
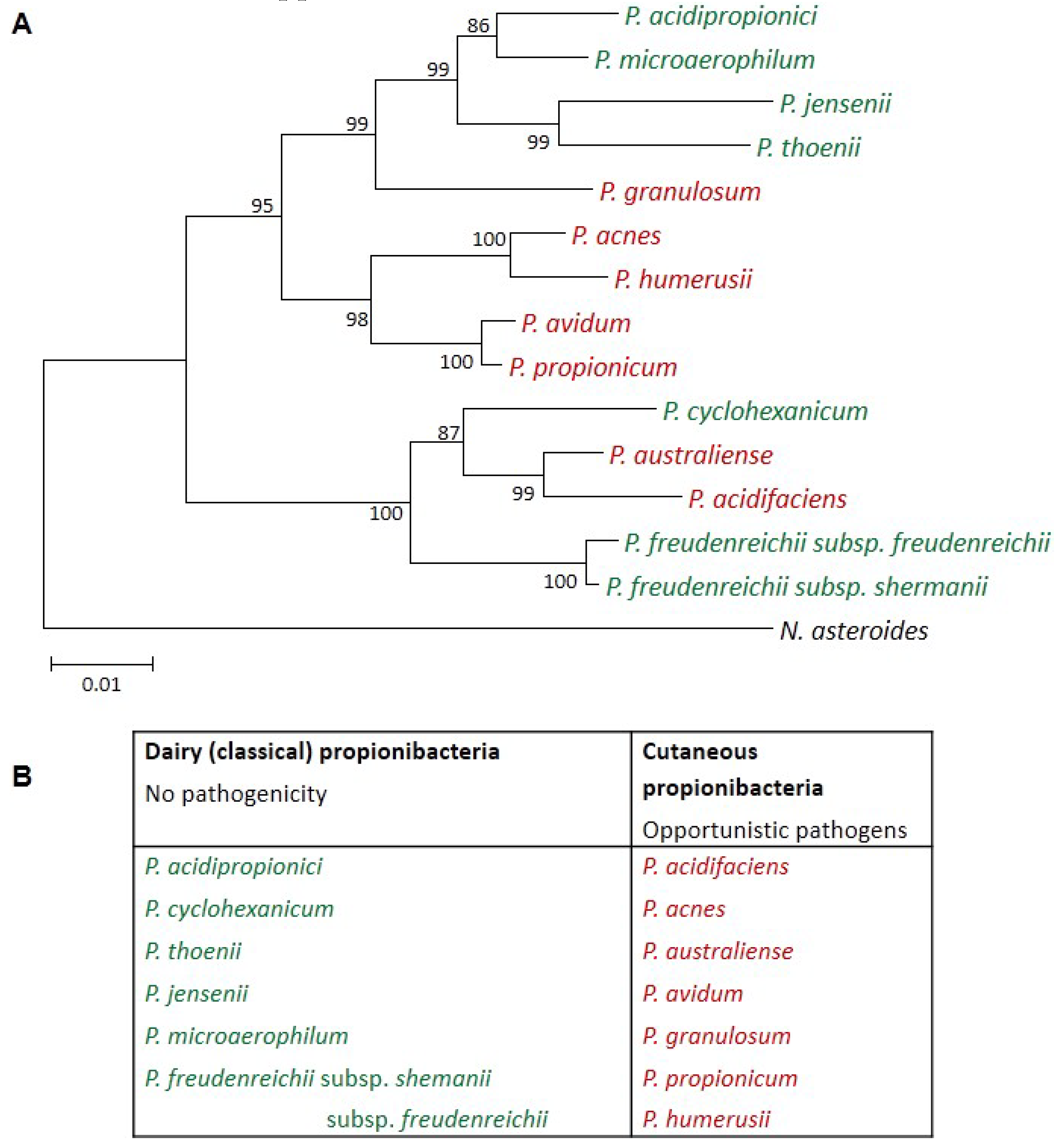
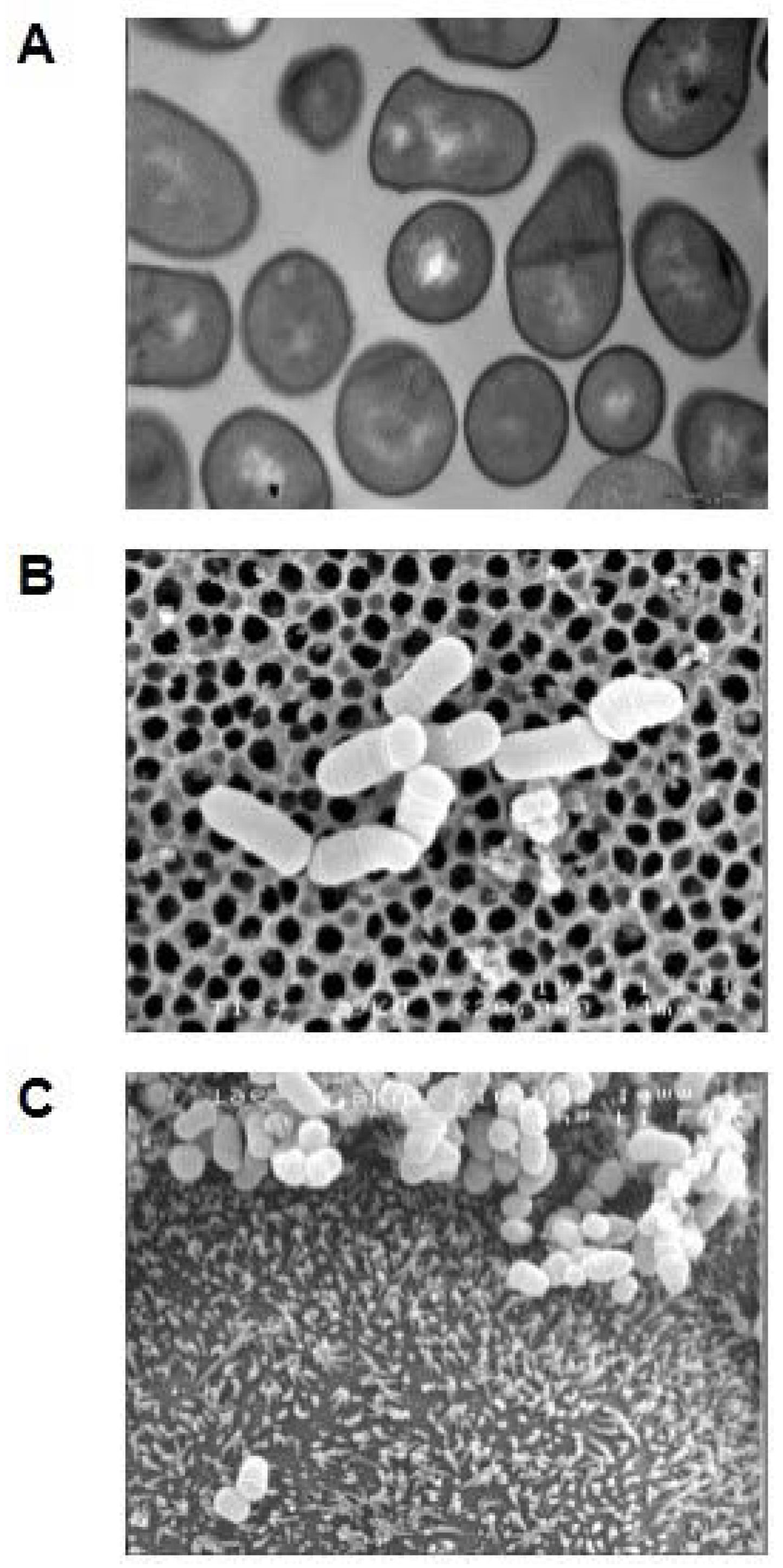
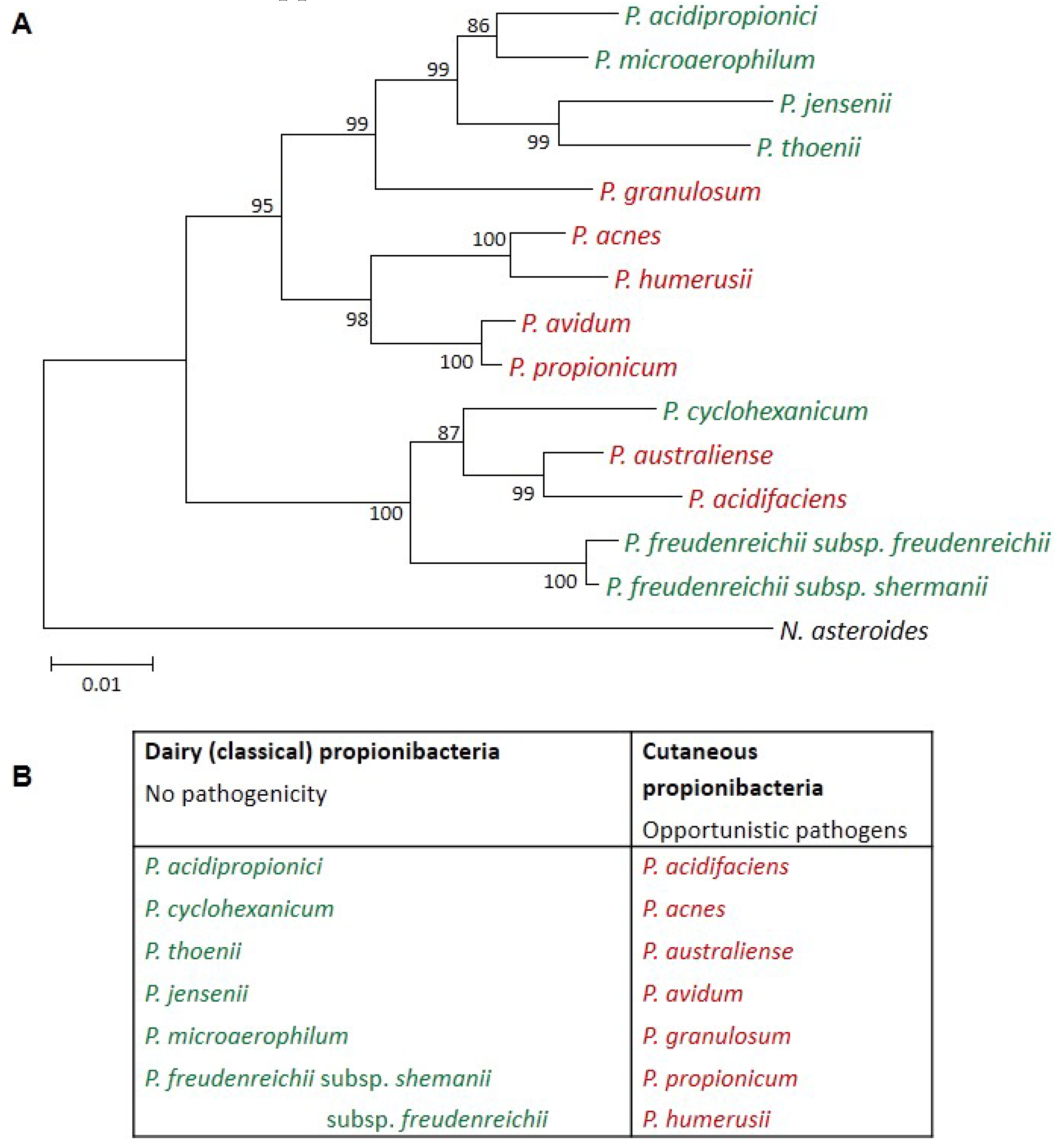
Dairy Propionibacteria: Versatile Probiotics

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Gut Persistence of Dairy Propionibacteria
2.1. Digestive Stress Tolerance
2.2. Adhesion to the Gut Epithelium
3. Microbiota Modulation by Dairy Propionibacteria
4. Immunomodulation by Dairy Propionibacteria
4.1. Surface Layer Proteins
4.2. Short Chain Fatty acids (SCFAs)
4.3. Conjugated Fatty Acids
4.4. DHNA
5. Anti-Cancerous Effect
5.1. Short Fatty Acids
5.2. Conjugated Fatty Acids
5.3. Bioremediation against Carcinogenic Components
6. Impact of Vectorization on Probiotic Properties
7. Technological Applications of Dairy Propionibacteria
7.1. Swiss-Cheese Manufacturing
7.2. Production of Nutritional Molecules
7.3. Production of Antimicrobial Molecules
8. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thierry, A.; Falentin, H.; Deutsch, S.M.; Jan, G. Propionibacterium spp. In Encyclopedia of Dairy Sciences; Fox, P.F., McSweeney, P.L., Fuquay, J.W., Eds.; Academic Press: San Diego, CA, USA, 2011; Volume 1, pp. 403–411. [Google Scholar]
- McDowell, A.; Nagy, I.; Magyari, M.; Barnard, E.; Patrick, S. The Opportunistic Pathogen Propionibacterium acnes: Insights into Typing, Human Disease, Clonal Diversification and CAMP Factor Evolution. PLoS ONE 2013, 8, e70897. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ) Scientific Opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed (2013 update): QPS 2013 update. EFSA J. 2013, 11, 3449.
- Food and Agriculture Organization. WHO Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO: Rome, Italy, 2006. [Google Scholar]
- Cousin, F.J.; Mater, D.D.G.; Foligne, B.; Jan, G. Dairy propionibacteria as human probiotics: A review of recent evidence. Dairy Sci. Technol. 2010, 91, 1–26. [Google Scholar] [CrossRef]
- Altieri, C. Dairy propionibacteria as probiotics: Recent evidences. World J. Microbiol. Biotechnol. 2016, 32, 172. [Google Scholar] [CrossRef] [PubMed]
- Plé, C.; Richoux, R.; Jardin, J.; Nurdin, M.; Briard-Bion, V.; Parayre, S.; Ferreira, S.; Pot, B.; Bouguen, G.; Deutsch, S.-M.; et al. Single-strain starter experimental cheese reveals anti-inflammatory effect of Propionibacterium freudenreichii CIRM BIA 129 in TNBS-colitis model. J. Funct. Foods 2015, 18, 575–585. [Google Scholar] [CrossRef]
- Cousin, F.J.; Foligné, B.; Deutsch, S.-M.; Massart, S.; Parayre, S.; Le Loir, Y.; Boudry, G.; Jan, G. Assessment of the Probiotic Potential of a Dairy Product Fermented by Propionibacterium freudenreichii in Piglets. J. Agric. Food Chem. 2012, 60, 7917–7927. [Google Scholar] [CrossRef] [PubMed]
- Cousin, F.J.; Louesdon, S.; Maillard, M.-B.; Parayre, S.; Falentin, H.; Deutsch, S.-M.; Boudry, G.; Jan, G. The first dairy product exclusively fermented by Propionibacterium freudenreichii: A new vector to study probiotic potentialities in vivo. Food Microbiol. 2012, 32, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Cousin, F.J.; Jouan-Lanhouet, S.; Théret, N.; Brenner, C.; Jouan, E.; Moigne-Muller, G.L.; Dimanche-Boitrel, M.-T.; Jan, G.; Cousin, F.J.; Jouan-Lanhouet, S.; et al. The probiotic Propionibacterium freudenreichii as a new adjuvant for TRAIL-based therapy in colorectal cancer. Oncotarget 2016, 7, 7161–7178. [Google Scholar] [PubMed]
- Okada, Y.; Tsuzuki, Y.; Narimatsu, K.; Sato, H.; Ueda, T.; Hozumi, H.; Sato, S.; Hokari, R.; Kurihara, C.; Komoto, S.; et al. 1,4-Dihydroxy-2-naphthoic acid from Propionibacterium freudenreichii reduces inflammation in interleukin-10-deficient mice with colitis by suppressing macrophage-derived proinflammatory cytokines. J. Leukoc. Biol. 2013, 94, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Plé, C.; Breton, J.; Richoux, R.; Nurdin, M.; Deutsch, S.-M.; Falentin, H.; Hervé, C.; Chuat, V.; Lemée, R.; Maguin, E.; et al. Combining selected immunomodulatory Propionibacterium freudenreichii and Lactobacillus delbrueckii strains: Reverse engineering development of an anti-inflammatory cheese. Mol. Nutr. Food Res. 2016, 60, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y. Propionibacterium freudenreichii component 1.4-dihydroxy-2-naphthoic acid (DHNA) attenuates dextran sodium sulphate induced colitis by modulation of bacterial flora and lymphocyte homing. Gut 2006, 55, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Falentin, H.; Deutsch, S.-M.; Jan, G.; Loux, V.; Thierry, A.; Parayre, S.; Maillard, M.-B.; Dherbécourt, J.; Cousin, F.J.; Jardin, J.; et al. The Complete Genome of Propionibacterium freudenreichii CIRM-BIA1T, a Hardy Actinobacterium with Food and Probiotic Applications. PLoS ONE 2010, 5, e11748. [Google Scholar] [CrossRef] [PubMed]
- Parizzi, L.P.; Grassi, M.C.B.; Llerena, L.A.; Carazzolle, M.F.; Queiroz, V.L.; Lunardi, I.; Zeidler, A.F.; Teixeira, P.J.; Mieczkowski, P.; Rincones, J.; et al. The genome sequence of Propionibacterium acidipropionici provides insights into its biotechnological and industrial potential. BMC Genom. 2012, 13, 562. [Google Scholar] [CrossRef] [PubMed]
- Amund, O.D. Exploring the relationship between exposure to technological and gastrointestinal stress and probiotic functional properties of lactobacilli and bifidobacteria. Can. J. Microbiol. 2016, 62, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, B.; Stanton, C.; Fitzgerald, G.; Ross, R. Life under Stress: The Probiotic Stress Response and How it may be Manipulated. Curr. Pharm. Des. 2008, 14, 1382–1399. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Adams, M.C. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria. Int. J. Food Microbiol. 2004, 91, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Campaniello, D.; Bevilacqua, A.; Sinigaglia, M.; Altieri, C. Screening of Propionibacterium spp. for potential probiotic properties. Anaerobe 2015, 34, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, V.; Jardin, J.; Rabah, H.; Briard-Bion, V.; Jan, G. Emmental Cheese Environment Enhances Propionibacterium freudenreichii Stress Tolerance. PLoS ONE 2015, 10, e0135780. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, A.; Brede, M.E.; Vegarud, G.E.; Østlie, H.M.; Narvhus, J.; Skeie, S.B. Survival of lactic acid and propionibacteria in low- and full-fat Dutch-type cheese during human digestion ex vivo. Lett. Appl. Microbiol. 2016, 62, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Rabah, H.; Jardin, J.; Briard-Bion, V.; Parayre, S.; Maillard, M.-B.; Le Loir, Y.; Chen, X.D.; Schuck, P.; Jeantet, R.; et al. Hyperconcentrated Sweet Whey, a New Culture Medium That Enhances Propionibacterium freudenreichii Stress Tolerance. Appl. Environ. Microbiol. 2016, 82, 4641–4651. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. Effect of dairy probiotic combinations on in vitro gastrointestinal tolerance, intestinal epithelial cell adhesion and cytokine secretion. J. Funct. Foods 2014, 8, 18–25. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. In vitro analysis of gastrointestinal tolerance and intestinal cell adhesion of probiotics in goat’s milk ice cream and yogurt, In vitro analysis of gastrointestinal tolerance and intestinal cell adhesion of probiotics in goat’s milk ice cream and yogurt. Food Res. Int. 2012, 49, 619–625. [Google Scholar] [CrossRef]
- Anastasiou, R.; Leverrier, P.; Krestas, I.; Rouault, A.; Kalantzopoulos, G.; Boyaval, P.; Tsakalidou, E.; Jan, G. Changes in protein synthesis during thermal adaptation of Propionibacterium freudenreichii subsp. shermanii. Int. J. Food Microbiol. 2006, 108, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Leverrier, P.; Pichereau, V.; Boyaval, P. Changes in Protein Synthesis and Morphology during Acid Adaptation of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2001, 67, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Leverrier, P.; Dimova, D.; Pichereau, V.; Auffray, Y.; Boyaval, P.; Jan, G. Susceptibility and Adaptive Response to Bile Salts in Propionibacterium freudenreichii: Physiological and Proteomic Analysis. Appl. Environ. Microbiol. 2003, 69, 3809–3818. [Google Scholar] [CrossRef] [PubMed]
- Leverrier, P.; Vissers, J.P.C.; Rouault, A.; Boyaval, P.; Jan, G. Mass spectrometry proteomic analysis of stress adaptation reveals both common and distinct response pathways in Propionibacterium freudenreichii. Arch. Microbiol. 2004, 181, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Guan, N.; Shin, H.; Chen, R.R.; Li, J.; Liu, L.; Du, G.; Chen, J. Understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the level of proteomics. Sci. Rep. 2014, 4, 6951. [Google Scholar] [CrossRef] [PubMed]
- Guan, N.; Liu, L.; Shin, H.; Chen, R.R.; Zhang, J.; Li, J.; Du, G.; Shi, Z.; Chen, J. Systems-level understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the microenvironment levels: Mechanism and application. J. Biotechnol. 2013, 167, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.S.; Gaspar, P.; Hugenholtz, J.; Ramos, A.; Santos, H. Enhancement of trehalose production in dairy propionibacteria through manipulation of environmental conditions. Int. J. Food Microbiol. 2004, 91, 195–204. [Google Scholar] [CrossRef]
- Lan, A.; Bruneau, A.; Philippe, C.; Rochet, V.; Rouault, A.; Hervé, C.; Roland, N.; Rabot, S.; Jan, G. Survival and metabolic activity of selected strains of Propionibacterium freudenreichii in the gastrointestinal tract of human microbiota-associated rats. Br. J. Nutr. 2007, 97, 714. [Google Scholar] [CrossRef] [PubMed]
- Argañaraz-Martínez, E.; Babot, J.D.; Apella, M.C.; Perez Chaia, A. Physiological and functional characteristics of Propionibacterium strains of the poultry microbiota and relevance for the development of probiotic products. Anaerobe 2013, 23, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hervé, C.; Fondrevez, M.; Chéron, A.; Barloy-Hubler, F.; Jan, G. Transcarboxylase mRNA: A marker which evidences P. freudenreichii survival and metabolic activity during its transit in the human gut. Int. J. Food Microbiol. 2007, 113, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Saraoui, T.; Parayre, S.; Guernec, G.; Loux, V.; Montfort, J.; Le Cam, A.; Boudry, G.; Jan, G.; Falentin, H. A unique in vivo experimental approach reveals metabolic adaptation of the probiotic Propionibacterium freudenreichii to the colon environment. BMC Genom. 2013, 14, 911. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G.; Palacios, J.M.; Villena, J.; Zúñiga-Hansen, M.E. Inhibition of enteropathogens adhesion to human enterocyte-like HT-29 cells by a dairy strain of Propionibacterium acidipropionici. Benef. Microbes 2016, 7, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G.; Morata De Ambrosini, V.; Perez Chaia, A.; González, S. Some factors affecting the adherence of probiotic Propionibacterium acidipropionici CRL 1198 to intestinal epithelial cells. Can. J. Microbiol. 2002, 48, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Tölkkö, S.; Kulmala, J.; Salminen, S.; Salminen, E. Adhesion of inactivated probiotic strains to intestinal mucus. Lett. Appl. Microbiol. 2000, 31, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Tuomola, E.M.; Ouwehand, A.C.; Salminen, S.J. Human ileostomy glycoproteins as a model for small intestinal mucus to investigate adhesion of probiotics. Lett. Appl. Microbiol. 1999, 28, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Ganan, M.; Martinez-Rodriguez, A.J.; Carrascosa, A.V.; Vesterlund, S.; Salminen, S.; Satokari, R. Interaction of Campylobacter spp. and Human Probiotics in Chicken Intestinal Mucus: Adhesion of Campylobacter and Interaction with Probiotics. Zoonoses Public Health 2013, 60, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Darilmaz, D.O.; Beyatli, Y.; Yuksekdag, Z.N. Aggregation and Hydrophobicity Properties of 6 Dairy Propionibacteria Strains Isolated from Homemade Turkish Cheeses. J. Food Sci. 2012, 77, M20–M24. [Google Scholar] [CrossRef] [PubMed]
- Vesterlund, S. Staphylococcus aureus adheres to human intestinal mucus but can be displaced by certain lactic acid bacteria. Microbiology 2006, 152, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Hajfarajollah, H.; Mokhtarani, B.; Noghabi, K.A. Newly Antibacterial and Antiadhesive Lipopeptide Biosurfactant Secreted by a Probiotic Strain, Propionibacterium freudenreichii. Appl. Biochem. Biotechnol. 2014, 174, 2725–2740. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.V.T.; Kollanoor-Johny, A. Effect of Propionibacterium freudenreichii on Salmonella multiplication, motility, and association with avian epithelial cells. Poult. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Le Maréchal, C.; Peton, V.; Plé, C.; Vroland, C.; Jardin, J.; Briard-Bion, V.; Durant, G.; Chuat, V.; Loux, V.; Foligné, B.; et al. Surface proteins of Propionibacterium freudenreichii are involved in its anti-inflammatory properties. J. Proteom. 2015, 113, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Falentin, H.; Deutsch, S.-M.; Loux, V.; Hammani, A.; Buratti, J.; Parayre, S.; Chuat, V.; Barbe, V.; Aury, J.-M.; Jan, G.; et al. Permanent draft genome sequence of the probiotic strain Propionibacterium freudenreichii CIRM-BIA 129 (ITG P20). Stand. Genom. Sci. 2016, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.E.; Anderson, S.E.; Corbett, A.H.; Lamb, T.J. Gleaning Insights from Fecal Microbiota Transplantation and Probiotic Studies for the Rational Design of Combination Microbial Therapies. Clin. Microbiol. Rev. 2017, 30, 191–231. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Toshimitsu, T.; Matsuoka, S.; Maruyama, A.; Oh-oka, K.; Takamura, T.; Nakamura, Y.; Ishimaru, K.; Fujii-Kuriyama, Y.; Ikegami, S.; et al. Identification of a probiotic bacteria-derived activator of the aryl hydrocarbon receptor that inhibits colitis. Immunol. Cell Biol. 2014, 92, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, K.; Masuda, J.; Yamasaki, H.; Kuwaki, K.; Kitazaki, S.; Koga, H.; Uchida, M.; Sata, M. Treatment of Ulcerative Colitis with Milk Whey Culture with Propionibacterium freudenreichii. J. Intest. Microbiol. 2007, 21, 143–147. [Google Scholar]
- Hojo, K.; Yoda, N.; Tsuchita, H.; Ohtsu, T.; Seki, K.; Taketomo, N.; Murayama, T.; Iino, H. Effect of Ingested Culture of Propionibacterium freudenreichii ET-3 on Fecal Microflora and Stool Frequency in Healthy Females. Biosci. Microflora 2002, 21, 115–120. [Google Scholar] [CrossRef]
- Seki, K.; Nakao, H.; Umino, H.; Isshiki, H.; Yoda, N.; Tachihara, R.; Ohuchi, T.; Saruta, H.; Suzuki, K.; Mitsuoka, T. Effects of Fermented Milk Whey Containing Novel Bifidogenic Growth Stimulator Produced by Propionibacterium on Fecal Bacteria, Putrefactive Metabolite, Defecation Frequency and Fecal Properties in Senile Volunteers Needed Serious Nursing-Care Taking Enteral Nutrition by Tube Feeding. J. Intest. Microbiol. 2004, 18, 107–115. [Google Scholar]
- Martínez, E.A.; Babot, J.D.; Lorenzo-Pisarello, M.J.; Apella, M.C.; Chaia, A.P. Feed supplementation with avian Propionibacterium acidipropionici contributes to mucosa development in early stages of rearing broiler chickens. Benef. Microbes 2016, 7, 687–698. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed]
- Isawa, K.; Hojo, K.; Yoda, N.; Kamiyama, T.; Makino, S.; Saito, M.; Sugano, H.; Mizoguchi, C.; Kurama, S.; Shibasaki, M.; et al. Isolation and Identification of a New Bifidogenic Growth Stimulator Produced by Propionibacterium freudenreichii ET-3. Biosci. Biotechnol. Biochem. 2002, 66, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, K.; Hojo, K.; Katakura, Y.; Ninomiya, K.; Shioya, S. Aerobic culture of Propionibacterium freudenreichii ET-3 can increase production ratio of 1,4-dihydroxy-2-naphthoic acid to menaquinone. J. Biosci. Bioeng. 2006, 101, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T. A Novel Bifidogenic Growth Stimulator Produced by Propionibacterium freudenreichii. Biosci. Microflora 1999, 18, 73–80. [Google Scholar] [CrossRef]
- Yamazaki, S.; Kano, K.; Ikeda, T.; Isawa, K.; Kaneko, T. Role of 2-amino-3-carboxy-1,4-naphthoquinone, a strong growth stimulator for bifidobacteria, as an electron transfer mediator for NAD(P)+ regeneration in Bifidobacterium longum. Biochim. Biophys. Acta BBA Gen. Subj. 1999, 1428, 241–250. [Google Scholar] [CrossRef]
- Kaneko, T.; Mori, H.; Iwata, M.; Meguro, S. Growth Stimulator for Bifidobacteria Produced by Propionibacterium freudenreichii and Several Intestinal Bacteria. J. Dairy Sci. 1994, 77, 393–404. [Google Scholar] [CrossRef]
- Uchida, M.; Tsuboi, H.; Takahashi, M.; Nemoto, A.; Seki, K.; Tsunoo, H.; Martyres, S.; Roberts, A. Safety of high doses of Propionibacterium freudenreichii ET-3 culture in healthy adult subjects. Regul. Toxicol. Pharmacol. 2011, 60, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Pisarello, M.J.; Gultemirian, M.L.; Nieto-Peñalver, C.; Perez Chaia, A. Propionibacterium acidipropionici CRL1198 influences the production of acids and the growth of bacterial genera stimulated by inulin in a murine model of cecal slurries. Anaerobe 2010, 16, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Ranadheera, C.S.; King, S.; Evans, C.A.; Baines, S.K. Potential influence of dairy propionibacteria on the growth and acid metabolism of Streptococcus bovis and Megasphaera elsdenii. Benef. Microbes 2016, 8, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, A.; Zielińska, M.; Storr, M.; Fichna, J. Beneficial Effects of Probiotics, Prebiotics, Synbiotics, and Psychobiotics in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- DuPont, A.; Richards, D.M.; Jelinek, K.A.; Krill, J.; Rahimi, E.; Ghouri, Y. Systematic review of randomized controlled trials of probiotics, prebiotics, and synbiotics in inflammatory bowel disease. Clin. Exp. Gastroenterol. 2014, 7, 473. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; Breton, J.; Mater, D.; Jan, G. Tracking the microbiome functionality: Focus on Propionibacterium species. Gut 2013, 62, 1227–1228. [Google Scholar] [CrossRef] [PubMed]
- Foligne, B.; Deutsch, S.-M.; Breton, J.; Cousin, F.J.; Dewulf, J.; Samson, M.; Pot, B.; Jan, G. Promising Immunomodulatory Effects of Selected Strains of Dairy Propionibacteria as Evidenced In Vitro and In Vivo. Appl. Environ. Microbiol. 2010, 76, 8259–8264. [Google Scholar] [CrossRef] [PubMed]
- Oksaharju, A.; Kooistra, T.; Kleemann, R.; van Duyvenvoorde, W.; Miettinen, M.; Lappalainen, J.; Lindstedt, K.A.; Kovanen, P.T.; Korpela, R.; Kekkonen, R.A. Effects of probiotic Lactobacillus rhamnosus GG and Propionibacterium freudenreichii ssp. shermanii JS supplementation on intestinal and systemic markers of inflammation in ApoE*3Leiden mice consuming a high-fat diet. Br. J. Nutr. 2013, 110, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Kajander, K.; Myllyluoma, E.; Rajilić-Stojanović, M.; Kyrönpalo, S.; Rasmussen, M.; Järvenpää, S.; Zoetendal, E.G.; De Vos, W.M.; Vapaatalo, H.; Korpela, R. Clinical trial: Multispecies probiotic supplementation alleviates the symptoms of irritable bowel syndrome and stabilizes intestinal microbiota. Aliment. Pharmacol. Ther. 2008, 27, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Mitsuyama, K.; Koga, H.; Tomiyasu, N.; Masuda, J.; Takaki, K.; Tsuruta, O.; Toyonaga, A.; Sata, M. Bifidogenic growth stimulator for the treatment of active ulcerative colitis: A pilot study. Nutrition 2006, 22, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Lortal, S.; Rouault, A.; Cesselin, B.; Sleytr, U.B. Paracrystalline surface layers of dairy propionibacteria. Appl. Environ. Microbiol. 1993, 59, 2369–2374. [Google Scholar] [PubMed]
- Fagan, R.P.; Fairweather, N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 2014, 12, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Ilk, N.; Egelseer, E.M.; Sleytr, U.B. S-layer fusion proteins—Construction principles and applications. Curr. Opin. Biotechnol. 2011, 22, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.-M.; Mariadassou, M.; Nicolas, P.; Parayre, S.; Le Guellec, R.; Chuat, V.; Peton, V.; Le Maréchal, C.; Burati, J.; Loux, V.; et al. Identification of proteins involved in the anti-inflammatory properties of Propionibacterium freudenreichii by means of a multi-strain study. Sci. Rep. 2017, 7, 46409. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.-M.; Parayre, S.; Bouchoux, A.; Guyomarc’h, F.; Dewulf, J.; Dols-Lafargue, M.; Bagliniere, F.; Cousin, F.J.; Falentin, H.; Jan, G.; et al. Contribution of Surface—Glucan Polysaccharide to Physicochemical and Immunomodulatory Properties of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2012, 78, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.M.; Bivic, P.L.; Herve, C.; Madec, M.N.; LaPointe, G.; Jan, G.; Loir, Y.L.; Falentin, H. Correlation of the Capsular Phenotype in Propionibacterium freudenreichii with the Level of Expression of gtf, a Unique Polysaccharide Synthase-Encoding Gene. Appl. Environ. Microbiol. 2010, 76, 2740–2746. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, Y.L.; Selle, K.; Yang, T.; Goh, Y.J.; Sahay, B.; Zadeh, M.; Owen, J.L.; Colliou, N.; Li, E.; Johannssen, T.; et al. SIGNR3-dependent immune regulation by Lactobacillus acidophilus surface layer protein A in colitis. EMBO J. 2015, 34, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.M.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [PubMed]
- Puertollano, E.; Kolida, S.; Yaqoob, P. Biological significance of short-chain fatty acid metabolism by the intestinal microbiome. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. Chapter Three—The Role of Short-Chain Fatty Acids in Health and Disease. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 121, pp. 91–119. [Google Scholar]
- Kim, C.H.; Park, J.; Kim, M. Gut microbiota-derived short-chain Fatty acids, T cells, and inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.A.; Singh, N.; Martin, P.M.; Thangaraju, M.; Ganapathy, V.; Waller, J.L.; Shi, H.; Robertson, K.D.; Munn, D.H.; Liu, K. Butyrate suppresses colonic inflammation through HDAC1-dependent Fas upregulation and Fas-mediated apoptosis of T cells. AJP Gastrointest. Liver Physiol. 2012, 302, G1405–G1415. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, F.M.; Shingfield, K.J.; Devillard, E.; Russell, W.R.; Wallace, R.J. Mechanism of conjugated linoleic acid and vaccenic acid formation in human faecal suspensions and pure cultures of intestinal bacteria. Microbiology 2009, 155, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Björck, L.; Fonden, R. Production of conjugated linoleic acid by dairy starter cultures. J. Appl. Microbiol. 1998, 85, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Boylston, T.D.; Glatz, B.A. Conjugated Linoleic Acid Content and Organoleptic Attributes of Fermented Milk Products Produced with Probiotic Bacteria. J. Agric. Food Chem. 2005, 53, 9064–9072. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, A.A.; Barrett, E.; Paul Ross, R.; Fitzgerald, G.F.; Devery, R.; Stanton, C. The Production of Conjugated α-Linolenic, γ-Linolenic and Stearidonic Acids by Strains of Bifidobacteria and Propionibacteria. Lipids 2012, 47, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, L.; Leroy, F.; De Vuyst, L.; De Smet, S.; Raes, K. Bacterial Production of Conjugated Linoleic and Linolenic Acid in Foods: A Technological Challenge. Crit. Rev. Food Sci. Nutr. 2015, 55, 1561–1574. [Google Scholar] [CrossRef] [PubMed]
- Rainio, A.; Vahvaselkä, M.; Suomalainen, T.; Laakso, S. Reduction of linoleic acid inhibition in production of conjugated linoleic acid by Propionibacterium freudenreichii ssp. shermanii. Can. J. Microbiol. 2001, 47, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Fontes, A.L.; Pimentel, L.L.; Simões, C.D.; Gomes, A.M.P.; Rodríguez-Alcalá, L.M. Evidences and perspectives in the utilization of CLNA isomers as bioactive compound in foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 2611–2622. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.-F.; Chen, X.-E.; Li, D. Conjugated linolenic acids and their bioactivities: A review. Food Funct. 2014, 5, 1360. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Chen, X.; Li, D. Modulation of Peroxisome Proliferator-Activated Receptor gamma (PPAR γ) by Conjugated Fatty Acid in Obesity and Inflammatory Bowel Disease. J. Agric. Food Chem. 2015, 63, 1883–1895. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, Y.; Kim, Y.J.; Park, Y. Conjugated Linoleic Acid: Potential Health Benefits as a Functional Food Ingredient. Annu. Rev. Food Sci. Technol. 2016, 7, 221–244. [Google Scholar] [CrossRef] [PubMed]
- Churruca, I.; Fernández-Quintela, A.; Portillo, M.P. Conjugated linoleic acid isomers: Differences in metabolism and biological effects. BioFactors 2009, 35, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Draper, E.; DeCourcey, J.; Higgins, S.C.; Canavan, M.; McEvoy, F.; Lynch, M.; Keogh, B.; Reynolds, C.; Roche, H.M.; Mills, K.H.G.; et al. Conjugated linoleic acid suppresses dendritic cell activation and subsequent Th17 responses. J. Nutr. Biochem. 2014, 25, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, A.A.; Ross, P.R.; Fitzgerald, G.F.; Stanton, C. Sources and Bioactive Properties of Conjugated Dietary Fatty Acids. Lipids 2016, 51, 377–397. [Google Scholar] [CrossRef] [PubMed]
- Viladomiu, M.; Hontecillas, R.; Bassaganya-Riera, J. Modulation of inflammation and immunity by dietary conjugated linoleic acid. Eur. J. Pharmacol. 2016, 785, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Interplay between diet, gut microbiota, epigenetic events, and colorectal cancer. Mol. Nutr. Food Res. 2017, 61, 1500902. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Belzacq, A.-S.; Haouzi, D.; Rouault, A.; Métivier, D.; Kroemer, G.; Brenner, C. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Lagadic-Gossmann, D.; Lemaire, C.; Brenner, C.; Jan, G. Acidic extracellular pH shifts colorectal cancer cell death from apoptosis to necrosis upon exposure to propionate and acetate, major end-products of the human probiotic propionibacteria. Apoptosis 2007, 12, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Cousin, F.J.; Jouan-Lanhouet, S.; Dimanche-Boitrel, M.-T.; Corcos, L.; Jan, G. Milk Fermented by Propionibacterium freudenreichii Induces Apoptosis of HGT-1 Human Gastric Cancer Cells. PLoS ONE 2012, 7, e31892. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Bruneau, A.; Bensaada, M.; Philippe, C.; Bellaud, P.; Rabot, S.; Jan, G. Increased induction of apoptosis by Propionibacterium freudenreichii TL133 in colonic mucosal crypts of human microbiota-associated rats treated with 1,2-dimethylhydrazine. Br. J. Nutr. 2008, 100, 1251. [Google Scholar] [CrossRef] [PubMed]
- Zarate, G.; Perez Chaia, A. Dairy bacteria remove in vitro dietary lectins with toxic effects on colonic cells. J. Appl. Microbiol. 2009, 106, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Collado, M.C.; El-Nezami, H.; Meriluoto, J.; Salminen, S. Combining strains of lactic acid bacteria may reduce their toxin and heavy metal removal efficiency from aqueous solution: Toxin removal with LAB combination. Lett. Appl. Microbiol. 2007, 46, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; El-Nezami, H.; Haskard, C.A.; Gratz, S.; Puong, K.Y.; Salminen, S.; Mykkänen, H. Kinetics of adsorption and desorption of aflatoxin B1 by viable and nonviable bacteria. J. Food Prot. 2003, 66, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.; Mykkänen, H.; El-Nezami, H. Aflatoxin B1 binding by a mixture of Lactobacillus and Propionibacterium: In vitro versus ex vivo. J. Food Prot. 2005, 68, 2470–2474. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.; Mykkanen, H.; Ouwehand, A.C.; Juvonen, R.; Salminen, S.; El-Nezami, H. Intestinal Mucus Alters the Ability of Probiotic Bacteria To Bind Aflatoxin B1 In Vitro. Appl. Environ. Microbiol. 2004, 70, 6306–6308. [Google Scholar] [CrossRef] [PubMed]
- El-Nezami, H.; Mykkänen, H.; Kankaanpää, P.; Salminen, S.; Ahokas, J. Ability of Lactobacillus and Propionibacterium strains to remove aflatoxin B, from the chicken duodenum. J. Food Prot. 2000, 63, 549–552. [Google Scholar] [CrossRef] [PubMed]
- El-Nezami, H.S.; Polychronaki, N.N.; Ma, J.; Zhu, H.; Ling, W.; Salminen, E.K.; Juvonen, R.O.; Salminen, S.J.; Poussa, T.; Mykkänen, H.M. Probiotic supplementation reduces a biomarker for increased risk of liver cancer in young men from Southern China. Am. J. Clin. Nutr. 2006, 83, 1199–1203. [Google Scholar] [PubMed]
- Kekkonen, R.A.; Holma, R.; Hatakka, K.; Suomalainen, T.; Poussa, T.; Adlercreutz, H.; Korpela, R. A Probiotic Mixture Including Galactooligosaccharides Decreases Fecal-Glucosidase Activity but Does Not Affect Serum Enterolactone Concentration in Men during a Two-Week Intervention. J. Nutr. 2011, 141, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, K.; Holma, R.; El-Nezami, H.; Suomalainen, T.; Kuisma, M.; Saxelin, M.; Poussa, T.; Mykkänen, H.; Korpela, R. The influence of Lactobacillus rhamnosus LC705 together with Propionibacterium freudenreichii ssp. shermanii JS on potentially carcinogenic bacterial activity in human colon. Int. J. Food Microbiol. 2008, 128, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Saxelin, M.; Lassig, A.; Karjalainen, H.; Tynkkynen, S.; Surakka, A.; Vapaatalo, H.; Järvenpää, S.; Korpela, R.; Mutanen, M.; Hatakka, K. Persistence of probiotic strains in the gastrointestinal tract when administered as capsules, yoghurt, or cheese. Int. J. Food Microbiol. 2010, 144, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Thierry, A.; Deutsch, S.-M.; Falentin, H.; Dalmasso, M.; Cousin, F.J.; Jan, G. New insights into physiology and metabolism of Propionibacterium freudenreichii. Int. J. Food Microbiol. 2011, 149, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Zárate, G. Dairy Propionibacteria: Less Conventional Probiotics to Improve the Human and Animal Health. In Probiotic in Animals; InTechOpen: Rijeka, Croatia, 2012; ISBN 978-953-51-0777-4. [Google Scholar]
- Yee, A.L.; Maillard, M.-B.; Roland, N.; Chuat, V.; Leclerc, A.; Pogačić, T.; Valence, F.; Thierry, A. Great interspecies and intraspecies diversity of dairy propionibacteria in the production of cheese aroma compounds. Int. J. Food Microbiol. 2014, 191, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.; Yamashita, M.; Kawaraichi, N.; Asegawa, R.; Ono, H.; Murooka, Y. Production of vitamin B12 in genetically engineered Propionibacterium freudenreichii. J. Biosci. Bioeng. 2004, 98, 167–173. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Z.; Jiao, Y.; Liu, S.; Wang, Y. Improved propionic acid and 5,6-dimethylbenzimidazole control strategy for vitamin B12 fermentation by Propionibacterium freudenreichii. J. Biotechnol. 2015, 193, 123–129. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabah, H.; Rosa do Carmo, F.L.; Jan, G. Dairy Propionibacteria: Versatile Probiotics. Microorganisms 2017, 5, 24. https://doi.org/10.3390/microorganisms5020024
Rabah H, Rosa do Carmo FL, Jan G. Dairy Propionibacteria: Versatile Probiotics. Microorganisms. 2017; 5(2):24. https://doi.org/10.3390/microorganisms5020024
Chicago/Turabian StyleRabah, Houem, Fillipe Luiz Rosa do Carmo, and Gwénaël Jan. 2017. "Dairy Propionibacteria: Versatile Probiotics" Microorganisms 5, no. 2: 24. https://doi.org/10.3390/microorganisms5020024
APA StyleRabah, H., Rosa do Carmo, F. L., & Jan, G. (2017). Dairy Propionibacteria: Versatile Probiotics. Microorganisms, 5(2), 24. https://doi.org/10.3390/microorganisms5020024