
The Effect of Recombinant Protein Production in Lactococcus lactis Transcriptome and Proteome



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. L. lactis Core Genome and Proteome
3. Transcriptome and Proteome Profiles of L. lactis in Response to Natural Stresses
4. Transcriptome and Proteome Profiles of L. lactis in Response to Plasmid DNA and Recombinant Protein Production
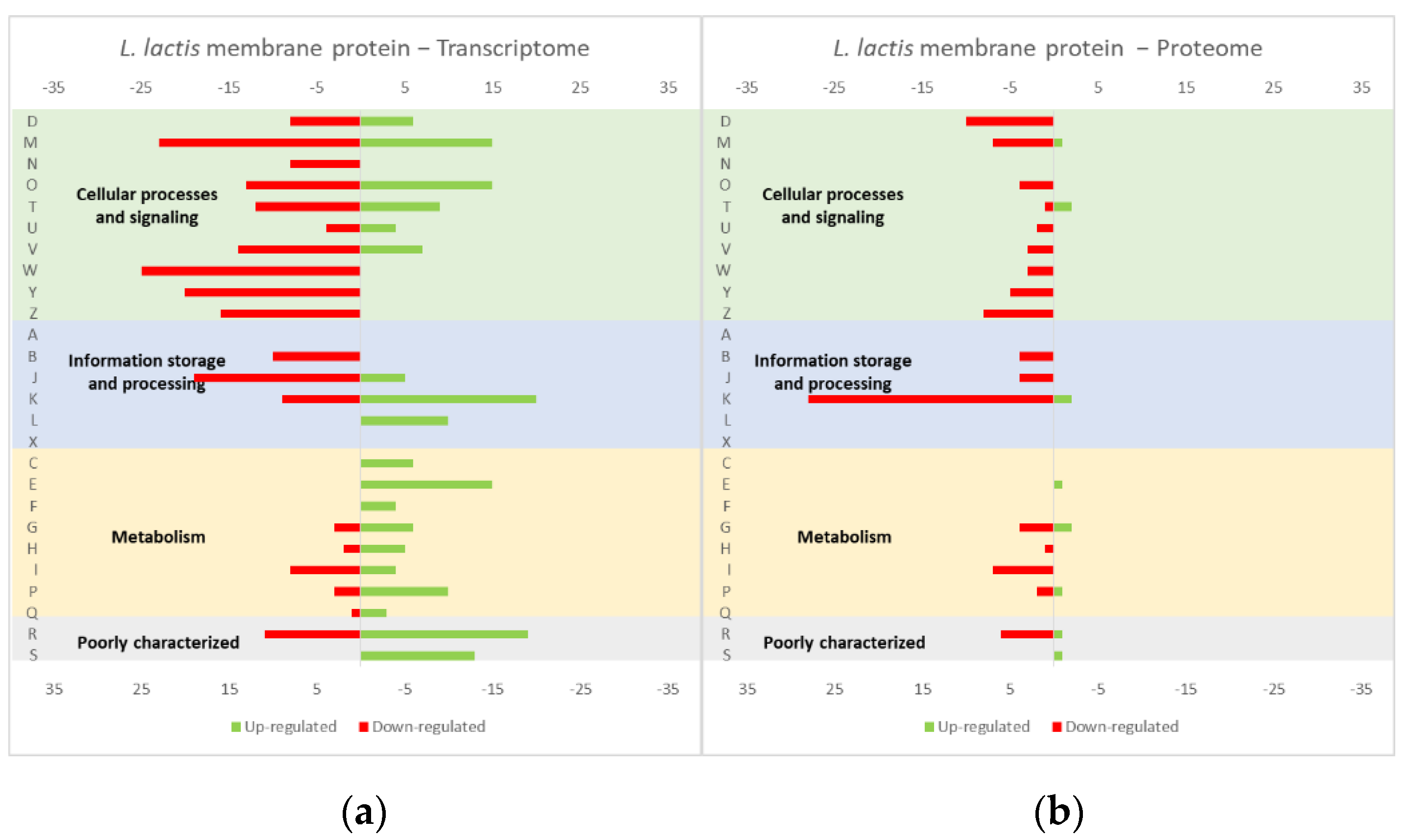
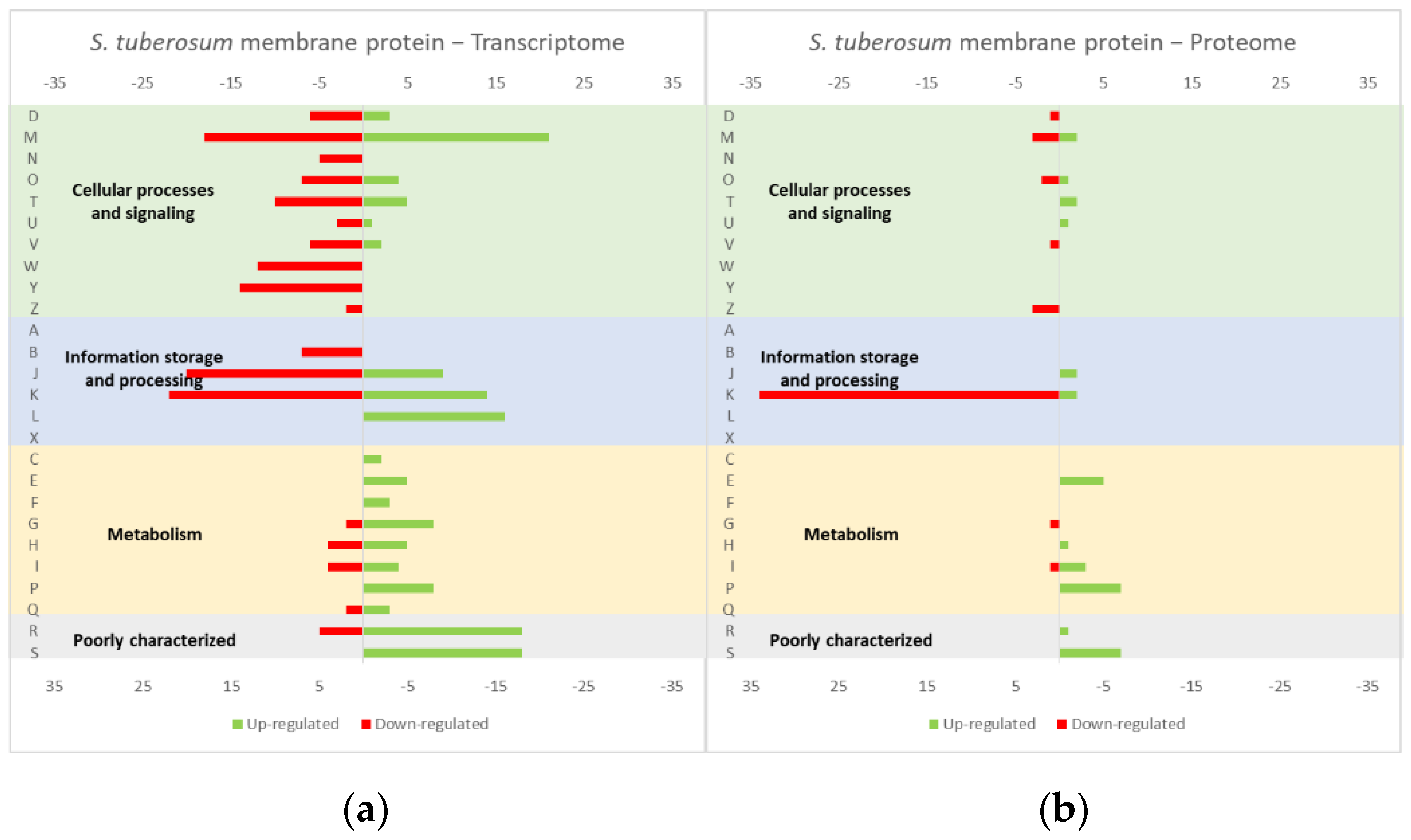
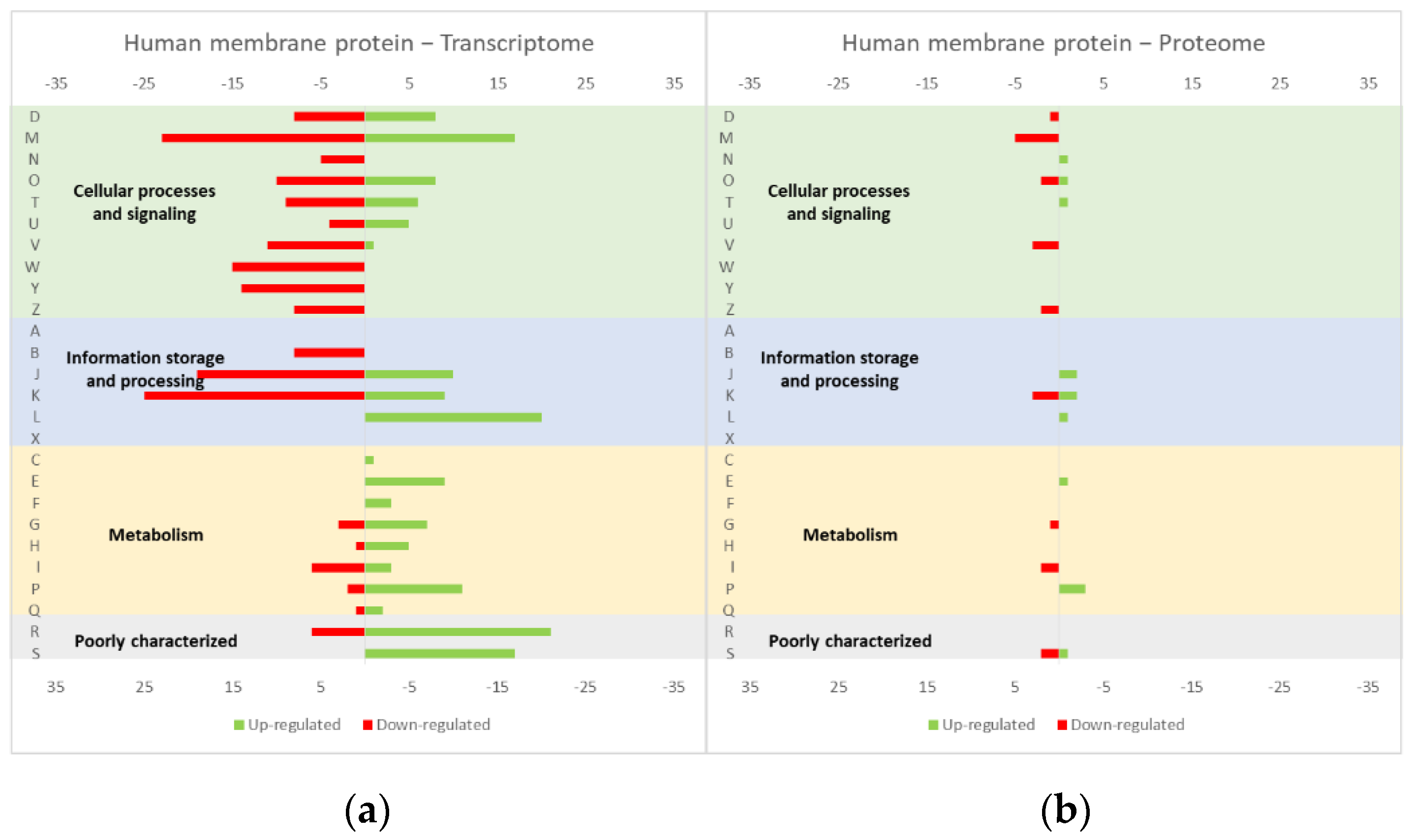
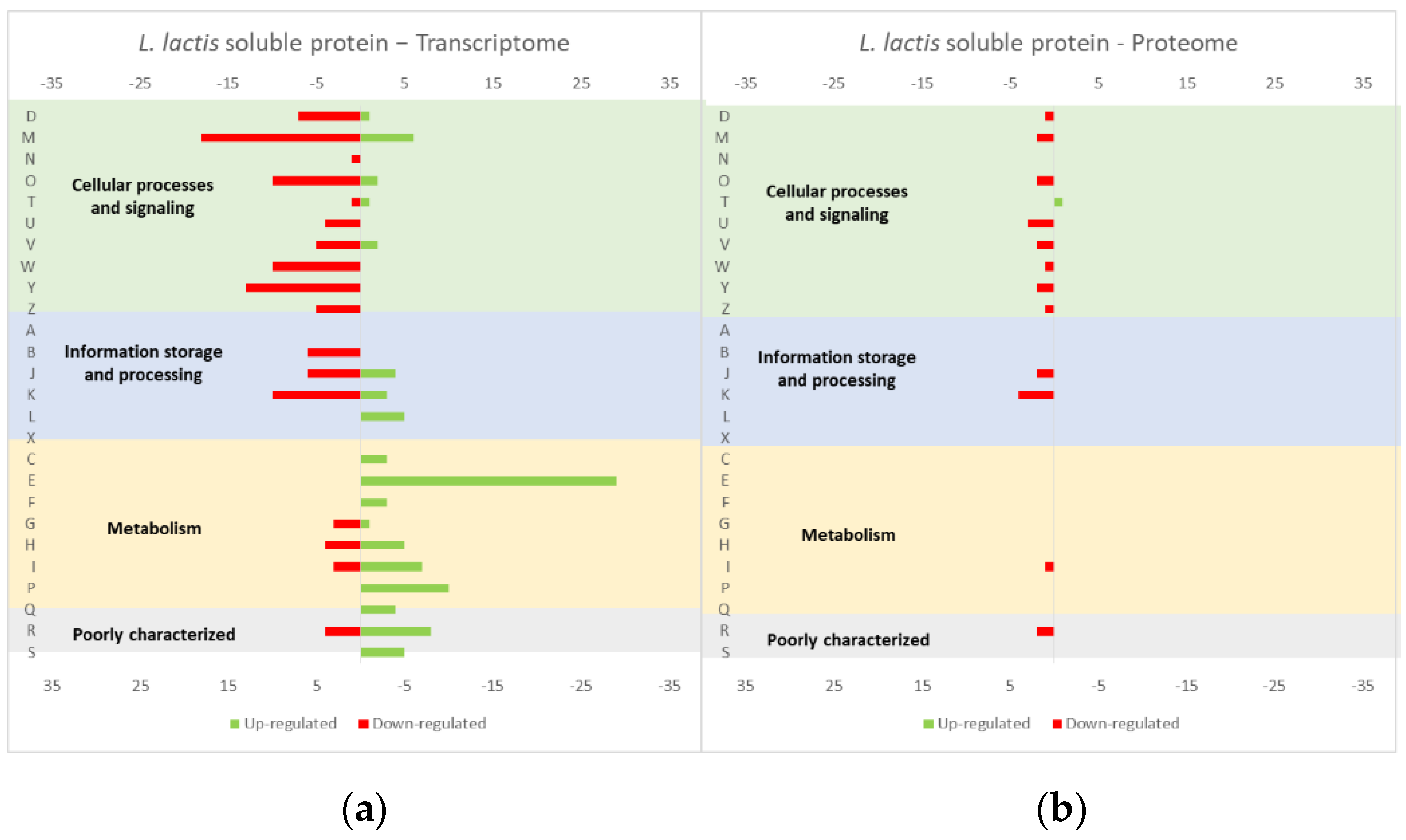
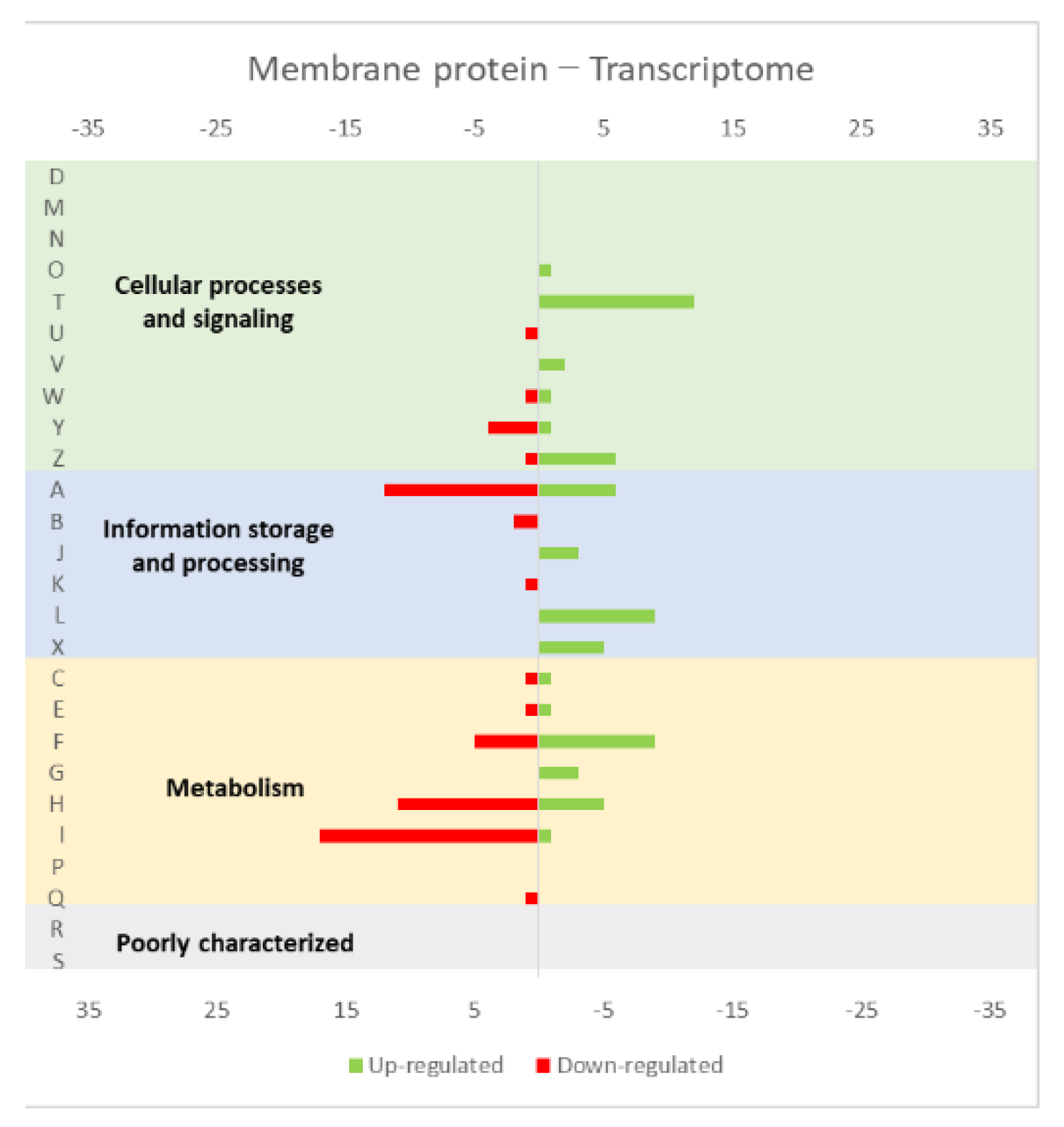
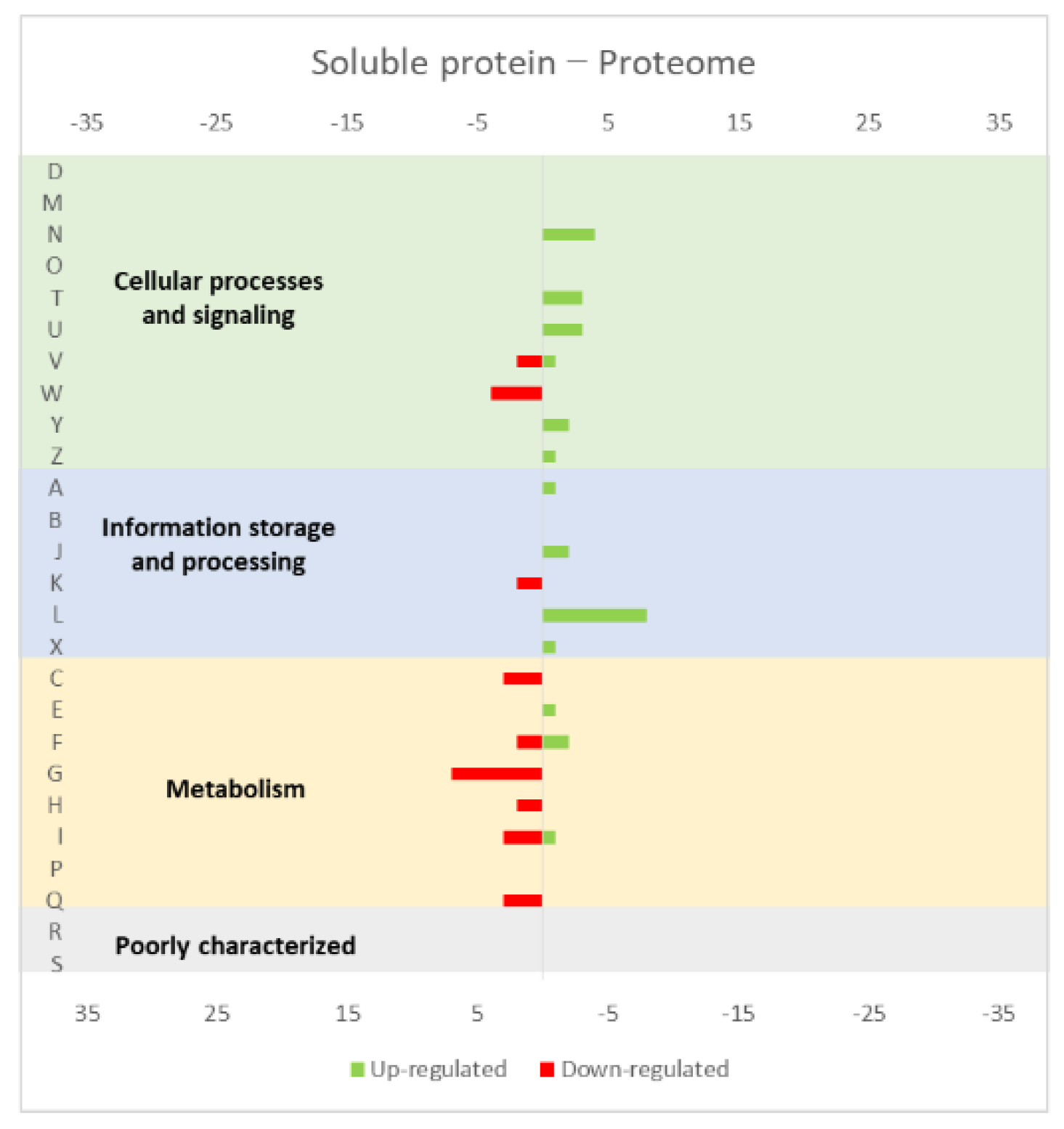
4.1. Effect of the Overexpression of Membrane and Soluble Proteins in the L. lactis Transcriptome and Proteome
4.2. Effect of the Growth Rate in the Proteome of L. lactis
4.3. Effect of Plasmid Copy Number in the Proteome of L. lactis
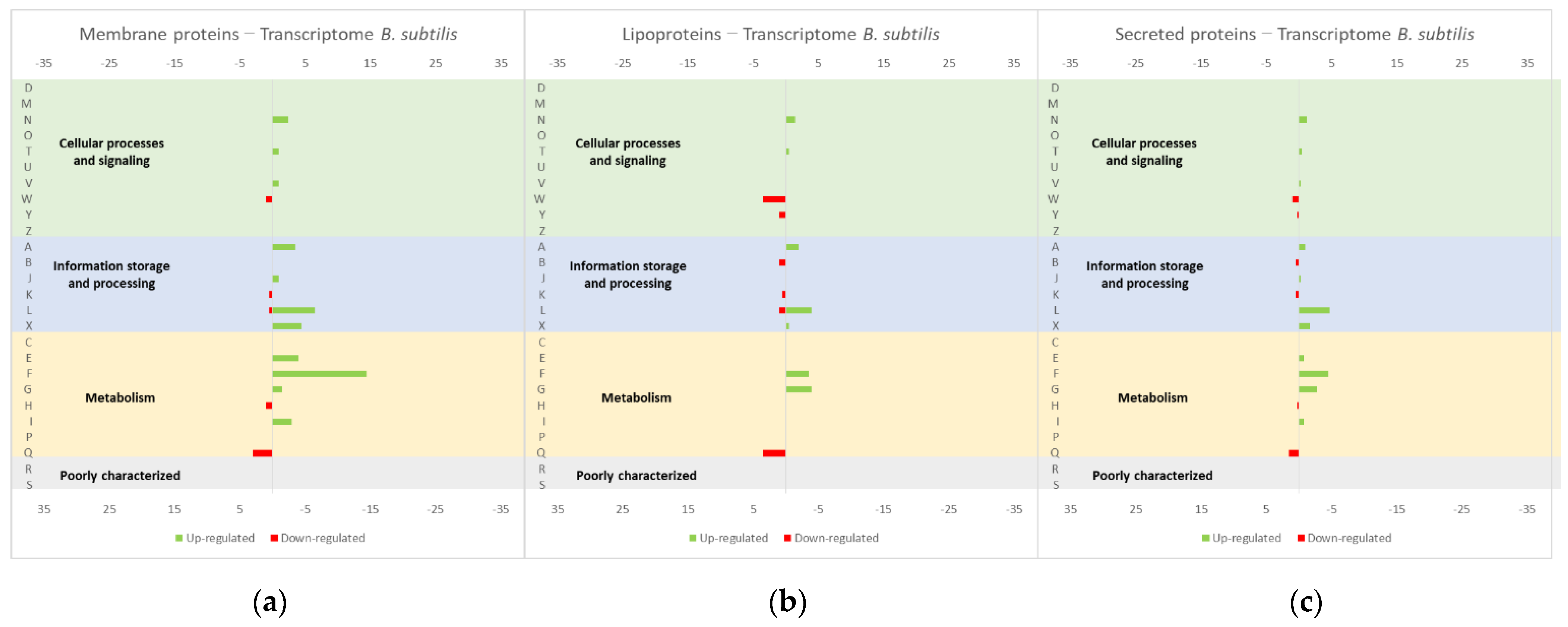
4.4. Insights from E. coli and B. subtilis Studies
4.5. Effect of Different Stresses in the L. lactis Transcriptome and Proteome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wood, B.J.B.; Holzapfel, W.H. The Genera of Lactic Acid Bacteria; Springer: Boston, MA, USA, 1995. [Google Scholar]
- Bolotin, A.; Wincker, P.; Mauger, S.; Jaillon, O.; Malarme, K.; Weissenbach, J.; Ehrlich, S.D.; Sorokin, A. The Complete Genome Sequence of the Lactic Acid Bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-HumaránL, G.; Kharrat, P.; Chatel, J.-M.; Langella, P. Lactococci and lactobacilli as mucosal delivery vectors for therapeutic proteins and DNA vaccines. Microb. Cell Factories 2011, 10, S4. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.; Pandeya, D.R.; Hong, S.T. Lactococcus lactis: An efficient gram positive cell factory for the production and secretion of recombinant protein. Biomed. Res. 2012, 23, 1. [Google Scholar]
- Le Loir, Y.; Azevedo, V.; Oliveira, S.C.; Freitas, D.A.; Miyoshi, A.; Bermúdez-Humaran, L.G.B.; Nouaille, S.; Ribeiro, A.L.; Leclercq, S.; Gabriel, E.J.; et al. Protein secretion in Lactococcus lactis: An efficient way to increase the overall heterologous protein production. Microb. Cell Factories 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Gram, G.J.; Fomsgaard, A.; Thorn, M.; Madsen, S.M.; Glenting, J. Immunological analysis of a Lactococcus lactis-based DNA vaccine expressing HIV gp120. Genet. Vaccines Ther. 2007, 5, 3. [Google Scholar] [CrossRef]
- Pedersen, M.L.; Arrived, K.R.; Johansen, E. Genetic analysis of the minimal replicon of the Lactococcus lactis subsp. lactis biovar diacetylactis citrate plasmid. Mol. Genet. Genom. 1994, 244, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.B.; Turk, M.Z.; Saraiva, T.D.L.; De Castro, C.P.; Souza, B.M.; Agresti, P.M.; Lima, F.A.; Pfeiffer, V.N.; de Azevedo, M.S.P.; Rocha, C.S.; et al. DNA Vaccines Approach: From Concepts to Applications. World J. Vaccines 2014, 4, 50–71. [Google Scholar] [CrossRef]
- SilvaI, N.; Duarte, S.; Moreira, L.M.; Monteiro, G.A. Draft Genome Sequence of the Plasmid-Free Lactococcus lactis subsp. lactis Strain LMG 19460. Genome Announc. 2017, 5, e00210-17. [Google Scholar] [CrossRef]
- Ideker, T.; Galitski, T.; Hood, L. A new approach to decoding life: Systems Biology. Annu. Rev. Genom. Hum. Genet. 2001, 2, 343–372. [Google Scholar] [CrossRef]
- Kitano, H. Systems Biology: A Brief Overview. Science 2002, 295, 1662–1664. [Google Scholar] [CrossRef]
- Rathore, A.S.; Chauhan, A. Transcriptomics and the production of recombinant therapeutics. Biopharm Int. 2018, 31, 22–28. [Google Scholar]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, P.; Bottacini, F.; Mahony, J.; Kilcawley, K.N.; Van Sinderen, U. Comparative and functional genomics of the Lactococcus lactis taxon; insights into evolution and niche adaptation. BMC Genom. 2017, 18, 267. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.M.; Sousa, C.; Oliveira, L.D.C.; Soares, S.C.; Souza, G.H.M.F.; Tavares, G.C.; Resende, C.P.; Folador, E.L.; Pereira, F.L.; Figueiredo, H.; et al. Comparative proteomic analysis of four biotechnological strains Lactococcus lactisthrough label-free quantitative proteomics. Microb. Biotechnol. 2019, 12, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Wels, M.; Siezen, R.; Van Hijum, S.; Kelly, W.J.; Bachmann, H. Comparative Genome Analysis of Lactococcus lactis Indicates Niche Adaptation and Resolves Genotype/Phenotype Disparity. Front. Microbiol. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Hols, P.; Kleerebezem, M.; Schanck, A.N.; Ferain, T.; Hugenholtz, J.; Delcour, J.; De Vos, W.M. Conversion of Lactococcus lactis from homolactic to homoalanine fermentation through metabolic engineering. Nat. Biotechnol. 1999, 17, 588–592. [Google Scholar] [CrossRef]
- Hugenholtz, J.; Kleerebezem, M.; Starrenburg, M.; Delcour, J.; de Vos, W.; Hols, P. Lactococcus lactis as a Cell Factory for High-Level Diacetyl Production. Appl. Environ. Microbiol. 2000, 66, 4112–4114. [Google Scholar] [CrossRef]
- Sybesma, W.; Starrenburg, M.; Kleerebezem, M.; Mierau, I.; de Vos, W.M.; Hugenholtz, J. Increased Production of Folate by Metabolic Engineering of Lactococcus lactis. Appl. Environ. Microbiol. 2003, 69, 3069–3076. [Google Scholar] [CrossRef]
- Sybesma, W.; Born, E.V.D.; Starrenburg, M.; Mierau, I.; Kleerebezem, M.; de Vos, W.M.; Hugenholtz, J. Controlled Modulation of Folate Polyglutamyl Tail Length by Metabolic Engineeringof Lactococcuslactis. Appl. Environ. Microbiol. 2003, 69, 7101–7107. [Google Scholar] [CrossRef]
- Sybesma, W.; Burgess, C.; Starrenburg, M.; van Sinderen, D.; Hugenholtz, J. Multivitamin production in Lactococcus lactis using metabolic engineering. Metab. Eng. 2003, 6, 109–115. [Google Scholar] [CrossRef]
- Badri, A.; Raman, K.; Jayaraman, G. Uncovering Novel Pathways for Enhancing Hyaluronan Synthesis in Recombinant Lactococcus lactis: Genome-Scale Metabolic Modeling and Experimental Validation. Processes 2019, 7, 343. [Google Scholar] [CrossRef]
- Zhu, D.; Fu, Y.; Liu, F.; Xu, H.; Saris, P.E.J.; Qiao, M. Enhanced heterologous protein productivity by genome reduction in Lactococcus lactis NZ9000. Microb. Cell Factories 2017, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhao, Y.; Du, Y.; Miao, S.; Liu, J.; Li, Y.; Caiyin, Q.; Qiao, J. Quantitative proteomics of Lactococcus lactis F44 under cross-stress of low pH and lactate. J. Dairy Sci. 2018, 101, 6872–6884. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, J.; Miao, S.; Zhao, Y.; Zhu, H.; Qiao, M.; Saris, P.E.J.; Qiao, J. Contribution of YthA, a PspC Family Transcriptional Regulator of Lactococcus lactis F44 Acid Tolerance and Nisin Yield: A Transcriptomic Approach. Appl. Environ. Microbiol. 2018, 84, e02483-17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, P.; Wu, Z.; Zhang, J.; Du, G. Systemic understanding of Lactococcus lactis response to acid stress using transcriptomics approaches. J. Ind. Microbiol. Biotechnol. 2019, 46, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; van Bokhorst-van de Veen, H.; Wels, M.; Kleerebezem, M. Stress Responses of Lactic Acid Bacteria; Tsakalidou, E., Papadimitriou, K., Eds.; Springer: Boston, MA, USA, 2011; Chapter 16; p. 369. [Google Scholar]
- Van Tilburg, A.Y.; Cao, H.; van der Meulen, S.B.; Solopova, A.; Kuipers, O.P. Metabolic engineering and synthetic biology employing Lactococcus lactis and Bacillus subtilis cell factories. Curr. Opin. Biotechnol. 2019, 59, 1–7. [Google Scholar] [CrossRef]
- Kim, M.; Jin, Y.; An, H.J.; Kim, J. Impact of High-Level Expression of Heterologous Protein on Lactococcus lactis Host. J. Microbiol. Biotechnol. 2017, 27, 1345–1358. [Google Scholar] [CrossRef]
- Marreddy, R.K.R.; Pinto, J.P.C.; Wolters, J.C.; Geertsma, E.R.; Fusetti, F.; Permentier, H.P.; Kuipers, O.P.; Kok, J.; Poolman, B. The Response of Lactococcus lactis to Membrane Protein Production. PLoS ONE 2011, 6, e24060. [Google Scholar] [CrossRef][Green Version]
- Teusink, B.; Bachmann, H.; Molenaar, D. Systems biology of lactic acid bacteria: A critical review. Microb. Cell Factories 2011, 10, S11. [Google Scholar] [CrossRef]
- Mairhofer, J.; Scharl, T.; Marisch, K.; Cserjan-Puschmann, M.; Striedner, G. Comparative Transcription Profiling and In-Depth Characterization of Plasmid-Based and Plasmid-Free Escherichia coli Expression Systems under Production Conditions. Appl. Environ. Microbiol. 2013, 79, 3802–3812. [Google Scholar] [CrossRef]
- Birnbaum, S.; Bailey, J.E. Plasmid presence changes the relative levels of many host cell proteins and ribosome components in recombinant Escherichia coli. Biotechnol. Bioeng. 1991, 37, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Neidhardt, I.C.; Ingraham, J.L.; Schaechter, M. Physiology of the Bacterial Cell—A Molecular Approach; Sinauer Associates Inc.: Sunderland, MA, USA, 1990. [Google Scholar]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A Genomic Perspective on Protein Families. Science 1997, 278, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef]
- Wallin, E.; Von Heijne, G. Genome-wide analysis of integral membrane proteins from eubacterial, archaean, and eukaryotic organisms. Protein Sci. 1998, 7, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Drews, J. Drug Discovery: A Historical Perspective. Science 2000, 287, 1960–1964. [Google Scholar] [CrossRef]
- Pinto, J.P.C.; Kuipers, O.P.; Marreddy, R.K.R.; Poolman, B.; Kok, J. Efficient Overproduction of Membrane Proteins in Lactococcus lactis Requires the Cell Envelope Stress Sensor/Regulator Couple CesSR. PLoS ONE 2011, 6, e21873. [Google Scholar] [CrossRef] [PubMed]
- Dressaire, C.; Gitton, C.; Loubière, P.; Monnet, V.; Queinnec, I.; Cocaign-Bousquet, M. Transcriptome and Proteome Exploration to Model Translation Efficiency and Protein Stability in Lactococcus lactis. PLoS Comput. Biol. 2009, 5, e1000606. [Google Scholar] [CrossRef]
- Dressaire, C.; Redon, E.; Milhem, H.; Besse, P.; Loubière, P.; Cocaign-Bousquet, M. Growth rate regulated genes and their wide involvement in the Lactococcus lactis stress responses. BMC Genom. 2008, 9, 343. [Google Scholar] [CrossRef]
- Mata, J.; Marguerat, S.; Bähler, J. Post-transcriptional control of gene expression: A genome-wide perspective. Trends Biochem. Sci. 2005, 30, 506–514. [Google Scholar] [CrossRef]
- Wu, G.; Nie, L.; Zhang, W. Integrative Analyses of Posttranscriptional Regulation in the Yeast Saccharomyces cerevisiae Using Transcriptomic and Proteomic Data. Curr. Microbiol. 2008, 57, 18–22. [Google Scholar] [CrossRef]
- Nie, L.; Wu, G.; Brockman, F.J.; Zhang, W. Integrated analysis of transcriptomic and proteomic data of Desulfovibrio vulgaris: Zero-inflated Poisson regression models to predict abundance of undetected proteins. Bioinformatics 2006, 22, 1641–1647. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nie, L.; Wu, G.; Zhang, W. Correlation of mRNA Expression and Protein Abundance Affected by Multiple Sequence Features Related to Translational Efficiency in Desulfovibrio vulgaris: A Quantitative Analysis. Genetics 2006, 174, 2229–2243. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Mills, D.A. Improvement of a nisin-inducible expression vector for use in lactic acid bacteria. Plasmid 2007, 58, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Jürgen, B.; Hanschke, R.; Sarvas, M.; Hecker, M.; Schweder, T. Proteome and transcriptome based analysis of Bacillus subtilis cells overproducing an insoluble heterologous protein. Appl. Microbiol. Biotechnol. 2001, 55, 326–332. [Google Scholar] [CrossRef]
- Aldor, I.S.; Krawitz, D.C.; Forrest, W.; Chen, C.; Nishihara, J.C.; Joly, J.C.; Champion, K.M. Proteomic Profiling of Recombinant Escherichia coli in High-Cell-Density Fermentations for Improved Production of an Antibody Fragment Biopharmaceutical. Appl. Environ. Microbiol. 2005, 71, 1717–1728. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Lee, W.-C. Protein solubility and differential proteomic profiling of recombinant Escherichia coli overexpressing double-tagged fusion proteins. Microb. Cell Factories 2010, 9, 63. [Google Scholar] [CrossRef]
- Marciniak, B.C.; Trip, H.; Veek, P.J.V.-D.; Kuipers, O.P. Comparative transcriptional analysis of Bacillus subtilis cells overproducing either secreted proteins, lipoproteins or membrane proteins. Microb. Cell Factories 2012, 11, 66. [Google Scholar] [CrossRef]
- Morello, E.; Humaran, L.G.B.; Llull, D.; Solé-Jamault, V.; Miraglio, N.; Langella, P.; Poquet, I. Lactococcus lactis, an Efficient Cell Factory for Recombinant Protein Production and Secretion. J. Mol. Microbiol. Biotechnol. 2007, 14, 48–58. [Google Scholar] [CrossRef]
- Poquet, I.; Bolotin, A.; Gruss, A. Optimising the production of heterologous exported proteins in Lactococcus lactis by inactivation of HtrA, the unique house-keeping surface protease. Lait 2001, 81, 37–47. [Google Scholar] [CrossRef]
- Van der Meulen, S.B.; de Jong, A.; Kok, J. Early Transcriptome Response of Lactococcus lactis to Environmental Stresses Reveals Differentially Expressed Small Regulatory RNAs and tRNAs. Front. Microbiol. 2017, 8, 1704. [Google Scholar] [CrossRef]
- Marreddy, R.K.R.; Geertsma, E.R.; Permentier, H.P.; Pinto, J.P.C.; Kok, J.; Poolman, B. Amino Acid Accumulation Limits the Overexpression of Proteins in Lactococcus lactis. PLoS ONE 2010, 5, e10317. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, A.; McDermaid, A.; Zhang, H.; Liu, C.; Cao, H.; Ma, Q. RECTA: Regulon Identification Based on Comparative Genomics and Transcriptomics Analysis. Genes 2018, 9, 278. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, G.A.; Duarte, S.O.D. The Effect of Recombinant Protein Production in Lactococcus lactis Transcriptome and Proteome. Microorganisms 2022, 10, 267. https://doi.org/10.3390/microorganisms10020267
Monteiro GA, Duarte SOD. The Effect of Recombinant Protein Production in Lactococcus lactis Transcriptome and Proteome. Microorganisms. 2022; 10(2):267. https://doi.org/10.3390/microorganisms10020267
Chicago/Turabian StyleMonteiro, Gabriel A., and Sofia O. D. Duarte. 2022. "The Effect of Recombinant Protein Production in Lactococcus lactis Transcriptome and Proteome" Microorganisms 10, no. 2: 267. https://doi.org/10.3390/microorganisms10020267
APA StyleMonteiro, G. A., & Duarte, S. O. D. (2022). The Effect of Recombinant Protein Production in Lactococcus lactis Transcriptome and Proteome. Microorganisms, 10(2), 267. https://doi.org/10.3390/microorganisms10020267