Abstract
Maize (Zea mays L.) is an important cereal crop species for food, feedstock and industrial material. Maize seeds host a suitable ecosystem for endophytic bacteria, facilitating seed germination and seedling growth. However, the inheritance, diversity and potential function of seed endophytic bacteria in maize remain largely unexplored. In this study, the endophytic bacteria in the seeds of maize inbred line WU109 collected during three consecutive seasons were identified using 16S rDNA sequencing. Core community composition was essentially consistent across three seed generations and two planting locations. In total, 212 operational taxonomic units (OTUs) belonging to 11 phyla were identified, among which proteobacteria was the dominant phylum. Fifty-six OTUs were conserved across three seed generations. Within them, 16 OTUs were core components and the dominant OTUs were Ralstonia solanacearum, Delftia tsuruhatensis, Bacillu svelezensis and Shigella boydii, accounting for 60% of the total abundance of OTUs. COG and KEGG analyses showed that the function of seed endophytic bacteria was mainly enriched in metabolic processes, especially in amino acid, carbohydrate and energy metabolism. Taken together, the results suggested that the community of maize seed endophytic bacteria was likely co-shaped by both genetic determination and the environment, while the core constitutes of seed endophytes were largely conserved due to transgenerational transmission. Establishing the mutualistic link between the maize seed and its endophytic bacteria enables the exploitation of the potential of endophytes for enhancing crop production. This finding provides a reference to better understand the inheritance and composition of seed core endophytic bacteria in maize.
1. Introduction
Endophytic bacteria ubiquitously exist in all terrestrial plants without visible detriment to the host [1]. Various plant tissues host large numbers of endophytic bacteria, which can be recruited either horizontally from external environments or acquired vertically through transgenerational transmission via seeds [2]. Bacterial endophytes preserve a symbiotic relationship with host plants, and they can facilitate the acquisition of nutrients, participate in the regulation of phytohormones to promote plant growth, and release antimicrobial compounds, competing for space and nutrients with phytopathogens to help host plants further mitigate the consequences of harsh conditions [3,4,5]. Endophytic strains affiliated with Bacillus, Enterobacter, Burkholderia, Pseudomonas, Rahnella, Azotobacter, Serratia, Arthrobacter, Streptomyces and Isoptericola had been successful when utilized in alleviating abiotic and biotic stress and promoting seedling growth in multiple crop species [6,7,8,9,10,11]. With the rapid development of high-throughput sequencing technology, investigation of the community composition of the microbiome inside plant tissues becomes feasible, providing a better opportunity to understand the mutualistic interactions between host plants and endophytic bacteria [3]. Nevertheless, current researches on plant endophytes mainly focus on the rhizosphere and phyllosphere, whereas the seed endophytes are inadvertently neglected.
The seed phase is an important phase in the crop lifespan, and seeds can be stored for several years when adverse conditions are met, becoming new seedlings in an appropriate environment [12]. Meanwhile, the seed is also an ideal habitat for endophytic bacteria. It has been reported that the dominate phyla of seed bacterial endophytes are Proteobacteria, Actinobacteria and Firmicutes, composed of 80, 25 and 20 genera, respectively, among which the genera Bacillus and Pseudomonas are ubiquitous [13,14]. The potential applications of seed endophytic bacteria are diverse, involving biofertilization, bioenergy production and bioremediation. Paenibacillus strains isolated from wheat seeds could promote seedling establishment and enhance resistance to Fusarium graminearum [15], while Bacillus velezensis isolated from maize seeds had the ability to promote seedling growth and prevent the infestation of F. verticillioides [16]. The inoculation of seeds with Methylobacterium sp. Cp3, a core strain of seed endophyte in Crotalaria pumila, could promote seed germination, root extension and enhance heavy metal tolerance [17]. Moreover, inoculation with Pantoea and Pseudomonas strains isolated from rice seeds could enhance plant height and root length [18], whereas elimination of the endophytic bacteria in rice seeds resulted in a remarkable suppression of seedling growth and root hair development [19]. However, it remains unclear where these functional endophytes come from and whether and how the community composition of seed endophytes is changed across seed generations and different ecosystems.
Core bacterial seed endophytes can be transmitted through seed generations, which is known as vertical transmission [20,21,22]. The vertical transmission of core seed endophytic bacteria benefits plant growth at an early stage and confers resistance to phytopathogens. Through an evaluation of two consecutive generations of rice seeds, Stenotrophomonas maltophilia and Mycobacterium abscessus were found to be conserved in both generations [23]. Sphingomonas melonis transmitted across generations in resistant rice seeds conferred resistance to bacterial diseases by generating anthranilic acid [24]. Moreover, Methylobacterium sp. Cp3 was found to be the dominant seed endophyte transmitted across seed generations of Crotalaria pumila, displaying the potential to promote seed germination and seedling growth [17]. In addition to vertically transmitted seed endophytes, alien seed endophytes transferred by horizontal transmission are also beneficial to host plants through symbiosis [25]. These alien seed endophytes are largely acquired from the environment. A number of studies have comprehensively investigated the community and function of maize seed endophytes, showing their capability to stimulate seedling growth and protect the host from various stresses [16,26,27]. However, details about the composition and conservation of maize seed core endophytic bacteria across consecutive generations remain unclear.
To illustrate the inheritance and potential functions of maize seed bacterial endophytes through diversity, community composition and functional analysis, 16S rDNA sequencing was deployed to identify the community structure of seed endophytic bacteria in three successive seed generations of maize inbred line WU109 in this study. It was found that the community of seed endophytic bacteria was determined by both plant genotype and the environment, and the core community was conserved across transgenerational transmission, which may contribute to the improvement of maize production via the regulation of the metabolism process. Our study elaborated upon the core seed-borne bacteria in maize seeds and provided genetic insights into the function of seed bacterial endophytes, which is helpful for further understanding the complexity of interactions among host plants, endophytes and the environment.
2. Materials and Methods
2.1. Plant Materials
Maize seeds (inbred line WU109) of three consecutive generations were collected from two planting locations. Seeds from the generations T0 and T1 were collected from Xinxiang, Henan province, China, located at 35°22′18.48″ N, 113°54′21.53″ E, in 2018 and 2019, respectively. T2 seeds were collected from Sanya, Hainan Province, China, located at 18°24′0.18″ N, 109°45′9.00″ E, in 2020. Collected seeds were stored at −80 °C freezer (Zhongkeduling, Hefei, China) for subsequent experiments.
2.2. Seed Surface Sterilization
The seeds were sterilized with 1% sodium hypochlorite (NaClO) in a 50 mL sterilized tube for 10 min and stirred with a glass stick constantly, under an aseptic environment and room temperature condition. Following the discarding of NaClO, the seeds were washed thoroughly using sterilized ddH2O, then immersed with 75% ethanol for 2 min and washed using sterilized ddH2O three times. To confirm the effect of sterilization, a small amount of the final washing water was dropped on the PDA plate, and the immersed water of unsterilized seeds was used as a positive control.
2.3. Identification of Seed Endophytic Bacteria by 16S rDNA Sequencing
The sterilized seeds were frozen with liquid nitrogen and thoroughly ground by a plant tissue crusher (ShanghaiJingxin, Shanghai, China). Extraction of total DNA was performed using the MolPure® Soil DNA Kit (18815ES70, YEASEN, Changsha, China) according to the manufacturer’s instructions. Two biological repeats with 10 seeds per replicate were included. The quantity and quality of the extracted DNA were assessed by agarose gel electrophoresis and NanoDrop one (Thermo Scientific China, Shanghai, China). Bacteria-specific primers (799F/1193R) were employed to amplify the V5–V7 region for amplicon sequencing using the Illumina NextSeq 2000 sequencing platform (Illumina China, Shanghai, China). The target sequences were amplified using PCR, and the resulting products were purified, quantified and standardized to generate a sequencing library. Library quality control was conducted to ensure the construction of high-quality libraries.
2.4. Data Analysis of 16S rDNA Amplicons
The acquired sequencing data were processed using Qiime 1.9.1. The quality of paired-end reads was checked and demultiplexed using Flash 1.2.11. Uparse 11, RDP classifier 2.13 and Usearch 11 were used to generate OTU information. To obtain reliable OTU data, mitochondrial and chloroplast reads, low-abundance OTUs (less than 0.01%) and that unclassified at the phylum level were removed. The α diversity analysis was conducted using R package vegan 2.6. Principle coordinate analysis (PCoA) for β diversity was conducted by R package scatterplot3d 0.3–44. Population community analysis at the phylum and genus levels was conducted by R package plyr 1.8.9, tidyverse 2.0.0 and ggalluvial 0.12.5. PICRUSt 1.1.0 software based on the COG (Cluster of Orthologous Groups of proteins) and KEGG (Kyoto Encyclopedia of Genes and Genome) databases was used to predict functional gene composition in the samples by comparing species composition information obtained from 16S sequencing data, and then functional differences between samples or groups were analyzed.
3. Results
3.1. Analysis of Pyrosequencing Data
Surface sterilization is paramount for analyzing endophytic bacteria. After culturing for 3 d, compared to the unsterilized control, no colony was found to be growing on the PDA plate, indicating that the sterilized seeds were suitable for constructing the 16S rDNA library. When the sequencing throughput reached 10 kb, the Shannon index at the OTU level was no longer increased, indicating that the data adequately captured the species diversity in the samples. In total, 257 OTUs and 86,584 reads were acquired. After filtering out the reads from the mitochondria/chloroplast and low-abundance OTUs (less than 0.01%), an average of 10,848 reads affiliated with 212 OTUs were obtained from all the samples.
3.2. Community Diversity of Seed Endophytic Bacteria
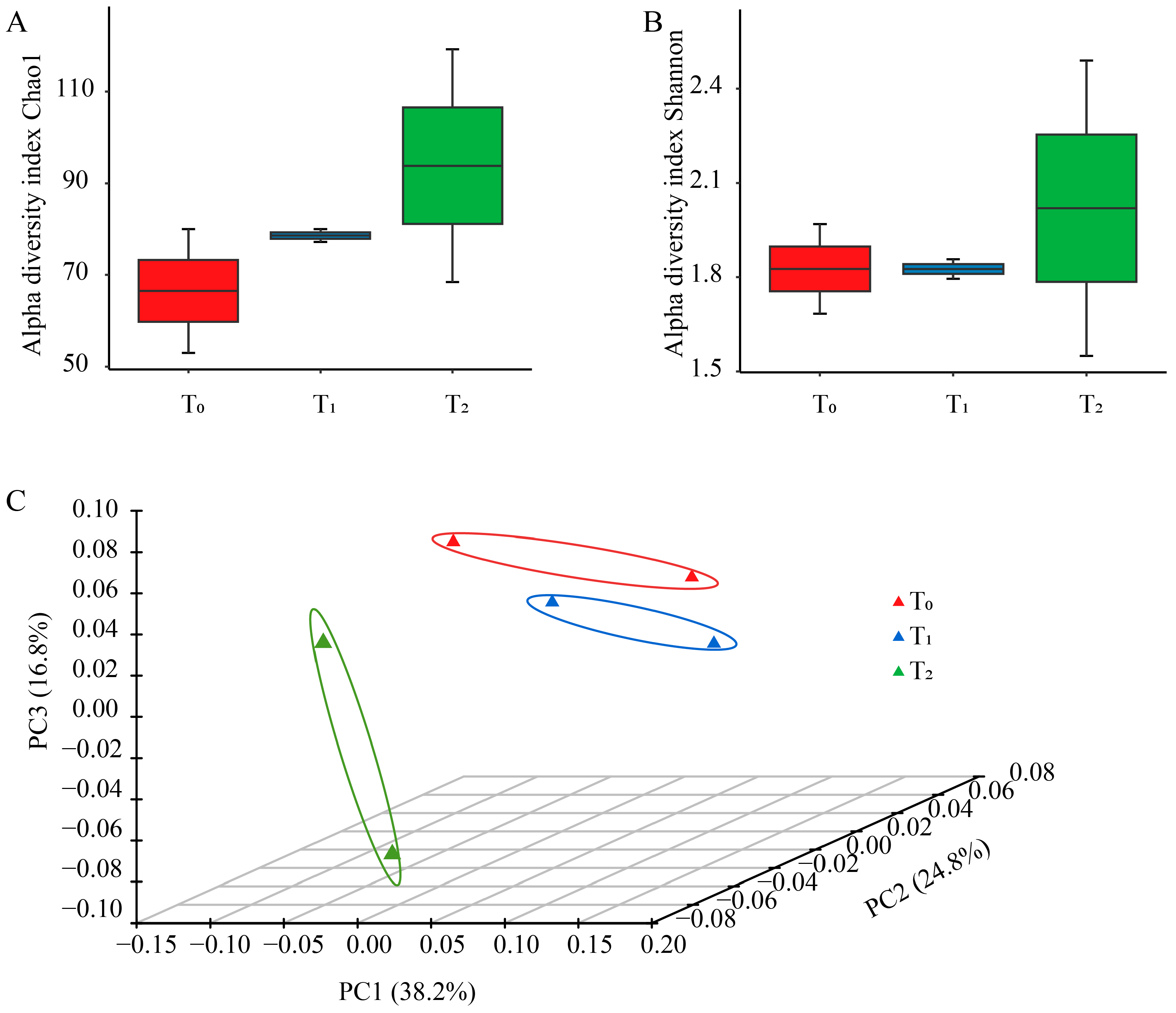
The Chao 1 and Shannon index affiliated with alpha diversity were utilized to reflect the richness and diversity of seed bacterial endophytes among the three seed generations. The means of Chao 1 richness calculated for consecutive generations were 66.5, 78.6 and 93.8 in T0, T1 and T2, respectively. Similarly, the mean of the Shannon index of T2 and was the highest among the three seed generations, larger than that of T0, and T1 (Figure 1A,B). For principal coordinate analysis, 79.8% of the variance was explained by the first three coordinates. The distance between T0 and T1 was short, while T2 was far from them (Figure 1C), suggesting that the community structure of T0 and T1 was more similar than that of T1 and T2, likely due to the difference between planting areas.
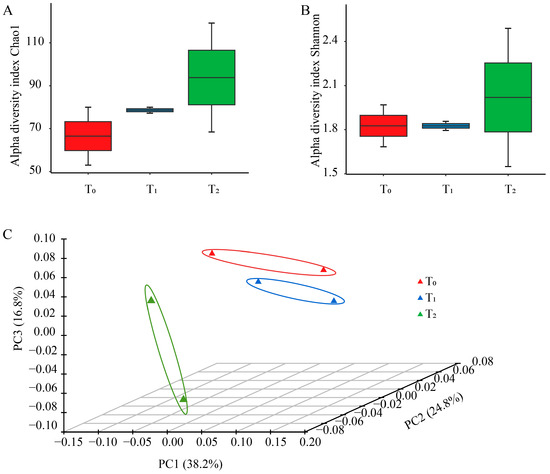
Figure 1.
Community diversity for maize seed bacterial endophytes of three seed generations. (A) Chao1 richness index of seed endophytic bacteria at OTU level. (B) Shannon index of seed endophytic bacteria at OTU level. (C) Principal coordinate analysis of total seed endophytic communities across three seed generations at OTU level. T0, T1 and T2 represent three consecutive seed generations, respectively.
3.3. Core Endophytic Bacteria Were Common Among Three Seed Generations
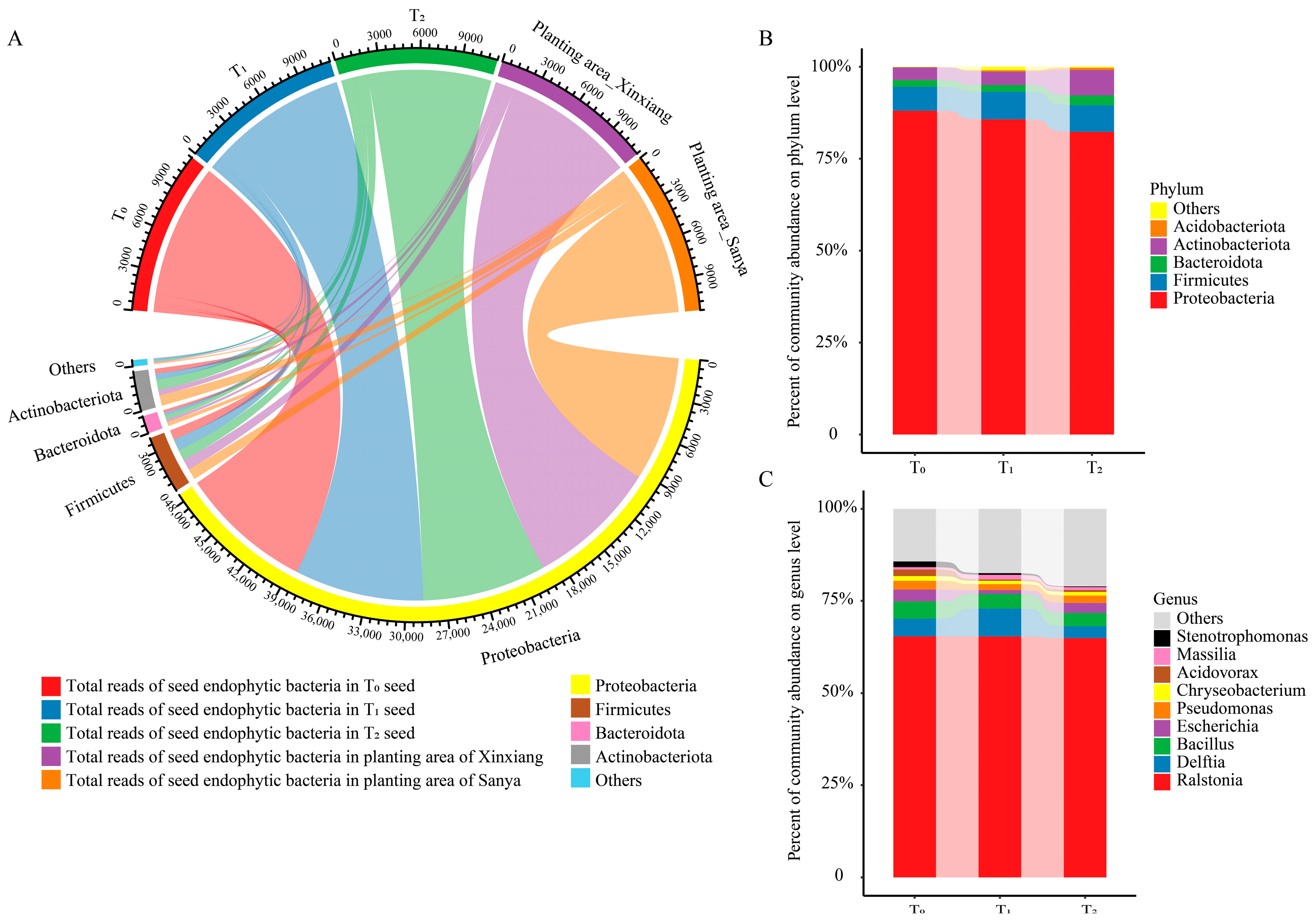
The dominant phylum was Proteobacteria, accounting for over 80% of total species, followed by Firmicutes, Actinobacteriota and Bacteroidota (Figure 2A,B). Moreover, there was no conspicuous difference among the three seed generations and the two planting areas at the phylum level (Figure 2A,B). Ralstonia, Delftia, Bacillus, Escherichia, Pseudomonas, Chryseobacterium, Acidovorax, Massilia, Stenotrophomonas and Sphingomonas were among the top 10 core genera, making up 79.66–85.58% of the total endophytic community (Figure 2C). Bacillus was affiliated with Firmicutes and Chryseobacterium was affiliated with Bacteroidota, while the other eight core genera belonged to Proteobacteria. Among them, Ralstonia was the dominant genus of maize seed endophytic bacteria, accounting for 65% of total OTUs.
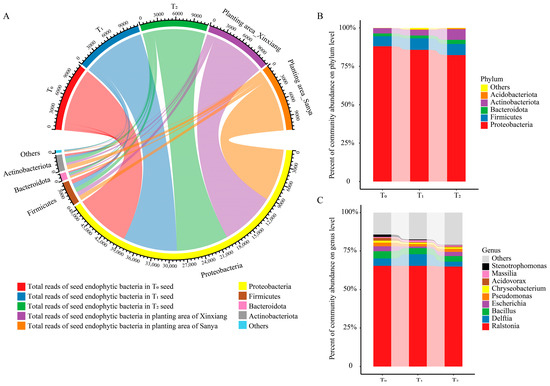
Figure 2.
Community composition and taxa classification among three successive seed generations. (A) Chordal diagram of the bacterial community structure at the phylum level among three seed generations and two planting areas. Taxa with a proportion lower than 1% were summarized as Others. (B) Community composition at the phylum level in three seed generations. (C) Community composition at the genus level in three seed generations. T0, T1 and T2 represent three consecutive seed generations, respectively.
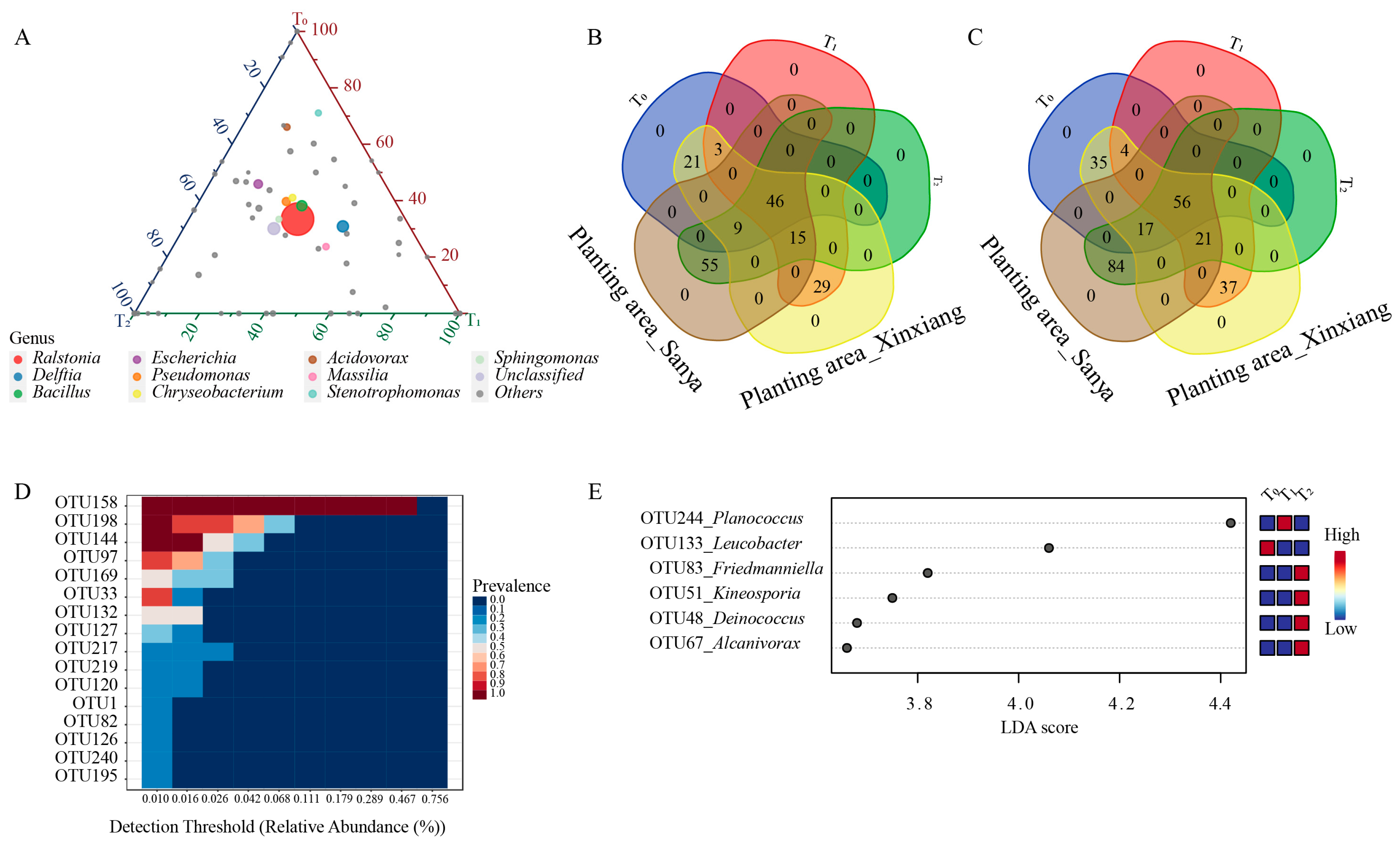
Within the top 10 genera, Ralstonia, Bacillus, Pseudomonas, Chryseobacterium and Sphingomonas were nearly equally distributed among the three seed generations (Figure 2C, Figure 3A). Acidovorax and Stenotrophomonas were more abundant in the T0 generation, while Delftia and Massilia were more enriched in the T1 generation (Figure 3A). The abundance of Escherichia was similar between the T0 and T1 generations, but it was relatively lower in the T2 generation (Figure 3A). Through comparing the similarities and differences of the three successive seed generations and the two planting areas, 46 genera and 56 OTUs were found to exist in all three generations and in both planting areas, suggesting that these OTUs were conserved and could be transferred through vertical transmission (Figure 3B,C). Within the 56 OTUs, 16 of them were core OTUs with high abundance and prevalence, but only 10 OTUs were annotated at the species level (Figure 3D, Table 1). Moreover, there were 156 OTUs that were flexible among different seed generations and planting locations. The dominant OTUs were annotated to Ralstonia solanacearum, Delftia tsuruhatensis and Bacillus velezensis (Table 1). Moreover, there were 29 genera and 37 OTUs that were unique to Xinxiang, and 55 genera and 84 OTUs were unique to Sanya (Figure 3B,C). Through LEfSe analysis, six OTUs (OTU48, OTU51, OTU67, OTU83, OTU133, OTU244) showed prominent differences among three generations (Figure 3E). Among them, OTU244, belonging to Planococcus, was more enriched in the T1 generation, and OTU133, affiliated with Leucobacter, was more abundant in the T0 generation, while OTU48, OTU51, OTU67 and OTU83 were more enriched in T2 generation. The above results indicate that the conserved core OTUs could be transmitted vertically through seed generations, while some unique OTUs that exist within different seed generations could be transmitted horizontally for adaptation to external environments.
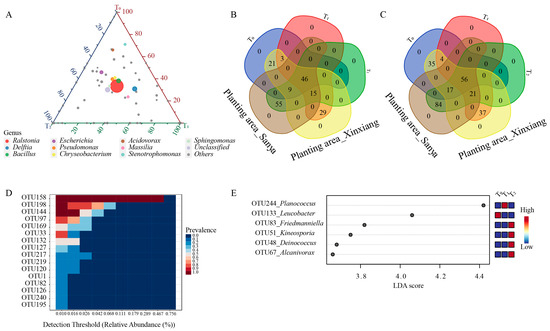
Figure 3.
Distribution of seed endophytic bacteria among three seed generations. (A) Ternary plot of top 10 genera in three seed generations. (B) Venn diagram for shared and unique genera. (C) Venn diagram for shared and unique OTUs. (D) Relative abundance of core OTUs. (E) LEfSe analysis at OTU level among successive seed generations. Linear discriminant analysis (LDA) scores were obtained through LEfSe analysis. T0, T1 and T2 represent three consecutive seed generations, respectively.

Table 1.
Annotation information of core OTUs at species level.
3.4. Three Seed Generations Shared Similar Functional Traits
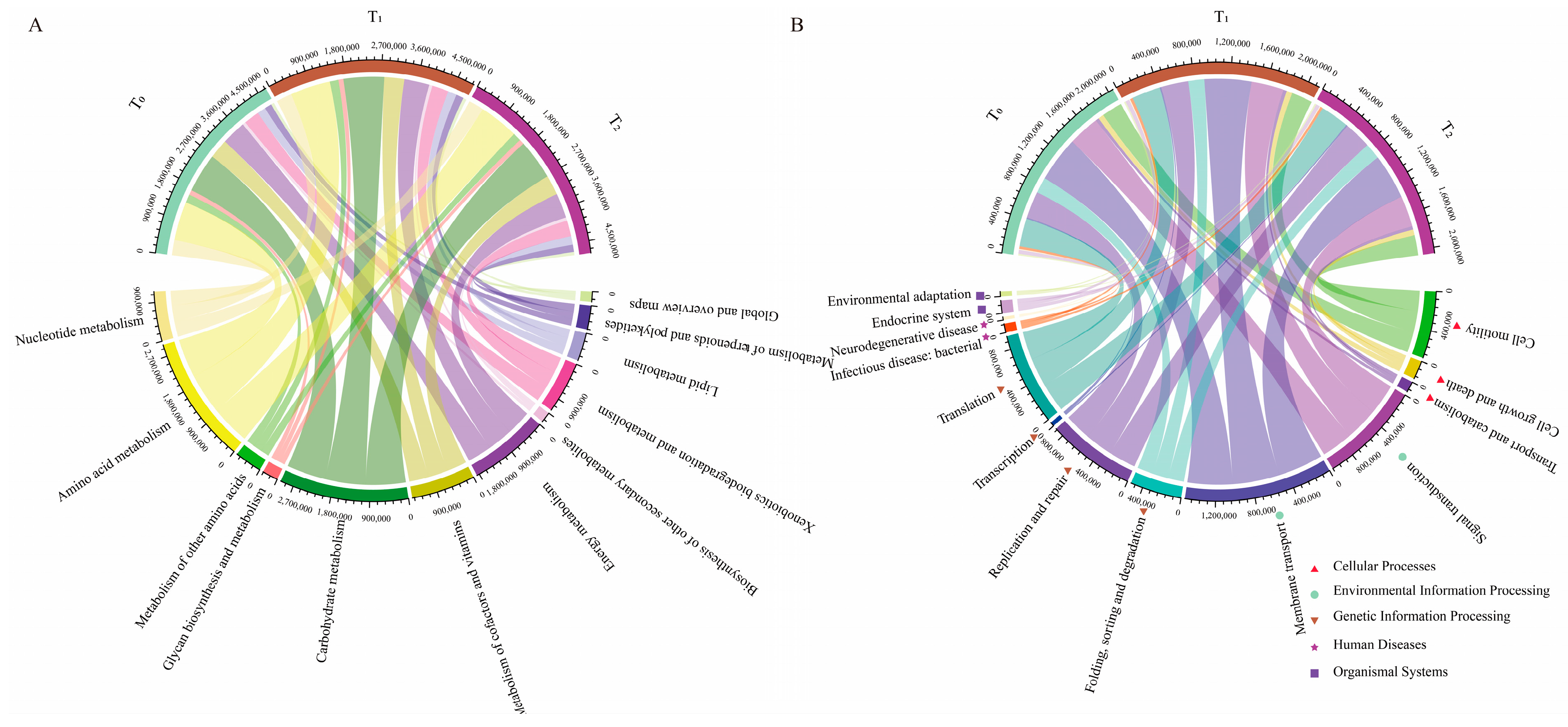
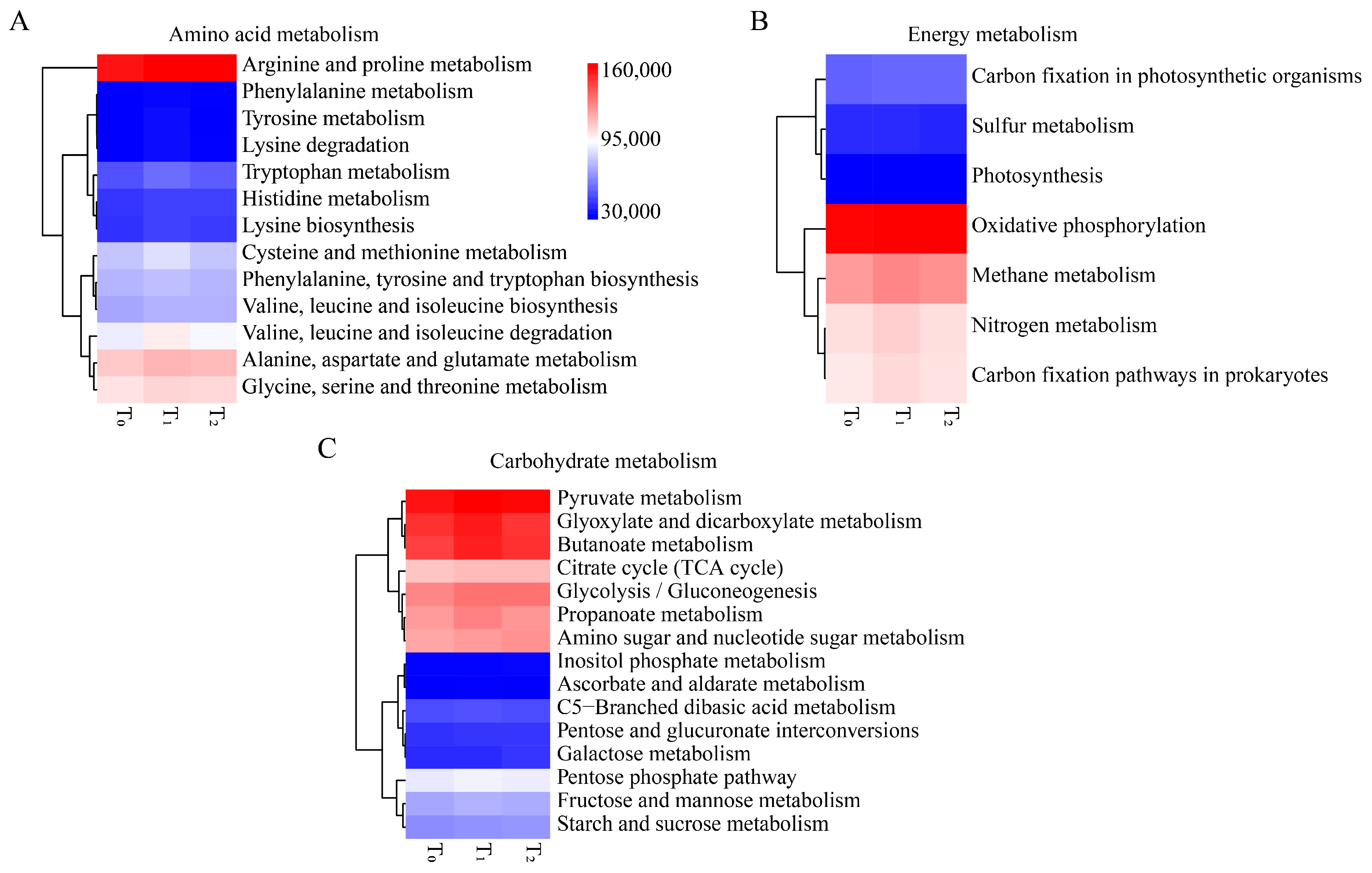
To better understand the functions of seed endophytic bacteria, PICRUSt analysis based on the COG and KEGG databases was conducted. The results showed that the predicted functions for maize seed bacterial endophytes of three seed generations were highly conserved. Based on the COG database, the top enriched pathways were those involving amino acid transport and metabolism, energy production and conservation, signal transduction mechanisms, inorganic ion transport and metabolism, carbohydrate transport and metabolism, etc. (Figure 4A). In KEGG enrichment analysis, metabolic functions were the most important, followed by genetic information processing and environmental information processing (Figure 4B). Within the metabolic functions of KEGG, amino acid metabolism, carbohydrate metabolism and energy metabolism were dominant (Figure 5A). Moreover, replication and repair, membrane transport and cell motility were dominant in the pathways of genetic information processing, environmental information processing and cellular processes, respectively (Figure 5B). Furthermore, arginine and proline metabolism were dominant in amino acid metabolism, while oxidative phosphorylation and pyruvate metabolism were prominent in energy metabolism and carbohydrate metabolism, respectively (Figure 6).

Figure 4.
Metataxonomic function prediction of seed endophytic bacteria of three consecutive seed generations. (A) Function predictions were categorized based on COG classification. The abscissa represents the relative abundance of different functional categories based on the COG database, while the ordinate represents three consecutive seed generations. (B) Function predictions were categorized based on KEGG classification at level 1. The variation in the ordinate represents the different function items on the KEGG pathway level 1. T0, T1 and T2 represent three consecutive seed generations, respectively. The values in the abscissa represent the relative abundance of corresponding functions in the samples.
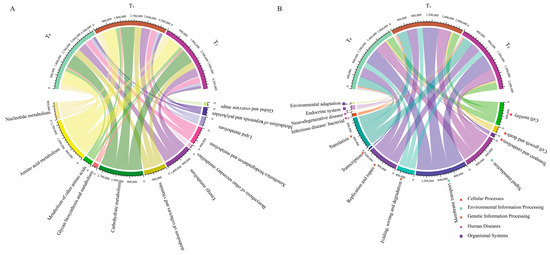
Figure 5.
The major functions predicted for seed bacterial endophytes were enriched in metabolic functions. (A) Function predictions about metabolism were categorized based on KEGG classification at level 2. (B) Function predictions about genetic information processing, environmental information processing and cellular processes were categorized based on KEGG classification at level 2. T0, T1 and T2 represent three consecutive seed generations, respectively. The values represent the relative abundance of corresponding functions in the samples.
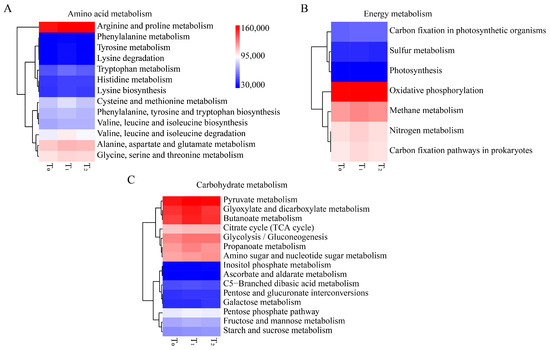
Figure 6.
Functional prediction of maize seed bacterial endophytes that might involve amino acid metabolism, energy metabolism and carbohydrate metabolism. (A) Heatmap of relative abundance of amino acid metabolism. (B) Heatmap of relative abundance of carbohydrate metabolism. (C) Heatmap of relative abundance of energy metabolism. T0, T1 and T2 represent three consecutive seed generations, respectively. The values represent the relative abundance of corresponding functions in the samples, and the colors blue, white and red represent the values at low, middle and high levels, respectively.
4. Discussion
The maize seed is a suitable habitat for endophytic bacteria. Through coevolution, seeds and seed-borne endophytes have established a close symbiotic relationship and have benefited each other. In the present study, we found that maize seed endophytic bacteria community structure showed no significant changes among three generations in community α diversity. The above results are supported by research from Sanchez-Lopez et al. [28] in Crotalaria pumila, suggesting that the community diversity of seed bacterial endophytes was conserved to a certain extent across progeny transmission via seeds.
In nine monocot and eight dicot crop species, the top five phyla of seed endophytic bacteria were identified to be Proteobacteria, Firmicutes, Cynaobacteria, Bacteroidota and Actinobacteriota [29]. Our results showed that the dominant phylum of seed endophytic bacteria in maize was Proteobacteria, followed by Firmicutes, Actinobacteriota and Bacteroidota, which is accordant with previous reports in highland barley [30], Crotalaria pumila [28], Solanum lycopersicum [31], Sophora davidii [32], Daucus carota [33], Elymus nutans [34] and Cistanche phelypaea [35]. By analyzing endophytic bacterial community structure and diversity in melon seeds, another study identified 492 OTUs affiliated with 8 phyla, and Proteobacteria and Firmicutes were dominant [36]. Therefore, there is a high degree of conservation in the community structure of maize seed endophytic bacteria at the phylum level across seed generations and multiple crop species.
In maize, Liu et al. [14] reported that Enterobacter, Shigella, Pseudomonas and Achromobacter were the core genera of seed endophytes. From the seeds of 30 maize cultivars, 80 bacterial endophytes were isolated, and the dominant genera were Bacillus and Staphylococcus [37]. In the present study, the core genera of seed endophytes in maize were identified, including Ralstonia, Delftia, Bacillus, Escherichia and Pseudomonas. It is likely that the core genera in maize were highly variable, with only Pseudomonas and Bacillus being conserved. Zeng et al. [36] found that the core genera of melon seed endophytes were Escherichia, Streptococcus, Bifidobacterium, Parabacteroides, Enterobacter and Roseburia. Bacillus, Massilia, Paenibacillus, Pantoea, Pseudomonas, Rhizobium and Sphingomonas were highly abundant genera in Daucus carota [33], while the dominant endophytic bacteria of Alpinia zerumbet seeds were affiliated with Pseudomonas, Kosakonia, Curtobacterium and Sphingobacterium [38]. Through collecting seeds of Elymus nutans from four planting areas, one study found 10 core genera that were likely transmitted vertically across seed generations, including Pseudomonas, Halomonas, Pantoea, Ochrobactrum, Massilia, Sphingomonas, etc. [34]. Taken together, the core genera are different among different crop species and can be different even in the same crop species. Within them, Pseudomonas, Bacillus, Massilia and Sphingomonas exist ubiquitously, while Ralstonia and Delftia are rarely reported as core genera of seed endophytic bacteria.
In this study, we found that three consecutive seed generations shared 56 common OTUs. The dominant OTUs were Ralstonia solanacearum, followed by Delftia tsuruhatensis, Bacillus velezensis, Shigella boydii, Acidovorax avenae, Pseudomonas mosselii, Stenotrophomonas maltophiliag and Staphylococcus hominis. Wallace [27] reviewed publications about bacterial endophytes isolated from maize seeds up until 2022 and concluded that seven Bacillus strains (B. altitudinis, B. amyloliquefaciens, B. aquimaris, B. cereus, pumilus, B. subtilis and B. velezensis), Paenibacillus dendritiformis, Staphylococcus arlettae, Burkholderia anthina and Pseudomonas aeruginosa could be isolated from maize seeds. Pal et al. [39] found that endophytic bacterium B. velezensis within maize seeds could generate lipopeptides, which promoted seedling growth and activated ISR (induced systemic resistance) to enhance antifungal activity against the fungal pathogen F. verticillioides. Moreover, Stenotrophomonas maltophilia isolated from rice seeds showed an inhibitory effect on rice blast disease by suppressing the growth of Magnaporthe grisea in vitro when S. maltophilia was applied as biofertilizer in soil [40], while Staphylococcus hominis isolated from jute seeds also displayed antibacterial capability against Staphylococcus aureus [41]. In addition to B. velezensis, Stenotrophomonas maltophilia and Staphylococcus hominis, the functions of other core OTUs identified in this study remain to be characterized in the future.
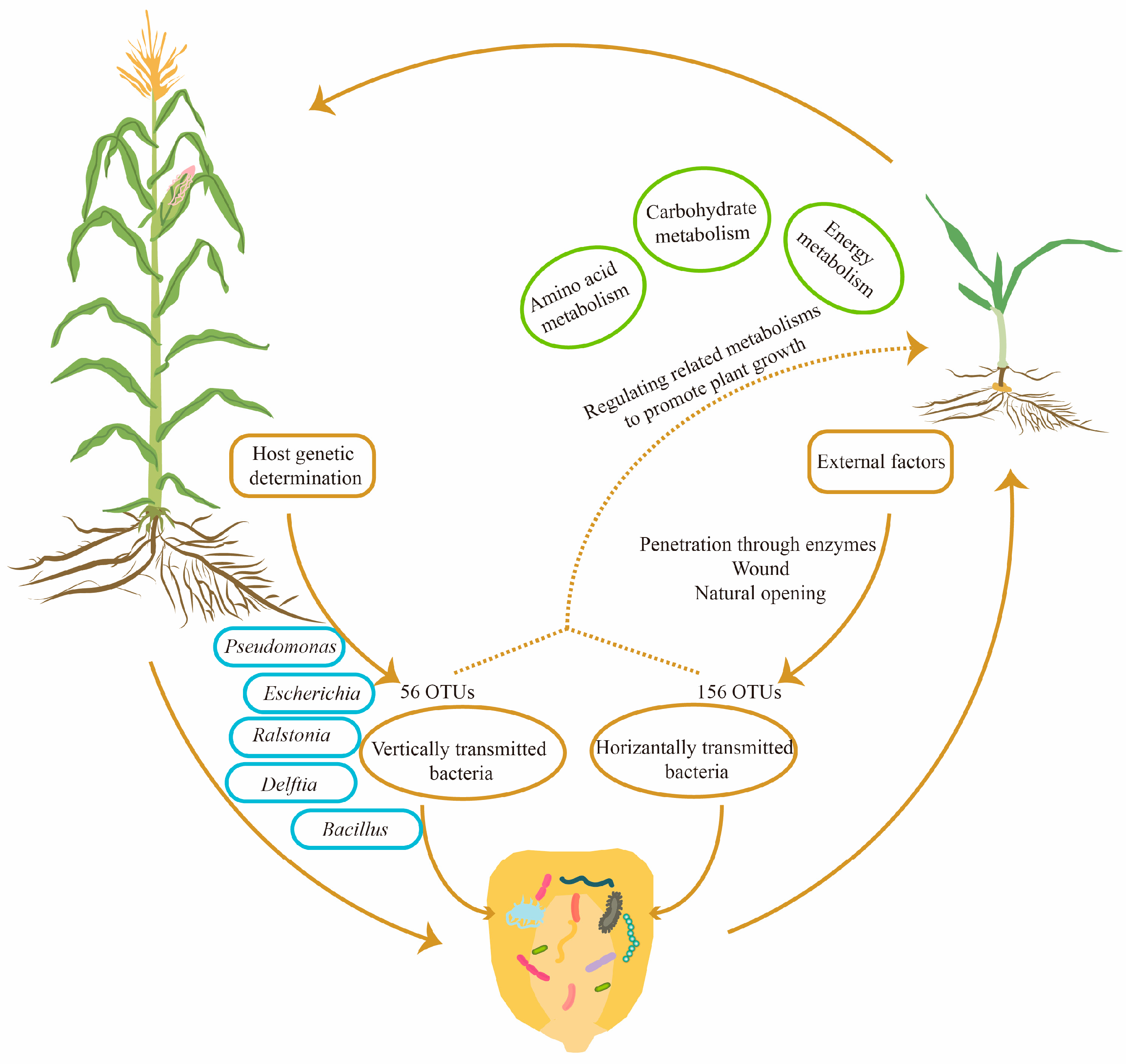
Through investigating the COG and KEGG databases, we found that the functions of seed endophytic bacteria in maize were mainly involved in metabolic processes, particularly in regulating amino acid metabolism, carbohydrate metabolism and energy metabolism. The results were essentially consistent with findings in Crotalaria pumila, which also illustrated that the above three metabolic pathways were major functions of seed bacterial endophytes [28]. In tomato, KEGG functional prediction also proposed primary metabolic processes as the major role of seed bacterial community [42]. In Cucumis melo, the results indicated that the function of seed endophytic bacteria was enriched in pathways of protein synthesis, carbohydrate metabolism, substance transport and metabolite synthesis [36]. Taken together, diverse crop species are capable of exerting their functions using the conserved primary metabolic pathways of seed endophytic bacteria to facilitate seedling growth and development, as well as progeny propagation. The potential mechanisms by which seed endophytic bacteria promote crop growth are summarized in Figure 7.
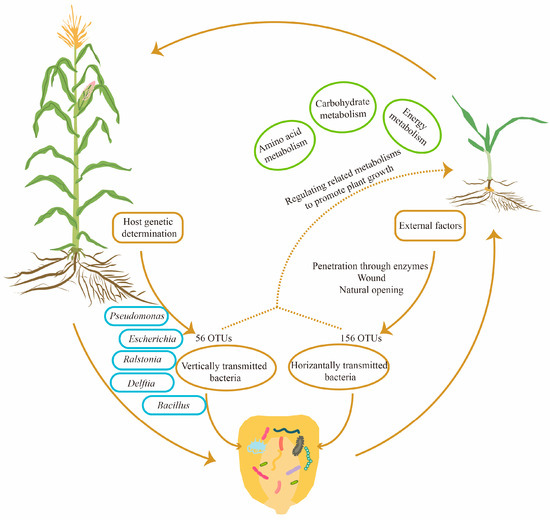
Figure 7.
A diagram of the inheritance and potential functions of maize seed endophytic bacteria. According to the analysis of community structure among three consecutive seed generations, 56 OTUs were found to be conserved and transmitted vertically, whereas another 156 OTUs were changeable and transmitted horizontally from the external environment. Within the conserved 56 OTUs, 10 core OTUs were annotated at the species level. Based on COG and KEGG predictions, seed bacterial endophytes of maize might promote seedling growth by regulating amino acid metabolism, carbohydrate metabolism and energy metabolism.
5. Conclusions
The symbiotic relationship between seeds and their bacterial endophytes is of vital importance for plant growth and development. The current study focused on illustrating the inheritance and function prediction of maize seed bacterial endophytes. Fifty-six OTUs were conserved across three successive seed generations. Within them, Ralstonia, Bacillus, Pseudomonas, Chryseobacterium and Sphingomonas are core genera of maize seed bacterial endophytes. Host plants could maintain these conserved core OTUs across progeny transmission, which may benefit seedling growth and survival in the face of environmental stresses or pathogen attacks through modulating the metabolic process.
Author Contributions
K.Z.: methodology, software, formal analysis and investigation, validation, data curation, writing—original draft preparation and revision; Y.Z.: methodology, formal analysis and investigation, data curation, writing—original draft preparation; C.Z.: investigation, data curation; Q.W.: resource, technical support; X.G., conceptualization, supervision, funding acquisition, project administration, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported financially by the grants from the JBGS [2021]002 project from the Jiangsu Government, the National Nature Science Foundation of China (No. 32472095), and Collaborative Innovation Center for Modern Crop Production (CIC-MCP) to X.G.
Data Availability Statement
Dataset available on request from the authors. The raw data supporting the conclusions of this article will be made available by the authors on request via dataset-file. The 16S rDNA sequences from the obtained isolates were submitted to the NCBI database with the URL of “https://www.ncbi.nlm.nih.gov/bioproject/1188770”.
Acknowledgments
The bioinformatics analysis in this work was supported in part by the high-performance computing platform of the Bioinformatics Center, Nanjing Agricultural University.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E.R.B.; Taghavi, S.; Mezgeay, M.; van der Lelie, D. Endophytic bacteria and their potential applications. Crit. Rev. Plant Sci. 2002, 21, 583–606. [Google Scholar] [CrossRef]
- Guha, T.; Biswas, S.M. Recent progress in the role of seed endophytic bacteria as plant growth-promoting microorganisms and biocontrol agents. World J. Microbiol. Biotechnol. 2024, 40, 218. [Google Scholar] [CrossRef] [PubMed]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Onyino, J.; Gao, X. Current advances in the functional diversity and mechanisms underlying endophyte-plant interactions. Microorganisms 2024, 12, 779. [Google Scholar] [CrossRef]
- Taghavi, S.; van der Lelie, D.; Hoffman, A.; Zhang, Y.-B.; Walla, M.D.; Vangronsveld, J.; Newman, L.; Monchy, S. Genome sequence of the plant growth promoting endophytic bacterium Enterobacter sp. 638. PLoS Genet. 2010, 6, e1000943. [Google Scholar] [CrossRef]
- Taghavi, S.; Garafola, C.; Monchy, S.; Newman, L.; Hoffman, A.; Weyens, N.; Barac, T.; Vangronsveld, J.; van der Lelie, D. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl. Environ. Microbiol. 2009, 75, 748–757. [Google Scholar] [CrossRef]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef]
- Castillo, U.F.; Strobel, G.A.; Ford, E.J.; Hess, W.M.; Porter, H.; Jensen, J.B.; Albert, H.; Robison, R.; Condron, M.A.M.; Teplow, D.B.; et al. Munumbicins, wide-spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscans. Microbiology 2002, 148, 2675–2685. [Google Scholar] [CrossRef]
- Semenzato, G.; Bernacchi, A.; Amata, S.; Bechini, A.; Berti, F.; Calonico, C.; Catania, V.; Esposito, A.; Puglia, A.M.; Piccionello, A.P.; et al. Antibacterial properties of bacterial endophytes isolated from the medicinal plant Origanum heracleoticum L. Front. Biosci. 2024, 29, 111. [Google Scholar] [CrossRef]
- He, Y.; Miao, X.; Xia, Y.; Chen, X.; Liu, J.; Zhou, G. The research of antagonistic endophytic bacterium Bacillus velezensis CSUFT-BV4 for growth promotion and induction of resistance to anthracnose in Camellia oleifera. Microorganisms 2024, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B. Microbial dynamics and interactions in the spermosphere. Annu. Rev. Phytopathol. 2004, 42, 271–309. [Google Scholar] [CrossRef] [PubMed]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, H.; Zhang, X.X.; Zhang, R.Y.; Li, M.; Xu, T.J.; Yang, F.Z.; Zheng, H.J.; Zhao, J.R. Investigating the endophytic bacterial diversity and community structures in seeds of genetically related maize (Zea mays L.) genotypes. 3 Biotech 2020, 10, 27. [Google Scholar] [CrossRef]
- Diaz Herrera, S.; Grossi, C.; Zawoznik, M.; Daniela Groppa, M. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiol. Res. 2016, 186, 37–43. [Google Scholar] [CrossRef]
- Pal, G.; Kumar, K.; Verma, A.; Verma, S.K. Seed inhabiting bacterial endophytes of maize promote seedling establishment and provide protection against fungal disease. Microbiol. Res. 2022, 255, 126926. [Google Scholar] [CrossRef]
- Sanchez-Lopez, A.S.; Pintelon, I.; Stevens, V.; Imperato, V.; Timmermans, J.-P.; Gonzalez-Chavez, C.; Carrillo-Gonzalez, R.; Van Hamme, J.; Vangronsveld, J.; Thijs, S. Seed endophyte microbiome of Crotalaria pumila unpeeled: Identification of plant-beneficial Methylobacteria. Int. J. Mol. Sci. 2018, 19, 291. [Google Scholar] [CrossRef]
- Hernandez, I.; Taule, C.; Perez-Perez, R.; Battistoni, F.; Fabiano, E.; Villanueva-Guerrero, A.; Napoles, M.C.; Herrera, H. Endophytic seed-associated bacteria as plant growth promoters of cuban rice (Oryza sativa L.). Microorganisms 2023, 11, 2317. [Google Scholar] [CrossRef]
- Verma, S.K.; Kingsley, K.; Irizarry, I.; Bergen, M.; Kharwar, R.N.; White, J.F., Jr. Seed-vectored endophytic bacteria modulate development of rice seedlings. J. Appl. Microbiol. 2017, 122, 1680–1691. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Raizada, M.N. Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS ONE 2011, 6, e20396. [Google Scholar] [CrossRef]
- Kaga, H.; Mano, H.; Tanaka, F.; Watanabe, A.; Kaneko, S.; Morisaki, H. Rice seeds as sources of endophytic bacteria. Microbes Environ. 2009, 24, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Ewald, P.W. Transmission modes and evolution of the parasitism-mutualism continuum. Ann. N. Y. Acad. Sci. 1987, 503, 295–306. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Hardoim, C.C.P.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial seed endophyte shapes disease resistance in rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Jones, S.E.; Lennon, J.T.; Martiny, A.C. Microbiomes in light of traits: A phylogenetic perspective. Science 2015, 350, aac9323. [Google Scholar] [CrossRef] [PubMed]
- Gomes Bomfim, C.S.; da Silva, V.B.; Santos Cursino, L.H.; Mattos, W.D.S.; Souza Santos, J.C.; Barbosa de Souza, L.S.; Dantas, B.F.; Santiago de Freitas, A.D.; Fernandes-Junior, P.I. Endophytic bacteria naturally inhabiting commercial maize seeds occupy different niches and are efficient plant growth-promoting agents. Symbiosis 2020, 81, 255–269. [Google Scholar] [CrossRef]
- Wallace, J.G. Maize seed endophytes. Mol. Plant Pathol. 2022, 24, 801–810. [Google Scholar] [CrossRef]
- Sanchez-Lopez, A.S.; Thijs, S.; Beckers, B.; Carmen Gonzalez-Chavez, M.; Weyens, N.; Carrillo-Gonzalez, R.; Vangronsveld, J. Community structure and diversity of endophytic bacteria in seeds of three consecutive generations of Crotalaria pumila growing on metal mine residues. Plant Soil 2018, 422, 51–66. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Gutierrez, J.P.; Lopez-Lavalle, L.A.B. Seed-transmitted bacteria and fungi dominate juvenile plant microbiomes. Front. Microbiol. 2021, 12, 737616. [Google Scholar] [CrossRef]
- Chen, Y.L.; Liang, J.P.; Zia, A.; Gao, X.; Wang, Y.; Zhang, L.Z.; Xiang, Q.J.; Zhao, K.; Yu, X.M.; Chen, Q.; et al. Culture dependent and independent characterization of endophytic bacteria in the seeds of highland barley. Front. Microbiol. 2022, 13, 981158. [Google Scholar] [CrossRef]
- Bergna, A.; Cernava, T.; Raendler, M.; Grosch, R.; Zachow, C.; Berg, G. Tomato seeds preferably transmit plant beneficial endophytes. Phytobiomes J. 2018, 2, 183–193. [Google Scholar] [CrossRef]
- Ai, J.M.; Yu, T.F.; Liu, X.D.; Jiang, Y.Y.; Wang, E.T.; Deng, Z.S. Seed associated microbiota and vertical transmission of bacterial communities from seed to nodule in Sophora davidii. Plant Soil 2023, 491, 285–302. [Google Scholar] [CrossRef]
- Kumar, S.; Johnson, L.J.J.; Teasdale, S.; Morozova, Y.; de Bonth, A.C.M.; Jauregui, R.; Hannaford, R.; Card, S.D.D. Survey of the endophytic bacteria inhabiting wild daucus seed using 16S rRNA gene amplicon sequencing. Microbiol. Resour. Announc. 2023, 12, 23. [Google Scholar] [CrossRef]
- Guo, J.X.; Bowatte, S.; Hou, F.J. Diversity of endophytic bacteria and fungi in seeds of Elymus nutans growing in four locations of Qinghai Tibet Plateau, China. Plant Soil 2021, 459, 49–63. [Google Scholar] [CrossRef]
- Petrosyan, K.; Thijs, S.; Piwowarczyk, R.; Ruraz, K.; Kaca, W.; Vangronsveld, J. Diversity and potential plant growth promoting capacity of seed endophytic bacteria of the holoparasite Cistanche phelypaea (Orobanchaceae). Sci. Rep. 2023, 13, 11835. [Google Scholar] [CrossRef]
- Zeng, K.; Li, Y.; Wang, Z.; Du, Y.; Fan, M.; Xie, L. Community structure and diversity of endophytic bacteria in Melon (Cucumis melo L.) seeds. Horticulturae 2023, 9, 1195. [Google Scholar] [CrossRef]
- Bodhankar, S.; Grover, M.; Hemanth, S.; Reddy, G.; Rasul, S.; Yadav, S.K.; Desai, S.; Mallappa, M.; Mandapaka, M.; Srinivasarao, C. Maize seed endophytic bacteria: Dominance of antagonistic, lytic enzyme-producing Bacillus spp. 3 Biotech 2017, 7, 232. [Google Scholar] [CrossRef]
- Yan, K.; He, L.M.; Zhou, W.H.; Feng, R.Z.; Meng, L.N.; Wei, Q.; Liu, Y. Determination of the community structure and diversity of endophytic bacteria from Alpinia zerumbet seeds. Int. J. Agric. Biol. 2020, 24, 420–428. [Google Scholar]
- Pal, G.; Saxena, S.; Kumar, K.; Verma, A.; Kumar, D.; Shukla, P.; Pandey, A.; Verma, S.K. Seed endophytic bacterium Bacillus velezensis and its lipopeptides acts as elicitors of defense responses against Fusarium verticillioides in maize seedlings. Plant Soil 2023, 492, 109–124. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A. Suppression of the fungal pathogen Magnaporthe grisea by Stenotrophomonas maltophilia, a seed-borne rice (Oryza sativa L.) endophytic bacterium. Arch. Agron. Soil Sci. 2016, 62, 1271–1284. [Google Scholar] [CrossRef]
- Uddin, M.A.; Akter, S.; Ferdous, M.; Haidar, B.; Amin, A.; Molla, A.H.M.S.I.; Khan, H.; Islam, M.R. A plant endophyte Staphylococcus hominis strain MBL_AB63 produces a novel lantibiotic, homicorcin and a position one variant. Sci. Rep. 2021, 11, 11211. [Google Scholar]
- Thomas, P.; Shaik, S.P. Molecular profiling on surface-disinfected tomato seeds reveals high diversity of cultivation-recalcitrant endophytic bacteria with low shares of spore-forming Firmicutes. Microb. Ecol. 2020, 79, 910–924. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).