Salmonella Virulence and Immune Escape




Abstract
1. Introduction
2. Origin, Classification, and Diseases Caused by Salmonella
3. The Virulence-Related Genes of SPI
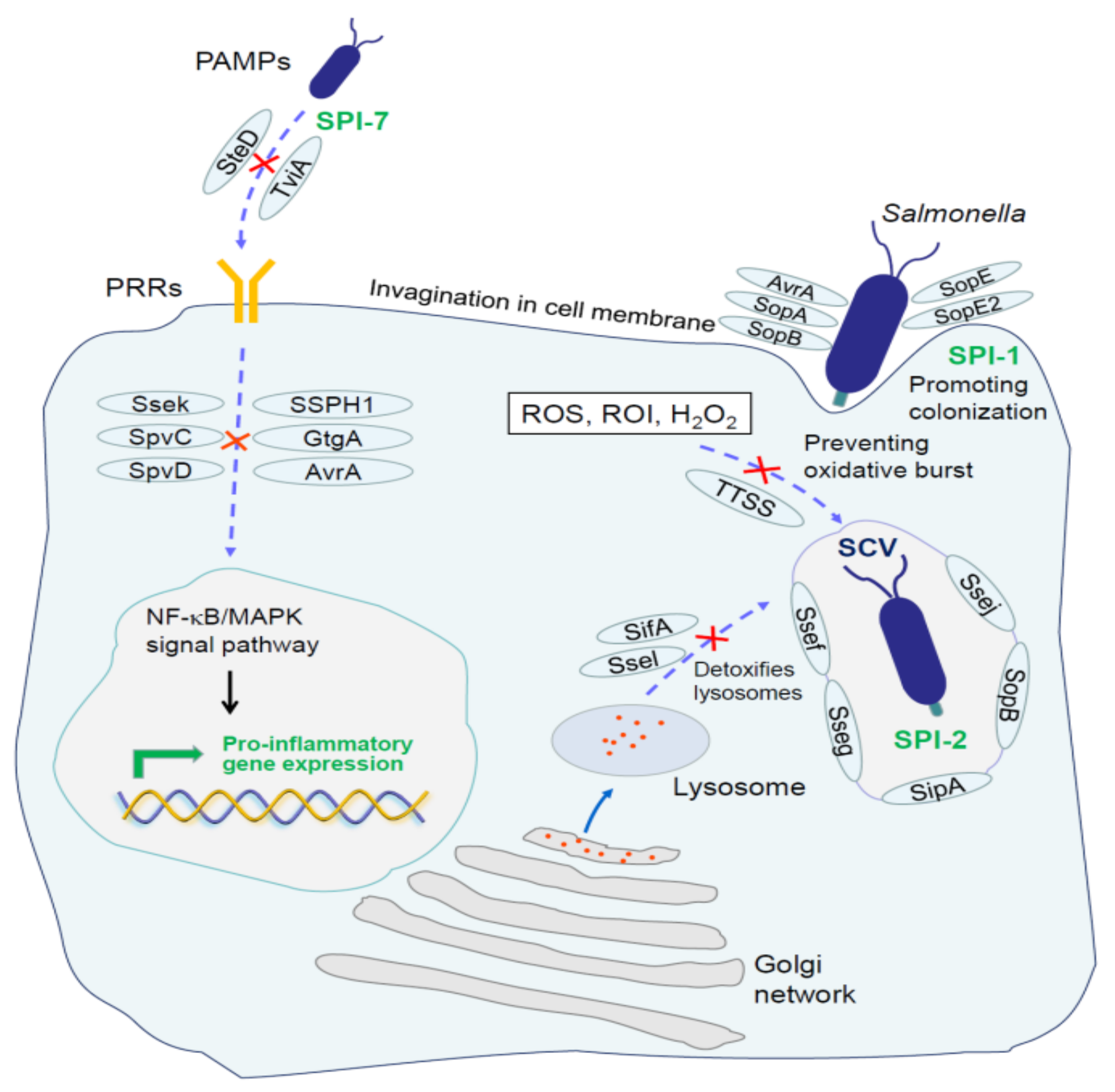
4. Molecular Mechanisms of Salmonella Immune Escape
4.1. Escape of Innate Immune System
4.2. Escape of Adaptive Immune Responses
5. Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- De Abrew Abeysundara, P.; Dhowlaghar, N.; Nannapaneni, R.; Schilling, M.W.; Mahmoud, B.; Sharma, C.S.; Ma, D.P. Salmonella enterica growth and biofilm formation in flesh and peel cantaloupe extracts on four food-contact surfaces. Int. J. Food Microbiol. 2018, 280, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Tadepalli, S.; Bridges, D.F.; Driver, R.; Wu, V.C.H. Effectiveness of different antimicrobial washes combined with freezing against Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes inoculated on blueberries. Food Microbiol. 2018, 74, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Al-Holy, M.A.; Abu Ghoush, M.; Al-Nabulsi, A.A.; Holley, R.A. Control of Salmonella enterica and Listeria monocytogenes in hummus using allyl isothiocyanate. Int. J. Food Microbiol. 2018, 278, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Parry-Hanson Kunadu, A.; Holmes, M.; Miller, E.L.; Grant, A.J. Microbiological quality and antimicrobial resistance characterization of Salmonella spp. in fresh milk value chains in Ghana. Int. J. Food Microbiol. 2018, 277, 41–49. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef]
- Wain, J.; Hendriksen, R.S.; Mikoleit, M.L.; Keddy, K.H.; Ochiai, R.L. Typhoid fever. Lancet 2015, 385, 1136–1145. [Google Scholar] [CrossRef]
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the host and disease: A brief review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Im, J.; Lee, J.-S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R. The global burden and epidemiology of invasive non-typhoidal infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Kingsley, R.A.; Townsend, S.M.; Ficht, T.A.; Adams, L.G.; Baumler, A.J. Of mice, calves, and men. Comparison of the mouse typhoid model with other Salmonella infections. Adv. Exp. Med. Biol. 1999, 473, 261–274. [Google Scholar]
- Adams, D.A.; Thomas, K.R.; Jajosky, R.A.; Foster, L.; Sharp, P.; Onweh, D.H.; Schley, A.W. Summary of Notifiable Infectious Diseases and Conditions—United States, 2014. Morb. Mortal. Wkly. Rep. 2016, 63, 1–152. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L. Clinical practice. Bacterial diarrhea. N. Engl. J. Med. 2009, 361, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Parisi, A.; Sarkar, K.; Blacker, B.F.; Reiner, R.C.; Hay, S.I.; Nixon, M.R.; Dolecek, C.; James, S.L.; Mokdad, A.H.; et al. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Dhowlaghar, N.; Bansal, M.; Schilling, M.W.; Nannapaneni, R. Scanning electron microscopy of Salmonella biofilms on various food-contact surfaces in catfish mucus. Food Microbiol. 2018, 74, 143–150. [Google Scholar] [CrossRef]
- Marin, C.; Torres, C.; Marco-Jimenez, F.; Cerda-Cuellar, M.; Sevilla, S.; Ayats, T.; Vega, S. Supplementary feeding stations for conservation of vultures could be an important source of monophasic Salmonella typhimurium 1,4,[5],12:i. Sci. Total Environ. 2018, 636, 449–455. [Google Scholar] [CrossRef]
- Spanò, S. Mechanisms of Salmonella Typhi Host Restriction. Adv. Exp. Med. Biol. 2016, 915, 283–294. [Google Scholar]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive virulence factors and pathogenesis. Cell. Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef]
- Feng, Y.; Lin, E.; Zou, S.; Chen, C.-L.; Chiu, C.-H. Complete genome sequence of Salmonella enterica serovar Sendai shows H antigen convergence with S. Miami and recent divergence from S. Paratyphi A. BMC Genom. 2019, 20, 398. [Google Scholar] [CrossRef]
- Ly, K.T.; Casanova, J.E. Mechanisms of Salmonella entry into host cells. Cell. Microbiol. 2007, 9, 2103–2111. [Google Scholar] [CrossRef]
- Haghjoo, E.; Galán, J.E. Salmonella typhi encodes a functional cytolethal distending toxin that is delivered into host cells by a bacterial-internalization pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 4614–4619. [Google Scholar] [CrossRef]
- Liston, S.D.; Ovchinnikova, O.G.; Whitfield, C. Unique lipid anchor attaches Vi antigen capsule to the surface of Salmonella enterica serovar Typhi. Proc. Natl. Acad. Sci. USA 2016, 113, 6719–6724. [Google Scholar] [CrossRef] [PubMed]
- Jepson, M.A.; Clark, M.A. The role of M cells in Salmonella infection. Microbes Infect. 2001, 3, 1183–1190. [Google Scholar] [CrossRef]
- Jones, B.D.; Falkow, S. Salmonellosis: Host immune responses and bacterial virulence determinants. Annu. Rev. Immunol. 1996, 14, 533–561. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Fang, F.C. Cellular routes of invasion by enteropathogens. Curr. Opin. Microbiol. 2000, 3, 54–59. [Google Scholar] [CrossRef]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, D.K.; Wong, V.K. Emergence of an Extensively Drug-Resistant Serovar Typhi Clone Harboring a Promiscuous Plasmid Encoding Resistance to Fluoroquinolones and Third-Generation Cephalosporins. MBio 2018, 9. [Google Scholar] [CrossRef]
- Holt, K.E.; Phan, M.D.; Baker, S.; Duy, P.T.; Nga, T.V.T.; Nair, S.; Turner, A.K. Emergence of a globally dominant IncHI1 plasmid type associated with multiple drug resistant typhoid. PLoS Negl. Trop. Dis. 2011, 5, e1245. [Google Scholar] [CrossRef]
- Wotzka, S.Y.; Nguyen, B.D.; Hardt, W.D. Salmonella Typhimurium Diarrhea Reveals Basic Principles of Enteropathogen Infection and Disease-Promoted DNA Exchange. Cell Host Microbe 2017, 21, 443–454. [Google Scholar] [CrossRef]
- Bumann, D.; Schothorst, J. Intracellular Salmonella metabolism. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef]
- Brumell, J.H.; Rosenberger, C.M.; Gotto, G.T.; Marcus, S.L.; Finlay, B.B. SifA permits survival and replication of Salmonella typhimurium in murine macrophages. Cell. Microbiol. 2001, 3, 75–84. [Google Scholar] [CrossRef]
- Wei, S.; Huang, J.; Liu, Z.; Wang, M.; Zhang, B.; Lian, Z.; Guo, Y. Differential immune responses of C57BL/6 mice to infection by Salmonella enterica serovar Typhimurium strain SL1344, CVCC541 and CMCC50115. Virulence 2019, 10, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, S.C.; Forest, C.G.; Lepage, C.; Leclerc, J.-M.; Daigle, F. So similar, yet so different: Uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol. Lett. 2010, 305, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Carden, S.E.; Walker, G.T.; Honeycutt, J.; Lugo, K.; Pham, T.; Jacobson, A.; Bouley, D. Pseudogenization of the Secreted Effector Gene sseI Confers Rapid Systemic Dissemination of S. Typhimurium ST313 within Migratory Dendritic Cells. Cell Host Microbe 2017, 21, 182–194. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. MBio 2019, 10. [Google Scholar] [CrossRef]
- Ahmed, H.A.; El-Hofy, F.I.; Shafik, S.M.; Abdelrahman, M.A.; Elsaid, G.A. Characterization of Virulence-Associated Genes, Antimicrobial Resistance Genes, and Class 1 Integrons in Salmonella enterica serovar Typhimurium Isolates from Chicken Meat and Humans in Egypt. Foodborne Pathog. Dis. 2016, 13, 281–288. [Google Scholar] [CrossRef]
- Hopkins, K.L.; Arnold, C.; Threlfall, E.J. Rapid detection of gyrA and parC mutations in quinolone-resistant Salmonella enterica using Pyrosequencing technology. J. Microbiol. Methods 2007, 68, 163–171. [Google Scholar] [CrossRef]
- Garmory, H.S.; Brown, K.A.; Titball, R.W. Salmonella vaccines for use in humans: Present and future perspectives. FEMS Microbiol. Rev. 2002, 26, 339–353. [Google Scholar]
- Edwards, P.R.; Kauffmann, F. A simplification of the Kauffmann-White schema. Am. J. Clin. Pathol. 1952, 22, 692–697. [Google Scholar] [CrossRef]
- Smith, N.H.; Selander, R.K. Sequence invariance of the antigen-coding central region of the phase 1 flagellar filament gene (fliC) among strains of Salmonella typhimurium. J. Bacteriol. 1990, 172, 603–609. [Google Scholar] [CrossRef]
- Winfield, M.D.; Groisman, E.A. Evolution and Ecology of Salmonella. EcoSal Plus 2004, 1. [Google Scholar] [CrossRef] [PubMed]
- Calenge, F.; Kaiser, P.; Vignal, A.; Beaumont, C. Genetic control of resistance to salmonellosis and to Salmonella carrier-state in fowl: A review. Genet. Sel. Evol. 2010, 42, 11. [Google Scholar] [CrossRef] [PubMed]
- Brenner, F.W.; Villar, R.G.; Angulo, F.J.; Tauxe, R.; Swaminathan, B. Salmonella nomenclature. J. Clin. Microbiol. 2000, 38, 2465–2467. [Google Scholar] [CrossRef] [PubMed]
- Reeves, M.W.; Evins, G.M.; Heiba, A.A.; Plikaytis, B.D.; Farmer, J.J. Clonal nature of Salmonella typhi and its genetic relatedness to other salmonellae as shown by multilocus enzyme electrophoresis, and proposal of Salmonella bongori comb. nov. J. Clin. Microbiol. 1989, 27, 313–320. [Google Scholar] [CrossRef]
- Li, J.; Nelson, K.; McWhorter, A.C.; Whittam, T.S.; Selander, R.K. Recombinational basis of serovar diversity in Salmonella enterica. Proc. Natl. Acad. Sci. USA 1994, 91, 2552–2556. [Google Scholar] [CrossRef]
- Beltran, P.; Musser, J.M.; Helmuth, R.; Farmer, J.J.; Frerichs, W.M.; Wachsmuth, I.K.; Ferris, K. Toward a population genetic analysis of Salmonella: Genetic diversity and relationships among strains of serotypes S. choleraesuis, S. derby, S. dublin, S. enteritidis, S. heidelberg, S. infantis, S. newport, and S. typhimurium. Proc. Natl. Acad. Sci. USA 1988, 85, 7753–7757. [Google Scholar] [CrossRef]
- Uzzau, S.; Brown, D.J.; Wallis, T.; Rubino, S.; Leori, G.; Bernard, S.; Casadesus, J. Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 2000, 125, 229–255. [Google Scholar] [CrossRef]
- Bale, J.; Meunier, D.; Weill, F.-X.; dePinna, E.; Peters, T.; Nair, S. Characterization of new Salmonella serovars by whole-genome sequencing and traditional typing techniques. J. Med. Microbiol. 2016, 65, 1074–1078. [Google Scholar] [CrossRef]
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Deatherage Kaiser, B.L.; Dinsmore, B.A.; Fitzgerald, C. Salmonella serotype determination utilizing high-throughput genome sequencing data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef]
- Gordon, M.A. Salmonella infections in immunocompromised adults. J. Infect. 2008, 56, 413–422. [Google Scholar] [CrossRef]
- Admassu, D.; Egata, G.; Teklemariam, Z. Prevalence and antimicrobial susceptibility pattern of Salmonella enterica serovar Typhi and Salmonella enterica serovar Paratyphi among febrile patients at Karamara Hospital, Jigjiga, eastern Ethiopia. SAGE Open Med. 2019, 7. [Google Scholar] [CrossRef]
- Schadich, E.; Dzubak, P.; Hajduch, M. Role of Salmonella Typhi Vi Antigen and Secretory Systems on Immune Response. Curr. Pharm. Design 2016, 22, 6251–6260. [Google Scholar] [CrossRef] [PubMed]
- Jorge, J.F.; Costa, A.B.; Rodrigues, J.L.; Girao, E.S.; Luiz, R.S.; Sousa, A.Q.; Moore, S.R. Salmonella typhi liver abscess overlying a metastatic melanoma. Am. J. Trop. Med. Hyg. 2014, 90, 716–718. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, V.; Eslick, G.D. Systematic review with meta-analysis: The relationship between chronic Salmonella typhi carrier status and gall-bladder cancer. Aliment. Pharmacol. Ther. 2014, 39, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Wang, Y.; Su, L.H.; Chiu, C.H. Nontyphoid salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Wan, J.; Xiong, Y.; Wu, K.; Cheong, W.C.; Zhou, G.; Wang, D. A quantitative study of charge carrier dynamics in well-defined WO3 nanowires and nanosheets: Insight into the crystal facet effect in photocatalysis. J. Am. Chem. Soc. 2018. [Google Scholar] [CrossRef]
- Kurtz, J.R.; Goggins, J.A.; McLachlan, J.B. Salmonella infection: Interplay between the bacteria and host immune system. Immunol. Lett. 2017, 190, 42–50. [Google Scholar] [CrossRef]
- Molbak, K. Human health consequences of antimicrobial drug-resistant Salmonella and other foodborne pathogens. Clin. Infect. Dis. 2005, 41, 1613–1620. [Google Scholar]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef]
- Gordon, M.A. Invasive nontyphoidal Salmonella disease: Epidemiology, pathogenesis and diagnosis. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef]
- Jones, T.F.; Ingram, L.A.; Cieslak, P.R.; Vugia, D.J.; Tobin-D’Angelo, M.; Hurd, S.; Medus, C. Salmonellosis outcomes differ substantially by serotype. J. Infect. Dis. 2008, 198, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Amuasi, J.H.; May, J. Non-typhoidal salmonella: Invasive, lethal, and on the loose. Lancet Infect. Dis. 2019, 19, 1267–1269. [Google Scholar] [CrossRef]
- Lim, S.H.; Methé, B.A.; Knoll, B.M.; Morris, A.; Obaro, S.K. Invasive non-typhoidal Salmonella in sickle cell disease in Africa: Is increased gut permeability the missing link? J. Transl. Med. 2018, 16, 239. [Google Scholar] [CrossRef]
- Gordon, M.A.; Kankwatira, A.M.; Mwafulirwa, G.; Walsh, A.L.; Hopkins, M.J.; Parry, C.M.; Faragher, E.B. Invasive non-typhoid salmonellae establish systemic intracellular infection in HIV-infected adults: An emerging disease pathogenesis. Clin. Infect. Dis. 2010, 50, 953–962. [Google Scholar] [CrossRef]
- Gilchrist, J.J.; MacLennan, C.A. Invasive Nontyphoidal Salmonella Disease in Africa. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef]
- Blondel, C.J.; Jimenez, J.C.; Contreras, I.; Santiviago, C.A. Comparative genomic analysis uncovers 3 novel loci encoding type six secretion systems differentially distributed in Salmonella serotypes. BMC Genom. 2009, 10, 354. [Google Scholar] [CrossRef]
- Sevellec, Y.; Vignaud, M.L.; Granier, S.A.; Lailler, R.; Feurer, C.; Le Hello, S.; Mistou, M.Y. Polyphyletic Nature of Salmonella enterica Serotype Derby and Lineage-Specific Host-Association Revealed by Genome-Wide Analysis. Front. Microbiol. 2018, 9, 891. [Google Scholar] [CrossRef]
- Rice, C.J.; Ramachandran, V.K.; Shearer, N.; Thompson, A. Transcriptional and Post-Transcriptional Modulation of SPI1 and SPI2 Expression by ppGpp, RpoS and DksA in Salmonella enterica sv Typhimurium. PLoS ONE 2015, 10, e0127523. [Google Scholar] [CrossRef]
- Buckner, M.M.; Croxen, M.A.; Arena, E.T.; Finlay, B.B. A comprehensive study of the contribution of Salmonella enterica serovar Typhimurium SPI2 effectors to bacterial colonization, survival, and replication in typhoid fever, macrophage, and epithelial cell infection models. Virulence 2011, 2, 208–216. [Google Scholar] [CrossRef]
- Blanc-Potard, A.B.; Solomon, F.; Kayser, J.; Groisman, E.A. The SPI-3 pathogenicity island of Salmonella enterica. J. Bacteriol. 1999, 181, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, I.; Karasova, D.; Sebkova, A.; Volf, J.; Sisak, F.; Havlickova, H.; Kummer, V. Virulence potential of five major pathogenicity islands (SPI-1 to SPI-5) of Salmonella enterica serovar Enteritidis for chickens. BMC Microbiol. 2009, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Morgan, E.; Nagy, G. Contribution of SPI-4 genes to the virulence of Salmonella enterica. FEMS Microbiol. Lett. 2007, 275, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Hensel, M. Salmonella pathogenicity island 2. Mol. Microbiol. 2000, 36, 1015–1023. [Google Scholar] [CrossRef]
- Hautefort, I.; Thompson, A.; Eriksson-Ygberg, S.; Parker, M.L.; Lucchini, S.; Danino, V.; Bongaerts, R.J. During infection of epithelial cells Salmonella enterica serovar Typhimurium undergoes a time-dependent transcriptional adaptation that results in simultaneous expression of three type 3 secretion systems. Cell. Microbiol. 2008, 10, 958–984. [Google Scholar] [CrossRef]
- Galan, J.E. Molecular genetic bases of Salmonella entry into host cells. Mol. Microbiol. 1996, 20, 263–271. [Google Scholar] [CrossRef]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef]
- Shea, J.E.; Hensel, M.; Gleeson, C.; Holden, D.W. Identification of a virulence locus encoding a second type III secretion system in Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1996, 93, 2593–2597. [Google Scholar] [CrossRef]
- Shappo, M.O.E.; Li, Q.; Lin, Z.; Hu, M.; Ren, J.; Xu, Z.; Pan, Z. SspH2 as anti-inflammatory candidate effector and its contribution in Salmonella Enteritidis virulence. Microb. Pathog. 2020, 142, 104041. [Google Scholar] [CrossRef]
- Zhang, K.; Riba, A.; Nietschke, M.; Torow, N.; Repnik, U.; Pütz, A.; Fulde, M. Minimal SPI1-T3SS effector requirement for Salmonella enterocyte invasion and intracellular proliferation in vivo. PLoS Pathog. 2018, 14, e1006925. [Google Scholar] [CrossRef]
- Yang, S.; Deng, Q.; Sun, L.; Dong, K.; Li, Y.; Wu, S.; Huang, R. Effector SpvB interferes with intracellular iron homeostasis regulation of transcription factor NRF2. FASEB J. 2019, 33, 13450–13464. [Google Scholar] [CrossRef] [PubMed]
- Knuff, K.; Finlay, B.B. What the SIF Is Happening-The Role of Intracellular-Induced Filaments. Front. Cell. Infect. Microbiol. 2017, 7, 335. [Google Scholar] [CrossRef]
- Rajashekar, R.; Liebl, D.; Chikkaballi, D.; Liss, V.; Hensel, M. Live cell imaging reveals novel functions of Salmonella enterica SPI2-T3SS effector proteins in remodeling of the host cell endosomal system. PLoS ONE. 2014, 9, e115423. [Google Scholar] [CrossRef] [PubMed]
- Liao, A.P.; Petrof, E.O.; Kuppireddi, S.; Zhao, Y.; Xia, Y.; Claud, E.C.; Sun, J. Salmonella type III effector AvrA stabilizes cell tight junctions to inhibit inflammation in intestinal epithelial cells. PLoS ONE 2008, 3, e2369. [Google Scholar] [CrossRef] [PubMed]
- Jennings, E.; Esposito, D.; Rittinger, K. Structure-function analyses of the bacterial zinc metalloprotease effector protein GtgA uncover key residues required for deactivating NF-kappaB. J. Biol. Chem. 2018, 293, 15316–15329. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Kamanova, J.; Lara-Tejero, M.; Galan, J.E. A Family of Salmonella Type III Secretion Effector Proteins Selectively Targets the NF-kappaB Signaling Pathway to Preserve Host Homeostasis. PLoS Pathog. 2016, 12, e1005484. [Google Scholar] [CrossRef]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 Type III Secretion System Effectors: Molecular Mechanisms and Physiological Consequences. Cell Host Microbe 2017, 22, 217–231. [Google Scholar] [CrossRef]
- Knodler, L.A.; Steele-Mortimer, O. The Salmonella effector PipB2 affects late endosome/lysosome distribution to mediate Sif extension. Mol. Biol. Cell 2005, 16, 4108–4123. [Google Scholar] [CrossRef]
- Henry, T.; Couillault, C.; Rockenfeller, P.; Boucrot, E.; Dumont, A.; Schroeder, N.; Hermant, A. The Salmonella effector protein PipB2 is a linker for kinesin-1. Proc. Natl. Acad. Sci. USA 2006, 103, 13497–13502. [Google Scholar] [CrossRef]
- McGourty, K.; Thurston, T.L.; Matthews, S.A.; Pinaud, L.; Mota, L.J.; Holden, D.W. Salmonella inhibits retrograde trafficking of mannose-6-phosphate receptors and lysosome function. Science 2012, 338, 963–967. [Google Scholar] [CrossRef]
- Brawn, L.C.; Hayward, R.D.; Koronakis, V. Salmonella SPI1 effector SipA persists after entry and cooperates with a SPI2 effector to regulate phagosome maturation and intracellular replication. Cell Host Microbe 2007, 1, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Myeni, S.K.; Wang, L.; Zhou, D. SipB-SipC complex is essential for translocon formation. PLoS ONE 2013, 8, e60499. [Google Scholar] [CrossRef] [PubMed]
- Glasgow, A.A.; Wong, H.T.; Tullman-Ercek, D. A Secretion-Amplification Role for Salmonella enterica Translocon Protein SipD. ACS Synth. Biol. 2017, 6, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Kamanova, J.; Sun, H.; Lara-Tejero, M.; Galan, J.E. The Salmonella Effector Protein SopA Modulates Innate Immune Responses by Targeting TRIM E3 Ligase Family Members. PLoS Pathog. 2016, 12, e1005552. [Google Scholar] [CrossRef] [PubMed]
- Perrett, C.A.; Zhou, D. Salmonella type III effector SopB modulates host cell exocytosis. Emerg. Microbes Infect. 2013, 2, e32. [Google Scholar] [CrossRef]
- D’Costa, V.M.; Braun, V.; Landekic, M.; Shi, R.; Proteau, A.; McDonald, L.; Cygler, M. Salmonella Disrupts Host Endocytic Trafficking by SopD2-Mediated Inhibition of Rab7. Cell Rep. 2015, 12, 1508–1518. [Google Scholar] [CrossRef]
- Vonaesch, P.; Sellin, M.E.; Cardini, S.; Singh, V.; Barthel, M.; Hardt, W.-D. The Salmonella Typhimurium effector protein SopE transiently localizes to the early SCV and contributes to intracellular replication. Cell. Microbiol. 2014, 16, 1723–1735. [Google Scholar] [CrossRef]
- Johnson, R.; Byrne, A.; Berger, C.N.; Klemm, E.; Crepin, V.F.; Dougan, G.; Frankel, G. The Type III Secretion System Effector SptP of Salmonella enterica Serovar Typhi. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [PubMed]
- Haneda, T.; Ishii, Y.; Shimizu, H.; Ohshima, K.; Iida, N.; Danbara, H.; Okada, N. Salmonella type III effector SpvC, a phosphothreonine lyase, contributes to reduction in inflammatory response during intestinal phase of infection. Cell. Microbiol. 2012, 14, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 158, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Hensel, M.; Shea, J.E.; Raupach, B.; Monack, D.; Falkow, S.; Gleeson, C.; Kubo, T. Functional analysis of ssaJ and the ssaK/U operon, 13 genes encoding components of the type III secretion apparatus of Salmonella Pathogenicity Island 2. Mol. Microbiol. 1997, 24, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Gallois, A.; Klein, J.R.; Allen, L.A.; Jones, B.D.; Nauseef, W.M. Salmonella pathogenicity island 2-encoded type III secretion system mediates exclusion of NADPH oxidase assembly from the phagosomal membrane. J. Immunol. 2001, 166, 5741–5748. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, G.L.; Müller, P.; Hensel, M. Functional dissection of SseF, a type III effector protein involved in positioning the salmonella-containing vacuole. Traffic 2006, 7, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejek, A.M.; Miller, S.I. Salmonella modulation of the phagosome membrane, role of SseJ. Cell. Microbiol. 2015, 17, 333–341. [Google Scholar] [CrossRef]
- Günster, R.A.; Matthews, S.A.; Holden, D.W.; Thurston, T.L.M. SseK1 and SseK3 Type III Secretion System Effectors Inhibit NF-κB Signaling and Necroptotic Cell Death in Salmonella-Infected Macrophages. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef]
- Le Negrate, G.; Faustin, B.; Welsh, K.; Loeffler, M.; Krajewska, M.; Hasegawa, P.; Mukherjee, S. Salmonella secreted factor L deubiquitinase of Salmonella typhimurium inhibits NF-kappaB, suppresses IkappaBalpha ubiquitination and modulates innate immune responses. J. Immunol. 2008, 180, 5045–5056. [Google Scholar] [CrossRef]
- Domingues, L.; Holden, D.W.; Mota, L.J. The Salmonella effector SteA contributes to the control of membrane dynamics of Salmonella-containing vacuoles. Infect. Immun. 2014, 82, 2923–2934. [Google Scholar] [CrossRef]
- Odendall, C.; Rolhion, N.; Förster, A.; Poh, J.; Lamont, D.J.; Liu, M.; Freemont, P.S. The Salmonella kinase SteC targets the MAP kinase MEK to regulate the host actin cytoskeleton. Cell Host Microbe 2012, 12, 657–668. [Google Scholar] [CrossRef]
- Chatterjee, S.; Chaudhury, S.; McShan, A.C.; Kaur, K.; De Guzman, R.N. Structure and biophysics of type III secretion in bacteria. Biochemistry 2013, 52, 2508–2517. [Google Scholar] [CrossRef]
- Cardenal-Munoz, E.; Gutierrez, G.; Ramos-Morales, F. Global impact of Salmonella type III secretion effector SteA on host cells. Biochem. Biophys. Res. Commun. 2014, 449, 419–424. [Google Scholar] [CrossRef]
- Young, A.M.; Palmer, A.E. Methods to Illuminate the Role of Salmonella Effector Proteins during Infection: A Review. Front. Cell. Infect. Microbiol. 2017, 7, 363. [Google Scholar] [CrossRef] [PubMed]
- Janakiraman, A.; Slauch, J.M. The putative iron transport system SitABCD encoded on SPI1 is required for full virulence of Salmonella typhimurium. Mol. Microbiol. 2000, 35, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Spector, M.P. The starvation-stress response (SSR) of Salmonella. Adv. Microb. Physiol. 1998, 40, 233–279. [Google Scholar] [PubMed]
- Hirvas, L.; Koski, P.; Vaara, M. Identification and sequence analysis of the gene mutated in the conditionally lethal outer membrane permeability mutant SS-C of Salmonella typhimurium. EMBO J. 1991, 10, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Hirvas, L.; Koski, P.; Vaara, M. Primary structure and expression of the Ssc-protein of Salmonella typhimurium. Biochem. Biophys. Res. Commun. 1990, 173, 53–59. [Google Scholar] [CrossRef]
- Hensel, M.; Shea, J.E.; Waterman, S.R.; Mundy, R.; Nikolaus, T.; Banks, G.; Vazquez-Torres, A. Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol. Microbiol. 1998, 30, 163–174. [Google Scholar] [CrossRef]
- Blanc-Potard, A.B.; Groisman, E.A. The Salmonella selC locus contains a pathogenicity island mediating intramacrophage survival. EMBO J. 1997, 16, 5376–5385. [Google Scholar] [CrossRef]
- Barlag, B.; Hensel, M. The giant adhesin SiiE of Salmonella enterica. Molecules 2015, 20, 1134–1150. [Google Scholar] [CrossRef]
- Kirchweger, P.; Weiler, S.; Egerer-Sieber, C.; Blasl, A.-T.; Hoffmann, S.; Schmidt, C.; Sander, N. Structural and functional characterization of SiiA, an auxiliary protein from the SPI4-encoded type 1 secretion system from Salmonella enterica. Mol. Microbiol. 2019, 112, 1403–1422. [Google Scholar] [CrossRef]
- Gerlach, R.G.; Claudio, N.; Rohde, M.; Jackel, D.; Wagner, C.; Hensel, M. Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell. Microbiol. 2008, 10, 2364–2376. [Google Scholar] [CrossRef]
- Gerlach, R.G.; Jackel, D.; Stecher, B.; Wagner, C.; Lupas, A.; Hardt, W.D.; Hensel, M. Salmonella Pathogenicity Island 4 encodes a giant non-fimbrial adhesin and the cognate type 1 secretion system. Cell. Microbiol. 2007, 9, 1834–1850. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.W.; Jones, M.A.; Watson, P.R.; Hedges, S.; Wallis, T.S.; Galyov, E.E. Identification of a pathogenicity island required for Salmonella enteropathogenicity. Mol. Microbiol. 1998, 29, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Celli, J.; Hardt, W.D.; Vallance, B.A.; Yip, C.; Finlay, B.B. Salmonella effectors within a single pathogenicity island are differentially expressed and translocated by separate type III secretion systems. Mol. Microbiol. 2002, 43, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Allard, M.; Strain, E.; Stones, R.; Zhao, S.; Brown, E.; Meng, J. Genetic diversity of Salmonella pathogenicity islands SPI-5 and SPI-6 in Salmonella Newport. Foodborne Pathog. Dis. 2014, 11, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Saroj, S.D.; Shashidhar, R.; Karani, M.; Bandekar, J.R. Distribution of Salmonella pathogenicity island (SPI)-8 and SPI-10 among different serotypes of Salmonella. J. Med. Microbiol. 2008, 57, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Bueno, S.M.; Santiviago, C.A.; Murillo, A.A.; Fuentes, J.A.; Trombert, A.N.; Rodas, P.I.; Youderian, P. Precise excision of the large pathogenicity island, SPI7, in Salmonella enterica serovar Typhi. J. Bacteriol. 2004, 186, 3202–3213. [Google Scholar] [CrossRef]
- Tomljenovic-Berube, A.M.; Henriksbo, B.; Porwollik, S.; Cooper, C.A.; Tuinema, B.R.; McClelland, M.; Coombes, B.K. Mapping and regulation of genes within Salmonella pathogenicity island 12 that contribute to in vivo fitness of Salmonella enterica Serovar Typhimurium. Infect. Immun. 2013, 81, 2394–2404. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, S.; Helmann, J.D.; Kim, B.H.; Park, Y.K. RaoN, a small RNA encoded within Salmonella pathogenicity island-11, confers resistance to macrophage-induced stress. Microbiology 2013, 159, 1366–1378. [Google Scholar] [CrossRef]
- Velasquez, J.C.; Hidalgo, A.A.; Villagra, N.; Santiviago, C.A.; Mora, G.C.; Fuentes, J.A. SPI-9 of Salmonella enterica serovar Typhi is constituted by an operon positively regulated by RpoS and contributes to adherence to epithelial cells in culture. Microbiology 2016, 162, 1367–1378. [Google Scholar] [CrossRef]
- Pickard, D.; Wain, J.; Baker, S.; Line, A.; Chohan, S.; Fookes, M.; Barron, A. Composition, acquisition, and distribution of the Vi exopolysaccharide-encoding Salmonella enterica pathogenicity island SPI-7. J. Bacteriol. 2003, 185, 5055–5065. [Google Scholar] [CrossRef]
- Desai, P.T.; Porwollik, S.; Long, F.; Cheng, P.; Wollam, A.; Bhonagiri-Palsikar, V.; Hallsworth-Pepin, K. Evolutionary Genomics of Salmonella enterica Subspecies. MBio 2013, 4. [Google Scholar] [CrossRef]
- Hensel, M. Evolution of pathogenicity islands of Salmonella enterica. Int. J. Med. Microbiol. 2004, 294, 95–102. [Google Scholar] [CrossRef]
- Parkhill, J.; Dougan, G.; James, K.D.; Thomson, N.R.; Pickard, D.; Wain, J.; Churcher, C. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 2001, 413, 848–852. [Google Scholar] [CrossRef]
- Espinoza, R.A.; Silva-Valenzuela, C.A.; Amaya, F.A.; Urrutia, I.M.; Contreras, I.; Santiviago, C.A. Differential roles for pathogenicity islands SPI-13 and SPI-8 in the interaction of Salmonella Enteritidis and Salmonella Typhi with murine and human macrophages. Biol. Res. 2017, 50, 5. [Google Scholar] [CrossRef] [PubMed]
- Mulder, D.T.; Cooper, C.A.; Coombes, B.K. Type VI secretion system-associated gene clusters contribute to pathogenesis of Salmonella enterica serovar Typhimurium. Infect. Immun. 2012, 80, 1996–2007. [Google Scholar] [CrossRef] [PubMed]
- Sana, T.G.; Flaugnatti, N.; Lugo, K.A.; Lam, L.H.; Jacobson, A.; Baylot, V.; Durand, E. Salmonella Typhimurium utilizes a T6SS-mediated antibacterial weapon to establish in the host gut. Proc. Natl. Acad. Sci. USA 2016, 113, E5044–E5051. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, D.; Wu, X.; Yi, Z.; Wang, Y.; Xin, S.; Wang, D. The ferric uptake regulator represses type VI secretion system function by binding directly to the clpV promoter in Salmonella enterica serovar Typhimurium. Infect. Immun. 2019. [Google Scholar] [CrossRef] [PubMed]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Dusold, R.; Rubino, S.; Baumler, A.J. The Vi capsular antigen of Salmonella enterica serotype Typhi reduces Toll-like receptor-dependent interleukin-8 expression in the intestinal mucosa. Infect. Immun. 2005, 73, 3367–3374. [Google Scholar] [CrossRef]
- Liaquat, S.; Sarwar, Y. Virulotyping of Salmonella enterica serovar Typhi isolates from Pakistan: Absence of complete SPI-10 in Vi negative isolates. PLoS Negl. Trop. Dis. 2018, 12, e0006839. [Google Scholar] [CrossRef]
- Faucher, S.P.; Viau, C.; Gros, P.P.; Daigle, F.; Le Moual, H. The prpZ gene cluster encoding eukaryotic-type Ser/Thr protein kinases and phosphatases is repressed by oxidative stress and involved in Salmonella enterica serovar Typhi survival in human macrophages. FEMS Microbiol. Lett. 2008, 281, 160–166. [Google Scholar] [CrossRef]
- Miller, S.I.; Kukral, A.M.; Mekalanos, J.J. A two-component regulatory system (phoP phoQ) controls Salmonella typhimurium virulence. Proc. Natl. Acad. Sci. USA 1989, 86, 5054–5058. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Alpuche-Aranda, C.M.; Loomis, W.P.; Belden, W.J.; Miller, S.I. Characterization of the Salmonella typhimurium pagC/pagD chromosomal region. J. Bacteriol. 1995, 177, 5040–5047. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.R.; Paul, N.C.; Burin, R.; Guard, J.; Shah, D.H. Genomic organization and role of SPI-13 in nutritional fitness of Salmonella. Int. J. Med. Microbiol. 2018, 308, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, X.; Lv, R.; Jiang, X.; Cao, H.; Du, Y.; Jiang, L. Global regulatory function of the low oxygen-induced transcriptional regulator LoiA in Salmonella Typhimurium revealed by RNA sequencing. Biochem. Biophys. Res. Commun. 2018, 503, 2022–2027. [Google Scholar] [CrossRef]
- Jiang, L.; Feng, L.; Yang, B.; Zhang, W.; Wang, P.; Jiang, X.; Wang, L. Signal transduction pathway mediated by the novel regulator LoiA for low oxygen tension induced Salmonella Typhimurium invasion. PLoS Pathog. 2017, 13, e1006429. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.S.; Parkhill, J. Interpolated variable order motifs for identification of horizontally acquired DNA: Revisiting the Salmonella pathogenicity islands. Bioinformatics 2006, 22, 2196–2203. [Google Scholar] [CrossRef]
- Bogomolnaya, L.M.; Santiviago, C.A.; Yang, H.J.; Baumler, A.J.; Andrews-Polymenis, H.L. ‘Form variation’ of the O12 antigen is critical for persistence of Salmonella Typhimurium in the murine intestine. Mol. Microbiol. 2008, 70, 1105–1119. [Google Scholar] [CrossRef]
- Faucher, S.P.; Forest, C.; Beland, M.; Daigle, F. A novel PhoP-regulated locus encoding the cytolysin ClyA and the secreted invasin TaiA of Salmonella enterica serovar Typhi is involved in virulence. Microbiology 2009, 155, 477–488. [Google Scholar] [CrossRef]
- Jofre, M.R.; Rodriguez, L.M.; Villagra, N.A.; Hidalgo, A.A.; Mora, G.C.; Fuentes, J.A. RpoS integrates CRP, Fis, and PhoP signaling pathways to control Salmonella Typhi hlyE expression. BMC Microbiol. 2014, 14, 139. [Google Scholar] [CrossRef]
- Levine, M.M.; Robins-Browne, R.M. Factors that explain excretion of enteric pathogens by persons without diarrhea. Clin. Infect. Dis. 2012, 55, S303–S311. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; Cavallo, I.; Pontone, M.; Toma, L.; Ensoli, F. Biofilm Producing Salmonella Typhi: Chronic Colonization and Development of Gallbladder Cancer. Int. J. Mol. Sci. 2017, 18, 1887. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Ohlson, M.B.; Monack, D.M. Innate immune response to Salmonella typhimurium, a model enteric pathogen. Gut Microbes 2012, 3, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Boyle, E.C.; Brown, N.F.; Finlay, B.B. Salmonella enterica serovar Typhimurium effectors SopB, SopE, SopE2 and SipA disrupt tight junction structure and function. Cell. Microbiol. 2006, 8, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigon, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front. Microbiol. 2018, 9, 736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gui, S.; Liang, Z.; Liu, A.; Chen, Z.; Tang, Y.; Xiao, M. Cecropin (Mdc) Alleviates-Induced Colonic Mucosal Barrier Impairment: Associating With Inflammatory and Oxidative Stress Response, Tight Junction as Well as Intestinal Flora. Front. Microbiol. 2019, 10, 522. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, X.; Xia, X.; Zhu, C.; Zhang, H.; Qin, W.; Xu, Y. Inhibitory Effects of Antimicrobial Peptide JH-3 on Serovar Typhimurium Strain CVCC541 Infection-Induced Inflammatory Cytokine Release and Apoptosis in RAW264.7 Cells. Molecules 2019, 24, 596. [Google Scholar] [CrossRef]
- Fan, J.; Petersen, E.M.; Hinds, T.R.; Zheng, N.; Miller, S.I. Structure of an Inner Membrane Protein Required for PhoPQ-Regulated Increases in Outer Membrane Cardiolipin. MBio 2020, 11. [Google Scholar] [CrossRef]
- Gayet, R.; Bioley, G.; Rochereau, N.; Paul, S.; Corthesy, B. Vaccination against Salmonella Infection: The Mucosal Way. Microbiol. Mol. Biol. Rev. 2017, 81. [Google Scholar] [CrossRef]
- Mabbott, N.A.; Donaldson, D.S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013, 6, 666–677. [Google Scholar] [CrossRef]
- Zhao, L.; Ezak, T.; Li, Z.Y.; Kawamura, Y.; Hirose, K.; Watanabe, H. Vi-Suppressed wild strain Salmonella typhi cultured in high osmolarity is hyperinvasive toward epithelial cells and destructive of Peyer’s patches. Microbiol. Immunol. 2001, 45, 149–158. [Google Scholar] [CrossRef]
- Wang, K.C.; Huang, C.H.; Huang, C.J.; Fang, S.B. Impacts of Salmonella enterica Serovar Typhimurium and Its speG Gene on the Transcriptomes of In Vitro M Cells and Caco-2 Cells. PLoS ONE 2016, 11, e0153444. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.H.; Staudt, L.M. Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 2013, 5, a011247. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Strowig, T.; Henao-Mejia, J.; Flavell, R.A. Regulation of the antimicrobial response by NLR proteins. Immunity 2011, 34, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, T.D.; Lamkanfi, M.; Nunez, G. Intracellular NOD-like receptors in host defense and disease. Immunity 2007, 27, 549–559. [Google Scholar] [CrossRef]
- Tam, M.A.; Rydström, A.; Sundquist, M.; Wick, M.J. Early cellular responses to Salmonella infection: Dendritic cells, monocytes, and more. Immunol. Rev. 2008, 225, 140–162. [Google Scholar] [CrossRef]
- Tükel, C.; Nishimori, J.H.; Wilson, R.P.; Winter, M.G.; Keestra, A.M.; van Putten, J.P.M.; Bäumler, A.J. Toll-like receptors 1 and 2 cooperatively mediate immune responses to curli, a common amyloid from enterobacterial biofilms. Cell. Microbiol. 2010, 12, 1495–1505. [Google Scholar] [CrossRef]
- Oppong, G.O.; Rapsinski, G.J.; Newman, T.N.; Nishimori, J.H.; Biesecker, S.G.; Tükel, Ç. Epithelial cells augment barrier function via activation of the Toll-like receptor 2/phosphatidylinositol 3-kinase pathway upon recognition of Salmonella enterica serovar Typhimurium curli fibrils in the gut. Infect. Immun. 2013, 81, 478–486. [Google Scholar] [CrossRef]
- Van Bergenhenegouwen, J.; Kraneveld, A.D.; Rutten, L.; Garssen, J.; Vos, A.P.; Hartog, A. Lipoproteins attenuate TLR2 and TLR4 activation by bacteria and bacterial ligands with differences in affinity and kinetics. BMC Immunol. 2016, 17, 42. [Google Scholar] [CrossRef]
- Rathinam, V.A.K.; Zhao, Y.; Shao, F. Innate immunity to intracellular LPS. Nat. Immunol. 2019, 20, 527–533. [Google Scholar] [CrossRef]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef]
- Mikołajczyk, A.; Kozłowska, A.; Gonkowski, S. Distribution and Neurochemistry of the Porcine Ileocaecal Valve Projecting Sensory Neurons in the Dorsal Root Ganglia and the Influence of Lipopolysaccharide from Different Serotypes of spp. on the Chemical Coding of DRG Neurons in the Cell Cultures. Int. J. Mol. Sci. 2018, 19, 2551. [Google Scholar] [CrossRef] [PubMed]
- Shaio, M.F.; Rowland, H. Bactericidal and opsonizing effects of normal serum on mutant strains of Salmonella typhimurium. Infect. Immun. 1985, 49, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Avraham, R.; Haseley, N.; Brown, D.; Penaranda, C.; Jijon, H.B.; Trombetta, J.J.; Satija, R. Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses. Cell 2015, 162, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Cian, M.B.; Giordano, N.P.; Masilamani, R.; Minor, K.E.; Dalebroux, Z.D. Salmonella enterica Serovar Typhimurium Uses PbgA/YejM To Regulate Lipopolysaccharide Assembly during Bacteremia. Infect. Immun. 2019, 88. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.D.; Groisman, E.A. The biology of the PmrA/PmrB two-component system: The major regulator of lipopolysaccharide modifications. Annu. Rev. Microbiol. 2013, 67. [Google Scholar] [CrossRef]
- Zeng, H.; Carlson, A.Q.; Guo, Y.; Yu, Y.; Collier-Hyams, L.S.; Madara, J.L.; Gewirtz, A.T. Flagellin is the major proinflammatory determinant of enteropathogenic Salmonella. J. Immunol. 2003, 171, 3668–3674. [Google Scholar] [CrossRef]
- Lahiri, A.; Lahiri, A.; Das, P.; Vani, J.; Shaila, M.S.; Chakravortty, D. TLR 9 activation in dendritic cells enhances salmonella killing and antigen presentation via involvement of the reactive oxygen species. PLoS ONE 2010, 5, e13772. [Google Scholar] [CrossRef]
- Tursi, S.A.; Tükel, Ç. Curli-Containing Enteric Biofilms Inside and Out: Matrix Composition, Immune Recognition, and Disease Implications. Microbiol. Mol. Biol. Rev. 2018, 82. [Google Scholar] [CrossRef]
- Yamamoto, M.; Takeda, K. Current views of toll-like receptor signaling pathways. Gastroenterol. Res. Pract. 2010, 2010, 240365. [Google Scholar] [CrossRef]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Tukel, C.; Akcelik, M.; Baumler, A.J. Capsule-mediated immune evasion: A new hypothesis explaining aspects of typhoid fever pathogenesis. Infect. Immun. 2006, 74, 19–27. [Google Scholar] [CrossRef]
- Wilson, R.P.; Raffatellu, M.; Chessa, D.; Winter, S.E.; Tukel, C.; Baumler, A.J. The Vi-capsule prevents Toll-like receptor 4 recognition of Salmonella. Cell. Microbiol. 2008, 10, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Tran, Q.T.; Gomez, G.; Khare, S.; Lawhon, S.D.; Raffatellu, M.; Baumler, A.J.; Ajithdoss, D. The Salmonella enterica serotype Typhi Vi capsular antigen is expressed after the bacterium enters the ileal mucosa. Infect. Immun. 2010, 78, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Raffatellu, M.; Wilson, R.P.; Russmann, H.; Baumler, A.J. The Salmonella enterica serotype Typhi regulator TviA reduces interleukin-8 production in intestinal epithelial cells by repressing flagellin secretion. Cell. Microbiol. 2008, 10, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Rolhion, N.; Furniss, R.C.; Grabe, G.; Ryan, A.; Liu, M.; Matthews, S.A.; Holden, D.W. Inhibition of Nuclear Transport of NF-kB p65 by the Salmonella Type III Secretion System Effector SpvD. PLoS Pathog. 2016, 12, e1005653. [Google Scholar] [CrossRef]
- Mazurkiewicz, P.; Thomas, J.; Thompson, J.A.; Liu, M.; Arbibe, L.; Sansonetti, P.; Holden, D.W. SpvC is a Salmonella effector with phosphothreonine lyase activity on host mitogen-activated protein kinases. Mol. Microbiol. 2008, 67, 1371–1383. [Google Scholar] [CrossRef]
- Li, H.; Xu, H.; Zhou, Y.; Zhang, J.; Long, C.; Li, S.; Chen, S. The phosphothreonine lyase activity of a bacterial type III effector family. Science 2007, 315, 1000–1003. [Google Scholar] [CrossRef]
- Alpuche-Aranda, C.M.; Racoosin, E.L.; Swanson, J.A.; Miller, S.I. Salmonella stimulate macrophage macropinocytosis and persist within spacious phagosomes. J. Exp. Med. 1994, 179, 601–608. [Google Scholar] [CrossRef]
- Cirillo, D.M.; Valdivia, R.H.; Monack, D.M.; Falkow, S. Macrophage-dependent induction of the Salmonella pathogenicity island 2 type III secretion system and its role in intracellular survival. Mol. Microbiol. 1998, 30, 175–188. [Google Scholar] [CrossRef]
- Groisman, E.A. The pleiotropic two-component regulatory system PhoP-PhoQ. J. Bacteriol. 2001, 183, 1835–1842. [Google Scholar] [CrossRef]
- Thompson, J.A.; Liu, M.; Helaine, S.; Holden, D.W. Contribution of the PhoP/Q regulon to survival and replication of Salmonella enterica serovar Typhimurium in macrophages. Microbiology 2011, 157, 2084–2093. [Google Scholar] [CrossRef]
- Klein, J.A.; Grenz, J.R.; Slauch, J.M. Controlled Activity of the Salmonella Invasion-Associated Injectisome Reveals Its Intracellular Role in the Cytosolic Population. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Liu, M.; Holden, D.W. Salmonella Effectors SseF and SseG Interact with Mammalian Protein ACBD3 (GCP60) To Anchor Salmonella-Containing Vacuoles at the Golgi Network. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Keestra, A.M.; Winter, M.G.; Klein-Douwel, D.; Xavier, M.N.; Winter, S.E.; Kim, A.; Tsolis, R.M. A Salmonella virulence factor activates the NOD1/NOD2 signaling pathway. MBio 2011, 2. [Google Scholar] [CrossRef]
- Geddes, K.; Rubino, S.; Streutker, C.; Cho, J.H.; Magalhaes, J.G.; Le Bourhis, L.; Selvanantham, T. Nod1 and Nod2 regulation of inflammation in the Salmonella colitis model. Infect. Immun. 2010, 78, 5107–5115. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef]
- Ferrand, J.; Ferrero, R.L. Recognition of Extracellular Bacteria by NLRs and Its Role in the Development of Adaptive Immunity. Front. Immunol 2013, 4, 344. [Google Scholar] [CrossRef]
- Reyes Ruiz, V.M.; Ramirez, J.; Naseer, N.; Palacio, N.M.; Siddarthan, I.J.; Yan, B.M.; Boyer, M.A. Broad detection of bacterial type III secretion system and flagellin proteins by the human NAIP/NLRC4 inflammasome. Proc. Natl. Acad. Sci. USA 2017, 114, 13242–13247. [Google Scholar] [CrossRef]
- Matusiak, M.; Van Opdenbosch, N.; Vande Walle, L.; Sirard, J.-C.; Kanneganti, T.-D.; Lamkanfi, M. Flagellin-induced NLRC4 phosphorylation primes the inflammasome for activation by NAIP5. Proc. Natl. Acad. Sci. USA 2015, 112, 1541–1546. [Google Scholar] [CrossRef]
- Qu, Y.; Misaghi, S.; Izrael-Tomasevic, A.; Newton, K.; Gilmour, L.L.; Lamkanfi, M.; Louie, S. Phosphorylation of NLRC4 is critical for inflammasome activation. Nature 2012, 490, 539–542. [Google Scholar] [CrossRef]
- Kayagaki, N.; Wong, M.T.; Stowe, I.B.; Ramani, S.R.; Gonzalez, L.C.; Akashi-Takamura, S.; Miyake, K. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science 2013, 341, 1246–1249. [Google Scholar] [CrossRef]
- Meunier, E.; Dick, M.S.; Dreier, R.F.; Schurmann, N.; Kenzelmann Broz, D.; Warming, S.; Roose-Girma, M. Caspase-11 activation requires lysis of pathogen-containing vacuoles by IFN-induced GTPases. Nature 2014, 509, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Bierschenk, D.; Monteleone, M.; Moghaddas, F.; Baker, P.J.; Masters, S.L.; Boucher, D.; Schroder, K. The Salmonella pathogenicity island-2 subverts human NLRP3 and NLRC4 inflammasome responses. J. Leukoc. Biol. 2019, 105, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Drecktrah, D.; Knodler, L.A.; Howe, D.; Steele-Mortimer, O. Salmonella trafficking is defined by continuous dynamic interactions with the endolysosomal system. Traffic 2007, 8, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.K.; Alpuche-Aranda, C.; Berthiaume, E.; Jinks, T.; Miller, S.I.; Swanson, J.A. Rapid and complete fusion of macrophage lysosomes with phagosomes containing Salmonella typhimurium. Infect. Immun. 1996, 64, 3877–3883. [Google Scholar] [CrossRef] [PubMed]
- Jantsch, J.; Chikkaballi, D.; Hensel, M. Cellular aspects of immunity to intracellular Salmonella enterica. Immunol. Rev. 2011, 240, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Xu, Y.; Jones-Carson, J.; Holden, D.W.; Lucia, S.M.; Dinauer, M.C.; Mastroeni, P. Salmonella pathogenicity island 2-dependent evasion of the phagocyte NADPH oxidase. Science 2000, 287, 1655–1658. [Google Scholar] [CrossRef]
- Fang, F.C. Antimicrobial actions of reactive oxygen species. MBio 2011, 2. [Google Scholar] [CrossRef]
- Bourret, T.J.; Song, M.; Vazquez-Torres, A. Codependent and independent effects of nitric oxide-mediated suppression of PhoPQ and Salmonella pathogenicity island 2 on intracellular Salmonella enterica serovar typhimurium survival. Infect. Immun. 2009, 77, 5107–5115. [Google Scholar] [CrossRef]
- Henard, C.A.; Vazquez-Torres, A. Nitric oxide and salmonella pathogenesis. Front. Microbiol. 2011, 2, 84. [Google Scholar] [CrossRef]
- Eswarappa, S.M.; Negi, V.D.; Chakraborty, S.; Chandrasekhar Sagar, B.K.; Chakravortty, D. Division of the Salmonella-containing vacuole and depletion of acidic lysosomes in Salmonella-infected host cells are novel strategies of Salmonella enterica to avoid lysosomes. Infect. Immun. 2010, 78, 68–79. [Google Scholar] [CrossRef]
- Van der Velden, A.W.; Lindgren, S.W.; Worley, M.J.; Heffron, F. Salmonella pathogenicity island 1-independent induction of apoptosis in infected macrophages by Salmonella enterica serotype typhimurium. Infect. Immun. 2000, 68, 5702–5709. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.L.; Cookson, B.T. Pyroptosis and host cell death responses during Salmonella infection. Cell. Microbiol. 2007, 9, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Cookson, B.T. Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol. Microbiol. 2000, 38, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Miao, E.A.; Alpuche-Aranda, C.M.; Dors, M.; Clark, A.E.; Bader, M.W.; Miller, S.I.; Aderem, A. Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1beta via Ipaf. Nat. Immunol. 2006, 7, 569–575. [Google Scholar] [CrossRef]
- Franchi, L.; Amer, A.; Body-Malapel, M.; Kanneganti, T.D.; Ozoren, N.; Jagirdar, R.; Inohara, N. Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1beta in salmonella-infected macrophages. Nat. Immunol. 2006, 7, 576–582. [Google Scholar] [CrossRef]
- Monack, D.M.; Detweiler, C.S.; Falkow, S. Salmonella pathogenicity island 2-dependent macrophage death is mediated in part by the host cysteine protease caspase-1. Cell. Microbiol. 2001, 3, 825–837. [Google Scholar] [CrossRef]
- Ygberg, S.E.; Clements, M.O.; Rytkonen, A.; Thompson, A.; Holden, D.W.; Hinton, J.C.; Rhen, M. Polynucleotide phosphorylase negatively controls spv virulence gene expression in Salmonella enterica. Infect. Immun. 2006, 74, 1243–1254. [Google Scholar] [CrossRef]
- Navarre, W.W.; Zychlinsky, A. Pathogen-induced apoptosis of macrophages: A common end for different pathogenic strategies. Cell. Microbiol. 2000, 2, 265–273. [Google Scholar] [CrossRef]
- Knodler, L.A. Salmonella enterica: Living a double life in epithelial cells. Curr. Opin. Microbiol. 2015, 23, 23–31. [Google Scholar] [CrossRef]
- Knodler, L.A.; Vallance, B.A.; Celli, J.; Winfree, S.; Hansen, B.; Montero, M.; Steele-Mortimer, O. Dissemination of invasive Salmonella via bacterial-induced extrusion of mucosal epithelia. Proc. Natl. Acad. Sci. USA 2010, 107, 17733–17738. [Google Scholar] [CrossRef]
- Malik-Kale, P.; Winfree, S.; Steele-Mortimer, O. The bimodal lifestyle of intracellular Salmonella in epithelial cells: Replication in the cytosol obscures defects in vacuolar replication. PLoS ONE 2012, 7, e38732. [Google Scholar] [CrossRef] [PubMed]
- Beuzon, C.R.; Salcedo, S.P.; Holden, D.W. Growth and killing of a Salmonella enterica serovar Typhimurium sifA mutant strain in the cytosol of different host cell lines. Microbiology 2002, 148, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Tattoli, I.; Sorbara, M.T.; Vuckovic, D.; Ling, A.; Soares, F.; Carneiro, L.A.M.; Yang, C. Amino acid starvation induced by invasive bacterial pathogens triggers an innate host defense program. Cell Host Microbe 2012, 11, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Cheminay, C.; Mohlenbrink, A.; Hensel, M. Intracellular Salmonella inhibit antigen presentation by dendritic cells. J. Immunol. 2005, 174, 2892–2899. [Google Scholar] [CrossRef]
- Brink, T.; Leiss, V.; Siegert, P.; Jehle, D.; Ebner, J.K.; Schwan, C.; Shymanets, A. Salmonella Typhimurium effector SseI inhibits chemotaxis and increases host cell survival by deamidation of heterotrimeric Gi proteins. PLoS Pathog. 2018, 14, e1007248. [Google Scholar] [CrossRef]
- Hormaeche, C.E.; Harrington, K.A.; Joysey, H.S. Natural resistance to salmonellae in mice: Control by genes within the major histocompatibility complex. J. Infect. Dis. 1985, 152, 1050–1056. [Google Scholar] [CrossRef]
- Nauciel, C.; Ronco, E.; Guenet, J.L.; Pla, M. Role of H-2 and non-H-2 genes in control of bacterial clearance from the spleen in Salmonella typhimurium-infected mice. Infect. Immun. 1988, 56, 2407–2411. [Google Scholar] [CrossRef]
- Mitchell, E.K.; Mastroeni, P.; Kelly, A.P.; Trowsdale, J. Inhibition of cell surface MHC class II expression by Salmonella. Eur. J. Immunol. 2004, 34, 2559–2567. [Google Scholar] [CrossRef]
- Bayer-Santos, E.; Durkin, C.H.; Rigano, L.A.; Kupz, A.; Alix, E.; Cerny, O.; Jennings, E. The Salmonella Effector SteD Mediates MARCH8-Dependent Ubiquitination of MHC II Molecules and Inhibits T cell Activation. Cell Host Microbe 2016, 20, 584–595. [Google Scholar] [CrossRef]
- Godlee, C.; Cerny, O.; Durkin, C.H.; Holden, D.W. SrcA is a chaperone for the Salmonella SPI-2 type three secretion system effector SteD. Microbiology 2019, 165, 15–25. [Google Scholar] [CrossRef]
- Tobar, J.A.; Carreno, L.J.; Bueno, S.M.; Gonzalez, P.A.; Mora, J.E.; Quezada, S.A.; Kalergis, A.M. Virulent Salmonella enterica serovar typhimurium evades adaptive immunity by preventing dendritic cells from activating T cells. Infect. Immun. 2006, 74, 6438–6448. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNgamma neutralization. J. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Pie, S.; Matsiota-Bernard, P.; Truffa-Bachi, P.; Nauciel, C. Gamma interferon and interleukin-10 gene expression in innately susceptible and resistant mice during the early phase of Salmonella typhimurium infection. Infect. Immun. 1996, 64, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Nauciel, C.; Espinasse-Maes, F. Role of gamma interferon and tumor necrosis factor alpha in resistance to Salmonella typhimurium infection. Infect. Immun. 1992, 60, 450–454. [Google Scholar] [CrossRef]
- Mastroeni, P.; Harrison, J.A.; Robinson, J.H.; Clare, S.; Khan, S.; Maskell, D.J.; Dougan, G. Interleukin-12 is required for control of the growth of attenuated aromatic-compound-dependent salmonellae in BALB/c mice: Role of gamma interferon and macrophage activation. Infect. Immun. 1998, 66, 4767–4776. [Google Scholar] [CrossRef]
- MacLennan, C.; Fieschi, C.; Lammas, D.A.; Picard, C.; Dorman, S.E.; Sanal, O.; MacLennan, J.M. Interleukin (IL)-12 and IL-23 are key cytokines for immunity against Salmonella in humans. J. Infect. Dis. 2004, 190, 1755–1757. [Google Scholar] [CrossRef]
- Staretz-Haham, O.; Melamed, R.; Lifshitz, M.; Porat, N.; Fieschi, C.; Casanova, J.L.; Levy, J. Interleukin-12 receptor beta1 deficiency presenting as recurrent Salmonella infection. Clin. Infect. Dis. 2003, 37, 137–140. [Google Scholar] [CrossRef]
- Mittrucker, H.W.; Kohler, A.; Kaufmann, S.H. Characterization of the murine T-lymphocyte response to Salmonella enterica serovar Typhimurium infection. Infect. Immun. 2002, 70, 199–203. [Google Scholar] [CrossRef]
- Eisenstein, T.K.; Huang, D.; Meissler, J.J., Jr.; al-Ramadi, B. Macrophage nitric oxide mediates immunosuppression in infectious inflammation. Immunobiology 1994, 191, 493–502. [Google Scholar] [CrossRef]
- Pie, S.; Truffa-Bachi, P.; Pla, M.; Nauciel, C. Th1 response in Salmonella typhimurium-infected mice with a high or low rate of bacterial clearance. Infect. Immun. 1997, 65, 4509–4514. [Google Scholar] [CrossRef]
- MacFarlane, A.S.; Schwacha, M.G.; Eisenstein, T.K. In vivo blockage of nitric oxide with aminoguanidine inhibits immunosuppression induced by an attenuated strain of Salmonella typhimurium, potentiates Salmonella infection, and inhibits macrophage and polymorphonuclear leukocyte influx into the spleen. Infect. Immun. 1999, 67, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Nauciel, C. Role of CD4+ T cells and T-independent mechanisms in acquired resistance to Salmonella typhimurium infection. J. Immunol. 1990, 145, 1265–1269. [Google Scholar] [PubMed]
- Mittrucker, H.W.; Kohler, A.; Mak, T.W.; Kaufmann, S.H. Critical role of CD28 in protective immunity against Salmonella typhimurium. J. Immunol. 1999, 163, 6769–6776. [Google Scholar] [PubMed]
- Mei, Y.; Zhao, L.; Liu, Y.; Gong, H.; Song, Y.; Lei, L.; Zhu, Y. Combining DNA Vaccine and AIDA-1 in Attenuated Salmonella Activates Tumor-Specific CD4(+) and CD8(+) T-cell Responses. Cancer Immunol. Res. 2017, 5, 503–514. [Google Scholar] [CrossRef]
- Wahid, R.; Fresnay, S.; Levine, M.M.; Sztein, M.B. Cross-reactive multifunctional CD4+ T cell responses against Salmonella enterica serovars Typhi, Paratyphi A and Paratyphi B in humans following immunization with live oral typhoid vaccine Ty21a. Clin. Immunol. 2016, 173, 87–95. [Google Scholar] [CrossRef]
- Mastroeni, P.; Villarreal-Ramos, B.; Hormaeche, C.E. Role of T cells, TNF alpha and IFN gamma in recall of immunity to oral challenge with virulent salmonellae in mice vaccinated with live attenuated aro-Salmonella vaccines. Microb. Pathog. 1992, 13, 477–491. [Google Scholar] [CrossRef]
- Mittrucker, H.W.; Raupach, B.; Kohler, A.; Kaufmann, S.H. Cutting edge: Role of B lymphocytes in protective immunity against Salmonella typhimurium infection. J. Immunol. 2000, 164, 1648–1652. [Google Scholar] [CrossRef]
- Perez-Shibayama, C.; Gil-Cruz, C.; Pastelin-Palacios, R.; Cervantes-Barragan, L.; Hisaki, E.; Chai, Q.; Onder, L. IFN-gamma-producing CD4+ T cells promote generation of protective germinal center-derived IgM + B cell memory against Salmonella Typhi. J. Immunol. 2014, 192, 5192–5200. [Google Scholar] [CrossRef]
- Kantele, A.; Arvilommi, H.; Jokinen, I. Specific immunoglobulin-secreting human blood cells after peroral vaccination against Salmonella typhi. J. Infect. Dis. 1986, 153, 1126–1131. [Google Scholar] [CrossRef]
- Tang, Y.; Foster, N.; Jones, M.A.; Barrow, P.A. A model of persistent Salmonella infection: Salmonella Pullorum modulates the immune response of the chicken from a Th17 towards a Th2-type response. Infect. Immun. 2018. [Google Scholar] [CrossRef]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Effectors | Pathogenicity Island | Function (s) | Key Reference (s) |
|---|---|---|---|
| AvrA | SPI-1/SPI-2 | Stabilizes the intestinal epithelial permeability and tight junctions; cysteine protease; inhibits NF-κB signaling | [84] |
| GogA | SPI-2 | Cleaves the subset of NF-κB subunits; inhibits NF-κB signaling | [85,86] |
| GogB | SPI-2 | Inhibits NF-κB signaling | [85] |
| GtgA | SPI-2 | Inhibits NF-κB signaling | [85] |
| GtgE | SPI-1/SPI-2 | Promotes replication inside murine macrophages | [87] |
| PipA | SPI-2 | Cleaves the subset of NF-κB subunits; inhibits NF-κB signaling | [85] |
| PipB | SPI-2 | Targeted to SIFs | [88] |
| PipB2 | SPI-2 | Resists extraction by high salt, high pH; implicated in recruitment of kinesin-1 to SCV | [89] |
| SifA | SPI-2 | Detoxifies lysosomes; subverts human NLRP3 and NLRC4 inflammasome; required for SCV membrane stability; SIF formation; contributes to T3SS1-independent inflammation | [90] |
| SifB | SPI-2 | Targeted to SIFs | [82,83] |
| SipA | SPI-1 | Enhances actin filament assembly; promotes proliferation of cytosolic Salmonella; disrupts tight junctions; SCV trafficking | [91] |
| SipB | SPI-1 | Cholesterol-binding translocon component; triggers apoptosis via caspase-1 activation in macrophages and DCs | [82] |
| SipC | SPI-1 | Translocon component: mediates effector molecule translocation; promotes actin polymerization and bundling | [92] |
| SipD | SPI-1 | Translocon component | [93] |
| Slrp | SPI-1/SPI-2 | Inhibits the release of IL-1β | [87] |
| SopA | SPI-1 | A HECT-like E3 ubiquitin ligase | [94] |
| SopB | SPI-1 | Modulates SCV trafficking; phosphoinositide phosphatase; involved in phagosomal closure; enhances RhoG activation; disrupts tight junctions; stimulates chloride secretion; prevents apoptosis through activation of Akt | [95] |
| SopD | SPI-1/SPI-2 | SIF formation, prevents accumulation of Rab32 on SCV and SIFs | [87] |
| SopD2 | SPI-2 | Targeted to SIFs and late endosomes | [96] |
| SopE | SPI-1 | Promotes colonization of Salmonella; induces remodeling of actin | [97] |
| SopE2 | SPI-1 | Guanine nucleotide exchange factor for Cdc42; promotes pro-inflammatory signaling | [80] |
| SpiC | SPI-2 | Interferes with vesicular trafficking in host cells to prevent SCV-lysosome fusion | [92] |
| SptP | SPI-1 | Rho GAP domain functions in downregulating host membrane ruffling after entry; tyrosine phosphatase domain acts on ACK; vimentin; and presumably other substrates | [98] |
| SpvB | SPI-2 | Promotes macrophage apoptosis and P-body disassembly | [81,87] |
| SpvC | SPI-1/SPI-2 | Inhibits MAPK signaling | [99] |
| SpvD | SPI-1/SPI-2 | Inhibits NF-κB signaling | [87] |
| SrfJ | SPI-2 | Responses to intracellular conditions | [100] |
| SsaJ | SPI-2 | Prevents the phagocyte NADPH oxidase from trafficking toward SCVs | [101] |
| Ssav | SPI-2 | Prevents the phagocyte NADPH oxidase from trafficking toward SCVs | [102] |
| SseB | SPI-2 | Prevents the phagocyte NADPH oxidase from trafficking toward SCVs | [102] |
| SseF | SPI-2 | Tethers SCV to the Golgi network; contributes to Sif formation; replication of Salmonella in SCV | [87,103] |
| SseG | SPI-2 | Tethers SCV to the Golgi network; contributes to Sif formation; replication of Salmonella in SCV | [87,103] |
| SseJ | SPI-2 | Acyl transferase; cholesterol esterification; SCV membrane dynamics | [87,104] |
| SseK1 | SPI-2 | Inhibits TNFα-stimulated NF-κB signaling | [105] |
| SseK2 | SPI-2 | Related effectors that inhibits NF-κB signaling | [105] |
| SseL | SPI-2 | Inhibits autophagic clearance of cytosolic aggregates; induces late macrophage cell death; inhibits directional migration of macrophages and DCs | [106] |
| SspH2 | SPI-2 | An E3 ubiquitin ligase; activates NOD1 signaling | [79,87] |
| SsrA | SPI-2 | Prevents the phagocyte NADPH oxidase from trafficking toward SCVs | [102] |
| SteA | SPI-1/SPI-2 | SIF formation, vacuolar membrane partitioning | [107] |
| SteC | SPI-2 | Induces assembly of F-actin meshwork around SCV | [108] |
| SteD | SPI-2 | Inhibits antigen presentation and T cell activation | [17] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Qazi, I.H.; Wang, L.; Zhou, G.; Han, H. Salmonella Virulence and Immune Escape. Microorganisms 2020, 8, 407. https://doi.org/10.3390/microorganisms8030407
Wang M, Qazi IH, Wang L, Zhou G, Han H. Salmonella Virulence and Immune Escape. Microorganisms. 2020; 8(3):407. https://doi.org/10.3390/microorganisms8030407
Chicago/Turabian StyleWang, Mengyao, Izhar Hyder Qazi, Linli Wang, Guangbin Zhou, and Hongbing Han. 2020. "Salmonella Virulence and Immune Escape" Microorganisms 8, no. 3: 407. https://doi.org/10.3390/microorganisms8030407
APA StyleWang, M., Qazi, I. H., Wang, L., Zhou, G., & Han, H. (2020). Salmonella Virulence and Immune Escape. Microorganisms, 8(3), 407. https://doi.org/10.3390/microorganisms8030407