
Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species




Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of Lacticaseibacillus spp. Genomes for Genes Involved in Survival in GIT and Adhesion
2.2. Bacterial Strains and Culture Conditions
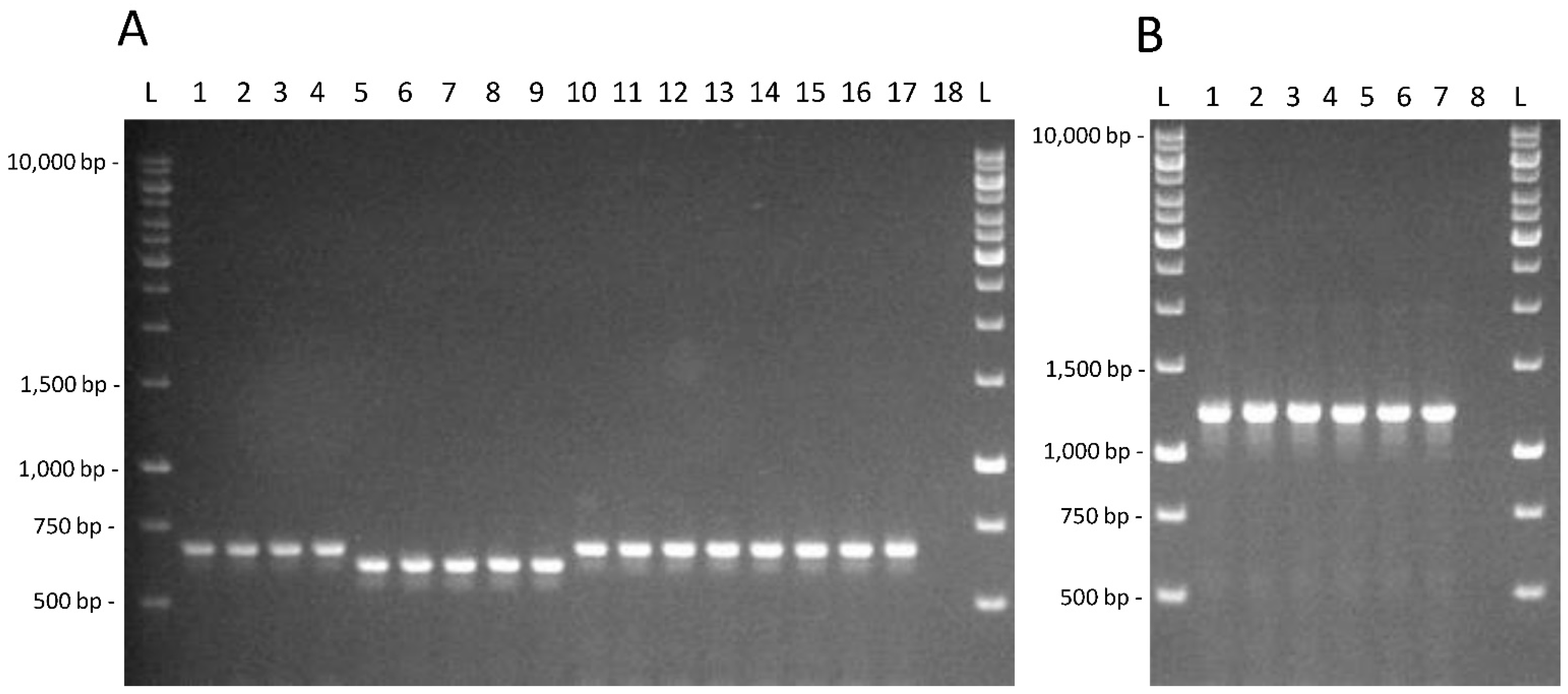
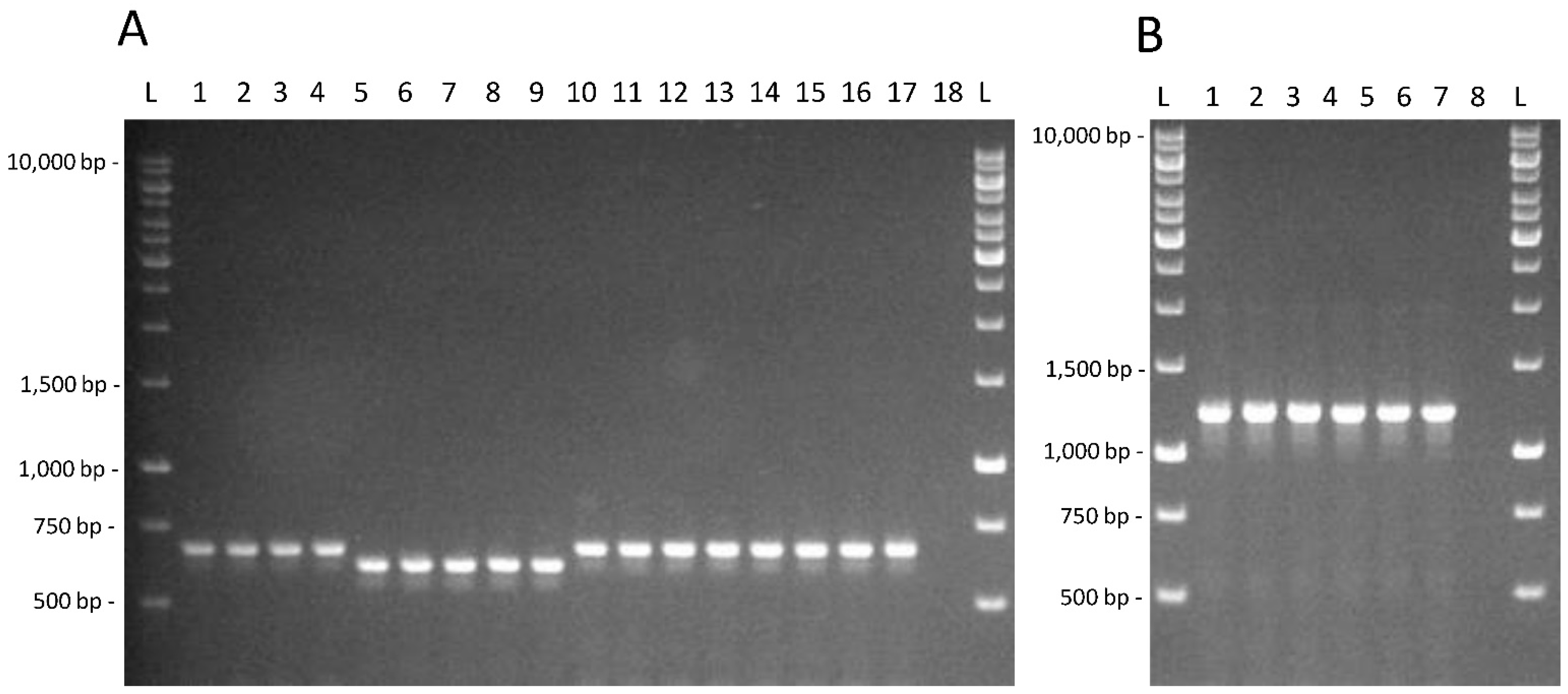
2.3. Molecular Techniques
2.4. Biofilm Forming Capacity
2.5. Bile Salt Tolerance Test
2.6. Statistical Analyses
3. Results
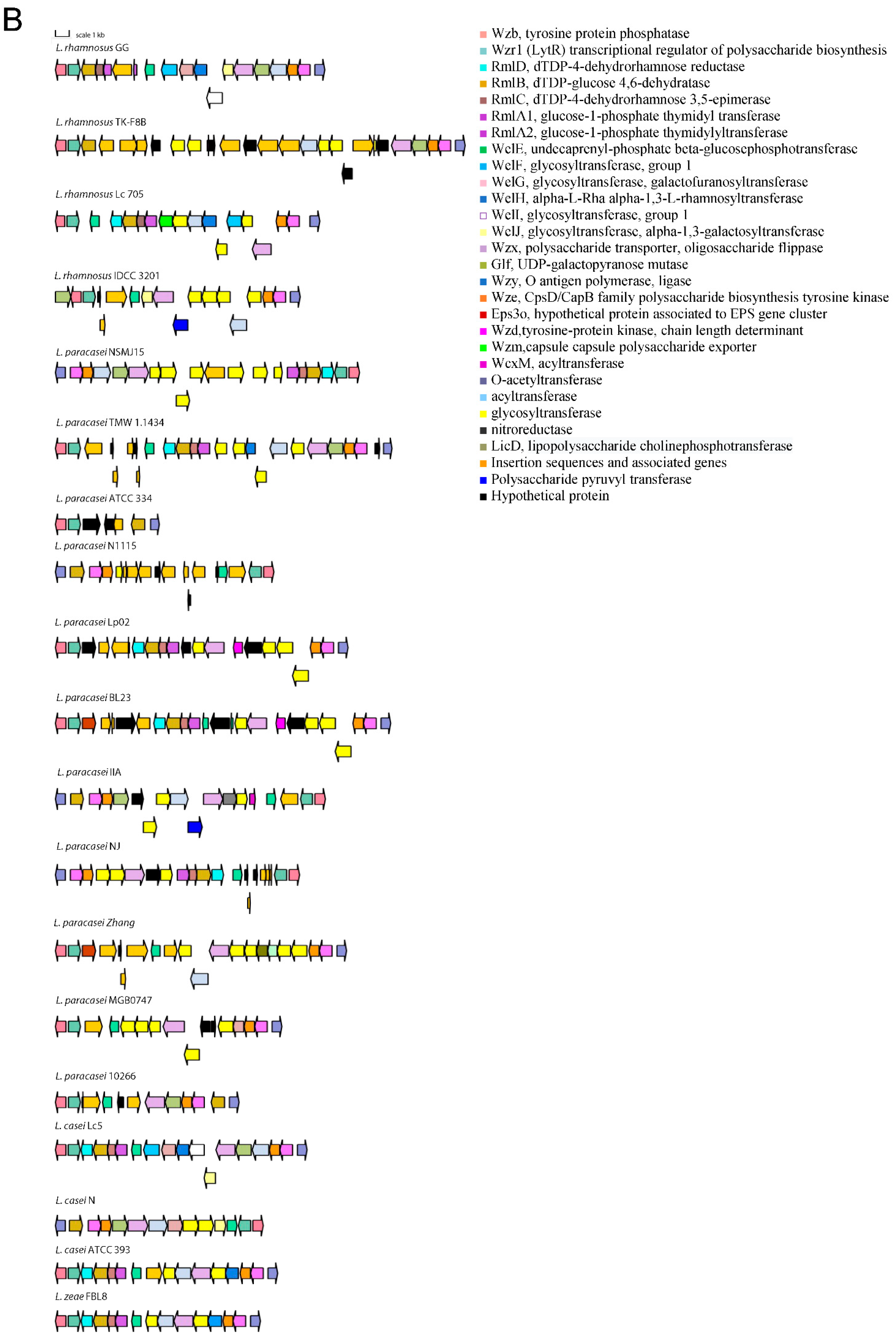
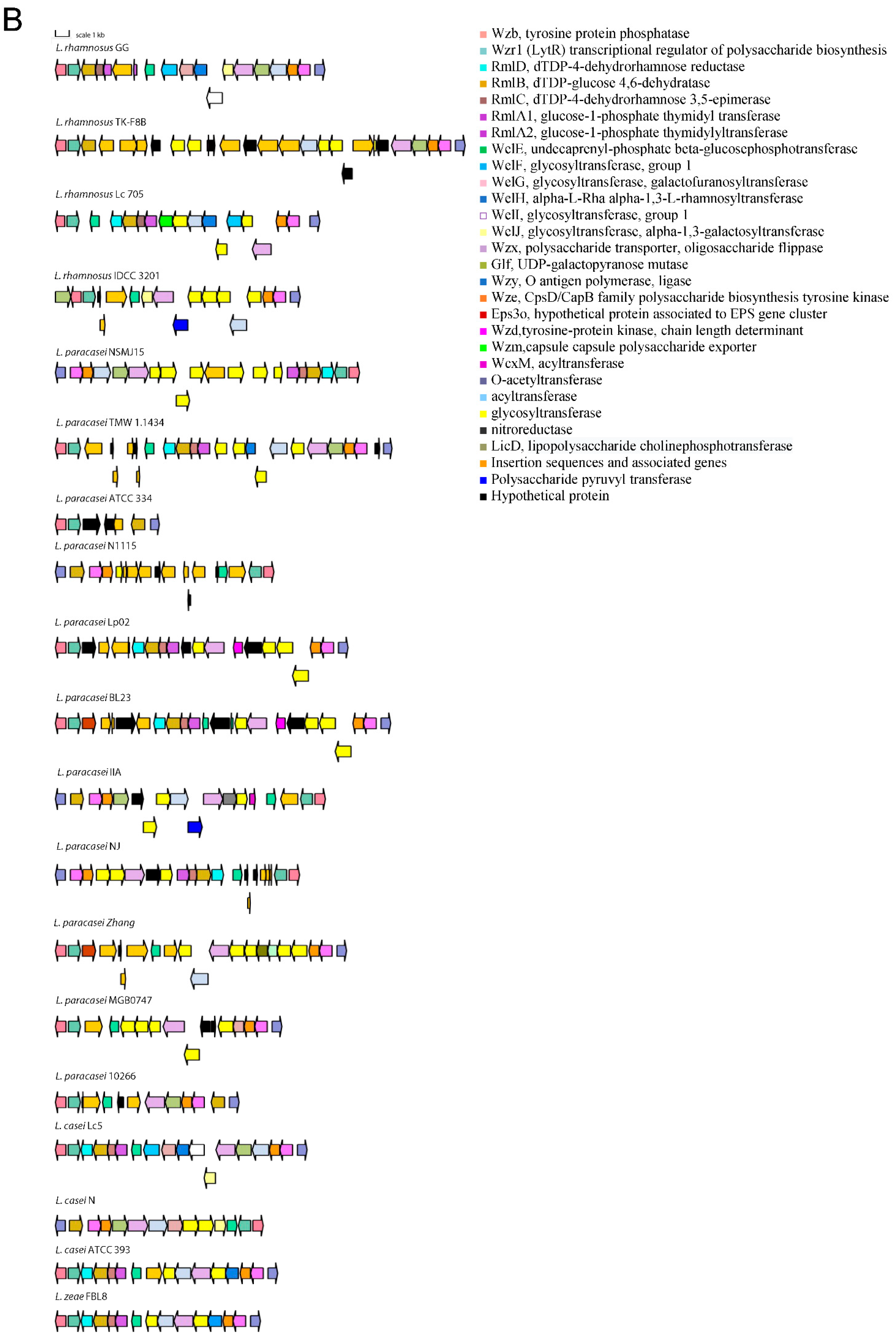
3.1. Distribution of Genetic Traits Required for Probiotic Activity in Lacticaseibacillus Species
3.2. Testing of Genetic and Physiological Features in Lacticaseibacillus Strains from Feces
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jacouton, E.; Chain, F.; Sokol, H.; Langella, P.; Bermúdez-Humarán, L.G. Probiotic Strain Lactobacillus casei BL23 Prevents Colitis-Associated Colorectal Cancer. Front. Immunol. 2017, 8, 1553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Cao, C.; Wu, Z.; Zhang, H.; Sun, Z.; Wang, M.; Xu, H.; Zhao, Z.; Wang, Y.; Pei, G.; et al. The probiotic L. casei Zhang slows the progression of acute and chronic kidney disease. Cell Metab. 2021, 33, 1926–1942.e8. [Google Scholar] [CrossRef] [PubMed]
- Colombo Pimentel, T.; Ramalho Brandão, L.; Pereira de Oliveira, M.; Almeida da Costa, W.K.; Magnani, M. Health benefits and technological effects of Lacticaseibacillus casei-01: An overview of the scientific literature. Trends Food Sci. Technol. 2021, 114, 722–737. [Google Scholar] [CrossRef]
- Capurso, L. Thirty Years of Lactobacillus rhamnosus GG: A Review. J. Clin. Gastroenterol. 2019, 53 (Suppl. 1), S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Del Matto, I.; Rossi, F.; Iannitto, G.; Petrone, D.; Mastrodomenico, M.T.; Alessiani, A.; Sacchini, L.; Amadoro, C.; Tucci, P.; Marino, L. Variability of the microbiota in traditional Caciocavallo, Scamorza and Caciotta cheeses manufactured with raw milk and natural cultures. Int. J. Dairy Technol. 2021, 74, 564–574. [Google Scholar] [CrossRef]
- Lu, Y.; Han, S.; Zhang, S.; Wang, K.; Lv, L.; McClements, D.J.; Xiao, H.; Berglund, B.; Yao, M.; Li, L. The role of probiotic exopolysaccharides in adhesion to mucin in different gastrointestinal conditions. Curr. Res. Food Sci. 2022, 5, 581–589. [Google Scholar] [CrossRef]
- Douillard, F.P.; Ribbera, A.; Kant, R.; Pietilä, T.E.; Järvinen, H.M.; Messing, M.; Randazzo, C.L.; Paulin, L.; Laine, P.; Ritari, J.; et al. Comparative genomic and functional analysis of 100 Lactobacillus rhamnosus strains and their comparison with strain GG. PLoS Gen. 2013, 9, e1003683. [Google Scholar] [CrossRef]
- Rodríguez-Díaz, J.; Monedero, V.; Yebra, M.J. Utilization of Natural Fucosylated Oligosaccharides by Three Novel α-l-Fucosidases from a Probiotic Lactobacillus casei Strain. Appl. Environ. Microbiol. 2011, 77, 703–705. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Yang, S.M.; Kim, D.; Kim, H.Y. Complete Genome Sequencing and Comparative Genomics of Three Potential Probiotic Strains, Lacticaseibacillus casei FBL6, Lacticaseibacillus chiayiensis FBL7, and Lacticaseibacillus zeae FBL8. Front. Microbiol. 2022, 12, 794315. [Google Scholar] [CrossRef]
- Amadoro, C.; Rossi, F.; Pallotta, M.L.; Gasperi, M.; Colavita, G. Traditional dairy products can supply beneficial microorganisms able to survive in the gastrointestinal tract. LWT 2018, 93, 376–383. [Google Scholar] [CrossRef]
- Bustos, A.Y.; Raya, R.; Bru, E.; de Valdez, G.F.; Taranto, M.P. Application of fluorescent techniques to evaluate the survival of probiotic lactobacilli to bile acid. Biotechnol. Lett. 2011, 33, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Frolova, M.; Yudin, S.; Makarov, V.; Glazunova, O.; Alikina, O.; Markelova, N.; Kolzhetsov, N.; Dzhelyadin, T.; Shcherbakova, V.; Trubitsyn, V.; et al. Lacticaseibacillus paracasei: Occurrence in the Human Gut Microbiota and K-Mer-Based Assessment of Intraspecies Diversity. Life 2021, 11, 1246. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Hill, C.; Gahan, C.G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Bäuerl, C.; Pérez-Martínez, G.; Yan, F.; Polk, D.B.; Monedero, V. Functional analysis of the p40 and p75 proteins from Lactobacillus casei BL23. J. Mol. Microbiol. Biotechnol. 2010, 19, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Domínguez Rubio, A.P.; Martínez, J.H.; Martínez Casillas, D.C.; Coluccio Leskow, F.; Piuri, M.; Pérez, O.E. Lactobacillus casei BL23 Produces Microvesicles Carrying Proteins That Have Been Associated with Its Probiotic Effect. Front. Microbiol. 2017, 8, 1783. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Scherbak, N.; Khalaf, H.; Olsson, P.E.; Jass, J. Substances released from probiotic Lactobacillus rhamnosus GR-1 potentiate NF-κB activity in Escherichia coli-stimulated urinary bladder cells. FEMS Immunol. Med. Microbiol. 2012, 66, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Chen, H.; Song, Y.; Chen, Y.Q.; Zhang, H.; Chen, W. Myosin-cross-reactive antigens from four different lactic acid bacteria are fatty acid hydratases. Biotechnol. Lett. 2013, 35, 75–81. [Google Scholar] [CrossRef]
- Jung, C.J.; Hsu, C.C.; Chen, J.W.; Cheng, H.W.; Yuan, C.T.; Kuo, Y.M.; Hsu, R.B.; Chia, J.S. PspC domain-containing protein (PCP) determines Streptococcus mutans biofilm formation through bacterial extracellular DNA release and platelet adhesion in experimental endocarditis. PLoS Pathog. 2021, 17, e1009289. [Google Scholar] [CrossRef]
- Naikare, H.; Palyada, K.; Panciera, R.; Marlow, D.; Stintzi, A. Major role for FeoB in Campylobacter jejuni ferrous iron acquisition, gut colonization, and intracellular survival. Infect. Immun. 2006, 74, 5433–5444. [Google Scholar] [CrossRef] [Green Version]
- Vélez, M.P.; Petrova, M.I.; Lebeer, S.; Verhoeven, T.L.; Claes, I.; Lambrichts, I.; Tynkkynen, S.; Vanderleyden, J.; De Keersmaecker, S.C. Characterization of MabA, a modulator of Lactobacillus rhamnosus GG adhesion and biofilm formation. FEMS Immunol. Med. Microbiol. 2010, 59, 386–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hörmannsperger, G.; von Schillde, M.A.; Haller, D. Lactocepin as a protective microbial structure in the context of IBD. Gut Microbes 2013, 4, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinster, S.; Furlan, S.; Serror, P. C-terminal WxL domain mediates cell wall binding in Enterococcus faecalis and other gram-positive bacteria. J. Bacteriol. 2007, 189, 1244–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, M.I.; Lievens, E.; Verhoeven, T.L.; Macklaim, J.M.; Gloor, G.; Schols, D.; Vanderleyden, J.; Reid, G.; Lebeer, S. The lectin-like protein 1 in Lactobacillus rhamnosus GR-1 mediates tissue-specific adherence to vaginal epithelium and inhibits urogenital pathogens. Sci. Rep. 2016, 6, 37437. [Google Scholar] [CrossRef]
- Gagic, D.; Wen, W.; Collett, M.A.; Rakonjac, J. Unique secreted-surface protein complex of Lactobacillus rhamnosus, identified by phage display. Microbiology 2013, 2, 1–17. [Google Scholar] [CrossRef]
- Chaurasia, P.; Pratap, S.; von Ossowski, I.; Palva, A.; Krishnan, V. New insights about pilus formation in gut-adapted Lactobacillus rhamnosus GG from the crystal structure of the SpaA backbone-pilin subunit. Sci. Rep. 2016, 6, 28664. [Google Scholar] [CrossRef] [Green Version]
- von Ossowski, I.; Reunanen, J.; Satokari, R.; Vesterlund, S.; Kankainen, M.; Huhtinen, H.; Tynkkynen, S.; Salminen, S.; de Vos, W.M.; Palva, A. Mucosal adhesion properties of the probiotic Lactobacillus rhamnosus GG SpaCBA and SpaFED pilin subunits. Appl. Environ. Microbiol. 2010, 76, 2049–2057. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Tomaro-Duchesneau, C.; Martoni, C.J.; Prakash, S. Cholesterol lowering with bile salt hydrolase-active probiotic bacteria, mechanism of action, clinical evidence, and future direction for heart health applications. Expert Opin. Biol. Ther. 2013, 13, 631–642. [Google Scholar] [CrossRef]
- Munoz-Provencio, D.; Perez-Martinez, G.; Monedero, V. Characterization of a fibronectin-binding protein from Lactobacillus casei BL23. J. Appl. Microbiol. 2010, 108, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Hymes, J.P.; Klaenhammer, T.R. Stuck in the Middle: Fibronectin-Binding Proteins in Gram-Positive Bacteria. Front. Microbiol. 2016, 7, 1504. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Washington, M.K.; Shi, Y.; Liu, L.; Chaturvedi, R.; Peek, R.M., Jr.; Wilson, K.T.; Polk, D.B. Colon-specific delivery of a probiotic-derived soluble protein ameliorates intestinal inflammation in mice through an EGFR-dependent mechanism. J. Clin. Investig. 2011, 121, 2242–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, L.; Moore, D.J.; Shen, X.; Peek, R.M.; Acra, S.A.; Li, H.; Ren, X.; Polk, D.B.; Yan, F. An LGG-derived protein promotes IgA production through upregulation of APRIL expression in intestinal epithelial cells. Mucosal. Immunol. 2017, 10, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Hao, Y.; Wang, L.; Lu, C.; Li, M.; Si, Z.; Wu, X.; Lu, Z. Characterization of a bacterial strain Lactobacillus paracasei LP10266 recovered from an endocarditis patient in Shandong, China. BMC Microbiol. 2021, 21, 183. [Google Scholar] [CrossRef] [PubMed]
- Koryszewska-Bagińska, A.; Gawor, J.; Nowak, A.; Grynberg, M.; Aleksandrzak-Piekarczyk, T. Comparative genomics and functional analysis of a highly adhesive dairy Lactobacillus paracasei subsp. paracasei IBB3423 strain. Appl. Microbiol. Biotechnol. 2019, 103, 7617–7634. [Google Scholar] [CrossRef] [Green Version]
- Fontana, A.; Zacconi, C.; Morelli, L. Genetic Signatures of Dairy Lactobacillus casei Group. Front. Microbiol. 2018, 9, 2611. [Google Scholar] [CrossRef]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Balzaretti, S.; Taverniti, V.; Guglielmetti, S.; Fiore, W.; Minuzzo, M.; Ngo, H.N.; Ngere, J.B.; Sadiq, S.; Humphreys, P.N.; Laws, A.P. A Novel Rhamnose-Rich Hetero-exopolysaccharide Isolated from Lactobacillus paracasei DG Activates THP-1 Human Monocytic Cells. Appl. Environ. Microbiol. 2017, 83, e02702-16. [Google Scholar] [CrossRef] [Green Version]
- Siezen, R.; Boekhorst, J.; Muscariello, L.; Molenaar, D.; Renckens, B.; Kleerebezem, M. Lactobacillus plantarum gene clusters encoding putative cell-surface protein complexes for carbohydrate utilization are conserved in specific gram-positive bacteria. BMC Genom. 2006, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. EFSA statement on the requirements for whole genome sequence analysis of microorganisms intentionally used in the food chain. EFSA J. 2021, 19, 6506. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Genetic Loci in L. rhamnosus GG |
|---|---|
| Anti-inflammatory protein | LGG_02734 b |
| Bile salt hydrolase | LGG_00501 * |
| Biofilm formation | LGG_00914; LGG_01827 |
| Cell wall anchored proteins with LPXTG domain | LGG_00434 b,f; LGG_00578 c,d,e; LGG_00584 |
| Fibrinogen binding | LGG_01590 c,d,e,f; LGG_02282 b |
| Fibronectin binding | LGG_0005 a,b; LGG_01450 |
| Fucose utilization | LGG_02652 a |
| Gut colonization | LGG_01877 g |
| Lectin-like protein | LGG_00183; LGG_00576 a; LGG_00579 b,c,d; LGG_00583 c,d,e,f; LGG_00585 c,d,e,f; LGG_00587 c,d,e,f; LGG_01765 c,d |
| Mucus binding | LGG_01883; LGG_02337e |
| Pilus protein | LGG_00422 c,d,f; LGG_00442 c,d,f; LGG_00443 c,d,f; LGG_00444 c,d,f; LGG_02370 c,d; LGG_02371 c,d; LGG_02372 c,d |
| Polysaccharide biosynthesis | LGG_00107 a,b,c,d; LGG_00278 a,c,d; LGG_00279 a,c,d; LGG_00280 a,c,d; LGG_00281 a,c,d; LGG_00282 a,c,d; LGG_00283 a,c,d; LGG_00295; LGG_00348 c,d,e; LGG_00349 e; LGG_00645; LGG_00695 e; LGG_00696; LGG_00697 e; LGG_00825; LGG_00826; LGG_00827; LGG_00830; LGG_00851; LGG_00998; LGG_00999; LGG_01057; LGG_01066; LGG_01069; LGG_01147; LGG_01366; LGG_01538 a,d; LGG_01586 e; LGG_01587 c,d,e; LGG_01990 †; LGG_02036 †; LGG_02144; LGG_02520; LGG_02869 h |
| Sortase | LGG_00441 c,f; LGG_02369 c,d |
| Surface adhesin | LGG_01591 e,f; LGG_01592 c,d,e,f; LGG_01865 b; LGG_02423; LGG_02426; LGG_02923 c,d,f |
| Surface antigen | LGG_00031; LGG_00324; LGG_00500 *; LGG_00503; LGG_01589 a,c,d; LGG_02016 |
| Taurine utilization | LGG_00172 a,c,d; LGG_00173 a,c,d; LGG_00174 a,c,d; LGG_00544 b |
| Toxin immunity | LGG_01002 |
| Strain | Taurine Utilization | α-L-Fucosidase LGG_02652 | % Membrane Potential Dissipation | Biofilm Formation (OD 620 nm) |
|---|---|---|---|---|
| L. casei | ||||
| AB-15-6 | + | 77 ± 0.5 a | 0.028 ± 0.01 a | |
| C-15-1 | + | 75 ± 0.5 a | 0.034 ± 0.01 a | |
| C-15-1b | + | + | 83.5 ± 0.5 b | 0.160 ± 0.05 b |
| G-0-6 | + | 86 ± 1 c | 0.065 ± 0.02 c | |
| L. paracasei | ||||
| AN-15-1 | 87 ± 0.5 c | 0.436 ± 0.05 d | ||
| AN-15-2 | + | 77 ± 1 a | 0.122 ± 0.05 e | |
| AN-15-3 | + | 80 ± 0.5 d | 0.051 ± 0.02 f | |
| AN-15-4 | 87 ± 0.5 c | 0.068 ± 0.01 c | ||
| J-7-1 | + | 83 ± 1 b | 0.115 ± 0.03 e | |
| J-7-3 | 79 ± 0.5 d | 0.534 ± 0.09 d | ||
| J-15-4 | + | 75 ± 1 a | 0.034 ± 0.02 a | |
| P-7-13 ¤ | + | + | 77 ± 0.5 a | 0.133 ± 0.02 b |
| 6-15-1 | 71.5 ± 0.5 | 0.043 ± 0.01 f | ||
| L. rhamnosus | ||||
| AN-0-1 | 83 ± 0.5 b | 0.171 ± 0.04 b | ||
| AN-7-1 | 76 ± 1 a | 0.111 ± 0.05 e | ||
| AN-7-4 | + | 87 ± 0.5 c | 0.119 ± 0.03 e | |
| AN-21-1 | + | 82.5 ± 0.5 b | 0.032 ± 0.01 a | |
| AN-21-2 | + | 81 ± 1 d | 0.076 ± 0.02 c | |
| C-0-4 | 75 ± 1 a | 0.025 ± 0.01 a | ||
| D-0-5 | + | + | 77 ± 0.5 a | 0.048 ± 0.02 f |
| G-7-14 | + | 80 ± 0.5 d | 0.270 ± 0.05 | |
| G-7-16 | + | 83 ± 0.5 b | 0.180 ± 0.02 b | |
| J-7-4 | + | 82 ± 0.5 b | 0.023 ± 0.01 a | |
| SA-7-6 | 83 ± 0.5 b | 0.613 ± 0.07 | ||
| Z-15-4 | + | + | 73 ± 0.5 | 0.024 ± 0.01 a |
| L. zeae | ||||
| J-7-2 | 95 ± 1 | 0.084 ± 0.02 c | ||
| L. rhamnosus GG | + | + | 79 ± 0.5 d | 0.138 ± 0.03 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, F.; Amadoro, C.; Pallotta, M.L.; Colavita, G. Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species. Microorganisms 2022, 10, 1023. https://doi.org/10.3390/microorganisms10051023
Rossi F, Amadoro C, Pallotta ML, Colavita G. Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species. Microorganisms. 2022; 10(5):1023. https://doi.org/10.3390/microorganisms10051023
Chicago/Turabian StyleRossi, Franca, Carmela Amadoro, Maria Luigia Pallotta, and Giampaolo Colavita. 2022. "Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species" Microorganisms 10, no. 5: 1023. https://doi.org/10.3390/microorganisms10051023
APA StyleRossi, F., Amadoro, C., Pallotta, M. L., & Colavita, G. (2022). Variability of Genetic Characters Associated with Probiotic Functions in Lacticaseibacillus Species. Microorganisms, 10(5), 1023. https://doi.org/10.3390/microorganisms10051023