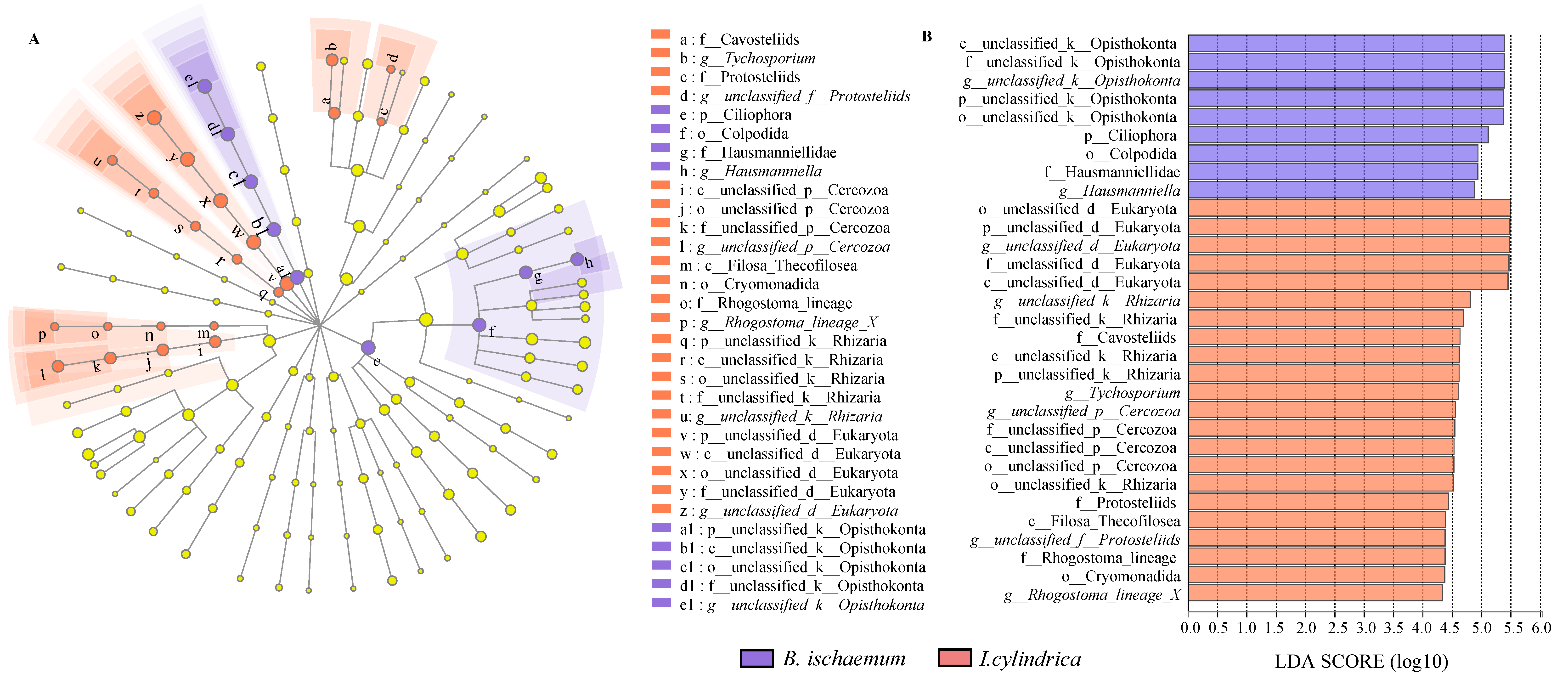
3.4. Difference Analysis of Litter Protozoa Community
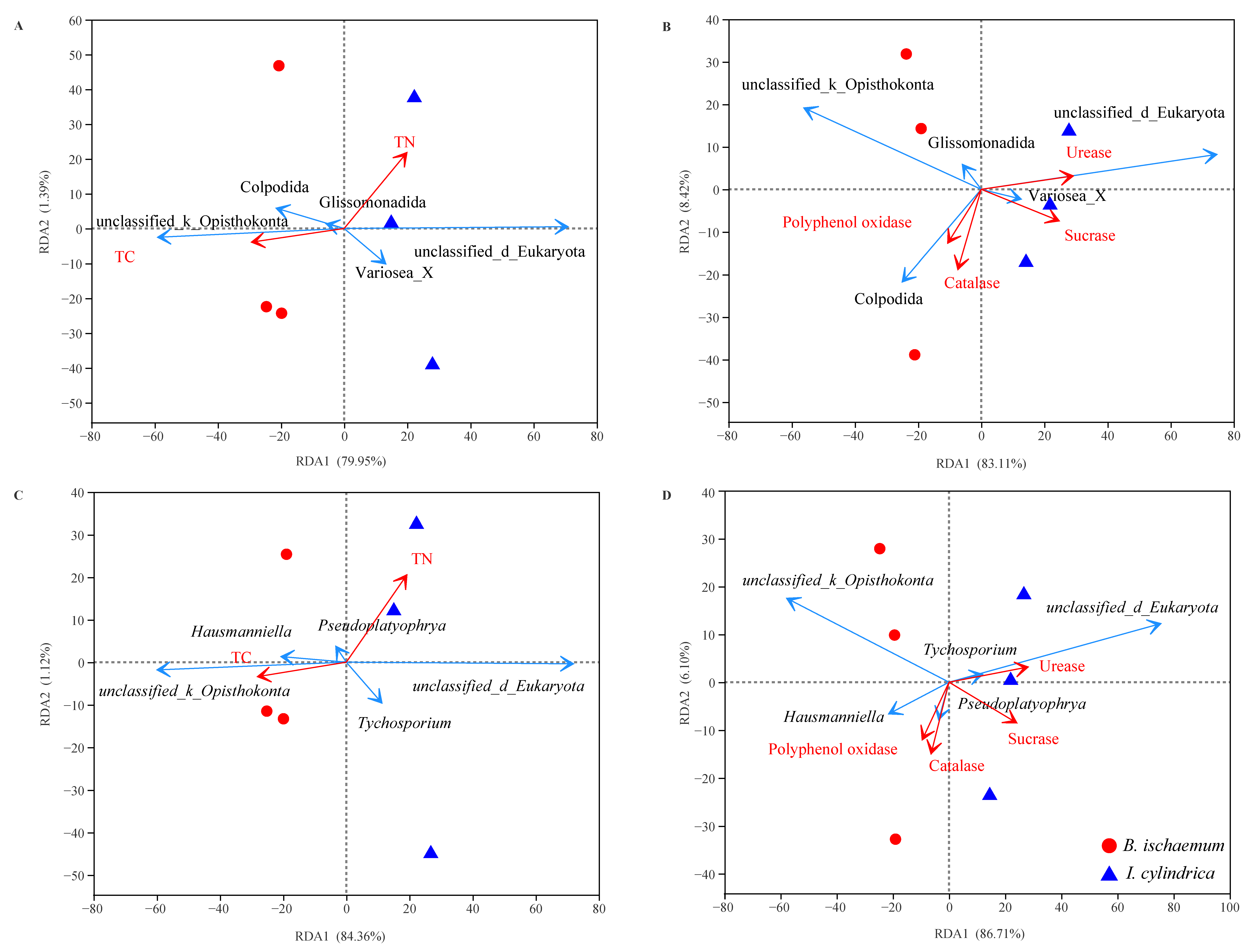
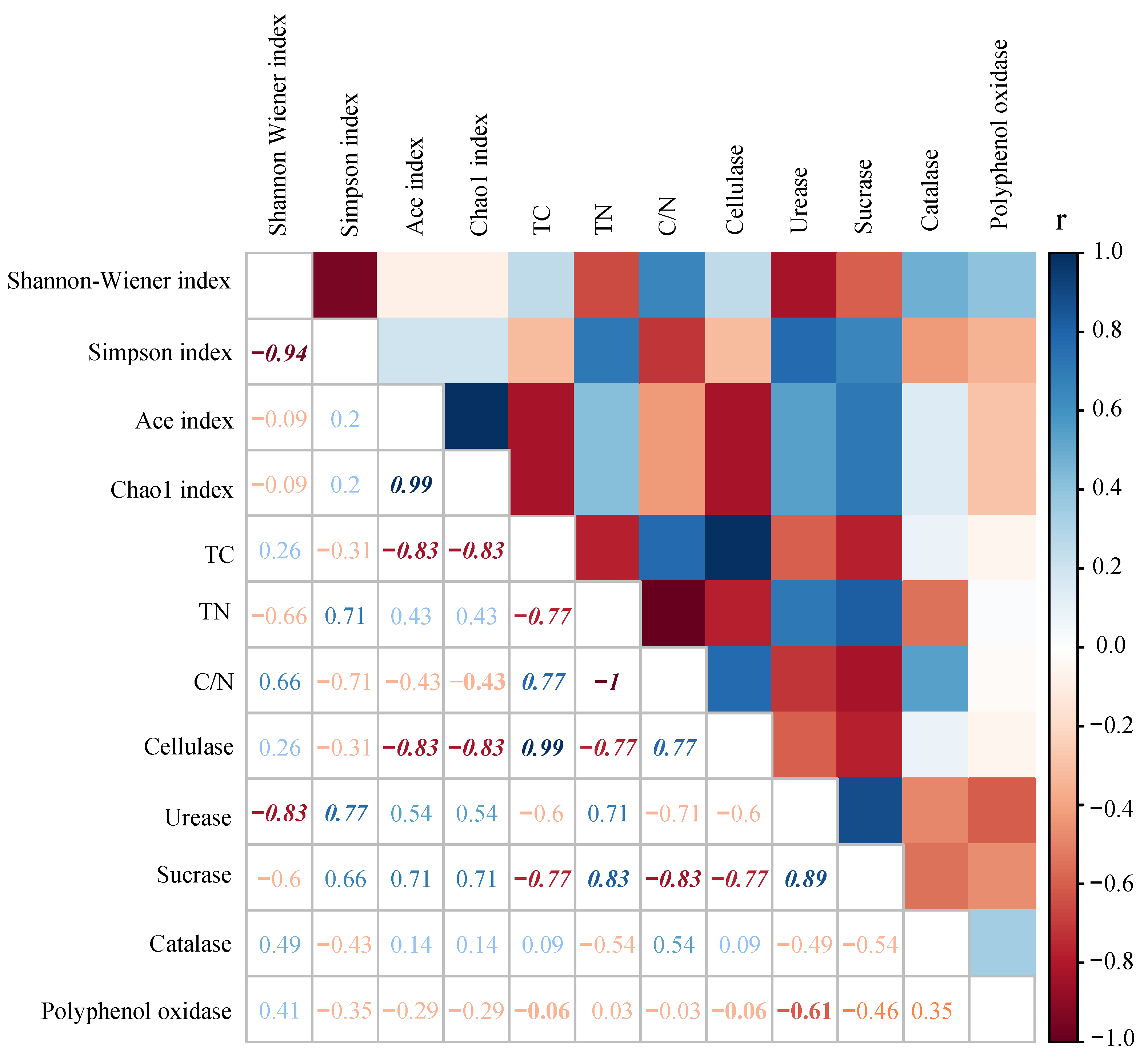
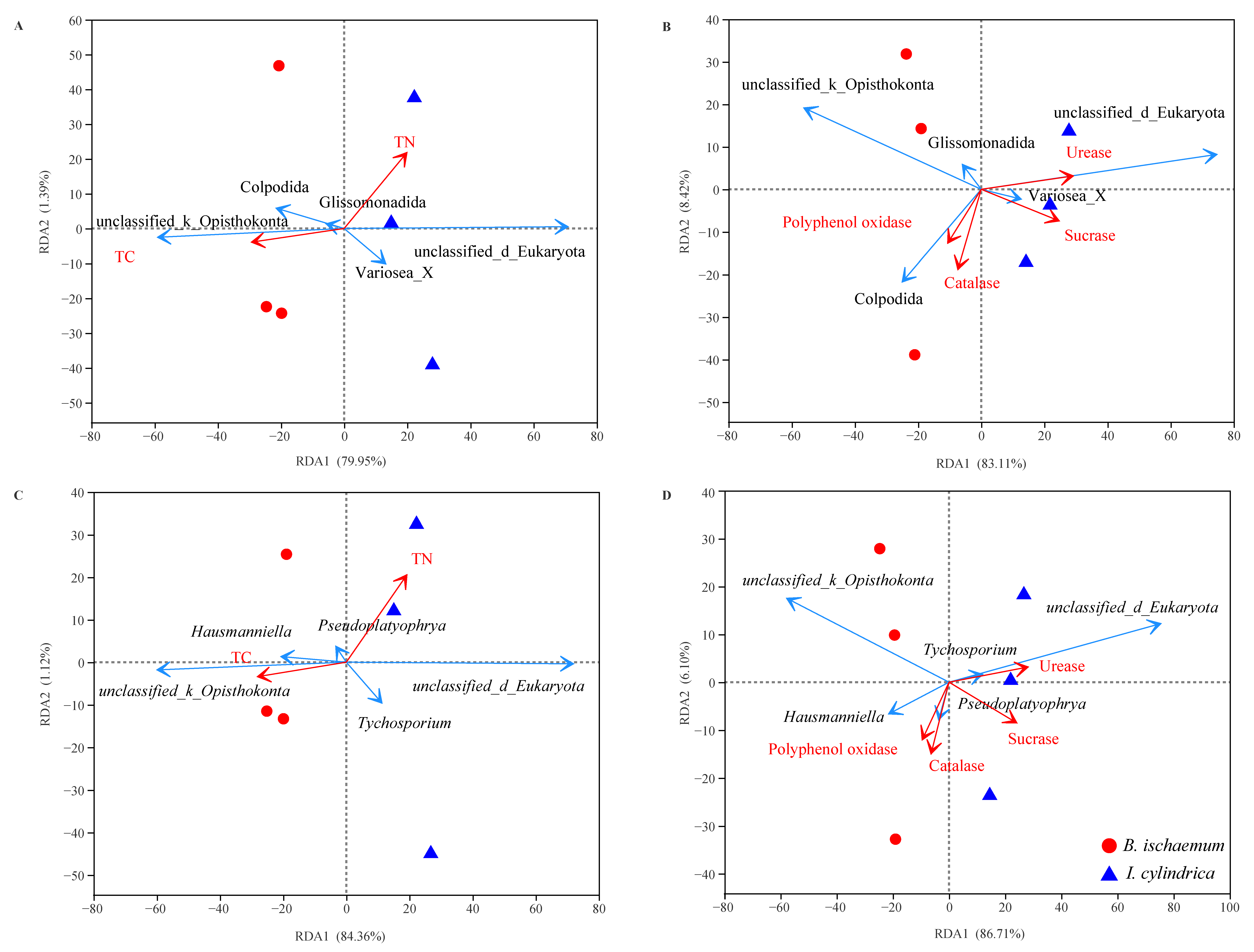
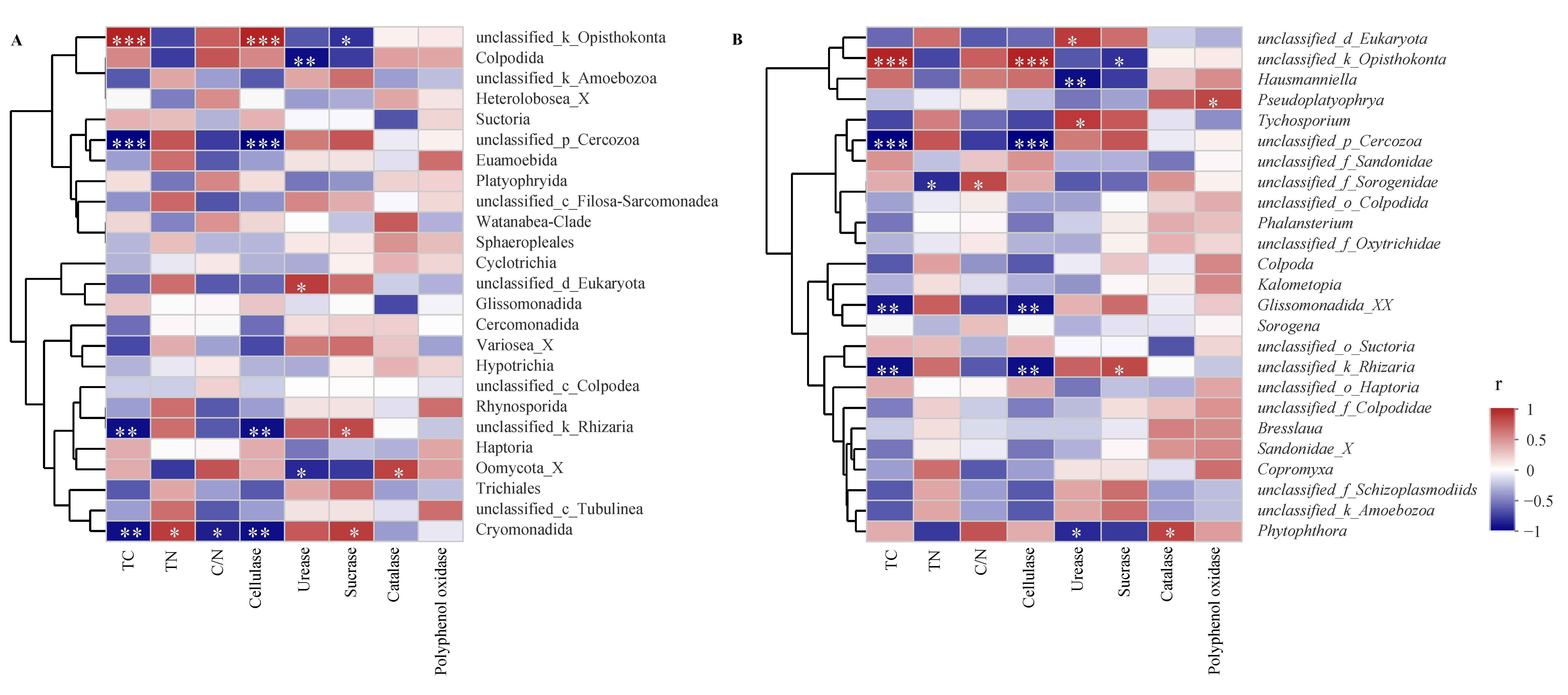
3.5. Litter Characteristic Effects on Protozoan Communities
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naldini, M.B.; Pérez Harguindeguy, N.; Kowaljow, E. Soil carbon release enhanced by increased litter input in a degraded semi-arid forest soil. J. Arid Environ. 2021, 186, 104400. [Google Scholar] [CrossRef]
- Wang, X.P.; Yang, X.; Yang, N.; Xin, X.J.; Qu, Y.B.; Zhao, N.X.; Gao, Y.B. Effects of litters diversity and composition an litter decomposition characteristics and soil microbial community II: Under the conditions of doubling ambient atmospheric CO2 concentration. Acta Ecol. Sin. 2020, 39, 6264–6272. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.W.; Yang, K.; Lyu, Z.T.; Zhu, J.J. Microbial groups and their functions control the decomposition of coniferous litter: A comparison with broadleaved tree litters. Soil Biol. Biochem. 2019, 133, 196–207. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Xu, M.P.; Lu, X.Q.; Xu, Y.D.; Zhong, Z.K.; Zhang, W.; Ren, C.J.; Han, X.H.; Yang, G.H.; Feng, Y.Z. Dynamics of bacterial community in litter and soil along a chronosequence of Robinia pseudoacacia plantations. Sci. Total. Environ. 2020, 703, 135613. [Google Scholar] [CrossRef]
- Geisen, S.; Mitchell, E.A.D.; Adl, S.M.; Bonkowski, M.; Dunthorn, M.; Ekelund, F.; Fernandez, L.D.; Jousset, A.; Krashevska, V.; Singer, D. Soil protists: A fertile frontier in soil biology research. FEMS Microbiol. Rev. 2018, 42, 293–323. [Google Scholar] [CrossRef] [PubMed]
- Shmakova, L.; Bondarenko, N.; Smirnov, A.V. Viable Species of Flamella (Amoebozoa: Variosea) Isolated from Ancient Arctic Permafrost Sediments. Protist 2016, 167, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Geisen, S.; Bandow, C.; Rombke, J.; Bonkowski, M. Soil water availability strongly alters the community composition of soil protists. Pedobiologia 2014, 57, 205–213. [Google Scholar]
- Wei, Z.; Song, Y.Q.; Xiong, W.; Xu, Y.C.; Shen, Q.R. Soil protozoa: Research methods and roles in the biocontrol of soil-borne diseases. Acta Pedol. Sin. 2021, 58, 1–10. [Google Scholar]
- Schmidt, O.; Dyckmans, J.; Schrader, S. Photoautotrophic microorganisms as a carbon source for temperate soil invertebrates. Biol. Lett. 2016, 12, 20150646. [Google Scholar] [CrossRef]
- Clarholm, M. Interactions of bacteria, protozoa and plants leading to mineralization of soil nitrogen. Soil Biol. Biochem. 1985, 17, 181–187. [Google Scholar] [CrossRef]
- Geisen, S. The bacterial-fungal energy channel concept challenged by enormous functional versatility of soil protists. Soil Biol. Biochem. 2016, 102, 22–25. [Google Scholar] [CrossRef]
- Geisen, S.; Koller, R.; Hünninghaus, M.; Dumack, K.; Urich, T.; Bonkowski, M. The soil food web revisited: Diverse and widespread mycophagous soil protists. Soil Biol. Biochem. 2016, 94, 10–18. [Google Scholar] [CrossRef]
- Long, J.J.; Jahn, C.E.; Sanchezhidalgo, A.; Wheat, W.H.; Jackson, M.; Gonzalezjuarrero, M.; Leach, J.E. Interactions of free-living amoebae with rice bacterial pathogens Xanthomonas oryzae pathovars oryzae and oryzicola. PLoS ONE 2018, 13, e0202941. [Google Scholar] [CrossRef] [Green Version]
- Geisen, S.; Fioredonno, A.M.; Walochnik, J.; Bonkowski, M. Acanthamoeba everywhere: High diversity of Acanthamoeba in soils. Parasitol. Res. 2014, 113, 3151–3158. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Milbau, A.; Beyens, L. Environmental factors influencing soil testate amoebae in herbaceous and shrubby vegetation along an altitudinal gradient in subarctic tundra (Abisko.; Sweden). Eur. J. Protistol. 2013, 49, 238–248. [Google Scholar] [CrossRef]
- Liu, Q.S.; Yan, S.Z.; Chen, S.L. Species diversity of myxomycetes associated with different terrestrial ecosystems, substrata (microhabitats) and environmental factors. Mycol. Prog. 2015, 14, 27. [Google Scholar] [CrossRef]
- Zhu, W.T.; Qin, C.X.; Ma, H.M.; Xi, S.G.; Zuo, T.; Pan, W.N.; Li, C.H. Response of protist community dynamics and co-occurrence patterns to the construction of artificial reefs: A case study in Daya Bay, China. Sci. Total. Environ. 2020, 742, 140575. [Google Scholar] [CrossRef]
- Acostamercado, D.; Lynn, D.H. Soil ciliate species richness and abundance associated with the rhizosphere of different subtropical plant species. J. Eukaryot. Microbiol. 2004, 51, 582–588. [Google Scholar] [CrossRef]
- Bischoff, P.J.; Connington, K. Winter abundances of naked amoebae in the soil system of the invasive species Japanese knotweed (Fallopia japonica) with comparative data from adjacent sites. Acta Protozool. 2016, 155–160. [Google Scholar]
- Dassen, S.; Cortois, R.; Martens, H.; De Hollander, M.; Kowalchuk, G.A.; Der Putten, W.H.V.; De Deyn, G.B. Differential responses of soil bacteria, fungi, archaea and protists to plant species richness and plant functional group identity. Mol. Ecol. 2017, 26, 4085–4098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, M.; Wetzel, C.E.; Ector, L.; Teuling, A.J.; Pfister, L. On the potential for terrestrial diatom communities and diatom indices to identify anthropic disturbance in soils. Ecol. Indic. 2017, 75, 73–81. [Google Scholar] [CrossRef]
- Liu, Y. The Influence of Photovoltaic Power Station on Two Typical Degraded Ecosystems in North of Shanxi; Shanxi University of Finance & Economics: Taiyuan, China, 2020. [Google Scholar]
- Ma, Z.X.; Gong, D.J.; Ning, Y.Z.; Niu, S.Q.; Liu, Z.J.; Cui, Y.D. Studies on the effect of soil pollution by waste material of lead and zinc ore mining an protozoa. J. Gansu Sci. 2002, 14, 53–57. [Google Scholar]
- Jing, J.H.; Liu, J.X.; Li, C.; Jia, T.; Wang, X.Y.; Chai, B.F. The structural characteristics of a soil bacterial community in a dam of copper mine tailings in Zhongtiaoshan mountains, Shanxi. Chin. J. Appl. Environ. Biol. 2017, 23, 527–534. [Google Scholar]
- Jia, T.; Cao, M.W.; Jing, J.H.; Liu, J.X.; Chai, B.F. Endophytic fungi and soil microbial community characteristics over different years of phytoremediation in a copper tailings dam of Shanxi, China. Sci. Total. Environ. 2017, 574, 881–888. [Google Scholar]
- Stoeck, T.; Breiner, H.; Filker, S.; Ostermaier, V.; Kammerlander, B.; Sonntag, B. A morphogenetic survey on ciliate plankton from a mountain lake pinpoints the necessity of lineage-specific barcode markers in microbial ecology. Environ. Microbiol. 2014, 16, 430–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, W.; Li, R.; Guo, S.; Karlsson, I.; Jiao, Z.X.; Xun, W.B.; Kowalchuk, G.A.; Shen, Q.R.; Geisen, S. Microbial amendments alter protist communities within the soil microbiome. Soil Biol. Biochem. 2019, 135, 379–382. [Google Scholar] [CrossRef]
- Santos, S.S.; Schöler, A.; Nielsen, T.K.; Hansen, L.H.; Schloter, M.; Winding, A. Land use as a driver for protist community structure in soils under agricultural use across Europe. Sci. Total. Environ. 2020, 717, 137228. [Google Scholar] [CrossRef]
- Dumack, K.; Flues, S.; Hermanns, K.; Bonkowski, M. Rhogostomidae (Cercozoa) from soils, roots and plant leaves (Arabidopsis thaliana): Description of Rhogostoma epiphylla sp. nov. and R. cylindrica sp. nov. Eur. J. Protistol. 2017, 60, 76–86. [Google Scholar] [CrossRef]
- Flues, S.; Bass, D.; Bonkowski, M. Grazing of leaf-associated Cercomonads (Protists: Rhizaria: Cercozoa) structures bacterial community composition and function. Environ. Microbiol. 2017, 19, 3297–3309. [Google Scholar] [CrossRef]
- Ning, Y.Z.; Wanf, F.G.; Du, H.F.; Wang, H.J.; Su, C. Ciliate species diversity and its relationships with environmental factors in plateau swamp wetlands of southern Gansu Province, Northwest China in autumn. Chin. J. Ecol. 2013, 32, 634–640. [Google Scholar]
- Ning, Y.Z.; Wang, X.J.; Li, Q.L.; Yu, J.H.; Ma, Z.X. Community characteristics of soil ciliates in the national forest park of the Taibai mountains. J. Northwest Norm. Univ. 2009, 45, 82–88. [Google Scholar]
- Niu, S.Q.; Ning, Y.Z.; Ma, Z.X.; Gong, D.J.; Liu, Z.J.; Xu, Y.G. Studies on the community characteristics of protozoa in the soil witj compoind pollution of heavy metals. J. Gansu Sci. 2002, 14, 44–48. [Google Scholar] [CrossRef]
- Johansen, J.L.; Ronn, R.; Ekelund, F. Toxicity of cadmium and zinc to small soil protists. Environ. Pollut. 2018, 242, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X. Pollution Ecology; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Spiegel, F.W.; Moore, D.L.; Feldman, J. Tychosporium acutostipes, a new protostelid which modifies the concept of the protosteliidae. Mycologia 1995, 87, 265–270. [Google Scholar] [CrossRef]
- Seppey, C.V.W.; Singer, D.; Dumack, K.; Fournier, B.; Belbahri, L.; Mitchell, E.A.D.; Lara, E. Distribution patterns of soil microbial eukaryotes suggests widespread algivory by phagotrophic protists as an alternative pathway for nutrient cycling. Soil Biol. Biochem. 2017, 112, 68–76. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TC (%) | TN (%) | C/N | Cellulase (mg·(g·72 h)−1) | Urease (mg·(g·24 h)−1) | Sucrase (mg·(g·24 h)−1) | Catalase (mg·(g·20 min)−1) | Polyphenol Oxidase (mL·g−1) | |
|---|---|---|---|---|---|---|---|---|
| B. ischaemum | 44.442 ± 0.057a | 0.468 ± 0.007 | 95.000 ± 1.512 | 1.200 ± 0.077a | 0.969 ± 0.478b | 1.395 ± 0.047 | 3.607 ± 0.866 | 5.867 ± 0.321 |
| I. cylindrica | 43.157 ± 0.113b | 0.583 ± 0.097 | 75.377 ± 12.721 | 0.772 ± 0.054b | 3.760 ± 0.308a | 3.072 ± 0.716 | 3.237 ± 0.231 | 5.500 ± 1.000 |
| Coverage | Shannon–Wiener Index | Simpson Index | Chao1 Index | ACE Index | |
|---|---|---|---|---|---|
| B. ischaemum | 0.997 ± 0.001 | 2.318 ± 0.454 | 0.178 ± 0.097 | 47.083 ± 15.261 | 48.467 ± 15.984 |
| I. cylindrica | 0.995 ± 0.001 | 1.733 ± 0.602 | 0.414 ± 0.178 | 66.917 ± 14.036 | 66.763 ± 10.907 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, T.; Liang, X.; Guo, T.; Chai, B. Impact of Nutrients on Protozoa Community Diversity and Structure in Litter of Two Natural Grass Species in a Copper Tailings Dam, China. Microorganisms 2021, 9, 2250. https://doi.org/10.3390/microorganisms9112250
Jia T, Liang X, Guo T, Chai B. Impact of Nutrients on Protozoa Community Diversity and Structure in Litter of Two Natural Grass Species in a Copper Tailings Dam, China. Microorganisms. 2021; 9(11):2250. https://doi.org/10.3390/microorganisms9112250
Chicago/Turabian StyleJia, Tong, Xiaoxia Liang, Tingyan Guo, and Baofeng Chai. 2021. "Impact of Nutrients on Protozoa Community Diversity and Structure in Litter of Two Natural Grass Species in a Copper Tailings Dam, China" Microorganisms 9, no. 11: 2250. https://doi.org/10.3390/microorganisms9112250
APA StyleJia, T., Liang, X., Guo, T., & Chai, B. (2021). Impact of Nutrients on Protozoa Community Diversity and Structure in Litter of Two Natural Grass Species in a Copper Tailings Dam, China. Microorganisms, 9(11), 2250. https://doi.org/10.3390/microorganisms9112250