Inhibition of Nitric Oxide Production, Oxidative Stress Prevention, and Probiotic Activity of Lactic Acid Bacteria Isolated from the Human Vagina and Fermented Food

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Microorganisms
2.2. Cell Culture
2.3. Nitric Oxide Assays
2.4. Biochemical Tests and Identification
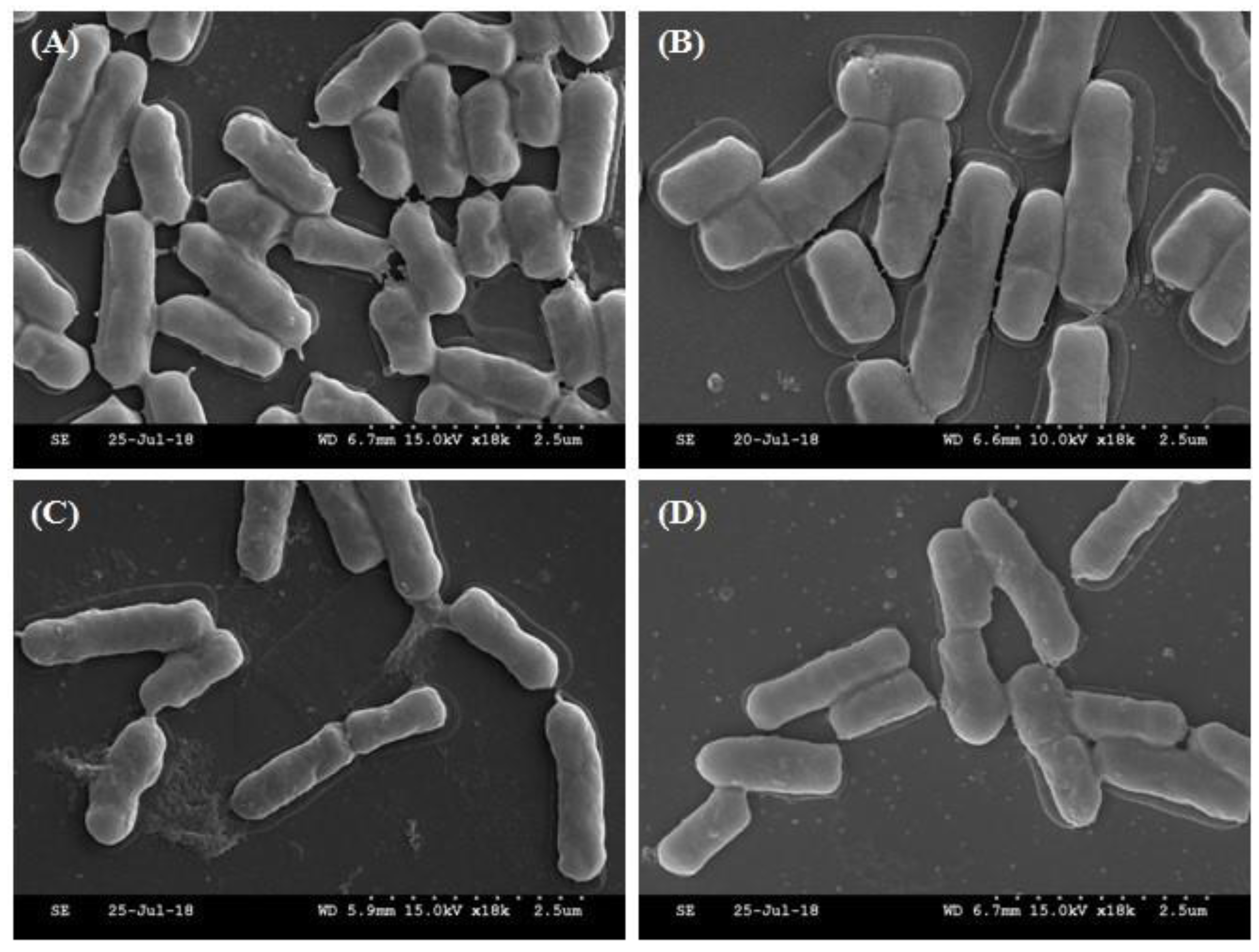
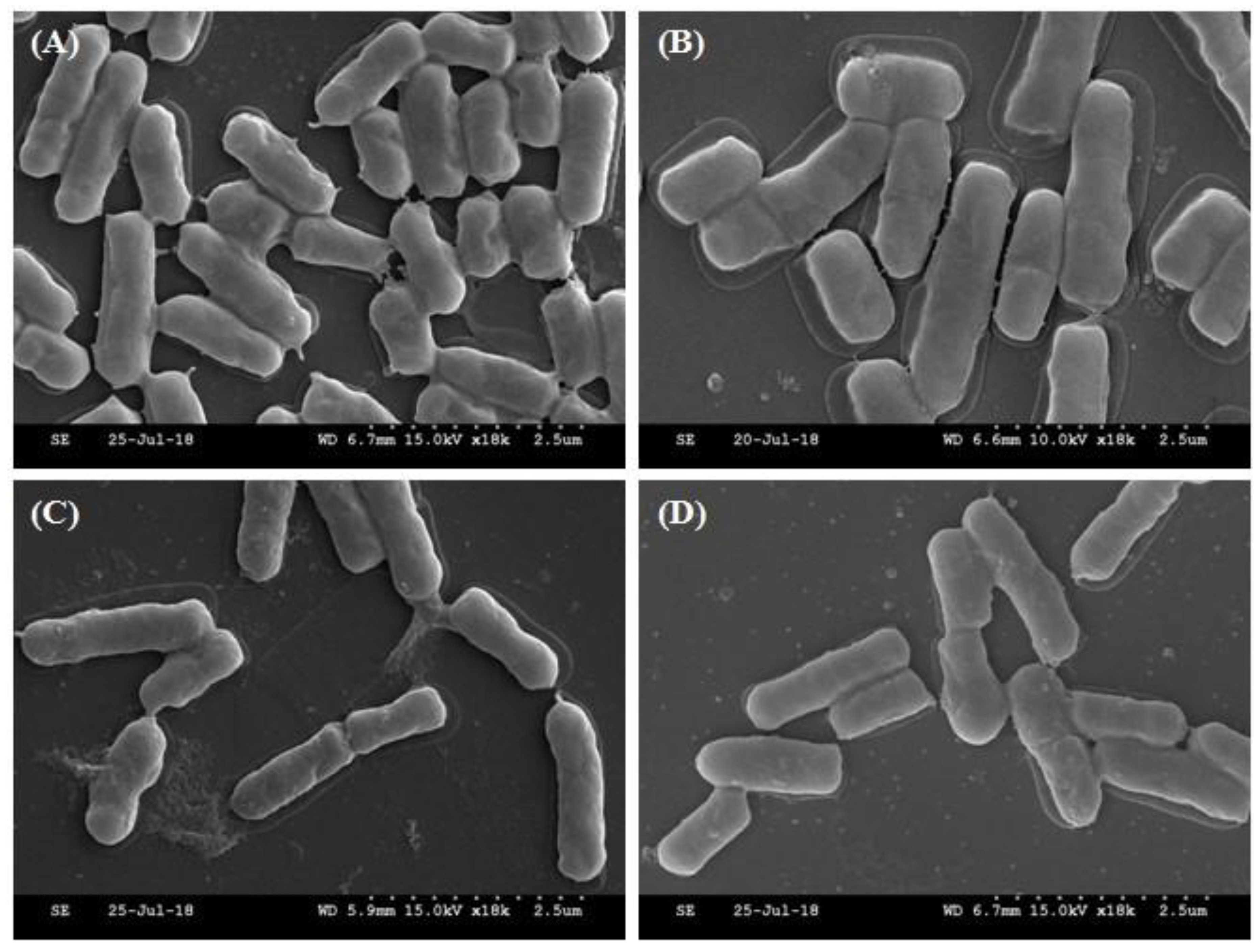
2.5. Morphological Analysis of Selected Strains by SEM
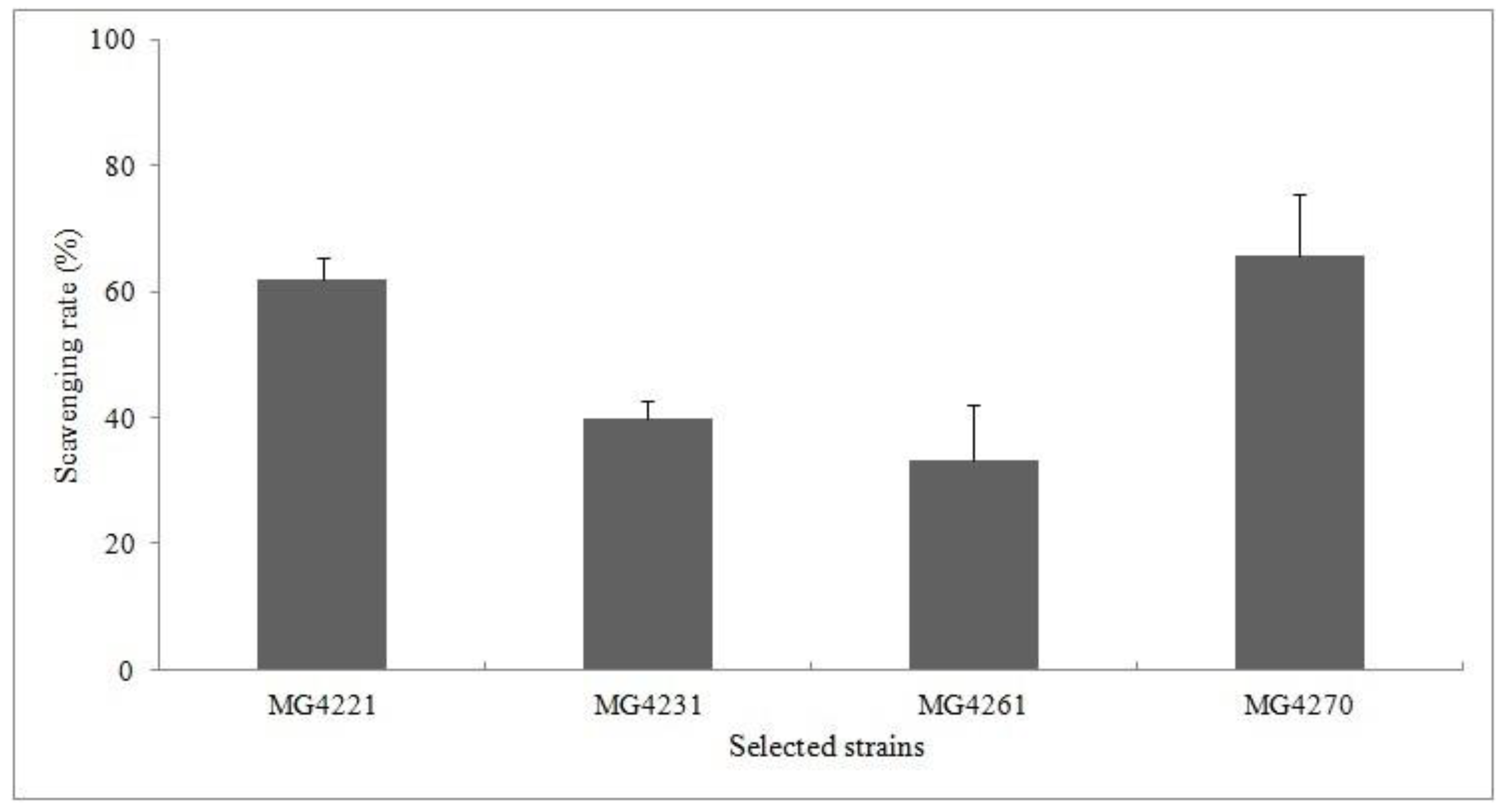
2.6. Measurement of 2,2-Diphenyl-1-Picryl-Hydrazyl (DPPH) Scavenging Activity
2.7. Strain Survival under Conditions Simulating the Human Gastrointestinal Tract
2.8. Strains Survival under Simulated Gastrointestinal Conditions
2.9. Antibiotic Sensitivity Test
2.10. Statistical Analysis
3. Results and Discussion
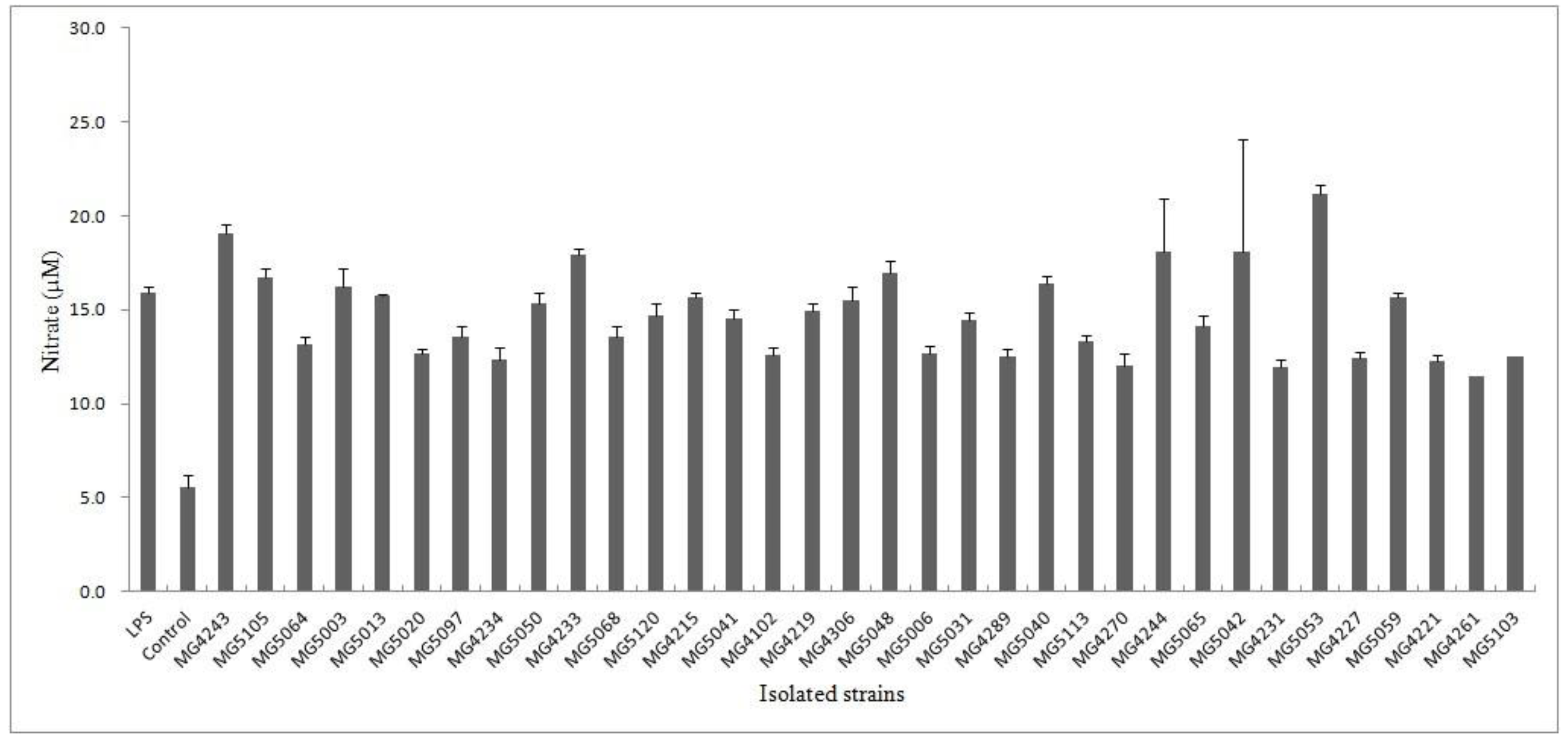
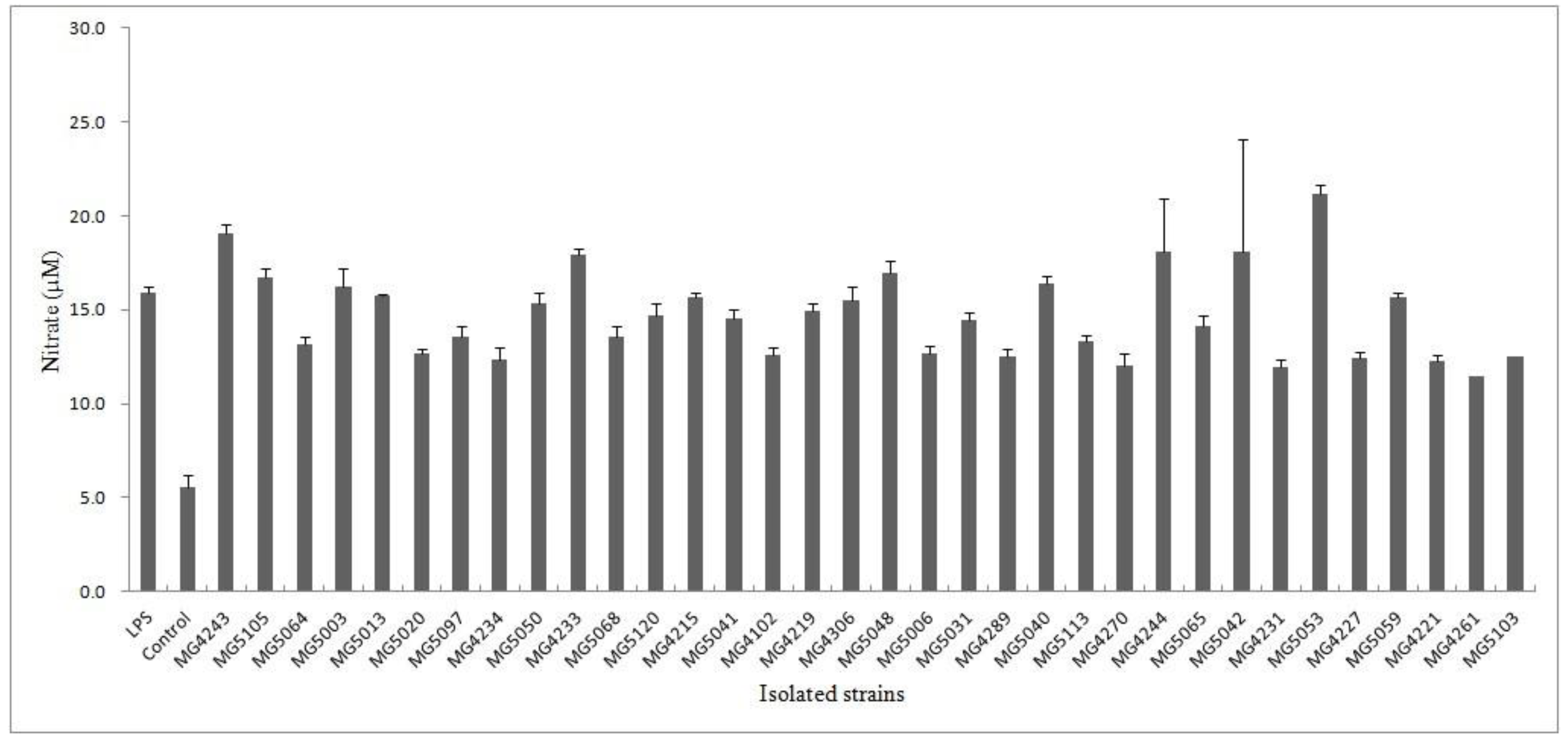
3.1. Inhibition of Isolated Strains on LPS-Induced NO Production
3.2. Identification of Selected Strains
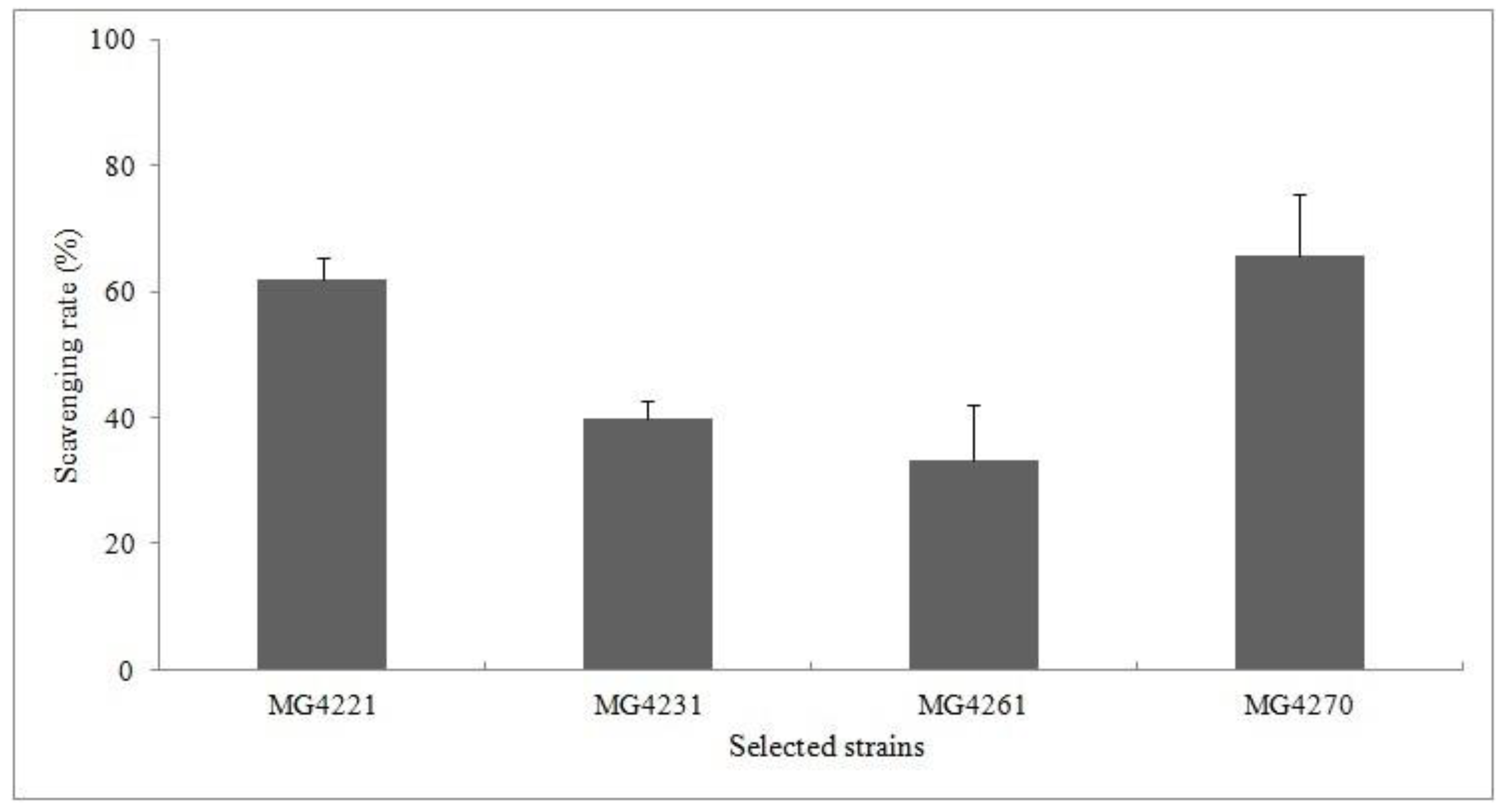
3.3. DPPH Scavenging Activity
3.4. Resistance to Simulated Gastrointestinal Juices
3.5. Antibiotic Sensitivity Test
Author Contributions
Funding
Conflicts of Interest
References
- Hawley, H.B.; Sheperd, P.A.; Wheather, D.M. Factors affection the implantation of lactobacilli in the intestine. J. Appl. Bacteriol. 1959, 22, 360–367. [Google Scholar]
- Klein, G.; Pack, A.; Bonaparte, C.; Reuter, G. Taxonomy and physiology of probiotic lactic acid bacteria. Int. J. Food Microbiol. 1998, 41, 103–125. [Google Scholar] [CrossRef]
- Schlundt, J. Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; Food and Agriculture Organization of the United Nations: Córdoba, Argentina, 2001; pp. 1–34. [Google Scholar]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Laursen, R.R.; Ouwehand, A.C. The production and delivery of probiotics: A review of a practical approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on the substantiation of health claims related to non-characterized microorganisms pursuant to Article 13 of Regulation (EC) No 1924/20061. EFSA J. 2009, 7, 1247. [Google Scholar] [CrossRef]
- EFSA. Guidance on the scientific. 462 requirements for health claims related to gut and immune function. EFSA J. 2011, 9, 1984. [Google Scholar] [CrossRef]
- Gill, H.S.; Guarner, F. Probiotics and human health: A clinical perspective. Postgrad. Med. J. 2004, 80, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Spahaak, S.; Havenaar, R.; Schaafsma, G. The effect of consumption of milk fermented by Lb. casei strain Shirota on the intestinal microflora and immune parameters in humans. Eur. J. Clin. Nutr. 1998, 52, 899–907. [Google Scholar] [CrossRef]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Clare, D.; Swaisgood, H. Bioactive milk peptides: A prospectus. J. Dairy Sci. 2000, 83, 1187–1195. [Google Scholar] [CrossRef]
- Khan, S.U. Probiotics in dairy foods: A review. Nutr. Food Sci. 2014, 44, 71–88. [Google Scholar] [CrossRef]
- Mishra, S.; Wang, S.; Nagpal, R.; Miller, B.; Singh, R.; Taraphder, S.; Yadav, H. Probiotics and prebiotics for the amelioration of type 1 diabetes: Present and future perspectives. Microorganisms 2019, 7, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; Anvari, S.; Anagnostou, K. The role of probiotics in preventing allergic disease. Children 2019, 6, 24. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Santos, J.A.; Bermúdez, M.G.; Campoy, C. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Burton, J.; Devillard, E. The rationale for probiotics in female urogenital healthcare. Med. Gen. Med. 2004, 6, 49–63. [Google Scholar]
- Mokoena, M.P. Lactic acid bacteria and their bacteriocins: Classification, biosynthesis and applications against uropathogens: A mini-review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Fares, C.; Longo, A.; Spano, G.; Capozzi, V. Lactobacillus plantarum with broad antifungal activity as a protective starter culture for bread production. Foods 2017, 6, 110. [Google Scholar] [CrossRef]
- Martin, R.; Soberon, N.; Vaneechoutte, M.; Florez, A.B.; Vazquez, F.; Suarez, J.E. Characterization of indigenous vaginal lactobacilli from healthy women as probiotic candidates. Int. Microbiol. 2008, 11, 261–266. [Google Scholar]
- Dimitonova, S.P.; Danova, S.T.; Serkedjieva, J.P.; Bakalov, B.V. Antimicrobial activity and protective properties of vaginal lactobacilli from healthy Bulgarian women. Anaerobe 2007, 13, 178–184. [Google Scholar] [CrossRef]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous food fermentations and potential risks for human health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Ahmend, A.A. In vitro screening of Lactobacillus species from homemade yogurt for antagonistic effects against common bacterial pathogens. Jordan J. Biol. Sci. 2013, 6, 211–216. [Google Scholar] [CrossRef]
- Chen, Y.S.; Liou, M.S.; Ji, S.H.; Yu, C.R.; Pan, S.F.; Yanagida, F. Isolation and characterization of lactic acid bacteria from Yantsai-shin (Fermented broccoli stems), a traditional fermented food in Taiwan. J. Appl. Microbiol. 2013, 115, 125–132. [Google Scholar] [CrossRef]
- Kalui, C.M.; Mathara, J.M.; Kutima, P.M. Probiotic potential of spontaneously fermented cereal based foods—A review. Afr. J. Biotechnol. 2010, 9, 2490–2498. [Google Scholar]
- Vincenzo, B.; Asif, I.J.; Nikolaos, P.; Francesco, M. Adaptive immunity and inflammation. Int. J. Inflam. 2015. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, A. The L-arginine-nitric oxide pathway. N. Engl. J. Med. 1993, 329, 2002–2012. [Google Scholar]
- Bredt, D.D.; Snydr, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682–685. [Google Scholar] [CrossRef]
- Chang, C.E.; Pavlova, S.I.; Tao, L.; Kim, E.; Kim, S.C.; Yun, H.S.; So, J.S. Molecular identification of vaginal Lactobacillus spp. isolated from Korean women. J. Microbiol. Biotechnol. 2002, 12, 312–317. [Google Scholar]
- Lyons, C.R.; Orloff, G.J.; Cunninggham, J.M. Molecular cloning and functional expression of an inducible nitric oxide synthase from a murine macrophage cell line. J. Biol. Chem. 1992, 9, 6370–6374. [Google Scholar]
- Claus, D. A standardized gram staining procedure. World J. Microb. Biot. 1992, 8, 451–452. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Jeung, M.K.; Park, M.H.; Lee, S.Y.; Lee, J.H. Effects of roasting conditions of sesame seeds on the oxidative stability of pressed oil during thermal oxidation. Food Chem. 2010, 118, 681–685. [Google Scholar] [CrossRef]
- Maragkoudakis, P.A.; Zoumpopoulou, G.; Miaris, C.; Kalantzopoulos, G.; Pot, B.; Tsakalidou, E. Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J. 2006, 6, 189–199. [Google Scholar] [CrossRef]
- Ga¨nzle, M.G.; Hertel, C.; van der Vossen, J.M.; Hammes, W.P. Effect of bacteriocin-producing lactobacilli on the survival of Escherichia coli and Listeria in a dynamic model of the stomach and the small intestine. Int. J. Food Microbiol. 1999, 48, 21–35. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard, 8th ed.; National Committee for Clinical Laboratory Standards Document M2–A8: Wayne, PA, USA, 2003. [Google Scholar]
- Lin, Y.L.; Lin, J.K. (−)-Epigallocatechin-3-gallate blocks the induction of nitric oxide systhase by down-regulating lipopolysaccharide-induced activity of transcription factor nuclear factor-kB. Mol. Pharmacol. 1997, 52, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Morita, H.; Yoshikawa, H.; Sakata, R.; Nagata, Y.; Tanaka, H. Synthesis of nitric oxide from the two equivalent guanidino nitrogens of L-arginine by Lactobacillus fermentum. J. Bacteriol. 1997, 79, 7812–7815. [Google Scholar] [CrossRef]
- Hugas, M.; Monfort, J.M. Bacterial starter cultures for meat fermentation. Food Chem. 1997, 59, 547–554. [Google Scholar] [CrossRef]
- Capozzi, V.; Weidmann, S.; Fiocco, D.; Reiu, A.; Hols, P.; Guzzo, J.; Spano, G. Inactivation of a small heat shock protein affects cell morphology and membrane fluidity in Lactobacillus plantarum WCFS1. Res. Microbiol. 2011, 162, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Fiocco, D.; Longo, A.; Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. How probiotics face food stress: They get by with a little help. Crit. Rev. Food Sci. Nutr. 2019. [Google Scholar] [CrossRef]
- Ji, K.; Jang, N.Y.; Kim, Y.T. Isolation of lactic acid bacteria showing antioxidative and probiotic activities from kimchi and infant feces. J. Microbiol. Biotechnol. 2015, 25, 1568–1577. [Google Scholar] [CrossRef]
- Morelli, L. In vitro assessment of probiotic bacteria: From survival to functionality. Int. Dairy J. 2007, 17, 1278–1283. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S. Bile salt and acid tolerance of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. Fems. Microbiol. 2005, 44, 129–137. [Google Scholar] [CrossRef]
- Dixit, G.; Samarth, D.; Tale, V.; Bhadekar, R. Comparative studies on potential probiotic characteristics of Lactobacillus acidophilus strains. Eur. J. Biosci. 2013, 7, 1–9. [Google Scholar] [CrossRef]
- Shivram, P.L.; Vishwanath, P.P. Assessment of probiotic potential of Lactobacillus sp. isolated from cheese and preparation of probiotic ice cream. Int. J. Res. Ayurveda Pharm. 2012, 3, 532–536. [Google Scholar]
- Liu, C.; Zhang, Z.Y.; Dong, K.; Yuan, J.P.; Guo, X.K. Antibiotic resistance of probiotic strains of lactic acid bacteria isolated from marketed foods and drugs. Biomed. Environ. Sci. 2009, 22, 401–412. [Google Scholar] [CrossRef]
- Saarela, M.; Mogensen, G.; Fonden, R.; Matto, J.; Mattila-Sandholm, T. Probiotic bacteria: Safety, functional and technological properties. J. Bacteriol. 2000, 84, 197–215. [Google Scholar] [CrossRef]
- Kirtzalidou, E.; Pramateftaki, P.; Kotsou, M.; Kyriacou, A. Screening for lactobacilli with probiotic properties in the infant gut microbiota. Anaerobe 2011, 17, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Ammor, M.S.; Belen Florezz, A.; Mayo, B. Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 2007, 24, 559–570. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selected Strains | Simulated Gastric Fluid (1) | Simulated Intestinal Fluid (2) | |||
|---|---|---|---|---|---|
| pH2 | pH3 | pH4 | pH7 | pH8 | |
| MG4221 | 4.20 ± 0.03 | 6.86 ± 0.06 | 7.91 ± 0.06 | 9.01 ± 0.08 | 9.02 ± 0.04 |
| MG4231 | 3.17 ± 0.02 | 6.02 ± 0.11 | 6.92 ± 0.07 | 9.04 ± 0.04 | 8.90 ± 0.01 |
| MG4261 | 3.46 ± 0.07 | 6.73 ± 0.07 | 6.91 ± 0.08 | 7.98 ± 0.02 | 7.99 ± 0.05 |
| MG4270 | 3.00 ± 0.02 | 5.92 ± 0.03 | 7.65 ± 0.04 | 9.00 ± 0.11 | 8.98 ± 0.06 |
| Antibiotics (µg/disc) | Selected Strains | |||
|---|---|---|---|---|
| MG4221 | MG4231 | MG4261 | MG4270 | |
| Sulphamethoxazole-trimethoprim (1.25/23.75) | R | S | S | R |
| Tetracyclin (30) | I | S | S | S |
| Cephalothin (30) | S | S | S | S |
| Gentamicin (10) | S | I | R | S |
| Erythromycin (15) | S | S | S | S |
| Vancomycin (30) | R | R | R | R |
| Ampicillin (10) | S | S | S | S |
| Rifampicin (5) | S | S | S | S |
| Ciprofloxacin (5) | R | I | R | I |
| Cefotaxime (30) | S | S | S | S |
| Cefepime (30) | S | R | S | S |
| Cefotetan (30) | R | R | R | R |
| Nalidixic acid (30) | R | R | R | R |
| Kanamycin (30) | I | I | I | S |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, C.-H.; Han, S.H.; Kim, J.-S.; Kim, Y.; Jeong, Y.; Park, H.M.; Paek, N.-S. Inhibition of Nitric Oxide Production, Oxidative Stress Prevention, and Probiotic Activity of Lactic Acid Bacteria Isolated from the Human Vagina and Fermented Food. Microorganisms 2019, 7, 109. https://doi.org/10.3390/microorganisms7040109
Kang C-H, Han SH, Kim J-S, Kim Y, Jeong Y, Park HM, Paek N-S. Inhibition of Nitric Oxide Production, Oxidative Stress Prevention, and Probiotic Activity of Lactic Acid Bacteria Isolated from the Human Vagina and Fermented Food. Microorganisms. 2019; 7(4):109. https://doi.org/10.3390/microorganisms7040109
Chicago/Turabian StyleKang, Chang-Ho, Seul Hwa Han, Jin-Seong Kim, YongGyeong Kim, Yulah Jeong, Hye Min Park, and Nam-Soo Paek. 2019. "Inhibition of Nitric Oxide Production, Oxidative Stress Prevention, and Probiotic Activity of Lactic Acid Bacteria Isolated from the Human Vagina and Fermented Food" Microorganisms 7, no. 4: 109. https://doi.org/10.3390/microorganisms7040109
APA StyleKang, C.-H., Han, S. H., Kim, J.-S., Kim, Y., Jeong, Y., Park, H. M., & Paek, N.-S. (2019). Inhibition of Nitric Oxide Production, Oxidative Stress Prevention, and Probiotic Activity of Lactic Acid Bacteria Isolated from the Human Vagina and Fermented Food. Microorganisms, 7(4), 109. https://doi.org/10.3390/microorganisms7040109