
Trypanosome Infections and Anemia in Cattle Returning from Transhumance in Tsetse-Infested Areas of Cameroon
 ,
,
Abstract
1. Introduction
- Nomadism: Herders do not have a fixed home and move throughout the year in search of water and fodder for their animals;
- Transhumance: Herders have a permanent residence but move seasonally, depending on the rainy and dry seasons, in search of better grasslands and water availability;
- Sedentary breeding: The breeders keep their cattle close to the village where they live. However, during the dry season, they sometimes entrust part of their herd to a transhumant herder. The sedentary population is typically made up of farmers–breeders, with livestock being a complement to crop production.
2. Methods
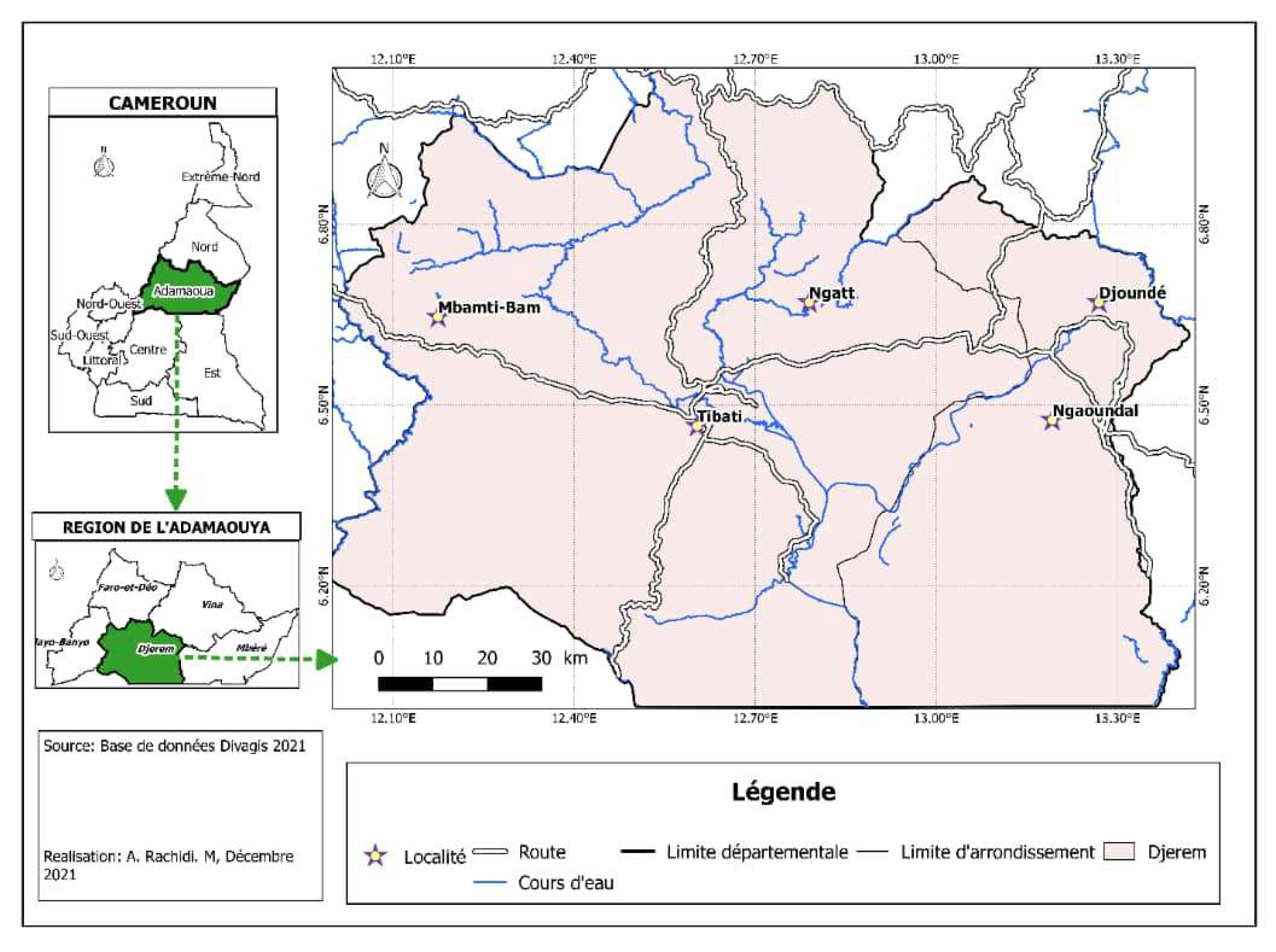
2.1. Sites of the Study
2.2. Blood Sample Collection in Cattle
2.3. Assessment of the Anemic Status of Cattle
2.4. Detection of Trypanosome Infections Using Immunological Test of the Very Diag Kit
2.5. Detection of Trypanosome Infections Using PCR-Based Method
2.5.1. DNA Extraction from Cattle Blood
2.5.2. Molecular Identification of Different Trypanosome Species in Cattle Blood
2.5.3. Molecular Identification of T. congolense Savannah and Forest “Types”
2.6. Data Analysis
3. Results
3.1. Trypanosome Infections Revealed by Immunological Test: Very Diag Test
3.2. Trypanosome Infections Revealed by Molecular Methods
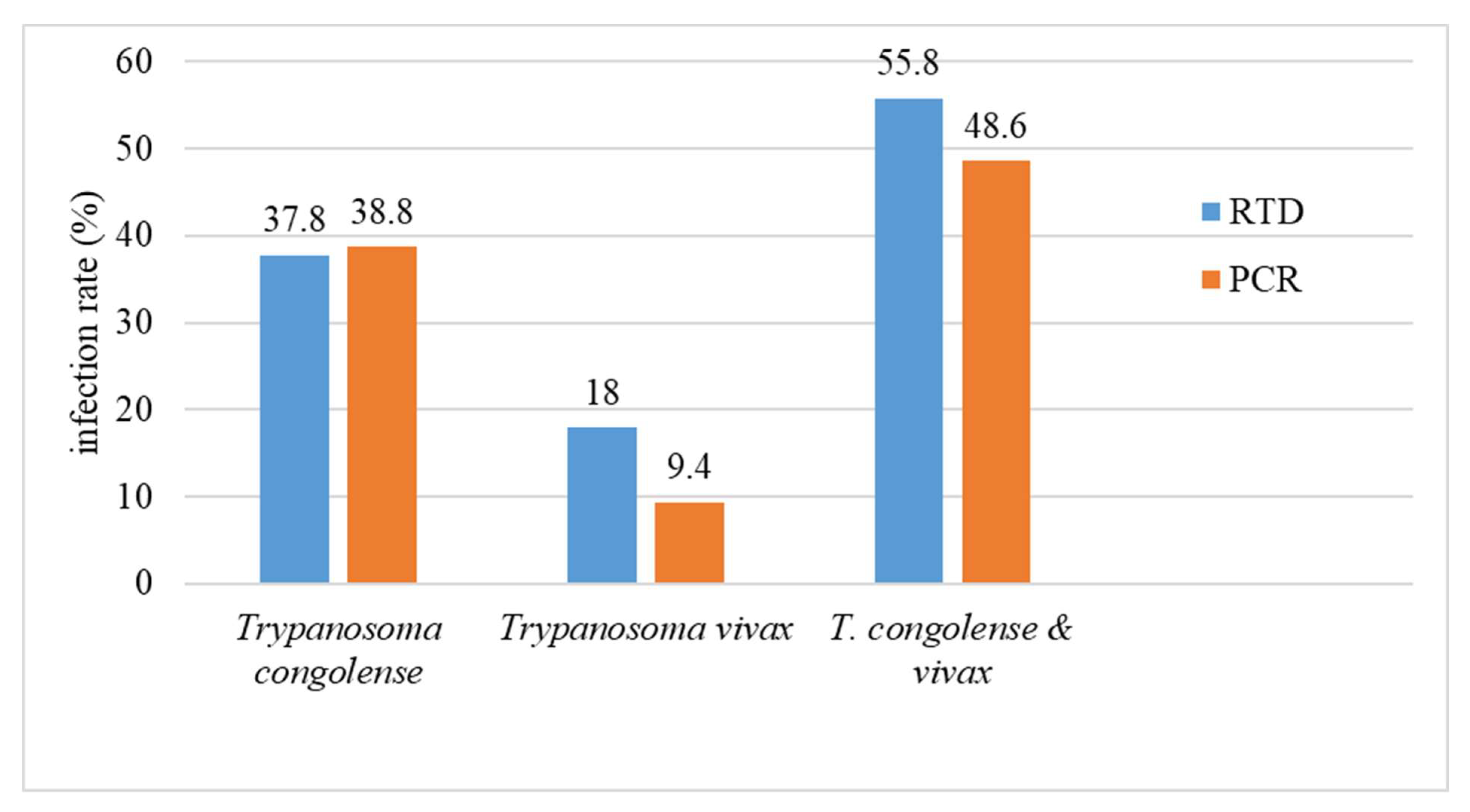
3.3. Comparison between the Infection Rates Recorded by Immunological Tests (Very Diag Kit) and PCR-Based Method
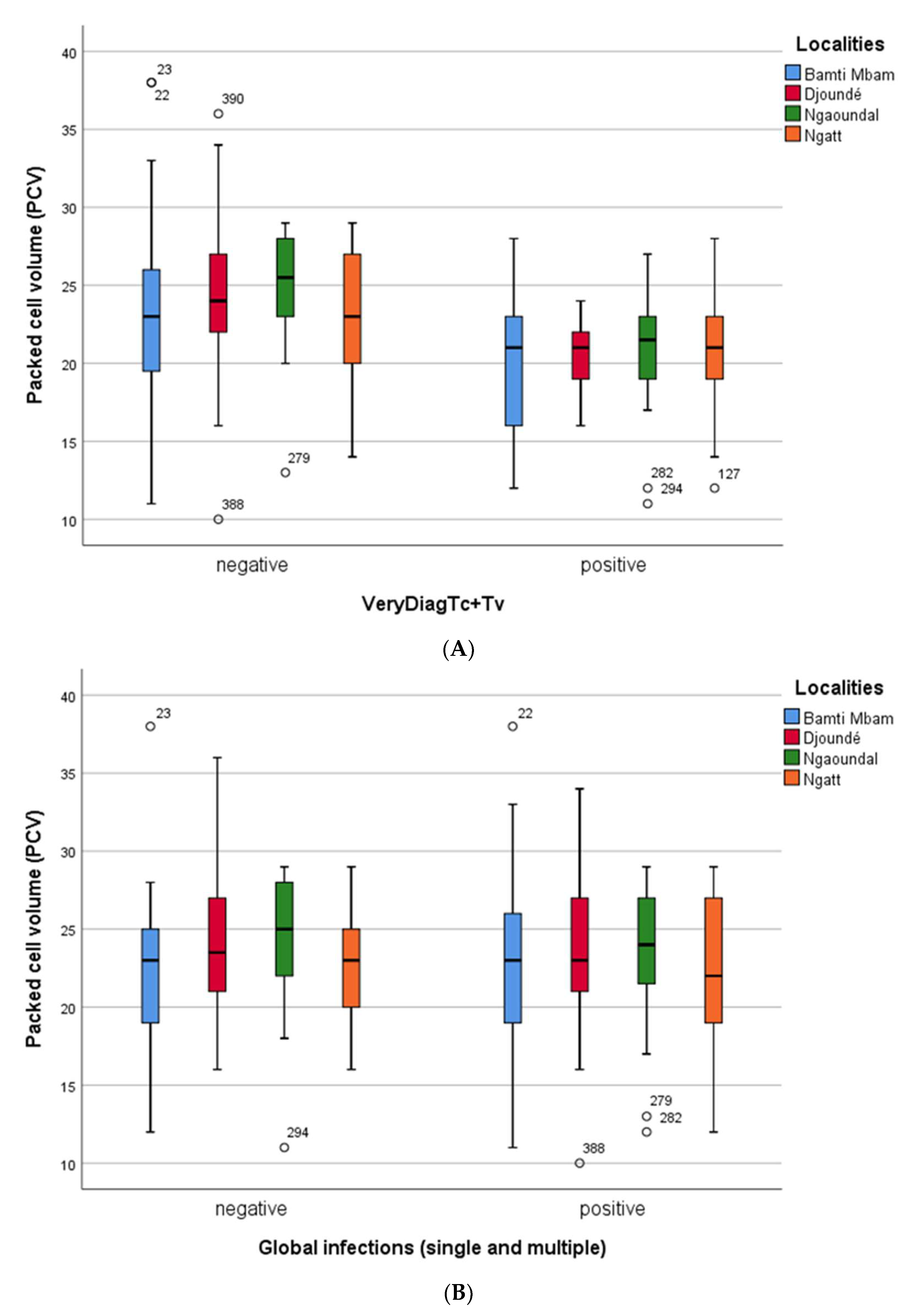
3.4. PCV Values or Anemia Status of Cattle from the Four Villages
3.5. Cattle Anemia: Effect of Trypanosome Infection on Cattle Anemia across the Sampling Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bronsvoort, M.; Tanya, V.N.; Kitching, R.P.; Nfon, C.; Hamman, S.M.; Morgan, K.L. Foot and mouth disease and livestock husbandry practices in the Adamawa Province of Cameroon. Trop. Anim. Health Prod. 2003, 35, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.F.; Hamman, S.M.; Morgan, K.L.; Nkongho, E.F.; Ngwa, V.N.; Tanya, V.; Andu, W.N.; Sander, M.; Ndip, L.; Handel, I.G.; et al. Knowledge of bovine tuberculosis, cattle husbandry and dairy practices amongst pastoralists and small-scale dairy farmers in Cameroon. PLoS ONE 2016, 11, 0146538. [Google Scholar] [CrossRef] [PubMed]
- Scoones, I.; Wolmer, W. Livestock, Disease, Trade and Markets: Policy Choices for the Livestock Sector in Africa; Technical Report; Institute for Development Studies, University of Sussex: Brighton, UK, 2006. [Google Scholar]
- Motta, P.; Porphyre, T.; Hamman, S.M.; Morgan, K.L.; Ngwa, V.N.; Vincent, N.; Tanya, V.N.; Raizman, E.; Handel, I.G.; Bronsvoort, B.M. Cattle transhumance and agropastoral nomadic herding practices in Central Cameroon. BMC Vet. Res. 2018, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.G. Livestock mobility and animal health policy in southern Africa: The impact of veterinary cordon fences on pastoralists. Hum. Ecol. 2002, 30, 215–226. [Google Scholar] [CrossRef]
- Pamo, E.T. Country Pasture/Forage Resource Profiles—Cameroon; Technical Report; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008. [Google Scholar]
- Tanenbe, C.; Gambo, H.; Musongong, G.A.; Boris, O.; Achukwi, M.D. Prévalence de la trypanosomose bovine dans les départements du Faro et Déo, et de la Vina au Cameroun: Bilan de vingt années de lutte contre les glossines. Rev. Elev. Med. Vet. Pays Trop. 2010, 63, 63–69. [Google Scholar] [CrossRef]
- Farikou, O.; Kame-Ngasse, G.I.; Simo, G.; Feudjio, S.S.; Banipé, L.; Njiokou, F.; Geiger, A. Diversity of tsetse flies and trypanosome species circulating in the highly infested cattle rearing area of the Faro and Deo subdivision, Adamawa region, Cameroon. Vet. Parasitol. Reg. Stud. Rep. 2022, 35, 100783. [Google Scholar] [CrossRef] [PubMed]
- Masiga, D.K.; Smyth, A.J.; Hayes, P.; Bromidge, T.J.; Gibson, W.C. Sensitive detection of trypanosomes in tsetse flies by DNA amplification. Int. J. Parasitol. 1992, 22, 909–918. [Google Scholar] [CrossRef]
- Moser, D.R.; Cook, G.A.; Ochs, D.E.; Bailey, C.P.; McKane, M.R.; Donelson, J.E. Detection of Trypanosoma congolense and Trypanosoma brucei subspecies by DNA amplification using the polymerase chain reaction. Parasitology 1989, 99, 57–66. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 3, 174. [Google Scholar] [CrossRef]
- Desquesnes, M.; Gonzatti, M.; Sazmand, M.; Thévenon, S.; Bossard, G.; Boulangé, A.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S.; et al. A review on the diagnosis of animal trypanosomoses. Parasit Vectors 2022, 15, 64. [Google Scholar] [CrossRef]
- Pillay, D.; Izotte, J.; Fikru, R.; Büscher, P.; Mucache, H.; Neves, L.; Boulangé, A.; Seck, M.T.; Bouyer, J.; Napier, G.B.; et al. Trypanosoma vivax GM6 antigen: A candidate antigen for diagnosis of African animal trypanosomosis in cattle. PLoS ONE 2013, 8, e78565. [Google Scholar] [CrossRef]
- Boulangé, A.; Pillay, D.; Chevtzoff, C.; Biteau, N.; Comé de Graça, V.; Rempeters, L.; Theodoridis, D.; Baltz, T. Development of a rapid antibody test for point-of-care diagnosis of animal African trypanosomosis. Vet. Parasitol. 2017, 233, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Lumbala, C.; Biéler, S.; Kayembe, S.; Makabuza, J.; Ongarello, S.; Ndung’u, J.M. Prospective evaluation of a rapid diagnostic test for Trypanosoma brucei gambiense infection developed using recombinant antigens. PLoS Negl. Trop. Dis. 2018, 12, e0006386. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Albizu, C.; Danesi, E.; Piorno, P.; Fernandez, M.; Campos, F.G.; Scollo, K.; Crud, F. Rapid Diagnostic Tests for Trypanosoma cruzi Infection: Field Evaluation of Two Registered Kits in a Region of Endemicity and a Region of Nonendemicity in Argentina. J. Clin. Microbiol. 2020, 58, e01140-20. [Google Scholar] [CrossRef]
- Whitman, J.D.; Bulman, C.A.; Gunderson, E.L.; Irish, A.M.; Townsend, R.L.; Stramer, S.L.; Sakanari, J.A.; Bern, C. Chagas disease serological test performance in U.S. blood donor specimens. J. Clin. Microbiol. 2019, 57, e01217-19. [Google Scholar] [CrossRef]
- Roddy, P.; Goiri, J.; Flevaud, L.; Palma, P.P.; Morote, S.; Lima, N.; Villa, L.; Torrico, F.; Albajar-Vinas, P. Field evaluation of a rapid immunochromatographic assay for detection of Trypanosoma cruzi infection by use of whole blood. J. Clin. Microbiol. 2008, 46, 2022–2027. [Google Scholar] [CrossRef]
- Luquetti, A.O.; Ponce, C.; Ponce, E.; Esfandiari, J.; Schijman, A.; Revollo, S.; Anez, N.; Zingales, B.; Ramgel-Aldao, R.; Gonzalez, A.; et al. Chagas’ disease diagnosis: A multicentric evaluation of Chagas Stat-Pak, a rapid immunochromatographic assay with recombinant proteins of Trypanosoma cruzi. Diagn. Microbiol. Infect. Dis. 2003, 46, 265–271. [Google Scholar] [CrossRef]
- Ponce, C.; Ponce, E.; Vinelli, E.; Montoya, A.; de Aguilar, V.; Gonzalez, A.; Zingales, B.; Rangel-Aldao, R.; Levin, M.J.; Esfandiari, J.; et al. Validation of a rapid and reliable test for diagnosis of Chagas’ disease by detection of Trypanosoma cruzi-specific antibodies in blood of donors and patients in Central America. J. Clin. Microbiol. 2005, 43, 5065–5068. [Google Scholar] [CrossRef]
- Chappuis, F.; Mauris, A.; Holst, M.; Albajar-Vinas, P.; Jannin, J.; Luquetti, A.O.; Jackson, Y. Validation of a rapid immunochromatographic assay for diagnosis of Trypanosoma cruzi infection among Latin-American Migrants in Geneva, Switzerland. J. Clin. Microbiol. 2010, 48, 2948–2952. [Google Scholar] [CrossRef]
- Sosa-Estani, S.; Gamboa-Leon, M.R.; Del Cid-Lemus, J.; Althabe, F.; Alger, J.; Almendares, O.; Cafferata, M.L.; Chippaux, J.P.; Dumonteil, E.; Gibbons, L.; et al. Use of a rapid test on umbilical cord blood to screen for Trypanosoma cruzi infection in pregnant women in Argentina, Bolivia, Honduras, and Mexico. Am. J. Trop. Med. Hyg. 2008, 79, 755–759. [Google Scholar] [CrossRef]
- Oumarou, F.; Flobert, N.; Gustave, S.; Irma, K.N.G.; Steve, S.; Louis, B.; Anne, G. Prevalence of trypanosome infection in three cattle breeds and effect on cattle annemia in the Dodeo cattle rearing area of Cameroun. Int. J. Vet. Sci. 2022, 3, 119. [Google Scholar]
- Paguem, A.; Abanda, B.; Ndjonka, D.; Weber, J.S.; Ngomtcho, S.C.H.; Manchang, K.T.; Achukwi, M.D. Widespread co-endemicity of Trypanosoma species infecting cattle in the Sudano-Sahelian and Guinea Savannah zones of Cameroon. BMC Vet. Res. 2019, 15, 344. [Google Scholar] [CrossRef] [PubMed]
- Kame-Ngasse, G.I.; Njiokou, F.; Melachio-Tanekou, T.T.; Farikou, O.; Simo, G.; Geiger, A. Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa Region of Cameroon. BMC Microbiol. 2018, 18, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Ngomtcho, S.C.H.; Weber, J.S.; Bum, E.N.; Gbem, T.T.; Kelm, S.; Achukwi, M.D. Molecular screening of tsetse flies and cattle reveal different Trypanosoma species including T. grayi and T. theileri in northern Cameroon. Parasit. Vectors 2017, 10, 631. [Google Scholar] [CrossRef]
- Sima, N.; Dujeancourt-Henry, A.; Perlaza, B.L.; Ungeheuer, M.N.; Rotureau, B.; Glovera, L. SHERLOCK4HAT: A CRISPR-based tool kit for diagnosis of Human African Trypanosomiasis. eBioMedicine 2022, 85, 104308. [Google Scholar] [CrossRef]
- Büscher, P.; Mertens, P.; Leclipteux, T.; Gilleman, Q.; Jacquet, D.; Mumba-Ngoyi, D.; Pyana, P.P.; Boelaert, M.; Lejon, V. Sensitivity and specificity of HAT Sero-K-SeT, a rapid diagnostic test for serodiagnosis of sleeping sickness caused by Trypanosoma brucei gambiense: A case-control study. Lancet Glob. Health 2014, 2, e359–e363. [Google Scholar] [CrossRef] [PubMed]
- Awa, D.N.; Achukwi, M.D. Livestock pathology in the central African region:some epidemiological considerations and control strategies. Anim. Health Res. Rev. 2010, 11, 235–244. [Google Scholar] [CrossRef]
- Njiokou, F.; Laveissière, C.; Simo, G.; Nkinin, S.; Grébaut, P.; Cuny, G.; Herder, S. Wild fauna as a probable animal reservoir for Trypanosoma brucei gambiense in Cameroon. Infect. Genet. Evol. 2006, 6, 147–153. [Google Scholar] [CrossRef]
- Simo, G.; Njiokou, F.; Mbida Mbida, J.A.; Njitchouang, G.R.; Herder, S.; Asonganyi, T.; Cuny, G. Tsetse fly host preference from sleeping sickness foci in Cameroon: Epidemiological implications. Infect. Genet. Evol. 2008, 8, 34–39. [Google Scholar] [CrossRef]
- Farikou, O.; Njiokou, F.; Simo, G.; Asonganyi, T.; Cuny, G.; Geiger, A. Tsetse fly blood meal modification and trypanosome identification in two sleeping sickness foci in the forest of Southern Cameroon. Acta Trop. 2010, 116, 81–88. [Google Scholar] [CrossRef]
- Ministère de l’Elevage, des Pêches et des Industries Animales (MINEPIA). Rapportdéfinitif du Plan de Gestion des Antiparasitaires et des Antimicrobiens (pg2a) du Projet de Développement de L’élevage (PRODEL); MINEPIA: Yaoundé, Cameroon, 2016; 64p. [Google Scholar]
- Macpherson, C.N.L. The effect of transhumance on the epidemiology of animal diseases. Prev. Vet. Med. 1995, 25, 213–224. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Villages | Nb of Samples | Tc s.l. (%) | Tvx (%) | Single Infections (%) | Mixed Infections Tc/Tvx (%) | Global Infection (%) |
|---|---|---|---|---|---|---|
| Bamti Mbam | 100 | 49 (49%) | 17 (17%) | 66 (66%) | 13 (13%) | 79 (79%) |
| Djoundé | 105 | 35 (33.3%) | 20 (19.0%) | 55 (52.4%) | 22 (20.9%) | 77 (73.3%) |
| Ngaoundal | 100 | 36 (36%) | 17 (17%) | 53 (53%) | 22 (22%) | 75 (75%) |
| Ngatt | 100 | 33 (33%) | 19 (19%) | 52 (52%) | 27 (27%) | 79 (79%) |
| Total (Nb and mean %) | 405 | 153 (37.8%) | 73 (18.0%) | 226 (55.8%) | 84 (20.7%) | 310 (76.5%) |
| χ2 | / | 7.346 | 0.281 | 5.619 | 6.128 | 1.408 |
| P | / | 0.062 | 0.964 | 0.132 | 0.106 | 0.704 |
| Villages | Nb Samples | Tcs (%) | Tcf (%) | Tvx (%) | Tbr (%) | Simple Infection (%) | Tcs/Tcf (%) | Tcs/Tbr (%) | Tcs/Tvx (%) | Tcf/Tbr (%) | Tcf/Tvx (%) | Tvx/Tbr (%) | Tcs/Tcf/Tbr (%) | Mixed Infection (%) | Global Infection (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bamti Mbam | 100 | 26 (26%) | 4 (4%) | 12 (12%) | 12 (12%) | 54 (54%) | 3 (3%) | 6 (6%) | 3 (3%) | 1 (1%) | 1 (1%) | 0 | 0 | 14 (14%) | 69 (69%) |
| Djoundé | 105 | 35 (33.3%) | 11 (10.5%) | 6 (5.7%) | 3 (2.8%) | 55 (52.4%) | 1 (0.95%) | 8 (7.6%) | 2 (1.9%) | 3 (2.8%) | 2 (1.9%) | 5 (4.8%) | 3 (2.8%) | 24 (22.9%) | 79 (75.2%) |
| Ngaoundal | 100 | 38 (38%) | 8 (8%) | 6 (6%) | 6 (6%) | 58 (58%) | 2 (2%) | 0 | 4 (4%) | 0 | 1 (1%) | 5 (5%) | 3 (3%) | 15 (15%) | 72 (72%) |
| Ngatt | 100 | 30 (30%) | 5 (5%) | 14 (14%) | 4 (4%) | 53 (53%) | 0 | 3 (3%) | 2 (2%) | 0 | 0 | 0 | 0 | 5 (5%) | 58 (58%) |
| Total | 405 | 129 (31.8%) | 28 (6.9%) | 38 (9.4%) | 25 (6.2%) | 220 (54.3%) | 6 (1.5%) | 17 (4.2%) | 11 (2.7%) | 4 (0.98%) | 4 (0.98%) | 10 (2.5%) | 6 (1.5%) | 58 (14.3%) | 278 (68.6%) |
| χ2 | / | 3.58 | 4.14 | 6.32 | 8.67 | 0.78 | 3.46 | 8.60 | 1.11 | 5.74 | 1.90 | 10.01 | 5.94 | 13.36 | 7.91 |
| P | / | 0.31 | 0.24 | 0.09 | 0.03 | 0.85 | 0.32 | 0.03 | 0.77 | 0.12 | 0.59 | 0.02 | 0.11 | 0.004 | 0.05 |
| Villages | Nb of Sample | Mean | Standard Deviation |
|---|---|---|---|
| Bamti Mbam | 100 | 22.36 | 5.206 |
| Djoundé | 105 | 23.58 | 4.104 |
| Ngaoundal | 100 | 24.12 | 3.836 |
| Ngatt | 100 | 22.54 | 4.014 |
| Total | 405 | 23.15 | 4.365 |
| F | / | 3.8 | / |
| p | / | 0.01 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farikou, O.; Simo, G.; Njiokou, F.; Kamé Ngassé, G.I.; Achiri Fru, M.; Geiger, A. Trypanosome Infections and Anemia in Cattle Returning from Transhumance in Tsetse-Infested Areas of Cameroon. Microorganisms 2023, 11, 712. https://doi.org/10.3390/microorganisms11030712
Farikou O, Simo G, Njiokou F, Kamé Ngassé GI, Achiri Fru M, Geiger A. Trypanosome Infections and Anemia in Cattle Returning from Transhumance in Tsetse-Infested Areas of Cameroon. Microorganisms. 2023; 11(3):712. https://doi.org/10.3390/microorganisms11030712
Chicago/Turabian StyleFarikou, Oumarou, Gustave Simo, Flobert Njiokou, Ginette Irma Kamé Ngassé, Martin Achiri Fru, and Anne Geiger. 2023. "Trypanosome Infections and Anemia in Cattle Returning from Transhumance in Tsetse-Infested Areas of Cameroon" Microorganisms 11, no. 3: 712. https://doi.org/10.3390/microorganisms11030712
APA StyleFarikou, O., Simo, G., Njiokou, F., Kamé Ngassé, G. I., Achiri Fru, M., & Geiger, A. (2023). Trypanosome Infections and Anemia in Cattle Returning from Transhumance in Tsetse-Infested Areas of Cameroon. Microorganisms, 11(3), 712. https://doi.org/10.3390/microorganisms11030712