Effects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. bovis

 , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Animals
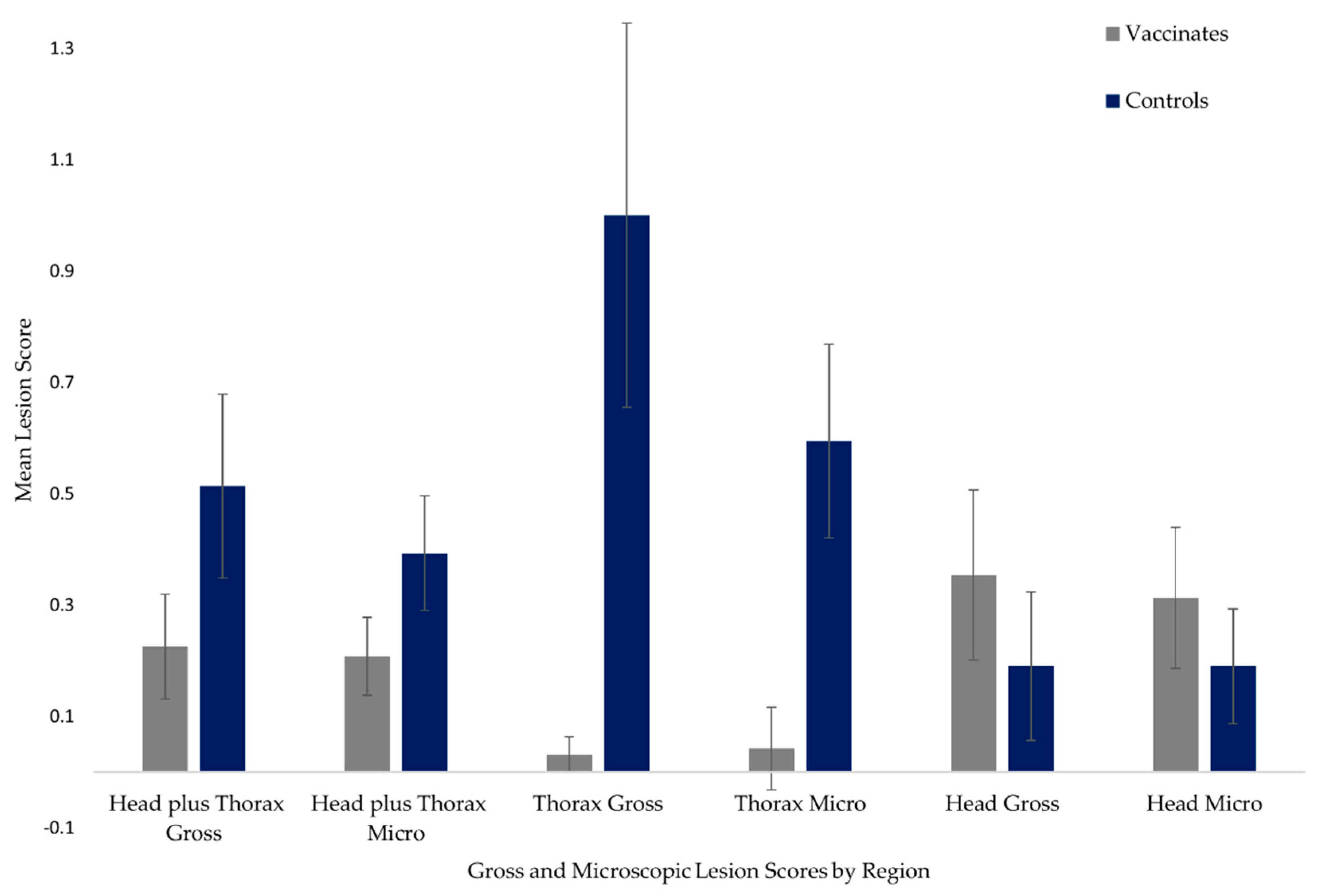
2.2. Gross and Microscopic Lesion Scores
2.3. Culture
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Vaccination
4.3. Challenge
4.4. Necropsy
4.5. Culture
4.6. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Palmer, M.V.; Waters, W.R. Bovine Tuberculosis and the Establishment of an Eradication Program in the United States: Role of Veterinarians. Veter. Med. Int. 2011, 2011, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Portacci, K.; Lombard, J.; Schoenbaum, M.; Orloski, K.; Camacho, M. The occurrence of M. bovis cases in U.S. cattle, 2001–2011. In Zoonotic tuberculosis: Mycobacterium Bovis and Other Pathogenic Mycobacteria; Thoen, C., Steele, J., Kaneene, J., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2014; pp. 253–261. [Google Scholar]
- Schmitt, S.M.; Fitzgerald, S.D.; Cooley, T.M.; Bruning-Fann, C.S.; Sullivan, L.; Berry, D.; Carlson, T.; Minnis, R.B.; Payeur, J.B.; Sikarskie, J. Bovine tuberculosis in free-ranging white-tailed deer from michigan. J. Wildl. Dis. 1997, 33, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Freier, J.E.; Miller, R.; Geter, K.D. Geospatial analysis and modelling in the prevention and control of animal diseases in the United States. Veter. Ital. 2010, 43. [Google Scholar]
- Rossi, S.; Staubach, C.; Blome, S.; Guberti, V.; Thulke, H.-H.; Vos, A.; Koenen, F.; Le Potier, M.-F. Controlling of CSFV in European wild boar using oral vaccination: A review. Front. Microbiol. 2015, 6, 274. [Google Scholar] [CrossRef]
- Freuling, C.M.; Hampson, K.; Selhorst, T.; Schröder, R.; Meslin, F.X.; Mettenleiter, T.C.; Müller, T. The elimination of fox rabies from Europe: Determinants of success and lessons for the future. Philos. Trans. R. Soc. B Boil. Sci. 2013, 368, 20120142. [Google Scholar] [CrossRef]
- Slate, D.; Algeo, T.P.; Nelson, K.M.; Chipman, R.B.; Donovan, D.; Blanton, J.D.; Niezgoda, M.; Rupprecht, C.E. Oral Rabies Vaccination in North America: Opportunities, Complexities, and Challenges. PLoS Neglected Trop. Dis. 2009, 3, e549. [Google Scholar] [CrossRef]
- Díez-Delgado, I.; Sevilla, I.A.; Romero, B.; Tanner, E.; Barasona, J.A.; White, A.R.; Lurz, P.W.; Boots, M.; De La Fuente, J.; Dominguez, L.; et al. Impact of piglet oral vaccination against tuberculosis in endemic free-ranging wild boar populations. Prev. Veter. Med. 2018, 155, 11–20. [Google Scholar] [CrossRef]
- Roy, Á.; Risalde, M.A.; Bezos, J.; Casal, C.; Romero, B.; Sevilla, I.A.; Díez-Guerrier, A.; Rodríguez-Bertos, A.; Domínguez, M.; Garrido, J.; et al. Response of goats to intramuscular vaccination with heat-killed Mycobacterium bovis and natural challenge. Comp. Immunol. Microbiol. Infect. Dis. 2018, 60, 28–34. [Google Scholar] [CrossRef]
- Arrieta-Villegas, C.; Perálvarez, T.; Vidal, E.; Puighibet, Z.; Moll, X.; Canturri, A.; Sevilla, I.A.; Espada, Y.; Juste, R.; Domingo, M.; et al. Efficacy of parenteral vaccination against tuberculosis with heat-inactivated Mycobacterium bovis in experimentally challenged goats. PLoS ONE 2018, 13, e0196948. [Google Scholar] [CrossRef]
- Thomas, J.; Risalde, M.Á.; Serrano, M.; Sevilla, I.A.; Geijo, M.; Ortíz, J.A.; Fuertes, M.; Ruíz-Fons, J.F.; De La Fuente, J.; Domínguez, L.; et al. The response of red deer to oral administration of heat-inactivated Mycobacterium bovis and challenge with a field strain. Veter. Microbiol. 2017, 208, 195–202. [Google Scholar] [CrossRef]
- Van Der Heijden, E.M.; Chileshe, J.; Vernooij, H.; Gortázar, C.; Juste, R.A.; Sevilla, I.A.; Crafford, J.E.; Rutten, V.P.M.G.; Michel, A.L. Immune response profiles of calves following vaccination with live BCG and inactivated Mycobacterium bovis vaccine candidates. PLoS ONE 2017, 12, e0188448. [Google Scholar] [CrossRef] [PubMed]
- Díez-Delgado, I.; Rodríguez, O.; Boadella, M.; Garrido, J.; Sevilla, I.A.; Bezos, J.; Juste, R.; Domínguez, L.; Gortazar, C. Parenteral Vaccination with Heat-Inactivated Mycobacterium Bovis Reduces the Prevalence of Tuberculosis-Compatible Lesions in Farmed Wild Boar. Transbound. Emerg. Dis. 2016, 64, e18–e21. [Google Scholar] [CrossRef] [PubMed]
- Beltran-Beck, B.; De La Fuente, J.; Garrido, J.M.; Aranaz, A.; Sevilla, I.A.; Villar, M.; Boadella, M.; Galindo, R.C.; De La Lastra, J.M.P.; Moreno-Cid, J.A.; et al. Oral Vaccination with Heat Inactivated Mycobacterium bovis Activates the Complement System to Protect against Tuberculosis. PLoS ONE 2014, 9, e98048. [Google Scholar] [CrossRef] [PubMed]
- Gortazar, C.; Beltran-Beck, B.; Garrido, J.M.; Aranaz, A.; Sevilla, I.A.; Boadella, M.; Lyashchenko, K.P.; Galindo, R.C.; Montoro, V.; Domínguez, L.; et al. Oral re-vaccination of Eurasian wild boar with Mycobacterium bovis BCG yields a strong protective response against challenge with a field strain. BMC Veter. Res. 2014, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, V.; Gortazar, C.; Vicente, J.; De La Fuente, J. Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex. Veter. Microbiol. 2008, 127, 1–9. [Google Scholar] [CrossRef]
- Rossi, S.; Toïgo, C.; Hars, J.; Pol, F.; Hamann, J.-L.; Depner, K.; Le Potier, M.-F. New Insights on the Management of Wildlife Diseases Using Multi-State Recapture Models: The Case of Classical Swine Fever in Wild Boar. PLoS ONE 2011, 6, e24257. [Google Scholar] [CrossRef]
- Barasona, J.A.; Díez-Delgado, I.; Aznar, J.; Vicente, J.; Gortazar, C.; Torres, M.J. Environmental Presence of Mycobacterium tuberculosis Complex in Aggregation Points at the Wildlife/Livestock Interface. Transbound. Emerg. Dis. 2016, 64, 1148–1158. [Google Scholar] [CrossRef]
- Fischer, J.W.; Blass, C.R.; Walter, W.D.; Anderson, C.W.; Lavelle, M.J.; Hall, W.H.; Vercauteren, K.C. Evaluating a strategy to deliver vaccine to white-tailed deer at a landscape level. Wildl. Soc. Bull. 2016, 40, 394–399. [Google Scholar] [CrossRef]
- Garrido, J.M.; Sevilla, I.A.; Beltran-Beck, B.; Minguijón, E.; Ballesteros, C.; Galindo, R.C.; Boadella, M.; Lyashchenko, K.P.; Romero, B.; Geijo, M.V.; et al. Protection against Tuberculosis in Eurasian Wild Boar Vaccinated with Heat-Inactivated Mycobacterium bovis. PLoS ONE 2011, 6, e24905. [Google Scholar] [CrossRef]
- Ellis, C.K.; Wehtje, M.E.; Wolfe, L.L.; Wolff, P.L.; Hilton, C.D.; Fisher, M.C.; Green, S.; Glow, M.P.; Halseth, J.M.; Lavelle, M.J.; et al. Comparison of the efficacy of four drug combinations for immobilization of wild pigs. Eur. J. Wildl. Res. 2019, 65, 78. [Google Scholar] [CrossRef]
- Ballesteros, C.; Garrido, J.; Vicente, J.; Romero, B.; Galindo, R.; Minguijón, E.; Villar, M.; Martín-Hernando, M.; Sevilla, I.A.; Juste, R.; et al. First data on Eurasian wild boar response to oral immunization with BCG and challenge with a Mycobacterium bovis field strain. Vaccine 2009, 27, 6662–6668. [Google Scholar] [CrossRef] [PubMed]
- Robbe-Austerman, S.; Bravo, D.; Harris, B. Comparison of the MGIT 960, BACTEC 460 TB and solid media for isolation of Mycobacterium bovis in United States veterinary specimens. BMC Veter. Res. 2013, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Dykema, P.E.; Stokes, K.D.; Beckwith, N.R.; Mungin, J.W.; Xu, L.; Vickers, D.J.; Reising, M.M.; Bravo, D.M.; Thomsen, B.V.; Robbe-Austerman, S. Development and validation of a direct real-time PCR assay for Mycobacterium bovis and implementation into the United States national surveillance program. PeerJ PrePrints 2016. [Google Scholar] [CrossRef]
- Mu, M.; Mauthe, A.; Tyson, G.; Cerqueira, E. Statistical analysis of ordinal user opinion scores. In Proceedings of the 2012 IEEE Consumer Communications and Networking Conference (CCNC), Las Vegas, NV, USA, 14–17 January 2012; pp. 331–336. [Google Scholar]
- Agresti, A. Analysis of Ordinal Categorical Data; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar]
- Fritz, C.O.; Morris, P.E.; Richler, J. Effect size estimates: Current use, calculations, and interpretation. J. Exp. Psychol. Gen. 2012, 141, 2–18. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Informa UK Limited: London, UK, 2013. [Google Scholar]


{kind=link}
| Tissue (Gross Lesion Scores/Microscopic Lesion Score/M. bovis Culture Outcome: +/-) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lung Lobe | Organ | Lymph Node/Lymphoid Tissue | |||||||||||||||||
| Treatment Group | Total Gross Lesion Score | Total Microscopic Lesion Score | ID | Caud2,3 | Cran2,4 | Card2,5 | Acc6 | Liver | Spleen | Kidney | Mand2,7 | Parotid2 | Retro-pharyngeal2 | Superf Cerv2,8 | Medias9 | Tracheo-bronch2,10 | Hep11 | Mes12 | Tonsil |
| Vaccinates | 18 | 21 | C | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 1,3/2,3/+ | 0/0/- | 1,4/1,3/+ | 0/0/- | 0/0/- | 0/0/+ | 0/0/+ | 0/0/- | 0/0/- |
| D | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 4/3/+ | 0/0/+ | 0/0/- | 0/0/- | 0/2/+ | 0/0/+ | 0/0/- | 0/0/- | |||
| E | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 1/3/- | 0/1/+ | 0/0/- | 0/0/- | |||
| H | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| K | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| L | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| M | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 4/3/+ | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| N | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| Controls | 36 | 33 | A | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- |
| B | 0/1/+13 | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 4/3,1/+ | 0/0/- | 0/1/- | 0/0/- | 0/0/- | 0/2/- | 0/0/- | 0/0/- | 0/0/- | |||
| F | 0/0/- | 0/0/- | 5/3,3/+ | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/+ | 4,3/3,3/+ | 0/0/- | 0/0/- | 0/0/- | |||
| G | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
| I | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 4/3/+ | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/2/- | 0/0/- | 0/0/- | 0/0/- | |||
| J | 0/0/- | 0/0/- | 5,5/2,3/+ | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 3,3/3/+ | 0/0/- | 0/0/- | 0/0/- | |||
| O | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | 0/0/- | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nol, P.; Wehtje, M.E.; Bowen, R.A.; Robbe-Austerman, S.; Thacker, T.C.; Lantz, K.; Rhyan, J.C.; Baeten, L.A.; Juste, R.A.; Sevilla, I.A.; et al. Effects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. bovis. Pathogens 2020, 9, 199. https://doi.org/10.3390/pathogens9030199
Nol P, Wehtje ME, Bowen RA, Robbe-Austerman S, Thacker TC, Lantz K, Rhyan JC, Baeten LA, Juste RA, Sevilla IA, et al. Effects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. bovis. Pathogens. 2020; 9(3):199. https://doi.org/10.3390/pathogens9030199
Chicago/Turabian StyleNol, Pauline, Morgan E. Wehtje, Richard A. Bowen, Suelee Robbe-Austerman, Tyler C. Thacker, Kristina Lantz, Jack C. Rhyan, Laurie A. Baeten, Ramón A. Juste, Iker A. Sevilla, and et al. 2020. "Effects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. bovis" Pathogens 9, no. 3: 199. https://doi.org/10.3390/pathogens9030199
APA StyleNol, P., Wehtje, M. E., Bowen, R. A., Robbe-Austerman, S., Thacker, T. C., Lantz, K., Rhyan, J. C., Baeten, L. A., Juste, R. A., Sevilla, I. A., Gortázar, C., & Vicente, J. (2020). Effects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. bovis. Pathogens, 9(3), 199. https://doi.org/10.3390/pathogens9030199