Lysis Profiles of Salmonella Phages on Salmonella Isolates from Various Sources and Efficiency of a Phage Cocktail against S. Enteritidis and S. Typhimurium


 and
and
Abstract
: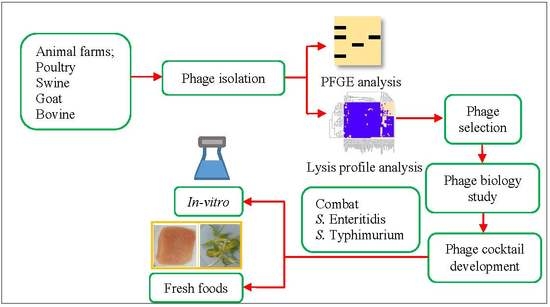
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Bacterial Strains Used in the Study
2.3. Isolation and Purification of Salmonella Phages
2.4. Lysate Preparation and Titer Determination of Salmonella Phages
2.5. Determination of Lysis Profiles of Salmonella Phages
2.6. Genome Size Determination of Salmonella Phages
2.7. Development of a Phage Cocktail Targeting Two Major Salmonella Serovars
2.7.1. Spotting Assay
2.7.2. Efficiency of Plating (EOP) Assay
2.8. One-Step Growth Curve
2.9. Efficiency of Phage Cocktail to Reduce S. Enteritidis and S. Typhimurium In Vitro and Evaluation of Phage-Resistance in Salmonella after Treated with Phage Cocktail
2.10. Efficiency of Phage Cocktail to Reduce S. Enteritidis and S. Typhimurium in Foods
2.11. Statistical Analysis
3. Results
3.1. Recovery of Salmonella Phages from Various Animal Farms in Thailand
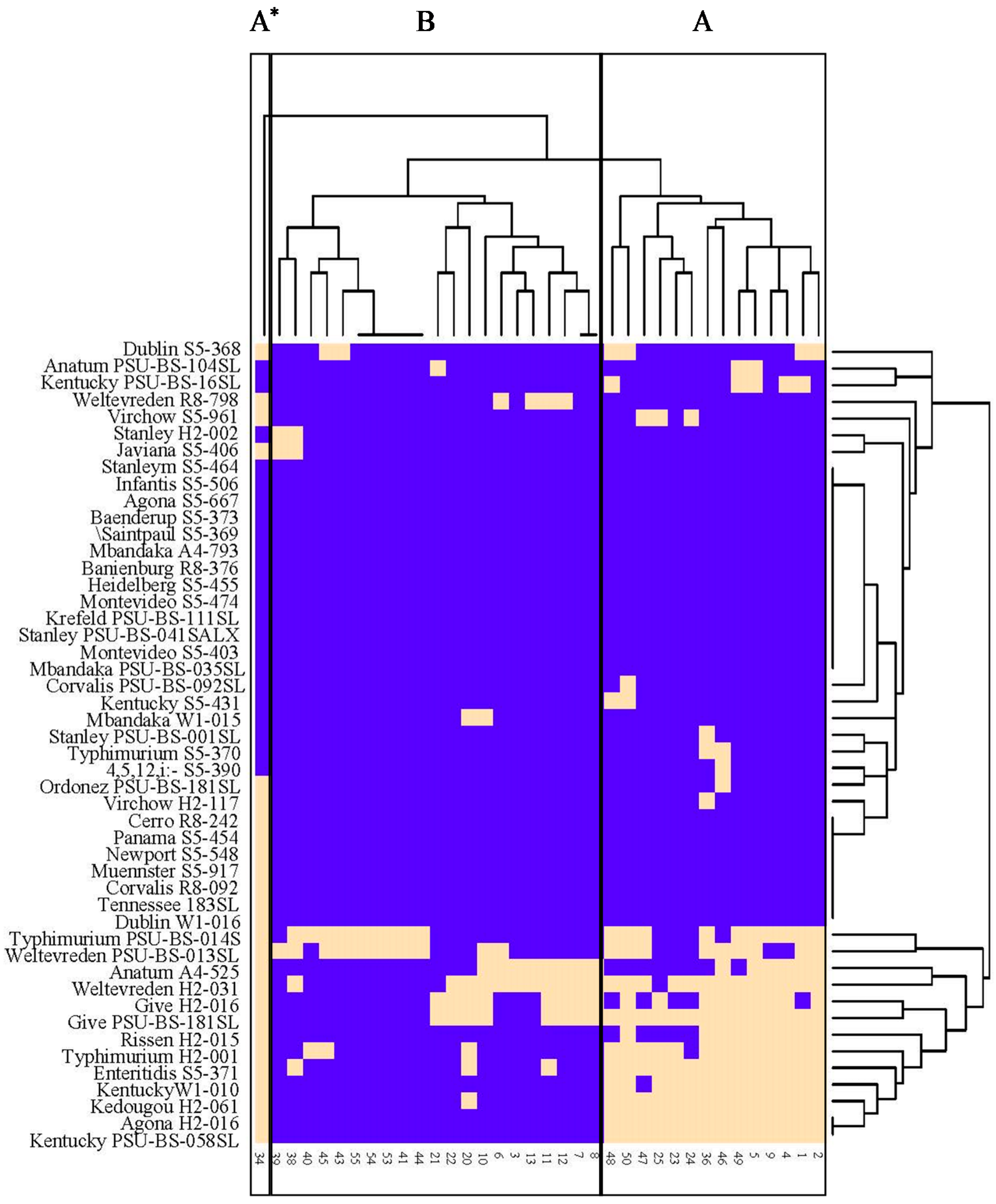
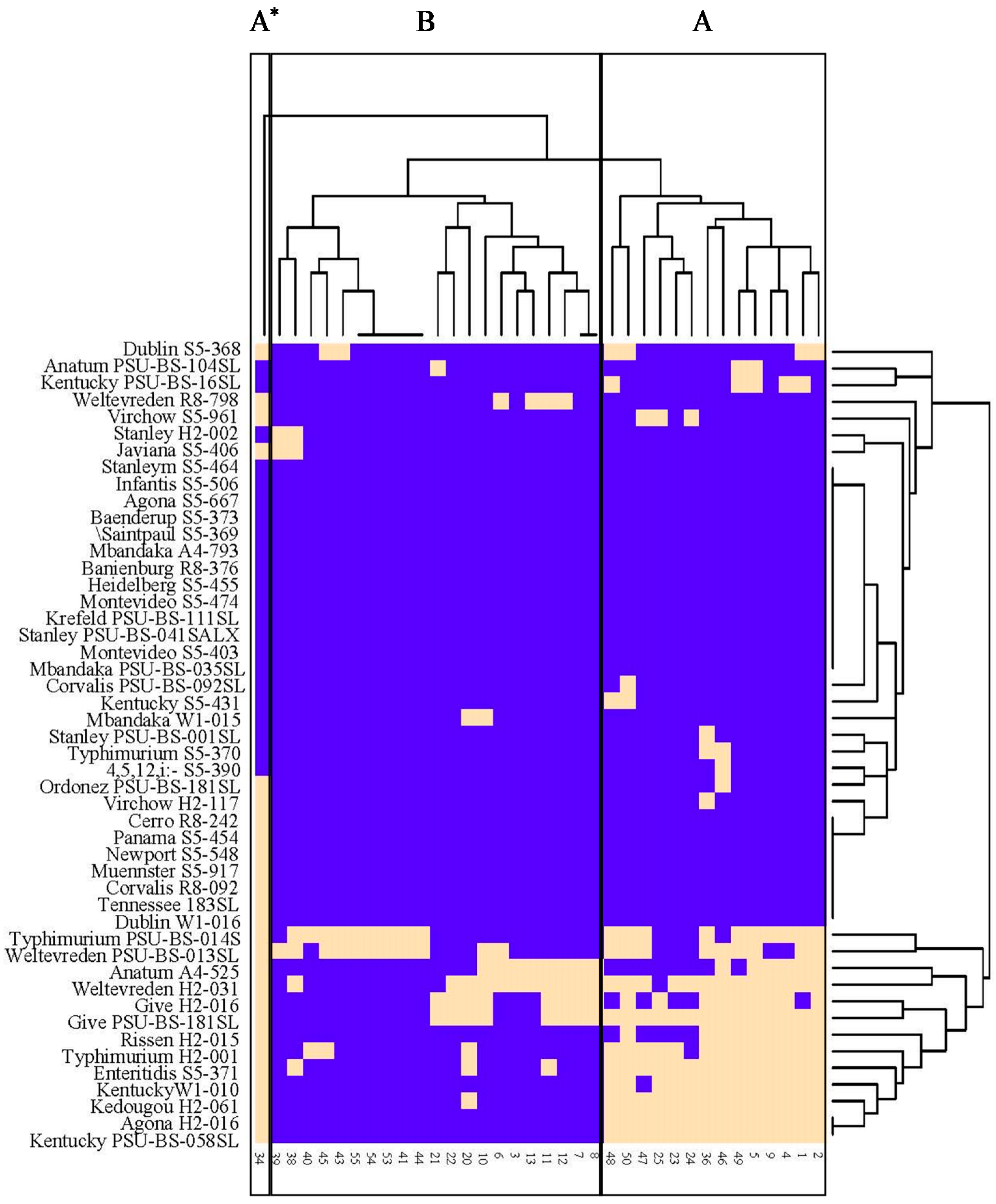
3.2. Lysis Profiles of Salmonella Phages from Animal Farms on Salmonella Strains from Various Sources in Thailand and USA
3.3. Phage Susceptibility of Different Salmonella Serovars from Various Sources
3.4. Development of a Phage Cocktail
3.5. Efficiency of Salmonella Phage Cocktail in Reducing S. Enteritidis and S. Typhimurium In Vitro and Different Fresh Foods, and Evidence of Phage-Resistant Development in Salmonella
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- García, V.; Mandomando, I.; Ruiz, J.; Herrera-León, S.; Alonso, P.L.; Rodicio, M.R. Salmonella enterica serovars Typhimurium and Enteritidis causing mixed infections in febrile children in Mozambique. Infect. Drug Resist. 2018, 11, 195–204. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention. Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 23 October 2018).
- Cummings, K.J.; Warnick, L.D.; Alexander, K.A.; Cripps, C.J.; Gröhn, Y.T.; James, K.L.; McDonough, P.L.; Reed, K.E. The duration of fecal Salmonella shedding following clinical disease among dairy cattle in the northeastern USA. Prev. Vet. Med. 2009, 92, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Moreno Switt, A.I.; den Bakker, H.C.; Vongkamjan, K.; Hoelzer, K.; Warnick, L.D.; Cummings, K.J.; Wiedmann, M. Salmonella bacteriophage diversity reflects host diversity on dairy farms. Food Microbiol. 2013, 36, 275–285. [Google Scholar]
- Shahzad, A.; Mahmood, M.Sh.; Hussain, I.; Siddique, F.; Abbas, R.Z. Prevalence of Salmonella species in hen eggs and egg storing-trays collected from poultry farms and marketing outlets of Faisalabad, Pakistan. Pak. J. Agric. Sci. 2012, 49, 565–568. [Google Scholar]
- United States Department of Agriculture. Serotypes Profile of Salmonella Isolates from Meat and Poultry Products January 1998 through December 2014. Available online: https://www.fsis.usda.gov/wps/wcm/connect/3866026a-582d-4f0e-a8ce-851b39c7390f/Salmonella-Serotype-Annual-2014.pdf?MOD=AJPERES (accessed on 23 October 2018).
- Reddy, S.P.; Wang, H.; Adams, J.K.; Feng, P.C.H. Prevalence and characteristics of Salmonella serotypes isolated from fresh produce marketed in the United States. J. Food Prot. 2016, 79, 6–16. [Google Scholar] [CrossRef]
- Ponce, E.; Khan, A.; Cheng, C.M.; Summage-West, C.; Cerniglia, C.E. Prevalence and characterization of Salmonella enterica serovar Weltevreden from imported seafood. Food Microbiol. 2008, 25, 29–35. [Google Scholar] [CrossRef]
- Shabarinath, S.; Sanath Kumar, H.; Khushiramani, R.; Karunasagar, I.; Karunasagar, I. Detection and characterization of Salmonella associated with tropical seafood. Int. J. Food Microbiol. 2007, 114, 227–233. [Google Scholar] [CrossRef]
- Espigares, E.; Bueno, A.; Espigares, M.; Gálvez, R. Isolation of Salmonella serotypes in wastewater and effluent: Effect of treatment and potential risk. Int. J. Hyg. Environ. Health 2006, 209, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Bangtrakulnonth, A.; Pornreongwong, S.; Pulsrikarn, Ch.; Sawanpanyalert, P.; Hendriksen, S.R.; Wong, D.; Arestrupt, F.M. Salmonella serovars from humans and other sources in Thailand 1993–2003. Emerg. Infect. Dis. 2004, 10, 131–135. [Google Scholar]
- Center for Disease Control and Prevention. Reports of Salmonella Outbreak Investigations from 2018. Available online: https://www.cdc.gov/salmonella/outbreaks-2018.html (accessed on 23 October 2018).
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis outbreaks in the United States due to fresh produce: Sources and potential intervention measures. Food Pathog. Dis. 2009, 6, 635–648. [Google Scholar] [CrossRef]
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Muñoz, S.L.; Koskella, B. Bacteria–phage interactions in natural environments. Adv. Appl. Microbiol. 2014, 89, 135–183. [Google Scholar]
- Kazi, M.; Annapure, U.S. Bacteriophage biocontrol of foodborne pathogens. J. Food Sci. Tech. 2016, 53, 1355–1362. [Google Scholar] [CrossRef]
- Fister, S.; Robben, C.; Witte, A.K.; Schoder, D.; Wagner, M.; Rossmanith, P. Influence of environmental factors on phage-bacteria interaction and on the efficacy and infectivity of phage P100. Front. Microbiol. 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Thung, T.T.; Premarathne, K.; Chang, W.S.; Loo, Y.Y.; Chin, Y. Zh.; Kuan, Ch.H.; Tan, Ch.W.; Basri, D.F.; Jasimah, Ch.W.; Radu, S. Use of a lytic bacteriophage to control Salmonella Enteritidis in retail food. LWT-Food Sci. Technol. 2017, 78, 222–225. [Google Scholar] [CrossRef]
- Kocharunchitt, C.; Ross, T.; McNeil, D.L. Use of bacteriophages as biocontrol agents to control Salmonella associated with seed sprouts. Int. J. Food. Microbiol. 2009, 128, 453–459. [Google Scholar] [CrossRef]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a bacteriophage cocktail to control Salmonella in food and the food industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Padungtod, P.; Kaneene, J.B. Salmonella in food animals and humans in northern Thailand. Int. J. Food Microbiol. 2006, 108, 346–354. [Google Scholar] [CrossRef]
- Vongkamjan, K.; Moreno Switt, A.I.; Den, H.C.; Fortes, E.D.; Wiedmann, M. Silage collected from dairy farms harbors an abundance of Listeria phages with considerable host range and genome size. Appl. Environ. Microbiol. 2012, 78, 8666–8675. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http:// www.R.-project.org/ (accessed on 20 January 2019).
- Mirzaei, M.K.; Nilsson, A.S. Correction: isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0127606. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, H.; Wang, R. Isolation and characterization of bacteriophages of Salmonella enterica serovar Pullorum. Poult. Sci. 2011, 90, 2370–2377. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations and World Health Organization. Joint FAO/WHO Food Standards Programme Codex Committee on Fish and Fishery Products, Discussion Paper on the Use of Chlorinated Water. 2000. CX/FFP 00/13. Available online: http://www.fao.org/tempref/codex/Meetings/CCCF/CCCF5/cf05_INF.pdf (accessed on 23 October 2018).
- Hungaro, M.H.; Santos Mendonca, R.C.; Gouvea, D.M.; Danatas Vanetti, M.C.; de Oliveira Pinto, C.L. Use of bacteriophages to reduce Salmonella in chicken skin in comparison with chemical agents. Food Res. Int. 2013, 52, 75–81. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Brabban, A.; Kutter, E.; Karriker, L.; Stahl, C.; Wagstrom, E.; Anderson, R.C.; Genovese, K.; McReynolds, J.; et al. Occurrence of Salmonella-specific bacteriophages in swine feces collected from commercial farms. Foodborne Pathog. Dis. 2010, 7, 851e856. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.R.; Balaa, M.F.; Sims, J.; King, R. Isolation of Salmonella bacteriophages from swine effluent lagoons. J. Environ. Qual. 2006, 35, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Shukla, K.S.; Hirpurkar, S.D.; Singh, S.K.; Rajoria, R. Isolation of phage from animal waste of different LSF and their utility in phage therapy. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 205–210. [Google Scholar]
- Wongsuntornpoj, S.; Moreno Switt, A.I.; Bergholz, P.; Wiedmann, M.; Chaturongakul, S. Salmonella phages isolated from dairy farms in Thailand show wider host range than a comparable set of phages isolated from U.S. dairy farms. Vet. Microbiol. 2014, 172, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Chibani-Chennoufi, S.; Bruttin, A.; Dillmann, M.L.; Brűssow, H. Phage-host interaction: An ecological perspective. J. Bacteriol. 2004, 186, 3677–3686. [Google Scholar] [CrossRef]
- Bailey, J.S.; Cosby, D.E. Salmonella prevalence in free-range and certified organic chickens. J. Food Prot. 2005, 68, 245–2453. [Google Scholar] [CrossRef]
- Chaturongakul, S.; Ounjai, P. Phage-host interplay: Examples from tailed phages and Gram-negative bacterial pathogens. Front. Microbiol. 2014, 5, 442. [Google Scholar] [CrossRef] [PubMed]
- Saengthongpinit, C.; Nane-Siri, D.; Aparachita, P.; Apiwannarat, P.; Buakhao, P.; Bowornnantiwath, W.; Thengchaisri, N.; Songserm, T. Longitudinal study of Salmonella and Campylobacter species from two laying duckling flocks in the central region of Thailand. Thai J. Vet. Med. 2014, 44, 355–361. [Google Scholar]
- Vaeteewootacharn, K.; Sutra, S.; Vaeteewootacharn, S.; Sithigon, D.; Jamjane, O.; Chomvarin, C.; Hahnvajanawong, C.; Thongskulpanich, N.; Thaewnon-giew, K. Salmonellosis and the food chain in Khon Kaen, northeastern Thailand. Southeast Asian J. Trop. Med. Public Health 2005, 36, 123–129. [Google Scholar]
- Hooton, S.P.; Atterbury, R.J.; Connerton, I.F. Application of a bacteriophage cocktail to reduce Salmonella Typhimurium U288 contamination on pig skin. Int. J. Food Microbiol. 2011, 151, 157–163. [Google Scholar] [CrossRef]
- Lee, J.H.; Shin, H.; Kim, H.; Ryu, S. Complete genome sequence of Salmonella bacteriophage SPN3US. J. Virol. 2011, 85, 13470–13471. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, J.H.; Lee, B.K.; Kwon, H.J.; Shin, J.H.; Kim, J.; Kim, S. Complete genome sequence of Salmonella bacteriophage SS3e. J. Virol. 2012, 86, 10253–10254. [Google Scholar] [CrossRef]
- Paradiso, R.; Orsini, M.; Bolletti Censi, S.; Borriello, G.; Galiero, G. Complete genome sequence of a Myoviridae bacteriophage infecting Salmonella enterica serovar Typhimurium. Genome Announc. 2016, 4, e00940-16. [Google Scholar] [CrossRef]
- Turner, D.; Hezwani, M.; Nelson, S.; Salisbury, V.; Reynolds, D. Characterization of the Salmonella bacteriophage vB_SenS-Ent1. J. Gen. Virol. 2012, 93, 2046–2056. [Google Scholar] [CrossRef]
- Santos, S.B.; Carvalho, C.; Azeredo, J.; Ferreira, E.C. Population dynamics of a Salmonella lytic phage and its host: Implications of the host bacterial growth rate in modelling. PLoS ONE 2014, 9, e102507. [Google Scholar] [CrossRef]
- Calsina, A.; Palmada, J.-M.; Ripoll, J. Optimal latent period in a bacteriophage population model structured by infection-age. Math. Models Methods Appl. Sci. 2011, 21, 1–26. [Google Scholar] [CrossRef]
- Abedon, S.T.; Hyman, P.; Thomas, C. Experimental examination of bacteriophage latent-period evolution as a response to bacterial availability. Appl. Environ. Microbiol. 2003, 69, 7499–7506. [Google Scholar] [CrossRef]
- Wommack, E.K.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef]
- Cao, F.; Wang, X.; Wang, L.; Li, Z.; Che, J.; Wang, L.; Li, X.; Cao, Z.; Zhang, J.; Jin, L.; Xu, Y. Evaluation of the efficacy of a bacteriophage in the treatment of Pneumonia induced by multidrug resistance Klebsiella Pneumoniae in mice. BioMed Res. Int. 2015, 2015, 752930. [Google Scholar]
- Kalatzis, P.G.; Bastías, R.; Kokkari, C.; Katharios, P. Isolation and characterization of two lytic bacteriophages, φSt2 and φGrn1; phage therapy application for biological control of Vibrio alginolyticus in aquaculture live feeds. PLoS ONE 2016, 11, e0151101. [Google Scholar] [CrossRef] [PubMed]
- Grant, A.; Salina, P.; Jurgen, S.; Fawzy, H.; Bob, V. Reduction of Salmonella in ground chicken using a bacteriophage. Poult. Sci. 2017, 96, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
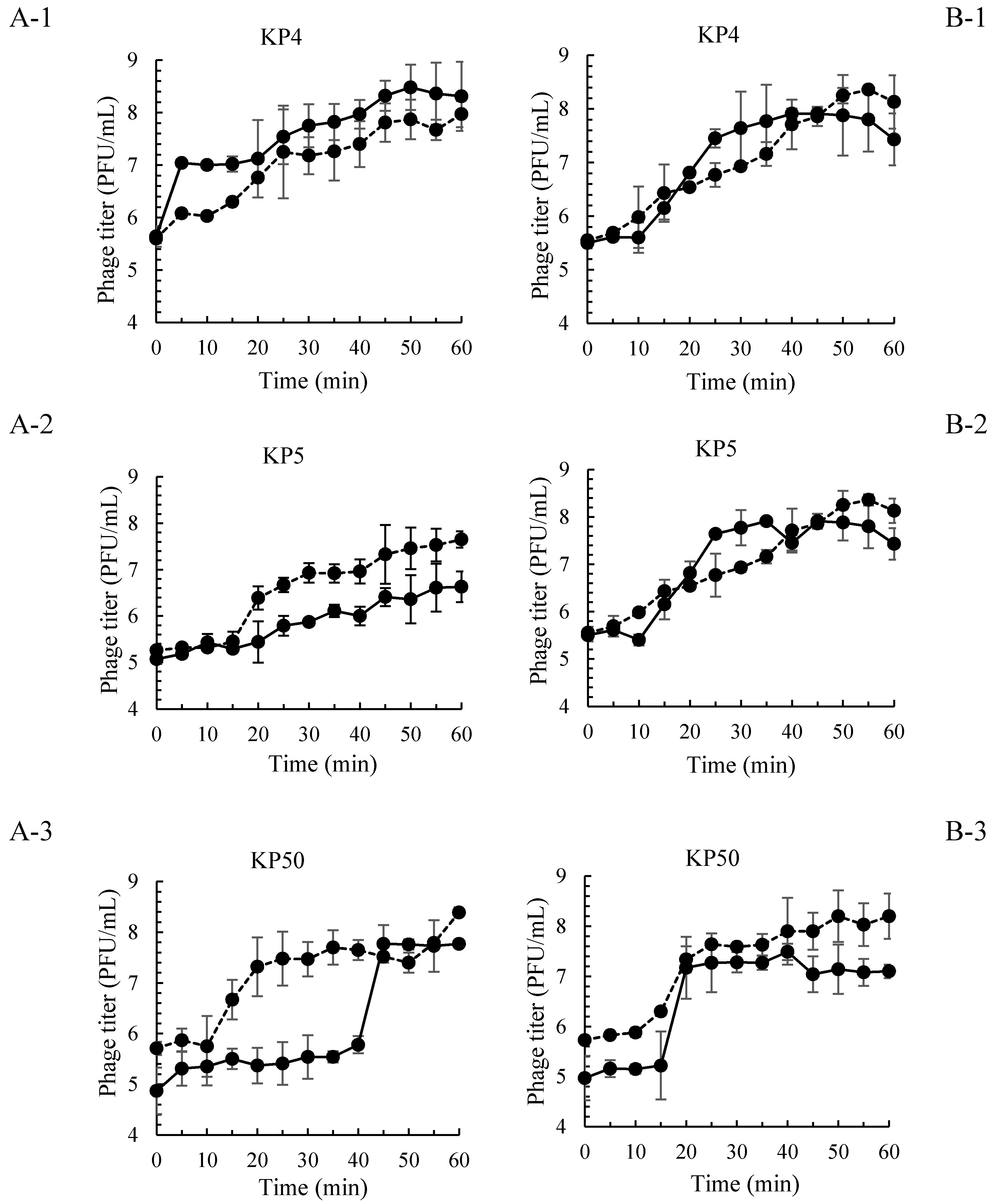
 and MOI 10 presented by
and MOI 10 presented by  . Bars represent the mean standard deviation (n = 3).
and MOI 10 presented by . Bars represent the mean standard deviation (n = 3).
. Bars represent the mean standard deviation (n = 3).
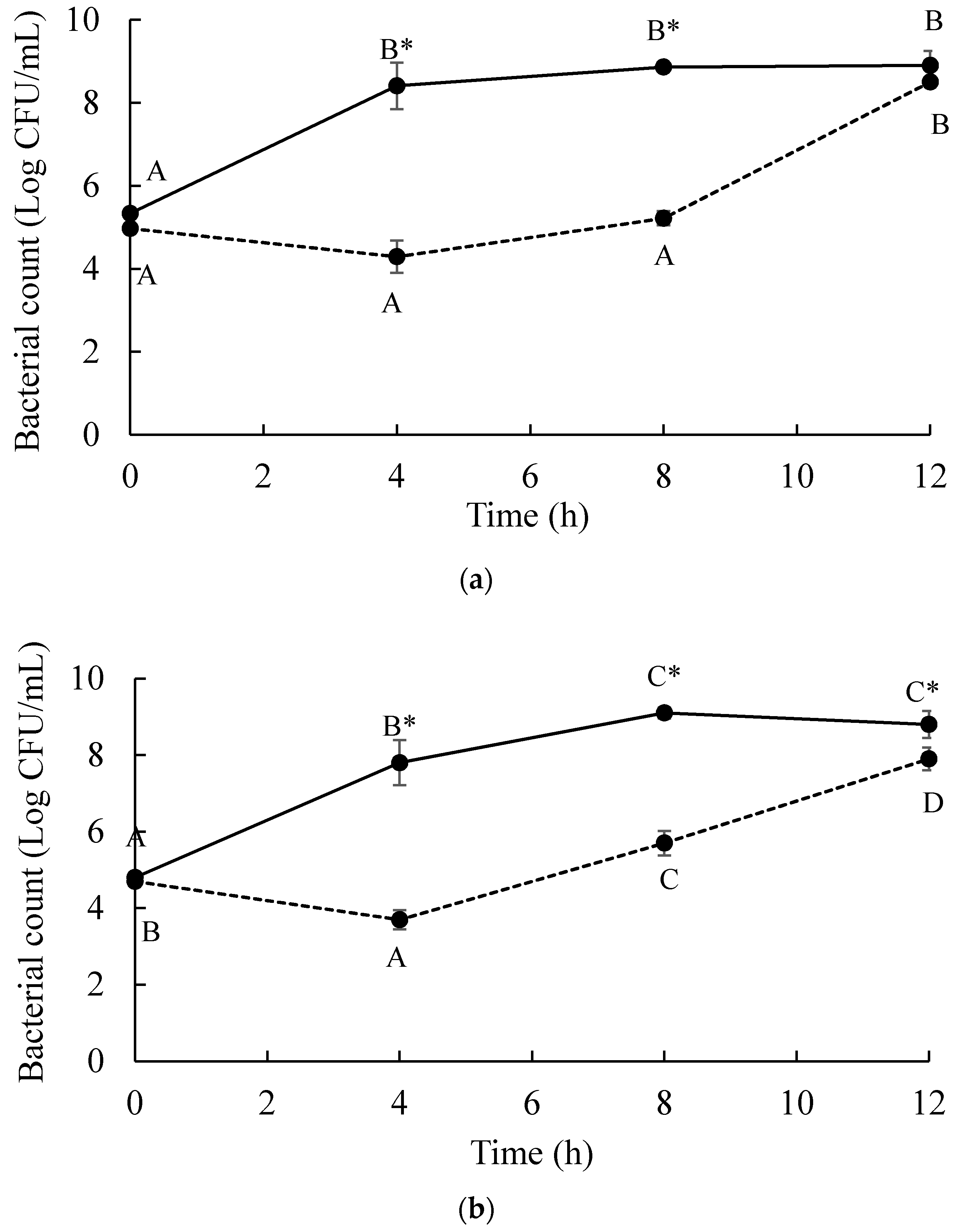
and MOI 10 presented by . Bars represent the mean standard deviation (n = 3).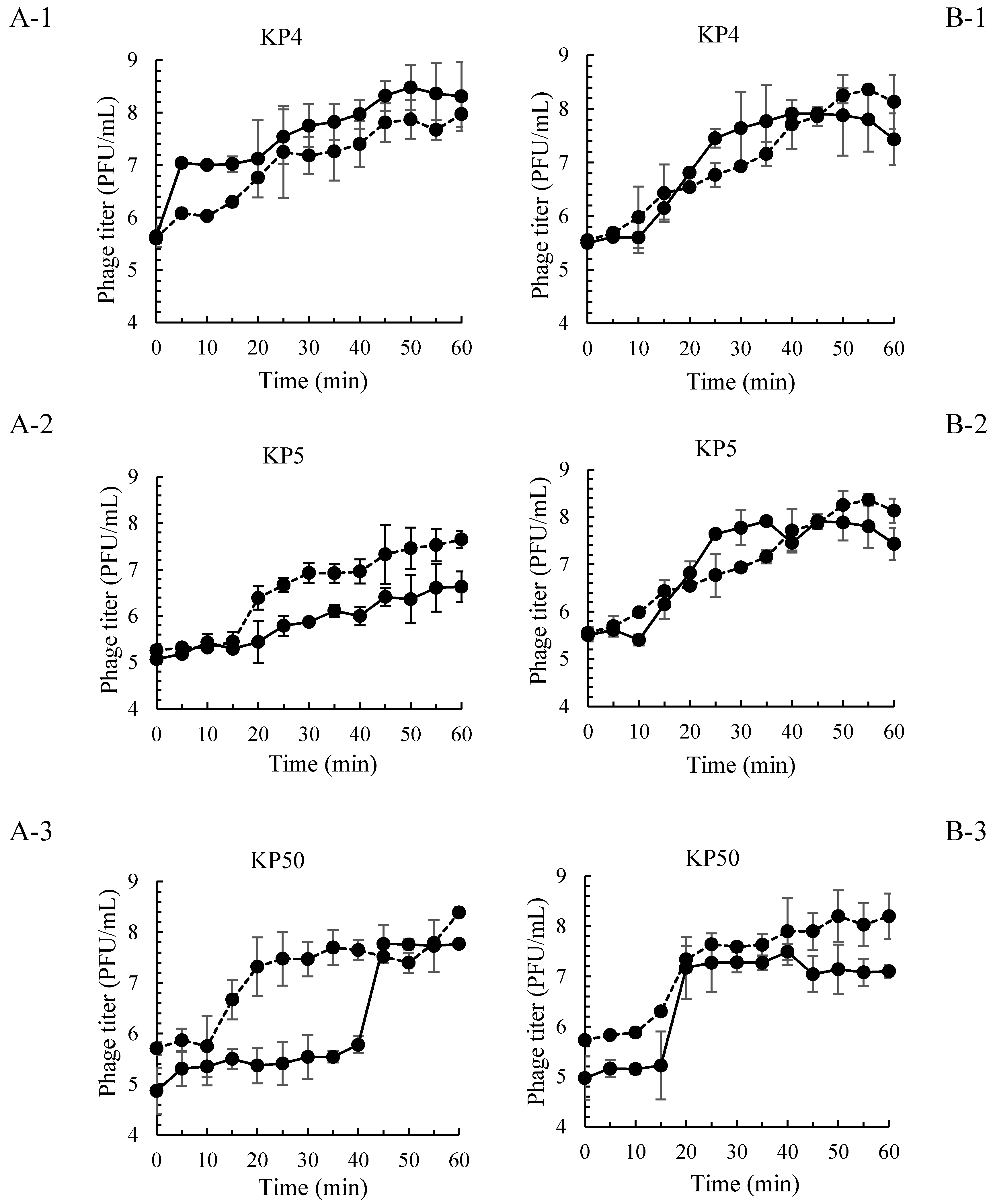 and treatment of a phage cocktail presented by . Bars represent the mean standard deviation (n = 3). The sign (*) on the lines indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the lines indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
and treatment of a phage cocktail presented by . Bars represent the mean standard deviation (n = 3). The sign (*) on the lines indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the lines indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
and treatment of a phage cocktail presented by . Bars represent the mean standard deviation (n = 3). The sign (*) on the lines indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the lines indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
and treatment of a phage cocktail presented by . Bars represent the mean standard deviation (n = 3). The sign (*) on the lines indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the lines indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.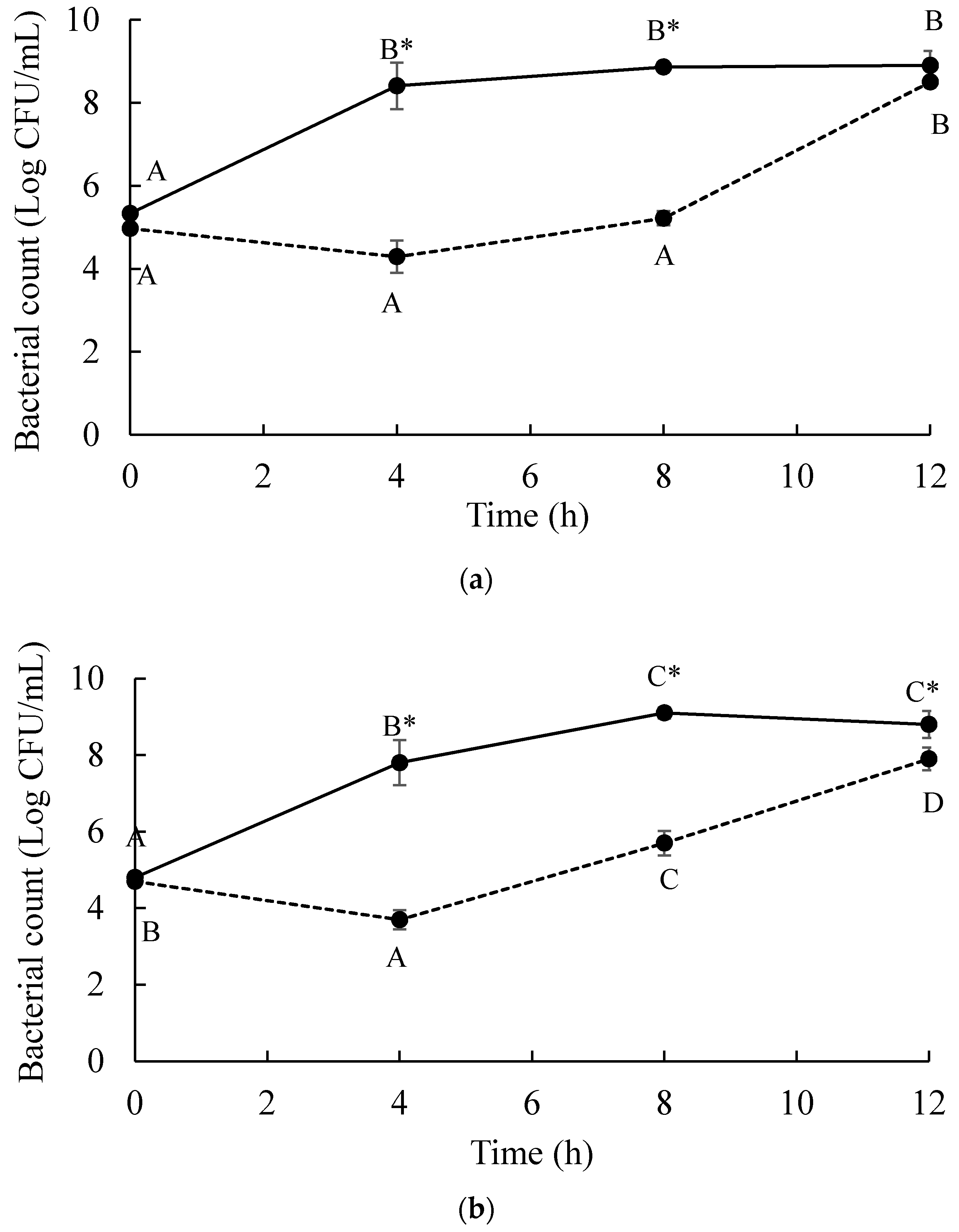
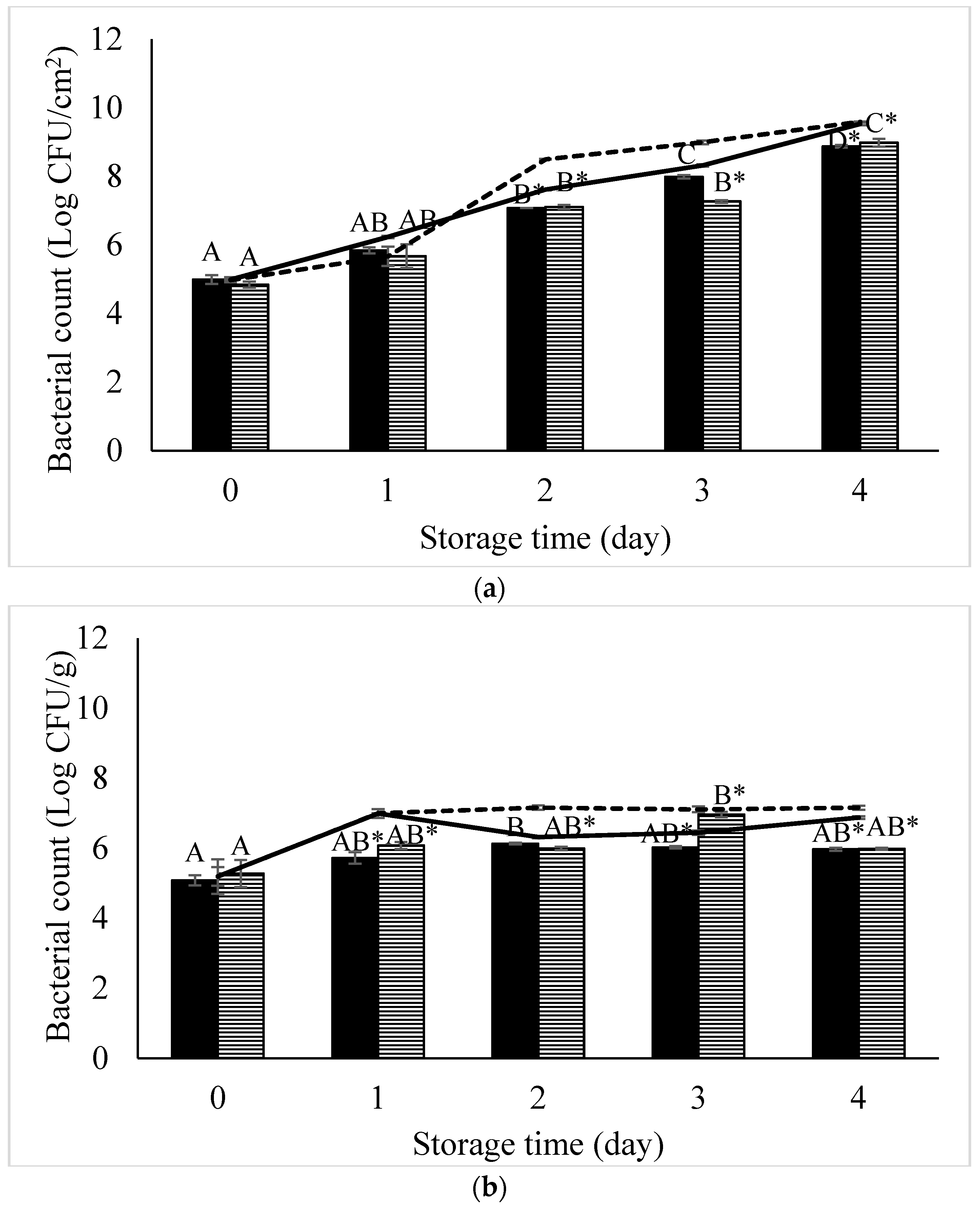
 ) and S. Typhimurium (
) and S. Typhimurium ( 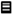 ) artificially contaminated in (a) chicken meat and (b) sunflower sprouts treated with a phage cocktail and stored at 4 °C for 4 days. Control (non-phage cocktail treated) of S. Enteritidis and S. Typhimurium inoculated on both samples presented by solid line and dashed line, respectively. Bars represent the mean standard deviation (n = 3). The sign (*) on the bars indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the bars indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
) and S. Typhimurium ( ) artificially contaminated in (a) chicken meat and (b) sunflower sprouts treated with a phage cocktail and stored at 4 °C for 4 days. Control (non-phage cocktail treated) of S. Enteritidis and S. Typhimurium inoculated on both samples presented by solid line and dashed line, respectively. Bars represent the mean standard deviation (n = 3). The sign (*) on the bars indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the bars indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
) artificially contaminated in (a) chicken meat and (b) sunflower sprouts treated with a phage cocktail and stored at 4 °C for 4 days. Control (non-phage cocktail treated) of S. Enteritidis and S. Typhimurium inoculated on both samples presented by solid line and dashed line, respectively. Bars represent the mean standard deviation (n = 3). The sign (*) on the bars indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the bars indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.
) and S. Typhimurium ( ) artificially contaminated in (a) chicken meat and (b) sunflower sprouts treated with a phage cocktail and stored at 4 °C for 4 days. Control (non-phage cocktail treated) of S. Enteritidis and S. Typhimurium inoculated on both samples presented by solid line and dashed line, respectively. Bars represent the mean standard deviation (n = 3). The sign (*) on the bars indicates significant differences (p < 0.05) of bacterial count between control and treatment during storage time for a given Salmonella strain. Different uppercase letters on the bars indicate significant differences (p < 0.05) of bacterial count among days of storage for a given Salmonella strain.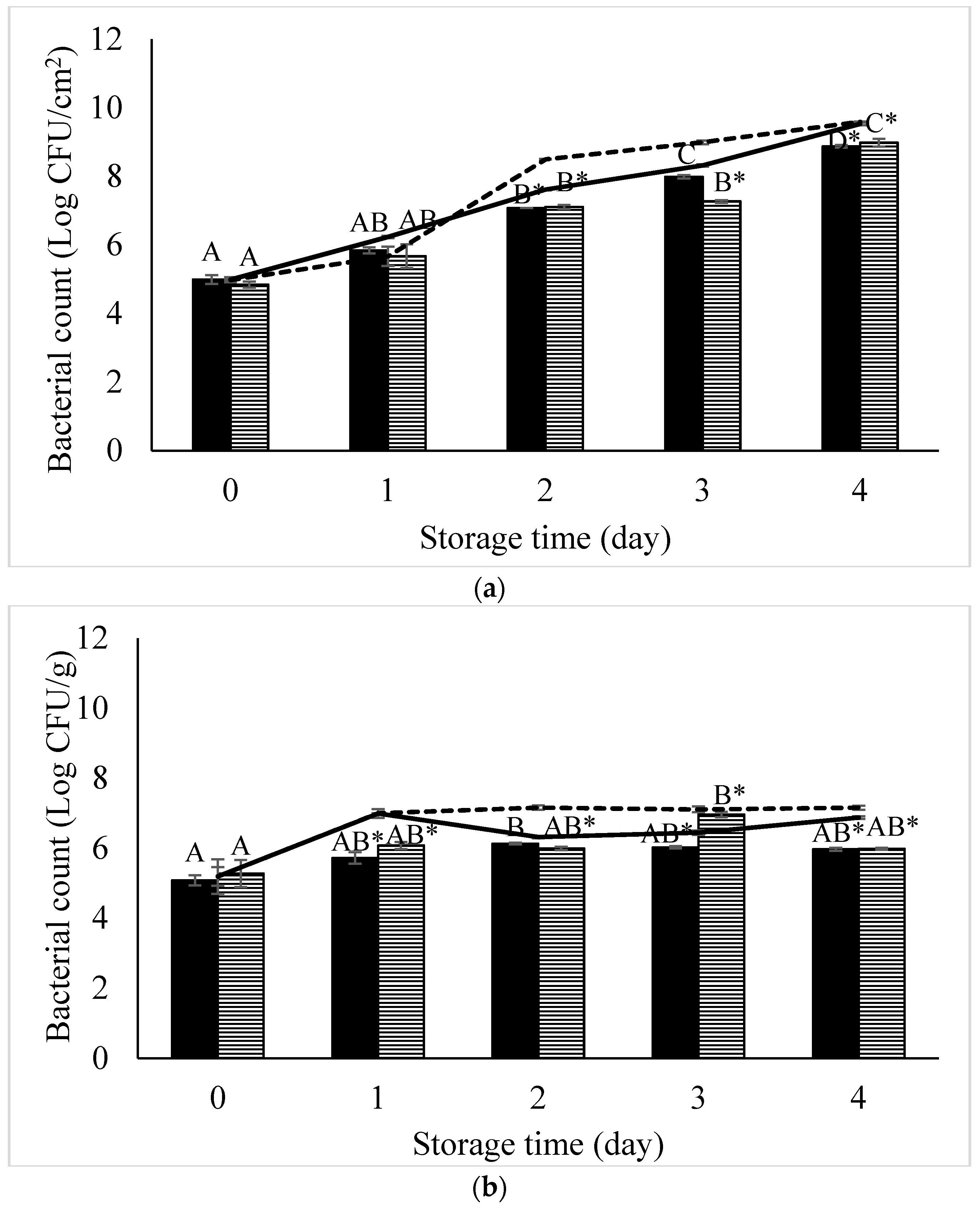

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serovars | Isolates ID | Source (Country) |
|---|---|---|
| Agona | H2-016 * | Pig slaughterhouse (Thailand) |
| FSL S5-667 | Bovine (USA) | |
| Anatum | PSU-BS-104SL | Seafood processing plant (Thailand) |
| FSL A4-525 * | Bovine (USA) | |
| Braenderup | FSL S5-373 | Human (USA) |
| Cerro | FSL R8-242 | Bovine (USA) |
| Corvolis | PSU-BS-092SL | Animal farm (Thailand) |
| FSL R8-092 | Human (USA) | |
| Dublin | W1-016 | Dairy farm (Thailand) |
| FSL S5-368 | Bovine (USA) | |
| Enteritidis | FSL S5-371 | Human (USA) |
| Give | PSU-BS-181SL | Animal farm (Thailand) |
| H2-018 * | Dairy farm (Thailand) | |
| Heidelberg | FSL S5-455 | Human (USA) |
| Infantis | FSL S5-506 | Human (USA) |
| Javiana | FSL S5-406 | Human (USA) |
| Kedougou | H2-061 | Pig slaughterhouse (Thailand) |
| Kentucky | W1-010 * | Dairy farm (Thailand) |
| PSU-BS-058SL | Chicken farm (Thailand) | |
| PSU-BS-116SL | Seafood processing plant (Thailand) | |
| FSL S5-431 | Bovine (USA) | |
| Krefeld | PSU-BS-111SL | Seafood processing plant (Thailand) |
| Mbandaka | W1-015 | Dairy farm (Thailand) |
| PSU-BS-035SL | Chicken farm (Thailand) | |
| FSL A4-793 | Bovine (USA) | |
| Montevideo | FSL S5-474 | Bovine (USA) |
| FSL S5-403 | Bovine (USA) | |
| Muennster | FSL S5-917 | Bovine (USA) |
| Newport | FSL S5-548 | Bovine (USA) |
| Oranienburg | FSL R8-376 | Human (USA) |
| Ordonez | PSU-BS-181SL | Seafood processing plant (Thailand) |
| Panama | FSL S5-454 | Human (USA) |
| Rissen | H2-015 | Pig slaughterhouse (Thailand) |
| Saintpaul | FSL S5-369 | Human (USA) |
| Stanley | PSU-BS-001SL | Swine farm (Thailand) |
| H2-002 * | Pig slaughterhouse (Thailand) | |
| PSU-BS-041SALX | Seafood processing plant (Thailand) | |
| FSL S5-464 | Human (USA) | |
| Tennessee | PSU-BS-183SL | Seafood processing plant (Thailand) |
| Typhimurium | H2-001 * | Pig slaughterhouse (Thailand) |
| PSU-BS-014SL | Chicken farm (Thailand) | |
| FSL S5-370 | Human (USA) | |
| Virchow | FSL H2-117 * | Human (Thailand) |
| FSL S5-961 | Human (USA) | |
| Weltevreden | H2-031 | Pig slaughterhouse (Thailand) |
| PSU-BS-013SL | Chicken farm (Thailand) | |
| FSL R8-798 | Human (USA) |
| Animal Farms (Sampling Date [mo/yr]) | Number of Sample (Number of Phages) |
|---|---|
| Sampling 1 (04/2014) | |
| Poultry farm; commercial | 1 (1) |
| Swine farm; commercial | 1 (0) |
| Goat farm; commercial | 1 (1) |
| Sampling 2 (08/2015) | |
| Bovine farm; free range | 1 (3) |
| Poultry farm; free range | 4 (15) |
| Sampling 3 (10/2016) | |
| Poultry farm; free range | 4 (16) |
| Total | 12 (36) |
| Salmonella Phage ID | Source | Estimated Genome Size (kb) |
|---|---|---|
| KP3 | Goat feces | 200 ± 2 |
| KP4 | Soil (poultry farm 2) | 105 ± 2 |
| KP5 | Poultry feces (poultry farm 3) | 60 ± 2 |
| KP6 | Poultry feed (poultry farm 2) | 72 ± 2 |
| KP7 | Soil (poultry farm 2) | 60 ± 2 |
| KP10 | Poultry feed (poultry farm 2) | 60 ± 2 |
| KP11 | Soil (poultry farm 2) | 60 ± 2 |
| KP34 | Poultry feces (poultry farm 1) | 60 ± 2 |
| KP38 | Bovine feces | 60 ± 2 |
| KP39 | Bovine feces | 50 ± 2 |
| KP41 | Drinking water (poultry farm 3) | 97 ± 2 |
| KP43 | Poultry feces (poultry farm 3) | 60 ± 2 |
| KP49 | Soil (poultry farm 3) | 60 ± 2 |
| KP50 | Poultry feces (poultry farm 3) | 103 ± 2 |
| KP53 | Poultry feces (poultry farm 3) | 50 ± 2 |
| KP54 | Soil (poultry farm 3) | 50 ± 2 |
| KP55 | Poultry feed (poultry farm 3) | 60 ± 2 |
| Serovars | Source | % Susceptibility (Level) a | |
|---|---|---|---|
| Thailand | USA | ||
| Agona | Pig slaughterhouse | 41.7 (H) | - |
| Anatum | Bovine | - | 41.7 (H) |
| Enteritidis | Human | - | 50.0 (H) |
| Give | Animal farm | 63.9 (H) | - |
| Dairy farm | 50.0 (H) | - | |
| Kedougou | Pig slaughterhouse | 44.4 (H) | - |
| Kentucky | Dairy farm | 38.9 (H) | - |
| Chicken farm | 41.7 (H) | - | |
| Typhimurium | Pig slaughterhouse | 47.2 (H) | - |
| Chicken farm | 55.6 (H) | - | |
| Weltevreden | Pig slaughterhouse | 69.4 (H) | - |
| Chicken farm | 58.3 (H) | - | |
| Dublin | Bovine | - | 19.4 (M) |
| Kentucky | Seafood processing plant | 13.9 (M) | - |
| Rissen | Pig slaughterhouse | 27.8 (M) | - |
| Virchow | Human | - | 11.1 (M) |
| Weltevreden | Human | - | 13.9 (M) |
| Agona | Bovine | - | No lysis |
| Anatum | Seafood processing plant | 8.3 (L) | - |
| Braenderup | Human | - | No lysis |
| Cerro | Bovine | 2.8 (L) | - |
| Corvalis | Animal farm | 2.8 (L) | - |
| Human | 2.8 (L) | - | |
| Dublin | Dairy farm | 2.8 (L) | - |
| Heidelberg | Human | - | No lysis |
| Infantis | Human | - | No lysis |
| Javiana | Human | - | 8.3 (L) |
| Kentucky | Bovine | - | 5.6 (L) |
| Krefeld | Seafood processing plant | No lysis | - |
| Mbandaka | Dairy farm | 5.6 (L) | - |
| Chicken farm | No lysis | - | |
| Bovine | - | No lysis | |
| Montevideo | Bovine | - | No lysis |
| Bovine | - | No lysis | |
| Muennster | Bovine | - | 2.8 (L) |
| Newport | Bovine | - | 2.8 (L) |
| Oranienburg | Human | - | No lysis |
| Ordonez | Seafood processing plant | 5.6 (L) | - |
| Panama | Human | - | 2.8 (L) |
| Saintpaul | Human | - | No lysis |
| Stanley | Swine farm | 2.8 (L) | - |
| Pig slaughterhouse | 5.6 (L) | - | |
| Seafood processing plant | No lysis | - | |
| Human | - | No lysis | |
| Tennessee | Seafood processing plant | 2.8 (L) | - |
| Typhimurium | Human | - | 5.6 (L) |
| Virchow | Human | 5.6 (L) | - |
| Salmonella Phage | Reference Salmonella Serovar | Target Salmonella Serovar | Lysis Ability a | Level of EOP b | |||||
|---|---|---|---|---|---|---|---|---|---|
| Phage Titer (PFU/mL) | |||||||||
| 107 | 106 | 105 | 104 | 103 | |||||
| KP1 | Anatum (A4-525) | Enteritidis | +++ | ++ | - | - | - | Medium | |
| Typhimurium | ++ | - | - | - | - | Medium | |||
| KP2 | Anatum (A4-525) | Enteritidis | +++ | +++ | ++ | - | - | Medium | |
| Typhimurium | +++ | ++ | - | - | - | Low | |||
| KP4 | Anatum (A4-525) | Enteritidis | +++ | +++ | +++ | + | - | Medium | |
| Typhimurium | +++ | +++ | +++ | + | - | Medium | |||
| KP5 | Anatum (A4-525) | Enteritidis | +++ | +++ | +++ | + | - | Medium | |
| Typhimurium | +++ | +++ | ++ | - | - | Medium | |||
| KP9 | Anatum (A4-525) | Enteritidis | +++ | +++ | +++ | ++ | - | High | |
| Typhimurium | +++ | + | - | - | - | Low | |||
| KP34 | Virchow (H2-117) | Enteritidis | +++ | +++ | ++ | - | - | Medium | |
| Typhimurium | ++ | - | - | - | - | Low | |||
| KP36 | Virchow (H2-117) | Enteritidis | +++ | +++ | +++ | ++ | - | Medium | |
| Typhimurium | ++ | - | - | - | - | Low | |||
| KP49 | Agona (H2-016) | Enteritidis | ++ | - | - | - | - | Low | |
| Typhimurium | ++ | - | - | - | - | Low | |||
| KP50 | Agona (H2-016) | Enteritidis | +++ | +++ | +++ | +++ | + | High | |
| Typhimurium | +++ | +++ | +++ | + | - | Medium | |||
| Salmonella Phage ID | Latent Period (min) | Burst Size (PFU/Cell) | ||||||
|---|---|---|---|---|---|---|---|---|
| S. Enteritidis | S. Typhimurium | S. Enteritidis | S. Typhimurium | |||||
| MOI 100 | MOI 10 | MOI 100 | MOI 10 | MOI 100 | MOI 10 | MOI 100 | MOI 10 | |
| KP4 | 5 | 10 | 15 | 10 | 25.1 | 16.6 | 70.8 | 19.1 |
| KP5 | 15 | 15 | 10 | 15 | 30.1 | 6.6 | 173.7 | 19.1 |
| KP50 | 40 | 10 | 15 | 10 | 97.7 | 37.2 | 112.2 | 28.8 |
| Treatment | Lysis Ability a | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cocktail | KP4 | KP5 | KP50 | ||||||
| Phage Titer (PFU/mL) | |||||||||
| 107 | 106 | 107 | 106 | 107 | 106 | 107 | 106 | ||
| Control SE (non-phage treatment) | ++ | ++ | ++ | ++ | ++ | ++ | ++ | + | |
| Control ST (non-phage treatment) | ++ | + | ++ | + | ++ | − | + | − | |
| Phage-treated SE | ++ | ++ | ++ | − | ++ | − | ++ | − | |
| Phage-treated ST | ++ | + | ++ | + | ++ | − | + | − | |
| Source of Isolation | Country | Estimated Genome Size (kb) | Reference |
|---|---|---|---|
| Goat feces | Thailand | 200 ± 2 | This study |
| Bovine feces | Thailand | 50 ± 2, 60 ± 2 | This study |
| Poultry feed | Thailand | 60 ± 2, 72 ± 2 | This study |
| Soil (poultry farm) | Thailand | 50 ± 2, 60 ± 2, 105 ± 2 | This study |
| Drinking water (poultry farm) | Thailand | 97 ± 2 | This study |
| Poultry feces | Thailand | 50 ± 2, 60 ± 2, 103 ± 2 | This study |
| Dairy farms | USA | 22 to 156 | [4] |
| Dairy farms | Thailand | 40 to 200 | [31] |
| Sewage effluent | UK | 40, 48.5 and 155 | [37] |
| Chicken feces | Korea | 240 | [38] |
| Sewage (poultry farm) | Korea | 40 | [39] |
| Water buffalo feces | Southern Italy | 39 | [40] |
| Swine lagoon effluent (poultry farm) | UK | 42 | [41] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petsong, K.; Benjakul, S.; Chaturongakul, S.; Switt, A.I.M.; Vongkamjan, K. Lysis Profiles of Salmonella Phages on Salmonella Isolates from Various Sources and Efficiency of a Phage Cocktail against S. Enteritidis and S. Typhimurium. Microorganisms 2019, 7, 100. https://doi.org/10.3390/microorganisms7040100
Petsong K, Benjakul S, Chaturongakul S, Switt AIM, Vongkamjan K. Lysis Profiles of Salmonella Phages on Salmonella Isolates from Various Sources and Efficiency of a Phage Cocktail against S. Enteritidis and S. Typhimurium. Microorganisms. 2019; 7(4):100. https://doi.org/10.3390/microorganisms7040100
Chicago/Turabian StylePetsong, Kantiya, Soottawat Benjakul, Soraya Chaturongakul, Andrea I. Moreno Switt, and Kitiya Vongkamjan. 2019. "Lysis Profiles of Salmonella Phages on Salmonella Isolates from Various Sources and Efficiency of a Phage Cocktail against S. Enteritidis and S. Typhimurium" Microorganisms 7, no. 4: 100. https://doi.org/10.3390/microorganisms7040100
APA StylePetsong, K., Benjakul, S., Chaturongakul, S., Switt, A. I. M., & Vongkamjan, K. (2019). Lysis Profiles of Salmonella Phages on Salmonella Isolates from Various Sources and Efficiency of a Phage Cocktail against S. Enteritidis and S. Typhimurium. Microorganisms, 7(4), 100. https://doi.org/10.3390/microorganisms7040100