Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica



{kind=link}
Abstract
1. Introduction
2. Protein Production
2.1. Recombinant Proteins
2.2. Metabolic Load: the Weight of Recombinant Expression on Host Cells
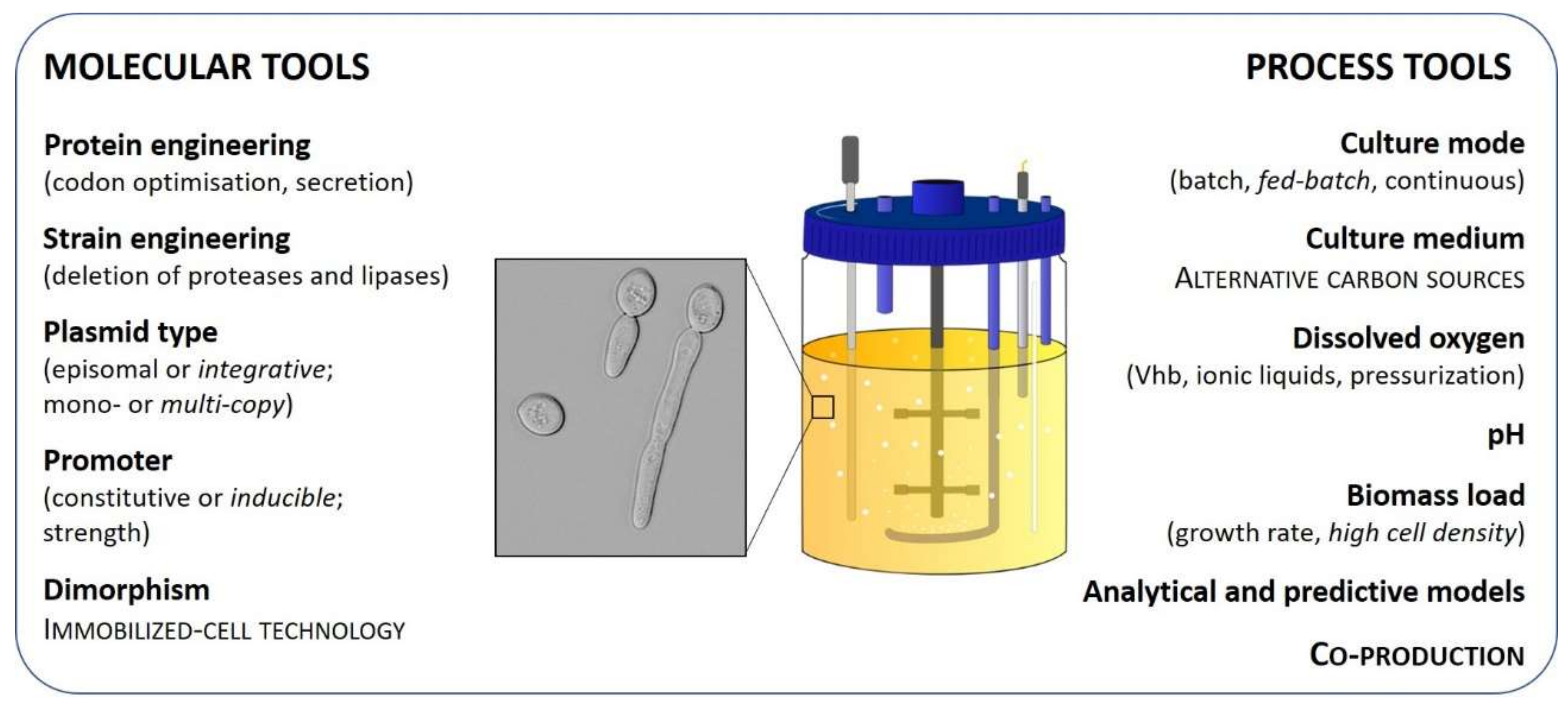
3. Molecular Strategies
3.1. Strains and Plasmids: Backbones of Recombinant Expression
3.2. Copy Number: Is More Better?
3.3. Promoters: an Inducible Advantage
3.4. Unpredictable Dimorphism
4. Process Strategies
4.1. The Predominance of Fed-Batch
4.2. Culture Medium: a Matter of Definition
4.3. Oxygen Supply: Toward Innovation
4.4. Multifaceted Implications of pH
4.5. Striking a Balance with Biomass Load
4.6. Working Volume: To Navigate along the Scale
4.7. Analytical and Predictive Bioprocess Models
4.8. Coupling Recombinant Protein Production with Another Bioprocess
5. Examples
5.1. Low Value Protein: The Lipase Lip2p for Industrial Catalysis
5.2. High Value Protein: The Human Interferon α2b for Medical Use
6. Conclusions and Prospects
Funding
Acknowledgments
Conflicts of Interest
References
- Mattanovich, D.; Branduardi, P.; Dato, L.; Gasser, B.; Sauer, M.; Porro, D. Recombinant Protein Production in Yeasts. In Recombinant Gene Expression; Methods in Molecular Biology; Lorence, A., Ed.; Humana Press: Totowa, NJ, USA, 2012; pp. 329–358. ISBN 978-1-61779-433-9. [Google Scholar]
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonça Bahia, F.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Nicaud, J.-M. Filamentous fungi-like secretory pathway strayed in a yeast system: Peculiarities of Yarrowia lipolytica100 secretory pathway underlying its extraordinary performance. Appl. Microbiol. Biotechnol. 2019, 103, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Nicaud, J.-M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Hamsa, P.V.; Chattoo, B.B. Cloning and growth-regulated expression of the gene encoding the hepatitis B virus middle surface antigen in Yarrowia lipolytica. Gene 1994, 143, 165–170. [Google Scholar] [CrossRef]
- Bhave, S.L.; Chattoo, B.B. Expression of Vitreoscilla hemoglobin improves growth and levels of extracellular enzyme in Yarrowia lipolytica. Biotechnol. Bioeng. 2003, 84, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Destain, J.; Thonart, P. Methyl Oleate Modulates LIP2 Expression in the Lipolytic Yeast Yarrowia lipolytica. Biotechnol. Lett. 2005, 27, 1751–1754. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Chen, Y.-F.; Chen, C.-Y.; Chen, W.-L.; Ciou, Y.-P.; Liu, W.-H.; Yang, C.-H. Production of ferulic acid from lignocellulolytic agricultural biomass by Thermobifida fusca thermostable esterase produced in Yarrowia lipolytica transformant. Bioresour. Technol. 2011, 102, 8117–8122. [Google Scholar] [CrossRef] [PubMed]
- Kar, T.; Destain, J.; Thonart, P.; Delvigne, F. Scale-down assessment of the sensitivity of Yarrowia lipolytica to oxygen transfer and foam management in bioreactors: Investigation of the underlying physiological mechanisms. J. Ind. Microbiol. Biotechnol. 2012, 39, 337–346. [Google Scholar] [CrossRef]
- Liu, S.; Wan, D.; Wang, M.; Madzak, C.; Du, G.; Chen, J. Overproduction of pro-transglutaminase from Streptomyces hygroscopicus in Yarrowia lipolytica and its biochemical characterization. BMC Biotechnol. 2015, 15, 75. [Google Scholar] [CrossRef]
- Chang, C.C.; Ryu, D.D.Y.; Park, C.S.; Kim, J.-Y. Improvement of heterologous protein productivity using recombinant Yarrowia lipolytica and cyclic fed-batch process strategy. Biotechnol. Bioeng. 1998, 59, 379–385. [Google Scholar] [CrossRef]
- Chang, C.C.; Ryu, D.D.Y.; Park, C.S.; Kim, J.-Y. Enhancement of rice α-amylase production in recombinant Yarrowia lipolytica. J. Ferment. Bioeng. 1997, 84, 421–427. [Google Scholar] [CrossRef]
- Chang, C.C.; Ryu, D.D.Y.; Park, C.S.; Kim, J.-Y.; Ogrydziak, D.M. Recombinant bioprocess optimization for heterologous protein production using two-stage, cyclic fed-batch culture. Appl. Microbiol. Biotechnol. 1998, 49, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; Park, T.J.; Ryu, D.D.Y.; Kim, J.-Y. High Cell Density Culture of Yarrowia lipolytica Using a One-Step Feeding Process. Biotechnol. Prog. 2000, 16, 657–660. [Google Scholar] [CrossRef]
- Gasmi, N.; Lassoued, R.; Ayed, A.; Tréton, B.; Chevret, D.; Nicaud, J.M.; Kallel, H. Production and characterization of human granulocyte–macrophage colony-stimulating factor (hGM-CSF) expressed in the oleaginous yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012, 96, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, N.; Ayed, A.; Nicaud, J.-M.; Kallel, H. Design of an efficient medium for heterologous protein production in Yarrowia lipolytica: Case of human interferon alpha 2b. Microb. Cell Fact. 2011, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, N.; Ayed, A.; Ammar, B.B.H.; Zrigui, R.; Nicaud, J.-M.; Kallel, H. Development of a cultivation process for the enhancement of human interferon alpha 2b production in the oleaginous yeast, Yarrowia lipolytica. Microb. Cell Fact. 2011, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C.; Tréton, B.; Blanchin-Roland, S. Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J. Mol. Microbiol. Biotechnol. 2000, 2, 207–216. [Google Scholar]
- Tiels, P.; Baranova, E.; Piens, K.; De Visscher, C.; Pynaert, G.; Nerinckx, W.; Stout, J.; Fudalej, F.; Hulpiau, P.; Tännler, S.; et al. A bacterial glycosidase enables mannose-6-phosphate modification and improved cellular uptake of yeast-produced recombinant human lysosomal enzymes. Nat. Biotechnol. 2012, 30, 1225–1231. [Google Scholar] [CrossRef]
- Botes, A.L.; Labuschagne, M.; Roth, R.; Mitra, R.K.; Lotter, J.; Lalloo, R.; Ramduth, D.; Rohitlall, N.; Simpson, C.; van Zyl, P. Recombinant Yeasts for Synthesizing Epoxide Hydrolases. U.S. Patent US 20080171359 A1, 17 July 2008. [Google Scholar]
- Maharajh, D.; Roth, R.; Lalloo, R.; Simpson, C.; Mitra, R.; Görgens, J.; Ramchuran, S. Multi-copy expression and fed-batch production of Rhodotorula araucariae epoxide hydrolase in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2008, 79, 235–244. [Google Scholar] [CrossRef]
- Maharajh, D.; Lalloo, R.; Görgens, J. Effect of an exponential feeding regime on the production of Rhodotorula araucariae epoxide hydrolase in Yarrowia lipolytica. Lett. Appl. Microbiol. 2008, 47, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Jolivet, P.; Bordes, F.; Fudalej, F.; Cancino, M.; Vignaud, C.; Dossat, V.; Burghoffer, C.; Marty, A.; Chardot, T.; Nicaud, J.M. Analysis of Yarrowia lipolytica extracellular lipase Lip2p glycosylation. FEMS Yeast Res. 2007, 7, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Kopečný, D.; Pethe, C.; Šebela, M.; Houba-Hérin, N.; Madzak, C.; Majira, A.; Laloue, M. High-level expression and characterization of Zea mays cytokinin oxidase/dehydrogenase in Yarrowia lipolytica. Biochimie 2005, 87, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- De Pourcq, K.; Vervecken, W.; Dewerte, I.; Valevska, A.; Van Hecke, A.; Callewaert, N. Engineering the yeast Yarrowia lipolytica for the production of therapeutic proteins homogeneously glycosylated with Man8GlcNAc2 and Man5GlcNAc2. Microb. Cell Fact. 2012, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, N.L.M.; Vervecken, W.; Pourcq, K.J.M.D.; Geysens, S.C.J.; Guerfal, M. Yarrowia Lipolytica and Pichia Pastoris HAC1 Nucleic Acids 2011. U.S. Patent US20090069232A1, 12 March 2009. [Google Scholar]
- Förster, A.; Aurich, A.; Mauersberger, S.; Barth, G. Citric acid production from sucrose using a recombinant strain of the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2007, 75, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Lazar, Z.; Rossignol, T.; Verbeke, J.; Crutz-Le Coq, A.-M.; Nicaud, J.-M.; Robak, M. Optimized invertase expression and secretion cassette for improving Yarrowia lipolytica growth on sucrose for industrial applications. J. Ind. Microbiol. Biotechnol. 2013, 40, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Wang, Q.; Zhang, F.; Zhang, S.-C.; Chi, Z.-M.; Madzak, C. Direct conversion of inulin into single cell protein by the engineered Yarrowia lipolytica carrying inulinase gene. Process Biochem. 2011, 46, 1442–1448. [Google Scholar] [CrossRef]
- Trassaert, M.; Vandermies, M.; Carly, F.; Denies, O.; Thomas, S.; Fickers, P.; Nicaud, J.-M. New inducible promoter for gene expression and synthetic biology in Yarrowia lipolytica. Microb. Cell Fact. 2017, 16, 141. [Google Scholar] [CrossRef]
- Glick, B.R. Metabolic load and heterologous gene expression. Biotechnol. Adv. 1995, 13, 247–261. [Google Scholar] [CrossRef]
- Görgens, J.F.; van Zyl, W.H.; Knoetze, J.H.; Hahn-Hägerdal, B. The metabolic burden of the PGK1 and ADH2 promoter systems for heterologous xylanase production by Saccharomyces cerevisiae in defined medium. Biotechnol. Bioeng. 2001, 73, 238–245. [Google Scholar] [CrossRef]
- Corchero, J.L.; Villaverde, A. Plasmid maintenance in Escherichia coli recombinant cultures is dramatically, steadily, and specifically influenced by features of the encoded proteins. Biotechnol. Bioeng. 1998, 58, 625–632. [Google Scholar] [CrossRef]
- Van Rensburg, E.; den Haan, R.; Smith, J.; van Zyl, W.H.; Görgens, J.F. The metabolic burden of cellulase expression by recombinant Saccharomyces cerevisiae Y294 in aerobic batch culture. Appl. Microbiol. Biotechnol. 2012, 96, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Roth, R.; Moodley, V.; van Zyl, P. Heterologous Expression and Optimized Production of an Aspergillus aculeatus Endo-1,4-β-mannanase in Yarrowia lipolytica. Mol. Biotechnol. 2009, 43, 112–120. [Google Scholar] [CrossRef] [PubMed]
- YaPing, W.; Ben, R.; Ling, Z.; Lixin, M. High-level expression of two thermophilic β-mannanases in Yarrowia lipolytica. Protein Expr. Purif. 2017, 133, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barth, G.; Gaillardin, C. Yarrowia lipolytica. In Nonconventional Yeasts in Biotechnology; Springer: Berlin/Heidelberg, Germany, 1996; pp. 313–388. ISBN 978-3-642-79858-0. [Google Scholar]
- Babour, A.; Kabani, M.; Boisramé, A.; Beckerich, J.-M. Characterization of Ire1 in the yeast Yarrowia lipolytica reveals an important role for the Sls1 nucleotide exchange factor in unfolded protein response regulation. Curr. Genet. 2008, 53, 337. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Cheon, S.A.; Kang, H.A.; Kim, J.-Y. Functional characterization of the unconventional splicing of Yarrowia lipolytica HAC1 mRNA induced by unfolded protein response. Yeast 2010, 27, 443–452. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, J.C.; Koskela, E.V.; Nandania, J.; Frey, A.D.; Velagapudi, V. Understanding the metabolic burden of recombinant antibody production in Saccharomyces cerevisiae using a quantitative metabolomics approach. Yeast 2018, 35, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Hanquier, J.; Sorlet, Y.; Desplancq, D.; Baroche, L.; Ebtinger, M.; Lefèvre, J.-F.; Pattus, F.; Hershberger, C.L.; Vertès, A.A. A single mutation in the activation site of bovine trypsinogen enhances its accumulation in the fermentation broth of the yeast Pichia pastoris. Appl. Environ. Microbiol. 2003, 69, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Białas, W.; Borkowska, M.; Grajek, W. Cloning, expression, and purification of insect (Sitophilus oryzae) alpha-amylase, able to digest granular starch, in Yarrowia lipolytica host. Appl. Microbiol. Biotechnol. 2015, 99, 2727–2739. [Google Scholar] [CrossRef]
- Angov, E. Codon usage: Nature’s roadmap to expression and folding of proteins. Biotechnol. J. 2011, 6, 650–659. [Google Scholar] [CrossRef]
- Celińska, E.; Borkowska, M.; Białas, W.; Korpys, P.; Nicaud, J.-M. Robust signal peptides for protein secretion in Yarrowia lipolytica: Identification and characterization of novel secretory tags. Appl. Microbiol. Biotechnol. 2018, 102, 5221–5233. [Google Scholar] [CrossRef] [PubMed]
- Le Dall, M.-T.; Nicaud, J.-M.; Gaillardin, C. Multiple-copy integration in the yeast Yarrowia lipolytica. Curr. Genet. 1994, 26, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Le Dall, M.; Gaillardin, C.; Thonart, P.; Nicaud, J.M. New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J. Microbiol. Methods 2003, 55, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Bordes, F.; Fudalej, F.; Dossat, V.; Nicaud, J.-M.; Marty, A. A new recombinant protein expression system for high-throughput screening in the yeast Yarrowia lipolytica. J. Microbiol. Methods 2007, 70, 493–502. [Google Scholar] [CrossRef] [PubMed]
- De Pourcq, K.; Tiels, P.; Hecke, A.V.; Geysens, S.; Vervecken, W.; Callewaert, N. Engineering Yarrowia lipolytica to Produce Glycoproteins Homogeneously Modified with the Universal Man3GlcNAc2 N-Glycan Core. PLoS ONE 2012, 7, e39976. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, L.; León, M.; Zueco, J. Isolation of the MNN9 gene of Yarrowia lipolytica (YlMNN9) and phenotype analysis of a mutant ylmnn9Δ strain. Yeast 2003, 20, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-N.; Song, Y.; Kim, J.-Y.; Oh, D.-B.; Kang, H.A. A Novel YlMPO1 Gene Derived from Yarrowia lipolytica and a Process for Preparing a Glycoprotein Not Being Mannosylphosphorylated by Using a Mutated Yarrowia lipolytica in Which YlMPO1 Gene Is Disrupted. World Patent Application WO2008136564 A1, 13 December 2008. [Google Scholar]
- Song, Y.; Choi, M.H.; Park, J.-N.; Kim, M.W.; Kim, E.J.; Kang, H.A.; Kim, J.-Y. Engineering of the Yeast Yarrowia lipolytica for the Production of Glycoproteins Lacking the Outer-Chain Mannose Residues of N-Glycans. Appl. Environ. Microbiol. 2007, 73, 4446–4454. [Google Scholar] [CrossRef]
- Nicaud, J.; Madzak, C.; Vandenbroek, P.; Gysler, C.; Duboc, P.; Niederberger, P.; Gaillardin, C. Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res. 2002, 2, 371–379. [Google Scholar]
- Borsenberger, V.; Onésime, D.; Lestrade, D.; Rigouin, C.; Neuvéglise, C.; Daboussi, F.; Bordes, F. Multiple Parameters Drive the Efficiency of CRISPR/Cas9-Induced Gene Modifications in Yarrowia lipolytica. J. Mol. Biol. 2018, 430, 4293–4306. [Google Scholar] [CrossRef]
- Gao, S.; Tong, Y.; Wen, Z.; Zhu, L.; Ge, M.; Chen, D.; Jiang, Y.; Yang, S. Multiplex gene editing of the Yarrowia lipolytica genome using the CRISPR-Cas9 system. J. Ind. Microbiol. Biotechnol. 2016, 43, 1085–1093. [Google Scholar] [CrossRef]
- Verbeke, J.; Beopoulos, A.; Nicaud, J.-M. Efficient homologous recombination with short length flanking fragments in Ku70 deficient Yarrowia lipolytica strains. Biotechnol. Lett. 2013, 35, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Holkenbrink, C.; Dam, M.I.; Kildegaard, K.R.; Beder, J.; Dahlin, J.; Belda, D.D.; Borodina, I. EasyCloneYALI: CRISPR/Cas9-Based Synthetic Toolbox for Engineering of the Yeast Yarrowia lipolytica. Biotechnol. J. 2018, 13, 1700543. [Google Scholar] [CrossRef] [PubMed]
- Juretzek, T.; Le Dall, M.-T.; Mauersberger, S.; Gaillardin, C.; Barth, G.; Nicaud, J.-M. Vectors for gene expression and amplification in the yeast Yarrowia lipolytica. Yeast 2001, 18, 97–113. [Google Scholar] [CrossRef]
- Heslot, H.; Nicaud, J.M.; Fabre, E.; Beckerich, J.M.; Fournier, P.; Gaillardin, C. Cloning of the alkaline extracellular protease gene of Yarrowia lipolytica and its use to express foreign genes. In Microbiological Application in Food Biotechnology; Nga, B.H., Lee, Y.K., Eds.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 27–45. [Google Scholar]
- Nicaud, J.-M.; Fabre, E.; Gaillardin, C. Expression of invertase activity in Yarrowia lipolytica and its use as a selective marker. Curr. Genet. 1989, 16, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Tharaud, C.; Ribet, A.-M.; Costes, C.; Gaillardin, C. Secretion of human blood coagulation factor XIIIa by the yeast Yarrowia lipolytica. Gene 1992, 121, 111–119. [Google Scholar] [CrossRef]
- James, L.C.; Strick, C.A. Multiple Integrative Vectors and Yarrowia lipolytica Transformants. U.S. Patent US5786212A, 28 July 1998. [Google Scholar]
- Ogrydziak, D.M.; Scharf, S.J. Alkaline extracellular protease produced by Saccharomycopsis lipolytica CX161-1B. J. Gen. Microbiol. 1982, 128, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Dalbøge, H. Yeast Promoters Suitable for Expression Cloning in Yeast and Heterologous Expression of Proteins in Yeast. U.S. Patent US6265185, 24 July 2001. [Google Scholar]
- Blazeck, J.; Liu, L.; Redden, H.; Alper, H. Tuning Gene Expression in Yarrowia lipolytica by a Hybrid Promoter Approach. Appl. Environ. Microbiol. 2011, 77, 7905–7914. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Reed, B.; Garg, R.; Gerstner, R.; Pan, A.; Agarwala, V.; Alper, H.S. Generalizing a hybrid synthetic promoter approach in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2013, 97, 3037–3052. [Google Scholar] [CrossRef] [PubMed]
- Dulermo, R.; Brunel, F.; Dulermo, T.; Ledesma-Amaro, R.; Vion, J.; Trassaert, M.; Thomas, S.; Nicaud, J.-M.; Leplat, C. Using a vector pool containing variable-strength promoters to optimize protein production in Yarrowia lipolytica. Microb. Cell Fact. 2017, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Shabbir Hussain, M.; Gambill, L.; Smith, S.; Blenner, M.A. Engineering Promoter Architecture in Oleaginous Yeast Yarrowia lipolytica. ACS Synth. Biol. 2016, 5, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Juretzek, T.; Wang, H.-J.; Nicaud, J.-M.; Mauersberger, S.; Barth, G. Comparison of promoters suitable for regulated overexpression of β-galactosidase in the alkane-utilizing yeast Yarrowia lipolytica. Biotechnol. Bioprocess Eng. 2000, 5, 320–326. [Google Scholar] [CrossRef]
- Aloulou, A.; Rodriguez, J.A.; Puccinelli, D.; Mouz, N.; Leclaire, J.; Leblond, Y.; Carrière, F. Purification and biochemical characterization of the Lip2 lipase from Yarrowia lipolytica. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2007, 1771, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Emond, S.; Montanier, C.; Nicaud, J.-M.; Marty, A.; Monsan, P.; André, I.; Remaud-Siméon, M. New Efficient Recombinant Expression System to Engineer Candida antarctica Lipase B. Appl. Environ. Microbiol. 2010, 76, 2684–2687. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Fudalej, F.; Nicaud, J.-M.; Destain, J.; Thonart, P. Selection of new over-producing derivatives for the improvement of extracellular lipase production by the non-conventional yeast Yarrowia lipolytica. J. Biotechnol. 2005, 115, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Sassi, H.; Delvigne, F.; Kallel, H.; Fickers, P. pH and Not Cell Morphology Modulate pLIP2 Induction in the Dimorphic Yeast Yarrowia lipolytica. Curr. Microbiol. 2017, 74, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Nicaud, J.M.; Gaillardin, C.; Destain, J.; Thonart, P. Carbon and nitrogen sources modulate lipase production in the yeast Yarrowia lipolytica. J. Appl. Microbiol. 2004, 96, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Fickers, P.; Marty, A.; Nicaud, J.M. The lipases from Yarrowia lipolytica: Genetics, production, regulation, biochemical characterization and biotechnological applications. Biotechnol. Adv. 2011, 29, 632–644. [Google Scholar] [CrossRef]
- Sassi, H.; Delvigne, F.; Kar, T.; Nicaud, J.-M.; Coq, A.-M.C.-L.; Steels, S.; Fickers, P. Deciphering how LIP2 and POX2 promoters can optimally regulate recombinant protein production in the yeast Yarrowia lipolytica. Microb. Cell Fact. 2016, 15, 159. [Google Scholar] [CrossRef]
- Park, Y.-K.; Korpys, P.; Kubiak, M.; Celińska, E.; Soudier, P.; Trébulle, P.; Larroude, M.; Rossignol, T.; Nicaud, J.-M. Engineering the architecture of erythritol-inducible promoters for regulated and enhanced gene expression in Yarrowia lipolytica. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef]
- Nicaud, J.-M.; Trassaert, M.; Thomas, S.; Fickers, P.; Vandermies, M.; Carly, F. Inducible Promoter for Gene Expression and Synthetic Biology. World Patent Application WO2018210867 A1, 22 November 2018. [Google Scholar]
- Timoumi, A.; Guillouet, S.E.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Impacts of environmental conditions on product formation and morphology of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2018, 102, 3831–3848. [Google Scholar] [CrossRef]
- Pérez-Campo, F.M.; Domínguez, A. Factors Affecting the Morphogenetic Switch in Yarrowia lipolytica. Curr. Microbiol. 2001, 43, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Palande, A.S.; Kulkarni, S.V.; León-Ramirez, C.; Campos-Góngora, E.; Ruiz-Herrera, J.; Deshpande, M.V. Dimorphism and hydrocarbon metabolism in Yarrowia lipolytica var. indica. Arch. Microbiol. 2014, 196, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Zinjarde, S.S.; Pant, A.; Deshpande, M.V. Dimorphic transition in Yarrowia lipolytica isolated from oil-polluted sea water. Mycol. Res. 1998, 102, 553–558. [Google Scholar] [CrossRef]
- Fickers, P.; Destain, J.; Thonart, P. Improvement of Yarrowia lipolytica lipase production by fed-batch fermentation. J. Basic Microbiol. 2009, 49, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C.; Otterbein, L.; Chamkha, M.; Moukha, S.; Asther, M.; Gaillardin, C.; Beckerich, J. Heterologous production of a laccase from the basidiomycete Pycnoporus cinnabarinus in the dimorphic yeast Yarrowia lipolytica. FEMS Yeast Res. 2005, 5, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.A.Z.; Amaral, P.F.F.; Belo, I. Yarrowia Lipolytica: An Industrial workhorse. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology Advances; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 930–940. [Google Scholar]
- Kraiem, H.; Manon, Y.; Anne-Archard, D.; Fillaudeau, L. In-situ and ex-situ rheometry of high density Yarrowia lipolytica broth: Determination of critical concentration and impact of yeast mycelial transition 2013. In Proceedings of the 48ème congrès annuel du Groupe Français de Rhéologie—GFR 2013, Nantes, France, 28 October—30 October 2013. [Google Scholar]
- Bellou, S.; Makri, A.; Triantaphyllidou, I.-E.; Papanikolaou, S.; Aggelis, G. Morphological and metabolic shifts of Yarrowia lipolytica induced by alteration of the dissolved oxygen concentration in the growth environment. Microbiology 2014, 160, 807–817. [Google Scholar] [CrossRef]
- Braga, A.; Mesquita, D.P.; Amaral, A.L.; Ferreira, E.C.; Belo, I. Aroma production by Yarrowia lipolytica in airlift and stirred tank bioreactors: Differences in yeast metabolism and morphology. Biochem. Eng. J. 2015, 93, 55–62. [Google Scholar] [CrossRef]
- Lopes, M.; Gomes, N.; Gonçalves, C.; Coelho, M.A.Z.; Mota, M.; Belo, I. Yarrowia lipolytica lipase production enhanced by increased air pressure. Lett. Appl. Microbiol. 2008, 46, 255–260. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Industrial derivative of tallow: A promising renewable substrate for microbial lipid, single-cell protein and lipase production by Yarrowia lipolytica. Electron. J. Biotechnol. 2007, 10. [Google Scholar] [CrossRef]
- Timoumi, A.; Cléret, M.; Bideaux, C.; Guillouet, S.E.; Allouche, Y.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Dynamic behavior of Yarrowia lipolytica in response to pH perturbations: Dependence of the stress response on the culture mode. Appl. Microbiol. Biotechnol. 2017, 101, 351–366. [Google Scholar] [CrossRef]
- Timoumi, A.; Bideaux, C.; Guillouet, S.E.; Allouche, Y.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Influence of oxygen availability on the metabolism and morphology of Yarrowia lipolytica: Insights into the impact of glucose levels on dimorphism. Appl. Microbiol. Biotechnol. 2017, 101, 7317–7333. [Google Scholar] [CrossRef] [PubMed]
- Bouchedja, D.N.; Danthine, S.; Kar, T.; Fickers, P.; Boudjellal, A.; Delvigne, F. Online flow cytometry, an interesting investigation process for monitoring lipid accumulation, dimorphism, and cells’ growth in the oleaginous yeast Yarrowia lipolytica JMY 775. Bioresour. Bioprocess. 2017, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Delvigne, F.; Zune, Q.; Lara, A.R.; Al-Soud, W.; Sørensen, S.J. Metabolic variability in bioprocessing: Implications of microbial phenotypic heterogeneity. Trends Biotechnol. 2014, 32, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Xie, D. Integrating Cellular and Bioprocess Engineering in the Non-Conventional Yeast Yarrowia lipolytica for Biodiesel Production: A Review. Front. Bioeng. Biotechnol. 2017, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Chávez, J.A.; Kronberg, F.; Passeron, S.; Ruiz-Herrera, J. Regulatory role of the PKA pathway in dimorphism and mating in Yarrowia lipolytica. Fungal Genet. Biol. 2009, 46, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Chávez, J.A.; Ruiz-Herrera, J. STE11 disruption reveals the central role of a MAPK pathway in dimorphism and mating in Yarrowia lipolytica. FEMS Yeast Res. 2006, 6, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Fagarasanu, A.; Rachubinski, R.A. Peroxisomal Peripheral Membrane Protein YlInp1p Is Required for Peroxisome Inheritance and Influences the Dimorphic Transition in the Yeast Yarrowia lipolytica. Eukaryot. Cell 2007, 6, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, C.A.; Rachubinski, R.A. MHY1 encodes a C2H2-type zinc finger protein that promotes dimorphic transition in the yeast Yarrowia lipolytica. J. Bacteriol. 1999, 181, 3051–3057. [Google Scholar]
- Hurtado, C.A.R.; Rachubinski, R.A. Isolation and Characterization of YlBEM1, a Gene Required for Cell Polarization and Differentiation in the Dimorphic Yeast Yarrowia lipolytica. Eukaryot. Cell 2002, 1, 526–537. [Google Scholar] [CrossRef]
- Hurtado, C.A.R.; Rachubinski, R.A. YlBMH1 encodes a 14-3-3 protein that promotes filamentous growth in the dimorphic yeast Yarrowia lipolytica. Microbiology 2002, 148, 3725–3735. [Google Scholar] [CrossRef]
- Hurtado, C.A.R.; Beckerich, J.-M.; Gaillardin, C.; Rachubinski, R.A. A Rac Homolog Is Required for Induction of Hyphal Growth in the Dimorphic Yeast Yarrowia lipolytica. J. Bacteriol. 2000, 182, 2376–2386. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, Y.-Q.; Zhao, X.-F.; Gao, X.-D. Roles of the three Ras proteins in the regulation of dimorphic transition in the yeast Yarrowia lipolytica. FEMS Yeast Res. 2014, 14, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Morales-Vargas, A.T.; Domínguez, A.; Ruiz-Herrera, J. Identification of dimorphism-involved genes of Yarrowia lipolytica by means of microarray analysis. Res. Microbiol. 2012, 163, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Torres-Guzman, J.C.; Domínguez, A. HOY1, a homeo gene required for hyphal formation in Yarrowia lipolytica. Mol. Cell. Biol. 1997, 17, 6283–6293. [Google Scholar] [CrossRef] [PubMed]
- Szabo, R. Cla4 protein kinase is essential for filament formation and invasive growth of Yarrowia lipolytica. Mol. Genet. Genom. 2001, 265, 172–179. [Google Scholar] [CrossRef]
- Vandermies, M.; Kar, T.; Carly, F.; Nicaud, J.-M.; Delvigne, F.; Fickers, P. Yarrowia lipolytica morphological mutant enables lasting in situ immobilization in bioreactor. Appl. Microbiol. Biotechnol. 2018, 102, 5473–5482. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, S.; Yang, X.; Lin, C.S.K. Green and sustainable succinic acid production from crude glycerol by engineered Yarrowia lipolytica via agricultural residue based in situ fibrous bed bioreactor. Bioresour. Technol. 2018, 249, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, S.; Li, X.; Yang, X.; Lin, C.S.K. Efficient metabolic evolution of engineered Yarrowia lipolytica for succinic acid production using a glucose-based medium in an in situ fibrous bioreactor under low-pH condition. Biotechnol. Biofuels 2018, 11, 236. [Google Scholar] [PubMed]
- Darvishi, F.; Moradi, M.; Jolivalt, C.; Madzak, C. Laccase production from sucrose by recombinant Yarrowia lipolytica and its application to decolorization of environmental pollutant dyes. Ecotoxicol. Environ. Saf. 2018, 165, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Darvishi, F.; Moradi, M.; Madzak, C.; Jolivalt, C. Production of Laccase by Recombinant Yarrowia lipolytica from Molasses: Bioprocess Development Using Statistical Modeling and Increase Productivity in Shake-Flask and Bioreactor Cultures. Appl. Biochem. Biotechnol. 2017, 181, 1228–1239. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Borkowska, M.; Białas, W. Enhanced production of insect raw-starch-digesting alpha-amylase accompanied by high erythritol synthesis in recombinant Yarrowia lipolytica fed-batch cultures at high-cell-densities. Process Biochem. 2017, 52, 78–85. [Google Scholar] [CrossRef]
- Van Zyl, P.J. A Method for Producing a Polypeptide in Yarrowia lipolytica. World Patent Application WO2011148339 A1, 1 December 2011. [Google Scholar]
- Park, C.S.; Chang, C.C.; Ryu, D.D.Y. Expression and High-Level Secretion of Trichoderma reesei Endoglucanase I in Yarrowia lipolytica. Appl. Biochem. Biotechnol. 2000, 87, 1–16. [Google Scholar] [CrossRef]
- Theron, C.W.; Berrios, J.; Delvigne, F.; Fickers, P. Integrating metabolic modeling and population heterogeneity analysis into optimizing recombinant protein production by Komagataella (Pichia) pastoris. Appl. Microbiol. Biotechnol. 2018, 102, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Y.; Jian, X.-X.; Lv, Y.-B.; Nian, K.-Q.; Gao, Q.; Chen, J.; Wei, L.-J.; Hua, Q. Enhanced squalene biosynthesis in Yarrowia lipolytica based on metabolically engineered acetyl-CoA metabolism. J. Biotechnol. 2018, 281, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Loira, N.; Dulermo, T.; Nicaud, J.-M.; Sherman, D.J. A genome-scale metabolic model of the lipid-accumulating yeast Yarrowia lipolytica. BMC Syst. Biol. 2012, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Lee, N.-R.; Lakshmanan, M.; Kim, M.; Kim, B.-G.; Lee, D.-Y. Genome-scale model-driven strain design for dicarboxylic acid production in Yarrowia lipolytica. BMC Syst. Biol. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Jian, X.; Chen, J.; Zhang, C.; Hua, Q. Reconstruction of genome-scale metabolic model of Yarrowia lipolytica and its application in overproduction of triacylglycerol. Bioresour. Bioprocess. 2017, 4, 51. [Google Scholar] [CrossRef]
- Barth, G.; Gaillardin, C. Physiology and genetics of the dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 1997, 19, 219–237. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Lazar, Z.; Rakicka, M.; Guo, Z.; Fouchard, F.; Crutz-Le Coq, A.-M.; Nicaud, J.-M. Metabolic engineering of Yarrowia lipolytica to produce chemicals and fuels from xylose. Metab. Eng. 2016, 38, 115–124. [Google Scholar] [CrossRef]
- Spagnuolo, M.; Shabbir Hussain, M.; Gambill, L.; Blenner, M. Alternative Substrate Metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef]
- Celińska, E.; Borkowska, M.; Białas, W. Evaluation of heterologous α-amylase production in two expression platforms dedicated for Yarrowia lipolytica: Commercial Po1g–pYLSC (php4d) and custom-made A18–pYLTEF (pTEF). Yeast 2016, 33, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Fakas, S.; Komaitis, M.; Aggelis, G. Citric acid production by Yarrowia lipolytica cultivated on olive-mill wastewater-based media. Bioresour. Technol. 2008, 99, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Katre, G.; Ajmera, N.; Zinjarde, S.; RaviKumar, A. Mutants of Yarrowia lipolytica NCIM 3589 grown on waste cooking oil as a biofactory for biodiesel production. Microb. Cell Fact. 2017, 16, 176. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Robin, J.; Duquesne, S.; O’Donohue, M.J.; Marty, A.; Bordes, F. Developing cellulolytic Yarrowia lipolytica as a platform for the production of valuable products in consolidated bioprocessing of cellulose. Biotechnol. Biofuels 2018, 11, 141. [Google Scholar]
- Lazar, Z.; Gamboa-Meléndez, H.; Le Coq, A.-M.C.; Neuvéglise, C.; Nicaud, J.-M. Awakening the endogenous Leloir pathway for efficient galactose utilization by Yarrowia lipolytica. Biotechnol. Biofuels 2015, 8, 185. [Google Scholar] [PubMed]
- Ledesma-Amaro, R.; Dulermo, T.; Nicaud, J.M. Engineering Yarrowia lipolytica to produce biodiesel from raw starch. Biotechnol. Biofuels 2015, 8, 1. [Google Scholar]
- Li, H.; Alper, H.S. Enabling xylose utilization in Yarrowia lipolytica for lipid production. Biotechnol. J. 2016, 11, 1230–1240. [Google Scholar] [CrossRef]
- Aunins, J.G.; Henzler, H.-J. Aeration in Cell Culture Bioreactors. In Biotechnology; John Wiley & Sons, Ltd.: New York, NY, USA, 2008; pp. 219–281. ISBN 978-3-527-62084-5. [Google Scholar]
- Routledge, S.J. Beyond de-foaming: The effects of antifoams on bioprocess productivity. Comput. Struct. Biotechnol. J. 2012, 3, e201210014. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; Sentandreu, R. Different effectors of dimorphism in Yarrowia lipolytica. Arch. Microbiol. 2002, 178, 477–483. [Google Scholar] [CrossRef]
- Kar, T.; Delvigne, F.; Masson, M.; Destain, J.; Thonart, P. Investigation of the effect of different extracellular factors on the lipase production by Yarrowia lipolytica on the basis of a scale-down approach. J. Ind. Microbiol. Biotechnol. 2008, 35, 1053–1059. [Google Scholar] [CrossRef]
- Kar, T.; Delvigne, F.; Destain, J.; Thonart, P. Bioreactor scale-up and design on the basis of physiologically relevant parameters: Application to the production of lipase by Yarrowia lipolytica. Biotechnol. Agron. Soc. Environ. 2011, 15, 585–595. [Google Scholar]
- Amaral, P.F.F.; Freire, M.G.; Rocha-Leão, M.H.M.; Marrucho, I.M.; Coutinho, J.A.P.; Coelho, M.A.Z. Optimization of oxygen mass transfer in a multiphase bioreactor with perfluorodecalin as a second liquid phase. Biotechnol. Bioeng. 2008, 99, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Biryukova, E.N.; Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Tolerance of the yeast Yarrowia lipolytica to oxidative stress. Microbiology 2006, 75, 243–247. [Google Scholar] [CrossRef]
- Lopes, M.; Mota, M.; Belo, I. Comparison of Yarrowia lipolytica and Pichia pastoris Cellular Response to Different Agents of Oxidative Stress. Appl. Biochem. Biotechnol. 2013, 170, 448–458. [Google Scholar] [CrossRef]
- Lopes, M.; Gomes, N.; Mota, M.; Belo, I. Yarrowia lipolytica Growth Under Increased Air Pressure: Influence on Enzyme Production. Appl. Biochem. Biotechnol. 2009, 159, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Ryu, S.; Trinh, C.T. Exceptional Solvent Tolerance in Yarrowia lipolytica Is Enhanced by Sterols. bioRxiv 2018. [Google Scholar] [CrossRef]
- United Nations Framework Convention on Climate Change. Kyoto Protocol Reference Manual on Accounting of Emissions and Assigned Amounts; United Nations Framework Convention on Climate Change: Rio de Janeiro, Brazil, 2008.
- Egermeier, M.; Russmayer, H.; Sauer, M.; Marx, H. Metabolic Flexibility of Yarrowia lipolytica Growing on Glycerol. Front. Microbiol. 2017, 8, 49. [Google Scholar] [CrossRef]
- Kebabci, Ö.; Ciïhangïr, N. Comparison of three Yarrowia lipolytica strains for lipase production: NBRC 1658, IFO 1195, and a local strain. Turk. J. Biol. 2012, 36, 15–24. [Google Scholar]
- Young, T.W.; Wadeson, A.; Glover, D.J.; Quincey, R.V.; Butlin, M.J.; Kamei, E.A. The extracellular acid protease gene of Yarrowia lipolytica: Sequence and pH-regulated transcription. Microbiology 1996, 142, 2913–2921. [Google Scholar] [CrossRef]
- Pinsach, J.; de Mas, C.; López-Santín, J. Induction strategies in fed-batch cultures for recombinant protein production in Escherichia coli: Application to rhamnulose 1-phosphate aldolase. Biochem. Eng. J. 2008, 41, 181–187. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Du, G.; Hua, Z.; Liu, L.; Li, J.; Chen, J. Enhancement of alkaline polygalacturonate lyase production in recombinant Pichia pastoris according to the ratio of methanol to cell concentration. Bioresour. Technol. 2009, 100, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Carly, F.; Steels, S.; Telek, S.; Vandermies, M.; Nicaud, J.-M.; Fickers, P. Identification and characterization of EYD1, encoding an erythritol dehydrogenase in Yarrowia lipolytica and its application to bioconvert erythritol into erythrulose. Bioresour. Technol. 2018, 247, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Destain, J.; Fickers, P.; Molitor, J.-P.; Moreau, B.; Thonart, P.; Weekers, F.; Weiss, A.D. Method for the Production of Lipase. European Patent Application, Eur. Patent 11 October 2004. [Google Scholar]
- Destain, J.; Roblain, D.; Thonart, P. Improvement of lipase production from Yarrowia lipolytica. Biotechnol. Lett. 1997, 19, 105–108. [Google Scholar] [CrossRef]
- Destain, J.; Fickers, P.; Weekers, F.; Moreau, B.; Thonart, P. Utilization of methyloleate in production of microbial lipase. Appl. Biochem. Biotechnol. 2005, 121, 9. [Google Scholar] [CrossRef]
- Fickers, P.; Ongena, M.; Destain, J.; Weekers, F.; Thonart, P. Production and down-stream processing of an extracellular lipase from the yeast Yarrowia lipolytica. Enzym. Microb. Technol. 2006, 38, 756–759. [Google Scholar] [CrossRef]
- Gull, I.; Samra, Z.Q.; Aslam, M.S.; Athar, M.A. Heterologous expression, immunochemical and computational analysis of recombinant human interferon alpha 2b. Springer Plus 2013, 2, 264. [Google Scholar] [CrossRef]
- Müller, S.; Sandal, T.; Kamp-Hansen, P.; Dalbøge, H. Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast 1998, 14, 1267–1283. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandermies, M.; Fickers, P. Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica. Microorganisms 2019, 7, 40. https://doi.org/10.3390/microorganisms7020040
Vandermies M, Fickers P. Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica. Microorganisms. 2019; 7(2):40. https://doi.org/10.3390/microorganisms7020040
Chicago/Turabian StyleVandermies, Marie, and Patrick Fickers. 2019. "Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica" Microorganisms 7, no. 2: 40. https://doi.org/10.3390/microorganisms7020040
APA StyleVandermies, M., & Fickers, P. (2019). Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica. Microorganisms, 7(2), 40. https://doi.org/10.3390/microorganisms7020040