
Optimization of Enzyme-Assisted Extraction of Bioactive Compounds from Sea Buckthorn (Hippophae rhamnoides L.) Leaves: Evaluation of Mixed-Culture Fermentation




 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals, Reagents, and Enzyme Products
2.3. Central Composite Design (CCD) and Response Surface Methodology (RSM)
2.4. Enzyme-Assisted Extraction (EAE)
2.5. Spontaneous Fermentation Using Tibetan Kefir Grains
2.6. Measurement of pH
2.7. Determination of Total Phenolic Content
2.8. Scanning Electron Microscopy (SEM)
2.9. Microbial Evaluation of Fermented Samples
2.10. Statistical Analysis
3. Results and Discussion
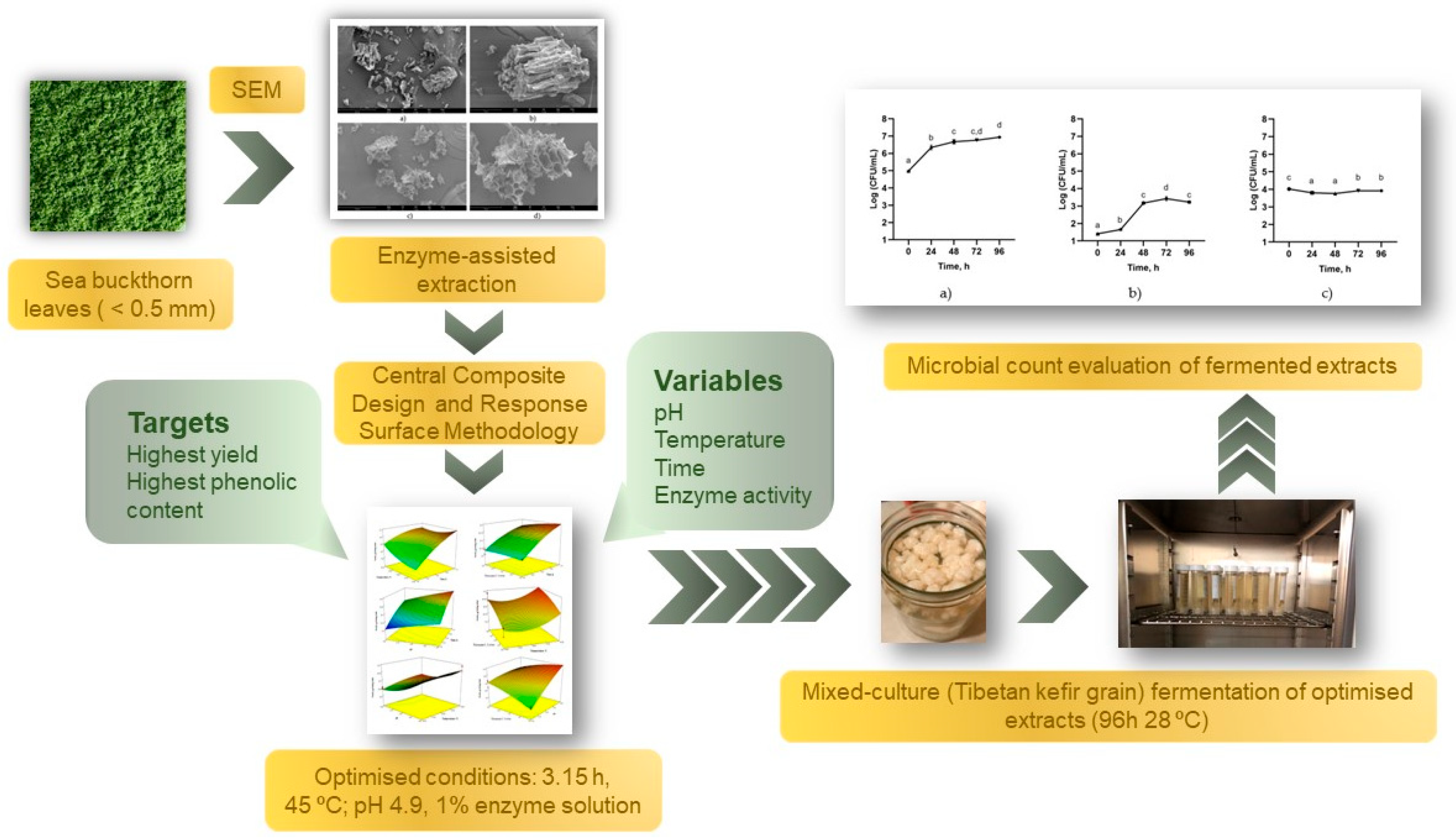
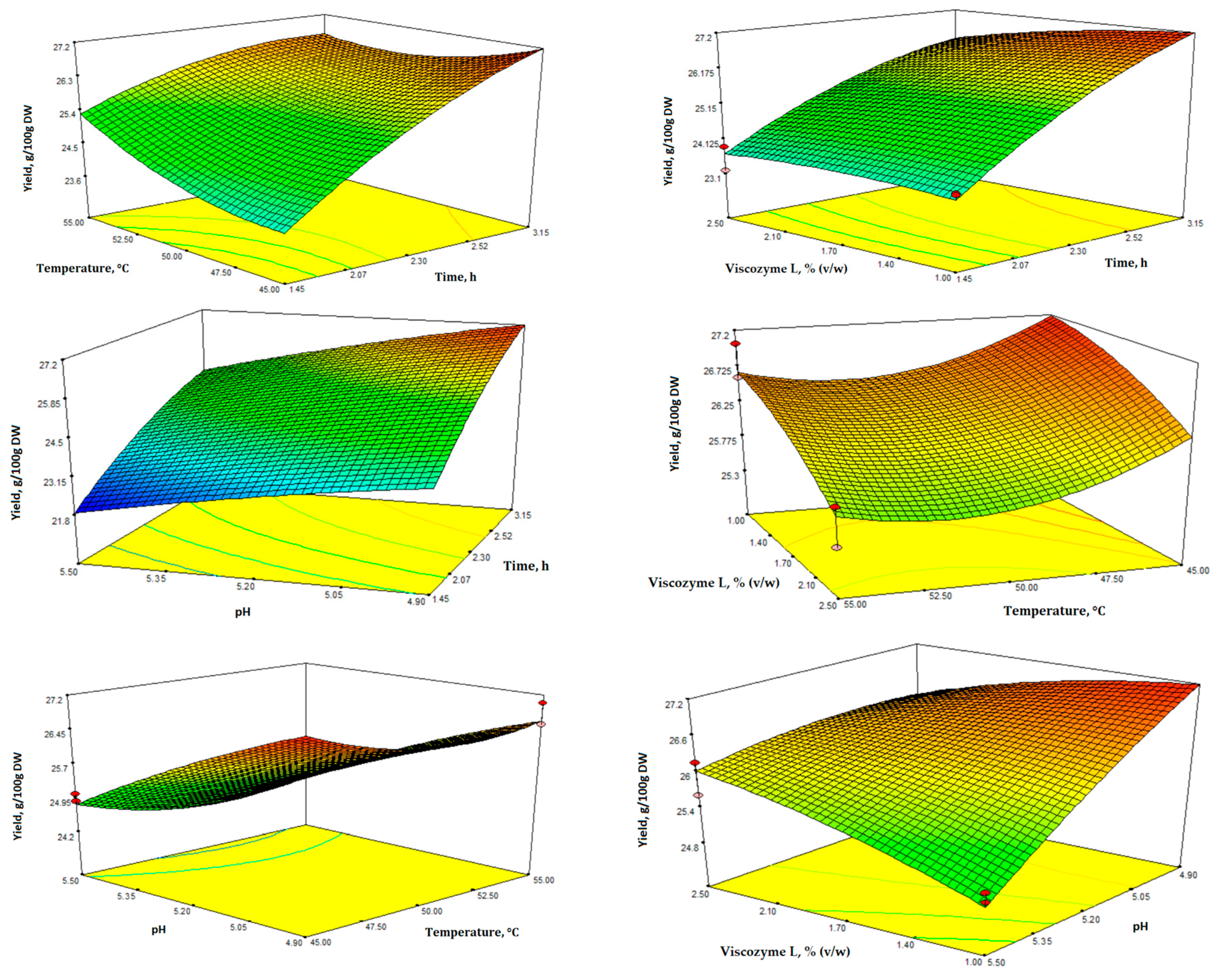
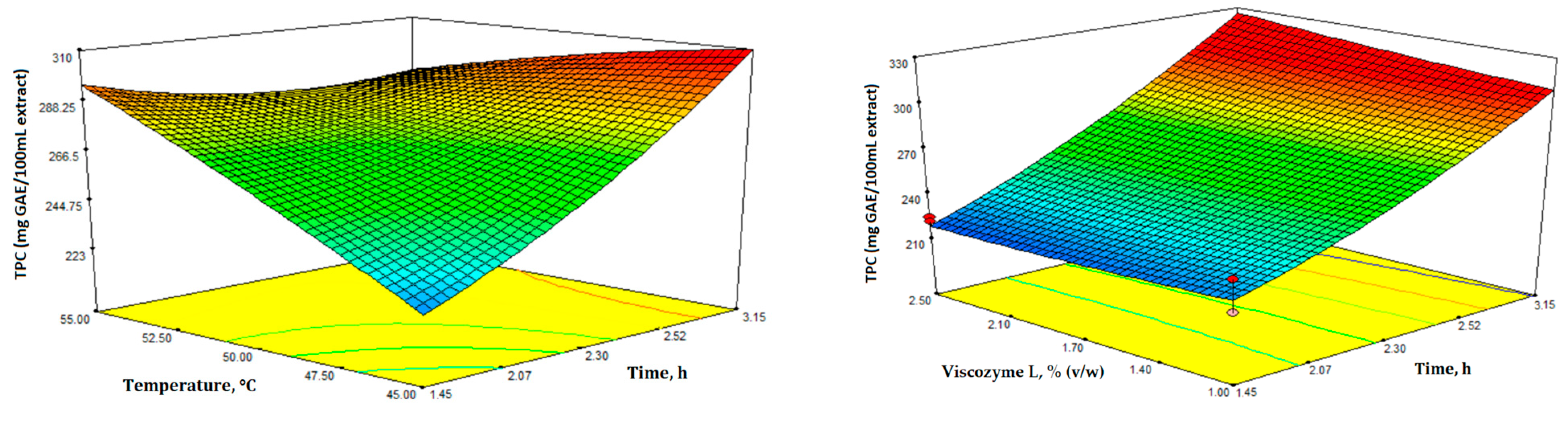
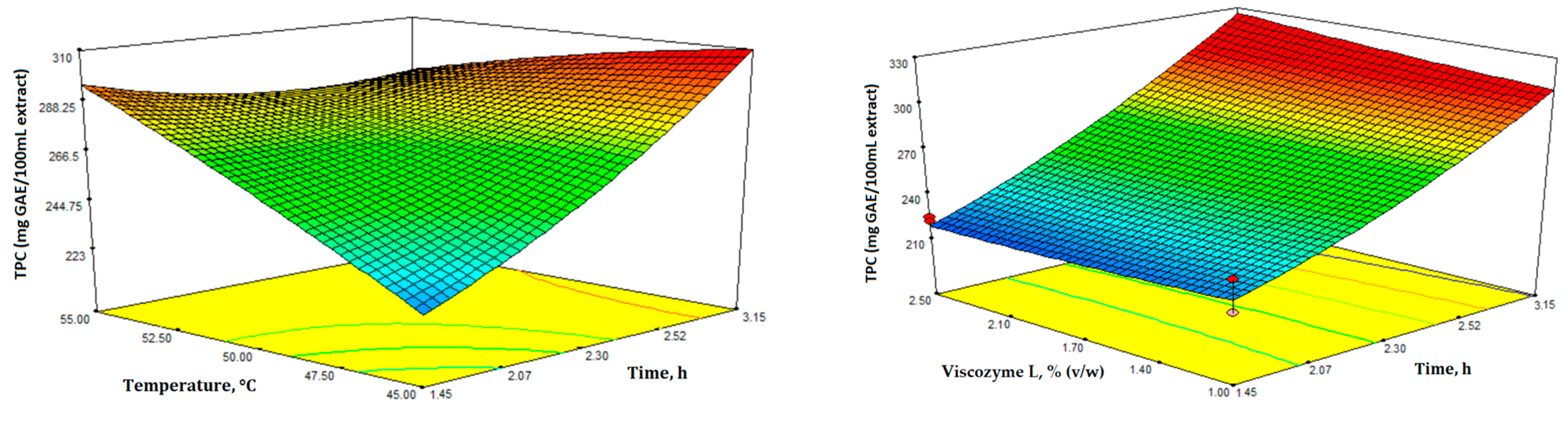
3.1. Optimization of Enzyme-Assisted Extraction (EAE)
3.2. Scanning Electron Microscopy Analysis of Plant Material before and after EAE
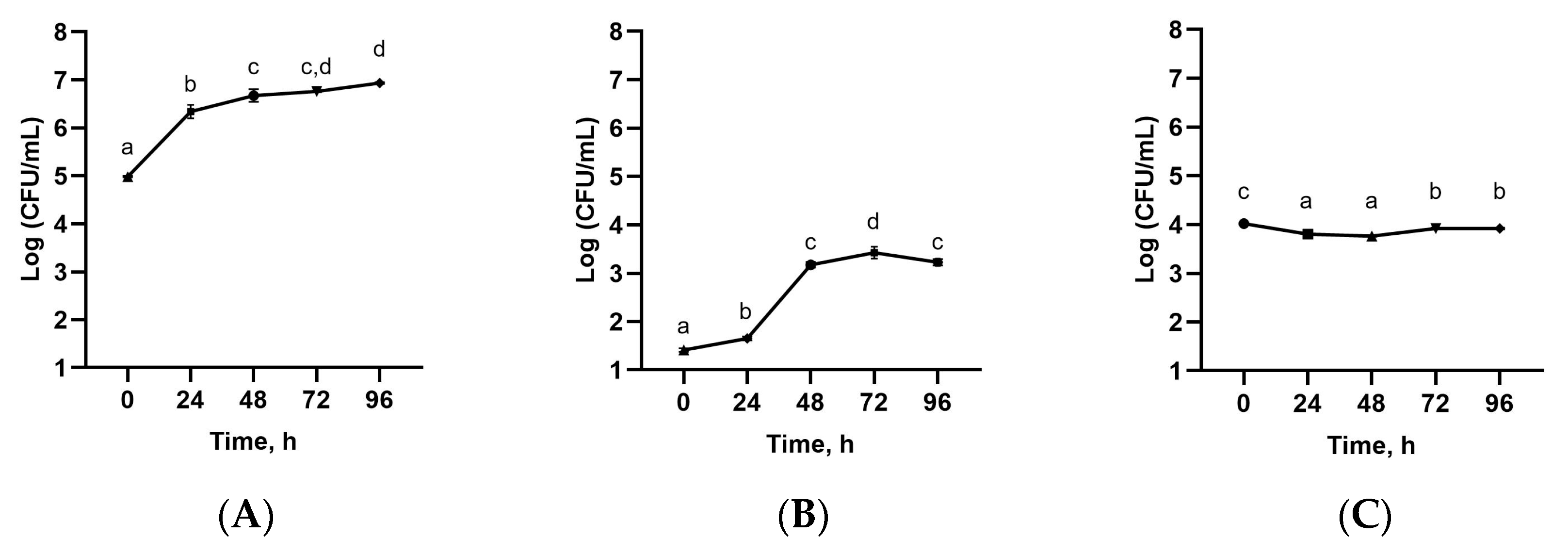
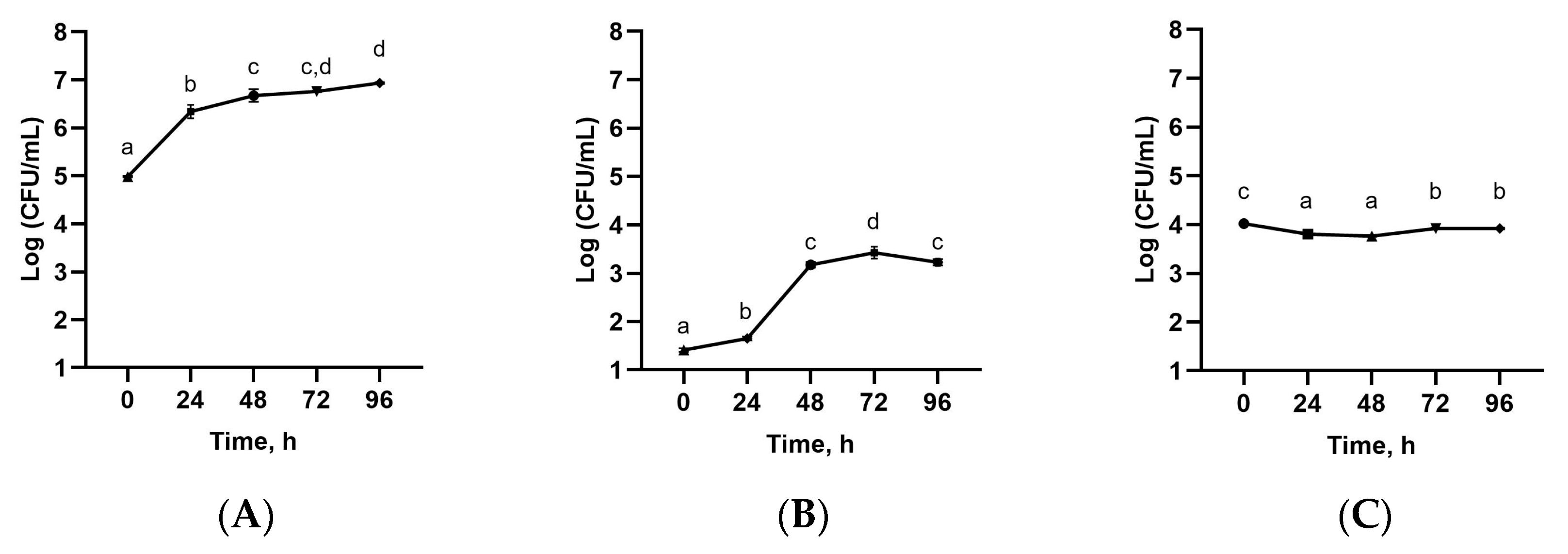
3.3. Microbial Count of Sea Buckthorn Leaves Extract Fermented with Tibetan Kefir Grains
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, T.S. 2. Taxinomie, répartition naturelle et botanique. In Production et Utilisation de l’Argousier (Hippophae rhamnoides L.); Presses Scientifiques du CNRC: Ottawa, ON, Canada, 2004; 145p. [Google Scholar]
- Letchamo, W.; Ozturk, M.; Altay, V.; Musayev, M.; Mamedov, N.A.; Hakeem, K.R. An alternative potential natural genetic resource: Sea buckthorn [Elaeagnus rhamnoides (syn.: Hippophae rhamnoides)]. In Global Perspectives on Underutilized Crops; Springer: Cham, Switzerland, 2018; pp. 25–82. [Google Scholar]
- Bartish, I.V. An ancient medicinal plant at the crossroads of modern horticulture and genetics: Genetic resources and biotechnology of sea buckthorn (Hippophae L., Elaeagnaceae). Gene Pool Divers. Crop Improv. 2016, 1, 415–446. [Google Scholar]
- Wang, Z.; Zhao, F.; Wei, P.; Chai, X.; Hou, G.; Meng, Q. Phytochemistry, health benefits, and food applications of sea buckthorn (Hippophae rhamnoides L.): A comprehensive review. Front. Nutr. 2022, 9, 1036295. [Google Scholar] [CrossRef]
- Żuchowski, J. Phytochemistry and pharmacology of sea buckthorn (Elaeagnus rhamnoides; syn. Hippophae rhamnoides): Progress from 2010 to 2021. Phytochem. Rev. 2023, 22, 3–33. [Google Scholar]
- Gradt, I.; Kühn, S.; Mörsel, J.-T.; Zvaigzne, G. Chemical composition of sea buckthorn leaves, branches and bark. Proc. Latv. Acad. Sci. Sect. B Nat. Exact. Appl. Sci. 2017, 71, 211–216. [Google Scholar]
- Sytařová, I.; Orsavová, J.; Snopek, L.; Mlček, J.; Byczyński, Ł.; Mišurcová, L. Impact of phenolic compounds and vitamins C and E on antioxidant activity of sea buckthorn (Hippophaë rhamnoides L.) berries and leaves of diverse ripening times. Food Chem. 2020, 310, 125784. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xu, Z.; Liao, X. Bioactive compounds, health benefits and functional food products of sea buckthorn: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6761–6782. [Google Scholar] [CrossRef] [PubMed]
- Raudone, L.; Puzerytė, V.; Vilkickyte, G.; Niekyte, A.; Lanauskas, J.; Viskelis, J.; Viskelis, P. Sea buckthorn leaf powders: The impact of cultivar and drying mode on antioxidant, phytochemical, and chromatic profile of valuable resource. Molecules 2021, 26, 4765. [Google Scholar] [CrossRef]
- He, Q.; Yang, K.; Wu, X.; Zhang, C.; He, C.; Xiao, P. Phenolic compounds, antioxidant activity and sensory evaluation of sea buckthorn (Hippophae rhamnoides L.) leaf tea. Food Sci. Nutr. 2023, 11, 1212–1222. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Q.; Wang, Y.; Zu, Y.; Wang, Z.; He, C.; Xiao, P. Application and modern research progress of sea buckthorn leaves. Zhongguo Zhong Yao Za Zhi 2021, 46, 1326–1332. [Google Scholar]
- Saracila, M.; Untea, A.E.; Panaite, T.D.; Varzaru, I.; Oancea, A.; Turcu, R.P.; Vlaicu, P.A. Effects of supplementing sea buckthorn leaves (Hippophae rhamnoides L.) and chromium (III) in broiler diet on the nutritional quality and lipid oxidative stability of meat. Antioxidants 2022, 11, 2220. [Google Scholar] [CrossRef] [PubMed]
- Marić, M.; Grassino, A.N.; Zhu, Z.; Barba, F.J.; Brnčić, M.; Brnčić, S.R. An overview of the traditional and innovative approaches for pectin extraction from plant food wastes and by-products: Ultrasound-, microwaves-, and enzyme-assisted extraction. Trends Food Sci. Technol. 2018, 76, 28–37. [Google Scholar] [CrossRef]
- Hernández Becerra, E.; De Jesús Pérez López, E.; Zartha Sossa, J.W. Recovery of biomolecules from agroindustry by solid-liquid enzyme-assisted extraction: A review. Food Anal. Methods 2021, 14, 1744–1777. [Google Scholar] [CrossRef]
- Costa, J.R.; Tonon, R.V.; Cabral, L.; Gottschalk, L.; Pastrana, L.; Pintado, M.E. Valorization of agricultural lignocellulosic plant byproducts through enzymatic and enzyme-assisted extraction of high-value-added compounds: A Review. ACS Sustain. Chem. Eng. 2020, 8, 13112–13125. [Google Scholar] [CrossRef]
- Patra, A.; Abdullah, S.; Pradhan, R.C. Review on the extraction of bioactive compounds and characterization of fruit industry by-products. Bioresour. Bioprocess. 2022, 9, 1–25. [Google Scholar] [CrossRef]
- Yuliarti, O.; Goh, K.K.; Matia-Merino, L.; Mawson, J.; Brennan, C. Extraction and characterisation of pomace pectin from gold kiwifruit (Actinidia chinensis). Food Chem. 2015, 187, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Han, A.; Park, S.; Cho, C.; Rhee, Y.; Hong, H. Effect of enzyme-assisted extraction on the physicochemical properties and bioactive potential of lotus leaf polysaccharides. Int. J. Biol. Macromol. 2020, 153, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Štreimikytė, P.; Urbonavičienė, D.; Balčiūnaitienė, A.; Viškelis, P.; Viškelis, J. Optimization of the multienzyme-assisted extraction procedure of bioactive compounds extracts from common buckwheat (Fagopyrum esculentum M.) and evaluation of obtained extracts. Plants 2021, 10, 2567. [Google Scholar] [CrossRef] [PubMed]
- Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-assisted extraction of plants for sustainable and functional applications. Int. J. Mol. Sci. 2022, 23, 2359. [Google Scholar] [CrossRef] [PubMed]
- Diez-Ozaeta, I.; Astiazaran, O.J. Fermented foods: An update on evidence-based health benefits and future perspectives. Food Res. Int. 2022, 156, 111133. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Śliżewska, K. Obesity as the 21st Century’s major disease: The role of probiotics and prebiotics in prevention and treatment. Food Biosci. 2021, 42, 101115. [Google Scholar] [CrossRef]
- Pimentel, T.C.; de Assis, B.B.T.; dos Santos Rocha, C.; Marcolino, V.A.; Rosset, M.; Magnani, M. Prebiotics in non-dairy products: Technological and physiological functionality, challenges, and perspectives. Food Biosci. 2022, 46, 101585. [Google Scholar] [CrossRef]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Feiqiao, B.Y.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]
- Kotzekidou, P. Improved traditional fermented foods of the Mediterranean Region–Health benefits as functional foods. In Functional Foods and Biotechnology; CRC Press: Boca Raton, FL, USA, 2020; pp. 135–150. [Google Scholar]
- Gao, W.; Zhang, L. Comparative analysis of the microbial community composition between Tibetan kefir grains and milks. Food Res. Int. 2019, 116, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zheng, Y.; Yang, T.; Mac Regenstein, J.; Zhou, P. Functional properties and sensory characteristics of kombucha analogs prepared with alternative materials. Trends Food Sci. Technol. 2022, 129, 608–616. [Google Scholar] [CrossRef]
- Streimikyte, P.; Kailiuviene, J.; Mazoniene, E.; Puzeryte, V.; Urbonaviciene, D.; Balciunaitiene, A.; Liapman, T.D.; Laureckas, Z.; Viskelis, P.; Viskelis, J. The Biochemical alteration of enzymatically hydrolysed and spontaneously fermented oat flour and its impact on pathogenic bacteria. Foods 2022, 11, 2055. [Google Scholar] [CrossRef]
- Zeng, X.; Wang, Y.; Jia, H.; Wang, Z.; Gao, Z.; Luo, Y.; Sheng, Q.; Yuan, Y.; Yue, T. Metagenomic analysis of microflora structure and functional capacity in probiotic Tibetan kefir grains. Food Res. Int. 2022, 151, 110849. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lu, Y.; Wang, J.; Yang, L.; Pan, C.; Huang, Y. Probiotic properties of Lactobacillus strains isolated from Tibetan kefir grains. PLoS ONE 2013, 8, e69868. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, T.; Ye, T.; Yang, X.; Xue, Y.; Shen, Y.; Zhang, Q.; Zheng, X. Effect of lactic acid bacteria and yeasts on the structure and fermentation properties of Tibetan kefir grains. Int. Dairy J. 2021, 114, 104943. [Google Scholar] [CrossRef]
- Plessas, S.; Nouska, C.; Mantzourani, I.; Kourkoutas, Y.; Alexopoulos, A.; Bezirtzoglou, E. Microbiological exploration of different types of kefir grains. Fermentation 2016, 3, 1. [Google Scholar] [CrossRef]
- Streimikyte, P.; Balciunaitiene, A.; Liapman, T.D.; Streimikyte-Mockeliune, Z.; Puzeryte, V.; Borkertas, S.; Viskelis, P.; Viskelis, J. Enzymatically hydrolysed common buckwheat (Fagopyrum esculentum M.) as a fermentable source of oligosaccharides and sugars. Appl. Sci. 2022, 12, 8210. [Google Scholar] [CrossRef]
- Ricci, A.; Bertani, G.; Maoloni, A.; Bernini, V.; Levante, A.; Neviani, E.; Lazzi, C. Antimicrobial activity of fermented vegetable byproduct extracts for food applications. Foods 2021, 10, 1092. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Bernini, V.; Maoloni, A.; Cirlini, M.; Galaverna, G.; Neviani, E.; Lazzi, C. Vegetable by-product lacto-fermentation as a new source of antimicrobial compounds. Microorganisms 2019, 7, 607. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Casas, D.E.; Aguilar, C.N.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Chávez-González, M.L.; Flores-Gallegos, A.C. Enzymatic hydrolysis and microbial fermentation: The most favorable biotechnological methods for the release of bioactive peptides. Food Chem. Mol. Sci. 2021, 3, 100047. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Bobinaitė, R.; Viškelis, P.; Venskutonis, P.R. Variation of total phenolics, anthocyanins, ellagic acid and radical scavenging capacity in various raspberry (Rubus spp.) cultivars. Food Chem. 2012, 132, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Qadir, R.; Anwar, F.; Gilani, M.A.; Zahoor, S.; ur Rehman, M.M.; Mustaqeem, M. RSM/ANN based optimized recovery of phenolics from mulberry leaves by enzyme-assisted extraction. Czech J. Food Sci. 2019, 37, 99–105. [Google Scholar] [CrossRef]
- Syrpas, M.; Valanciene, E.; Augustiniene, E.; Malys, N. Valorization of Bilberry (Vaccinium myrtillus L.) Pomace by enzyme-assisted extraction: Process optimization and comparison with conventional solid-liquid extraction. Antioxidants 2021, 10, 773. [Google Scholar] [CrossRef]
- Qadir, R.; Anwar, F.; Gilani, M.A.; Yaqoob, M.N.; Ahmad, M. Enzyme-assisted extraction for optimized recovery of phenolic bioactives from Peganum hermala leaves using response surface methodology. Curr. Top. Nutraceutical Res. 2019, 17, 49–354. [Google Scholar]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef]
- Islam, M.R.; Kamal, M.M.; Kabir, M.R.; Hasan, M.M.; Haque, A.R.; Hasan, S.K. Phenolic compounds and antioxidants activity of banana peel extracts: Testing and optimization of enzyme-assisted conditions. Meas. Food 2023, 10, 100085. [Google Scholar] [CrossRef]
- Gligor, O.; Mocan, A.; Moldovan, C.; Locatelli, M.; Crișan, G.; Ferreira, I.C. Enzyme-assisted extractions of polyphenols–A comprehensive review. Trends Food Sci. Technol. 2019, 88, 302–315. [Google Scholar] [CrossRef]
- Fernandes, A.; Mateus, N.; de Freitas, V. Polyphenol-dietary fiber conjugates from fruits and vegetables: Nature and biological fate in a food and nutrition perspective. Foods 2023, 12, 1052. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Ge, S.; Lin, S. Review of distribution, extraction methods, and health benefits of bound phenolics in food plants. J. Agric. Food Chem. 2020, 68, 3330–3343. [Google Scholar] [CrossRef]
- Thite, V.S.; Nerurkar, A.S. Valorization of sugarcane bagasse by chemical pretreatment and enzyme mediated deconstruction. Sci. Rep. 2019, 9, 15904. [Google Scholar] [CrossRef]
- Dong, M.; Wang, S.; Xu, F.; Wang, J.; Yang, N.; Li, Q.; Chen, J.; Li, W. Pretreatment of sweet sorghum straw and its enzymatic digestion: Insight into the structural changes and visualization of hydrolysis process. Biotechnol. Biofuels 2019, 12, 276. [Google Scholar] [CrossRef]
- Rafińska, K.; Wrona, O.; Krakowska-Sieprawska, A.; Walczak-Skierska, J.; Kiełbasa, A.; Rafiński, Z.; Pomastowski, P.; Kolankowski, M.; Buszewski, B. Enzyme-assisted extraction of plant material–New functional aspects of the process on an example of Medicago sativa L. Ind. Crops Prod. 2022, 187, 115424. [Google Scholar] [CrossRef]
- Łopusiewicz, Ł.; Śmietana, N.; Paradowska, D.; Drozłowska, E. Black cumin (Nigella sativa L.) seed press cake as a novel material for the development of new non-dairy beverage fermented with kefir grains. Microorganisms 2022, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Walton, G.; Sousa, S.; Rocha-Santos, T.A.; Duarte, A.C.; Freitas, A.C.; Gomes, A.M. In vitro fermentation and prebiotic potential of selected extracts from seaweeds and mushrooms. LWT 2016, 73, 131–139. [Google Scholar] [CrossRef]
- Gómez-García, R.; Vilas-Boas, A.A.; Oliveira, A.; Amorim, M.; Teixeira, J.A.; Pastrana, L.; Pintado, M.M.; Campos, D.A. Impact of simulated human gastrointestinal digestion on the bioactive fraction of upcycled pineapple by-products. Foods 2022, 11, 126. [Google Scholar] [CrossRef]
- Harper, A.R.; Dobson, R.C.; Morris, V.K.; Moggré, G. Fermentation of plant-based dairy alternatives by lactic acid bacteria. Microb. Biotechnol. 2022, 15, 1404–1421. [Google Scholar] [CrossRef]
- Montemurro, M.; Pontonio, E.; Coda, R.; Rizzello, C.G. Plant-based alternatives to yogurt: State-of-the-art and perspectives of new biotechnological challenges. Foods 2021, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to live yoghurt cultures and improved lactose digestion (ID 1143, 2976) pursuant to Article 13 (1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1763. [Google Scholar]
- Jan, G.; Tarnaud, F.; do Carmo, F.L.R.; Illikoud, N.; Canon, F.; Jardin, J.; Briard-Bion, V.; Guyomarc’h, F.; Gagnaire, V. The stressing life of Lactobacillus delbrueckii subsp. bulgaricus in soy milk. Food Microbiol 2022, 106, 104042. [Google Scholar] [CrossRef] [PubMed]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Dou, Z.; Chen, C.; Fu, X. Bioaccessibility, antioxidant activity and modulation effect on gut microbiota of bioactive compounds from Moringa oleifera Lam. leaves during digestion and fermentation in vitro. Food Funct. 2019, 10, 5070–5079. [Google Scholar] [CrossRef]
- Kaewkod, T.; Bovonsombut, S.; Tragoolpua, Y. Efficacy of kombucha obtained from green, oolong, and black teas on inhibition of pathogenic bacteria, antioxidation, and toxicity on colorectal cancer cell line. Microorganisms 2019, 7, 700. [Google Scholar] [CrossRef]
- Alimentarius, C. Codex Standard for Fermented Milks (CODEX STAN 243-2003). In Milk and Milk Products; FAO/WHO: Rome, Italy, 2015. [Google Scholar]
- Łopusiewicz, Ł.; Drozłowska, E.; Siedlecka, P.; Mężyńska, M.; Bartkowiak, A.; Sienkiewicz, M.; Zielińska-Bliźniewska, H.; Kwiatkowski, P. Development, characterization, and bioactivity of non-dairy kefir-like fermented beverage based on flaxseed oil cake. Foods 2019, 8, 544. [Google Scholar] [CrossRef]
- Spizzirri, U.G.; Loizzo, M.R.; Aiello, F.; Prencipe, S.A.; Restuccia, D. Non-dairy kefir beverages: Formulation, composition, and main features. J. Food Compos. Anal. 2023, 117, 105130. [Google Scholar] [CrossRef]
- Sousa, S.; Pinto, J.; Pereira, C.; Malcata, F.X.; Pacheco, M.B.; Gomes, A.M.; Pintado, M. In vitro evaluation of yacon (Smallanthus sonchifolius) tuber flour prebiotic potential. Food Bioprod. Process 2015, 95, 96–105. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Symbol | Coded Levels | ||||
|---|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | |||
| Time | h | A | 1:00 | 1:45 | 2:30 | 3:15 | 4:00 |
| Temperature | °C | B | 40 | 45 | 50 | 55 | 60 |
| Viscozyme L | % | C | 0.5 | 1.0 | 2.0 | 2.5 | 3.0 |
| pH | - | D | 4.5 | 4.9 | 5.3 | 5.6 | 6.0 |
| Time | Temperature | Viscozyme L | pH | Yield | TPC | |
|---|---|---|---|---|---|---|
| Run | min | °C | % | g/100 g DW | mg GAE/100 mL Extract | |
| 1 | 2:30 | 50 | 3.0 | 5.3 | 25.80 | 258.30 |
| 2 | 1:45 | 55 | 2.5 | 5.6 | 25.75 | 266.67 |
| 3 | 2:30 | 50 | 2.0 | 5.3 | 25.86 | 257.30 |
| 4 | 2:30 | 50 | 2.0 | 5.3 | 25.72 | 261.90 |
| 5 | 3:15 | 45 | 2.5 | 5.6 | 26.17 | 296.00 |
| 6 | 1:45 | 45 | 1.0 | 4.9 | 23.86 | 237.23 |
| 7 | 1:45 | 45 | 2.5 | 4.9 | 23.19 | 224.30 |
| 8 | 2:30 | 50 | 2.0 | 4.5 | 25.66 | 286.05 |
| 9 | 2:30 | 50 | 2.0 | 6.0 | 23.71 | 214.55 |
| 10 | 2:30 | 50 | 2.0 | 5.3 | 25.27 | 258.13 |
| 11 | 1:45 | 55 | 2.5 | 5.6 | 25.29 | 264.43 |
| 12 | 2:30 | 40 | 2.0 | 5.3 | 26.23 | 267.05 |
| 13 | 3:15 | 45 | 2.5 | 5.6 | 25.61 | 299.50 |
| 14 | 3:15 | 55 | 2.5 | 4.9 | 25.79 | 304.80 |
| 15 | 1:45 | 55 | 1.0 | 5.6 | 23.87 | 250.05 |
| 16 | 3:15 | 45 | 1.0 | 5.6 | 25.05 | 220.80 |
| 17 | 1:00 | 50 | 2.0 | 5.3 | 21.87 | 281.65 |
| 18 | 2:30 | 60 | 2.0 | 5.3 | 27.43 | 209.10 |
| 19 | 2:30 | 50 | 2.0 | 5.3 | 24.41 | 261.15 |
| 20 | 2:30 | 50 | 2.0 | 5.3 | 24.84 | 232.00 |
| 21 | 4:00 | 50 | 2.0 | 5.3 | 25.42 | 269.90 |
| 22 | 3:15 | 55 | 1.0 | 4.9 | 26.58 | 283.60 |
| 23 | 1:45 | 45 | 1.0 | 4.9 | 23.82 | 216.35 |
| 24 | 2:30 | 50 | 0.5 | 5.3 | 23.49 | 245.10 |
| 25 | 3:15 | 55 | 1.0 | 4.9 | 27.03 | 291.80 |
| 26 | 3:15 | 45 | 1.0 | 5.6 | 24.89 | 302.20 |
| 27 | 1:45 | 45 | 2.5 | 4.9 | 23.90 | 221.20 |
| 28 | 3:15 | 55 | 2.5 | 4.9 | 25.30 | 291.95 |
| 29 | 1:45 | 55 | 1.0 | 5.6 | 24.20 | 271.10 |
| Time, h | |||||
|---|---|---|---|---|---|
| 0 | 24 | 48 | 72 | 98 | |
| pH | 4.95 ± 0.020 c | 4.68 ± 0.021 a | 4.71 ± 0.041 a | 4.81 ± 0.013 b | 4.82 ± 0.042 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puzeryte, V.; Martusevice, P.; Sousa, S.; Balciunaitiene, A.; Viskelis, J.; Gomes, A.M.; Viskelis, P.; Cesoniene, L.; Urbonaviciene, D. Optimization of Enzyme-Assisted Extraction of Bioactive Compounds from Sea Buckthorn (Hippophae rhamnoides L.) Leaves: Evaluation of Mixed-Culture Fermentation. Microorganisms 2023, 11, 2180. https://doi.org/10.3390/microorganisms11092180
Puzeryte V, Martusevice P, Sousa S, Balciunaitiene A, Viskelis J, Gomes AM, Viskelis P, Cesoniene L, Urbonaviciene D. Optimization of Enzyme-Assisted Extraction of Bioactive Compounds from Sea Buckthorn (Hippophae rhamnoides L.) Leaves: Evaluation of Mixed-Culture Fermentation. Microorganisms. 2023; 11(9):2180. https://doi.org/10.3390/microorganisms11092180
Chicago/Turabian StylePuzeryte, Viktorija, Paulina Martusevice, Sérgio Sousa, Aiste Balciunaitiene, Jonas Viskelis, Ana Maria Gomes, Pranas Viskelis, Laima Cesoniene, and Dalia Urbonaviciene. 2023. "Optimization of Enzyme-Assisted Extraction of Bioactive Compounds from Sea Buckthorn (Hippophae rhamnoides L.) Leaves: Evaluation of Mixed-Culture Fermentation" Microorganisms 11, no. 9: 2180. https://doi.org/10.3390/microorganisms11092180
APA StylePuzeryte, V., Martusevice, P., Sousa, S., Balciunaitiene, A., Viskelis, J., Gomes, A. M., Viskelis, P., Cesoniene, L., & Urbonaviciene, D. (2023). Optimization of Enzyme-Assisted Extraction of Bioactive Compounds from Sea Buckthorn (Hippophae rhamnoides L.) Leaves: Evaluation of Mixed-Culture Fermentation. Microorganisms, 11(9), 2180. https://doi.org/10.3390/microorganisms11092180