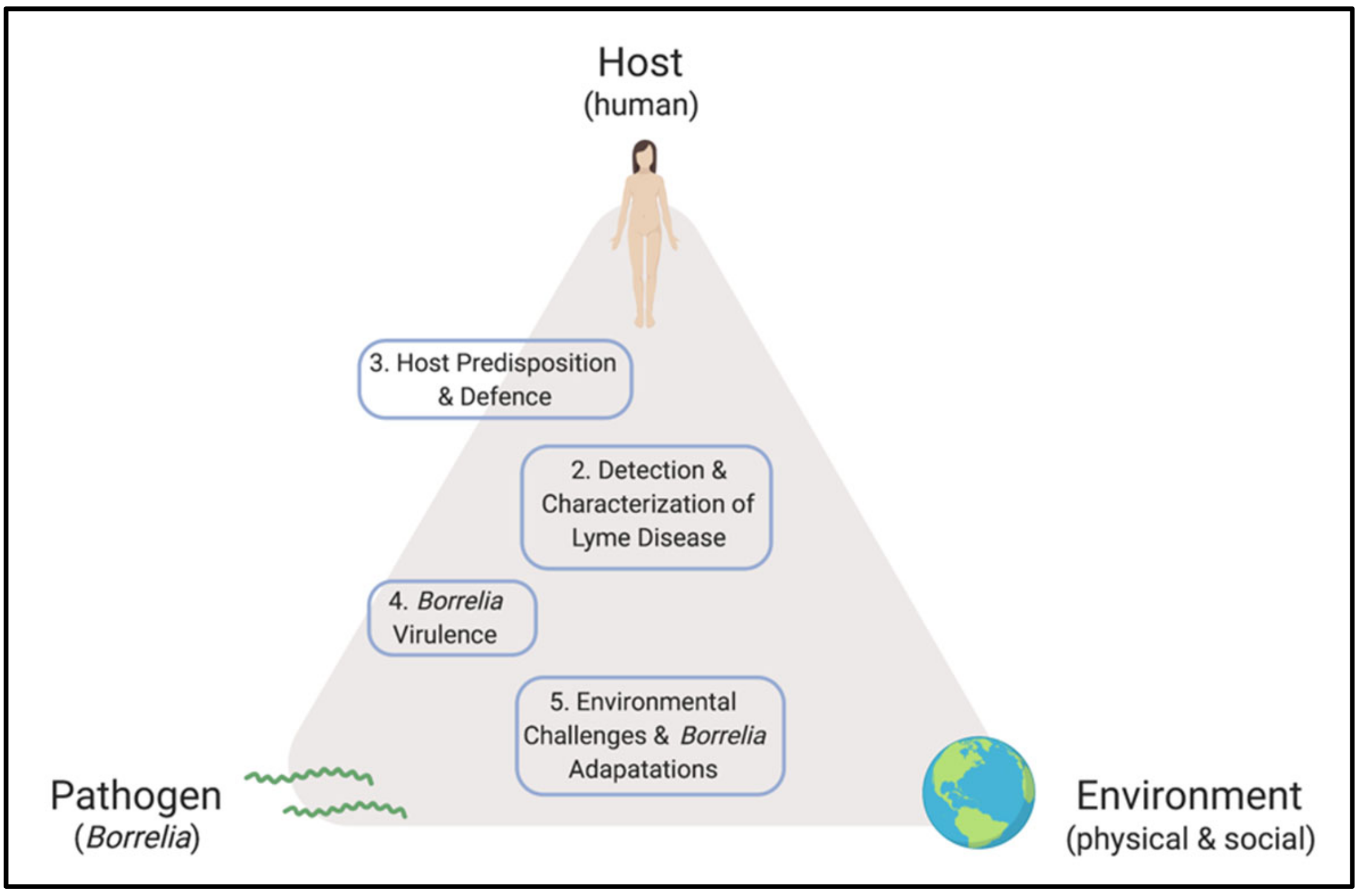
Lyme Disease Frontiers: Reconciling Borrelia Biology and Clinical Conundrums

 , and
, and
Abstract
1. Introduction
2. Detecting and Characterizing Lyme Disease
2.1. Diagnostic Challenges
2.2. Classifying Lyme Disease
Protracted Lyme Disease: Defining Chronic and Post-Treatment Conditions
2.3. Implications for Research
3. Pathogenicity: Host Predisposition and Defense
3.1. Genetic Susceptibility to Disease and Autoimmunity
3.2. Influence of Host Diet and Lifestyle: Hypercholesterolemia and Eicosanoids
4. Pathogenicity: Borrelia Virulence
4.1. Borrelia Biodiversity and Disease
4.1.1. Genospecies, Geography, and Disease Manifestation
4.1.2. Serotypes and Invasion
4.2. Host Colonization and Survival Strategies
4.2.1. Immune Modulation
4.2.2. Host Evasion
Host-Borrelia Interactions and Internalization
5. Environmental Challenges and Microbial Adaptations
5.1. Antibiotics and Borrelia Burgdorferi
5.1.1. Laboratory Determination of Antibiotic Susceptibilities
5.1.2. Resistance: Known Mechanisms and Generation
5.1.3. Borrelial Antibiotic Persistence
5.1.4. Clinical Antibiotic Treatment
5.1.5. Alternative Anti-Microbial Treatments
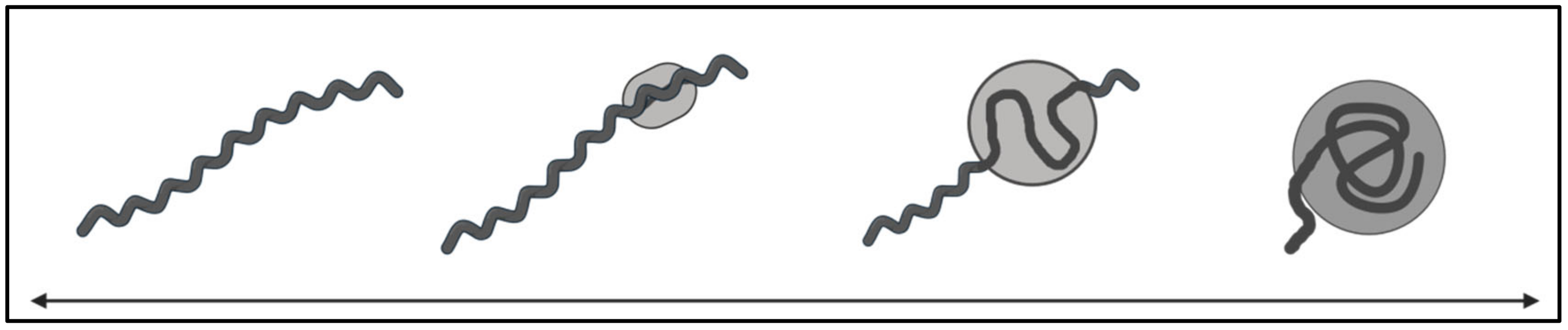
5.2. Pleomorphy
6. Conclusions and Future Frontiers
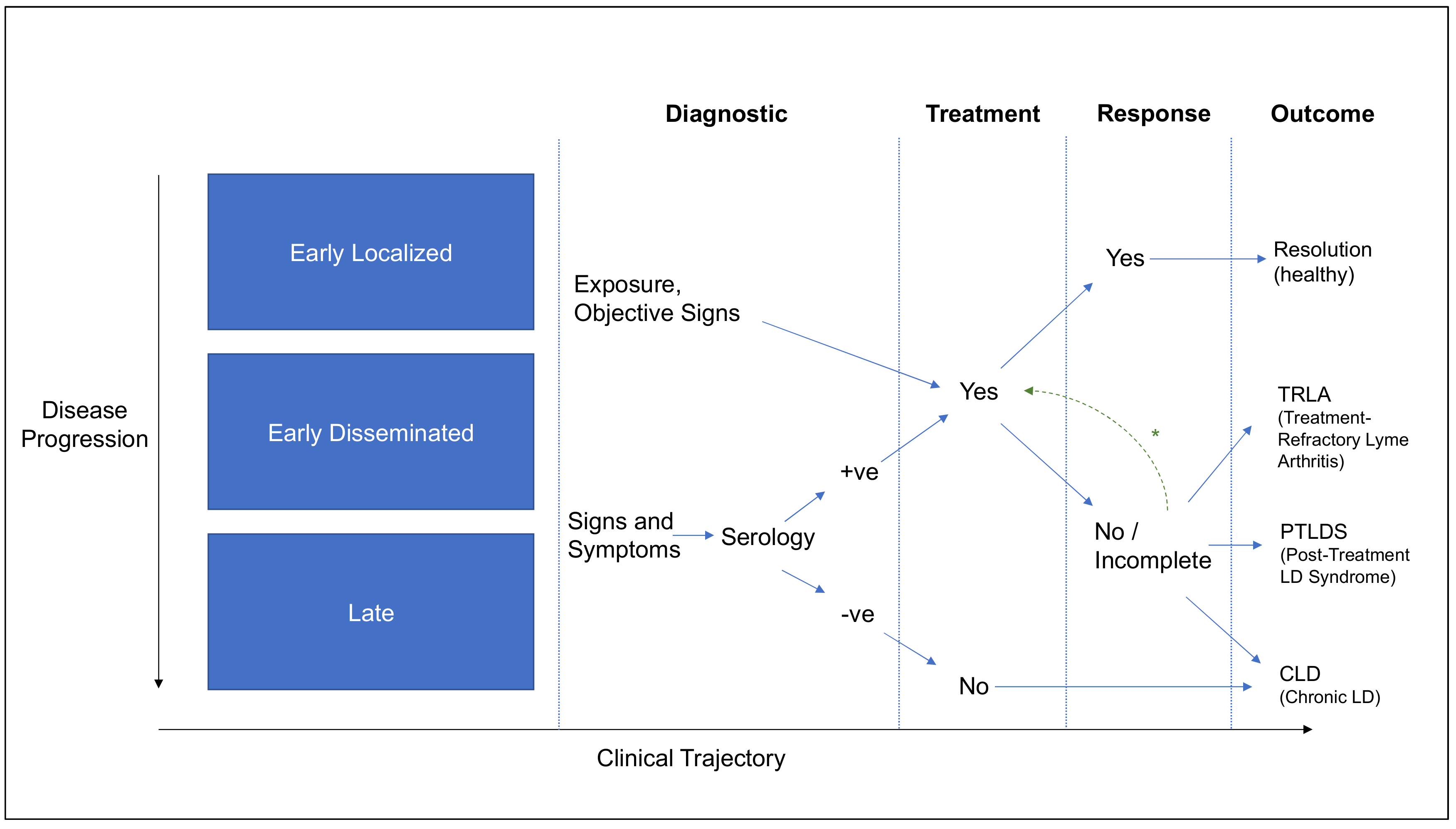
Clinical signs and two-tiered serology consistent with U.S. CDC recommendations are the most common approaches used in diagnosis and research study inclusion criteria. (Section 2.1)
|
LD presentation has been classified by stage of infection (early localized to late disease) and also by outcome (treatment refractory Lyme arthritis (TRLA), post-treatment Lyme disease syndrome (PTLDS), chronic Lyme disease (CLD)). (Section 2.2 and Section 2.3)
|
Host genetic predisposition to treatment-refractory LA has been proposed based on an autoimmune mechanism associated with HLA-DR alleles. (Section 3.1)
|
Host diet and blood cholesterol levels could be implicated in the disease progression. (Section 3.2)
|
The invasive potential of Bb is associated with serotype. (Section 4.1)
|
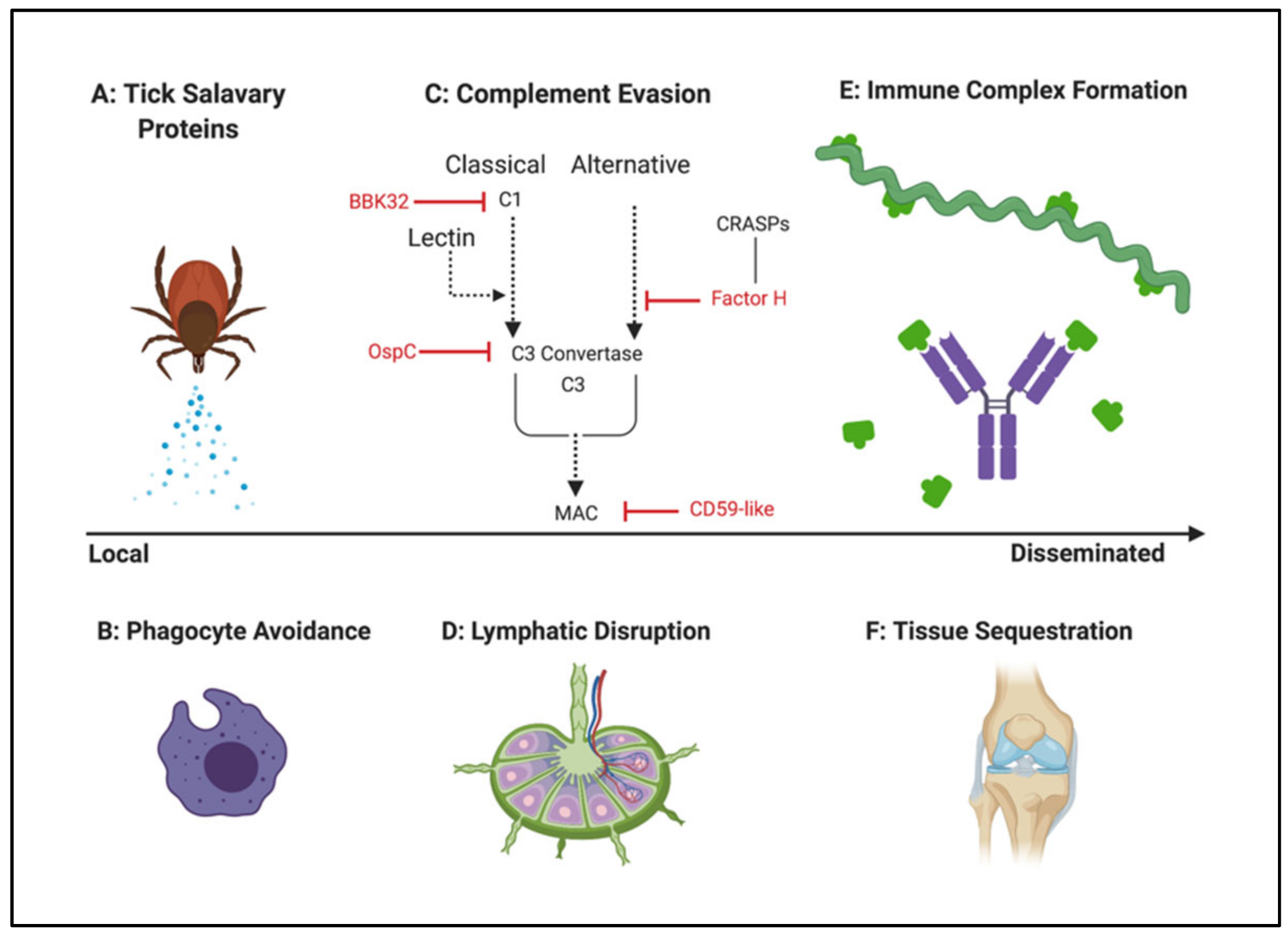
Mammalian host colonization requires an arsenal of strategies to move through the body, evade and subvert the immune system, and invade distal tissues. (Section 4.2)
|
Signalling pathways governing Bb population dynamics are incompletely characterized. (Section 5)
|
Bb antibiotic resistance and persistence have been described in vitro. (Section 5.1)
|
Doxycycline, amoxicillin, or cefuroxime axetil are first-line agents for treatment of early infection in North America. (Section 5.1.4)
|
Bb is capable of adopting pleomorphic forms under stressful conditions, in vitro. Morphologically similar forms have been identified in tissue samples from patients with confirmed infections with Borrelia. (Section 5.2)
|
6.1. Classifying and Studying Lyme Disease
6.2. Clinical Microbiology, Pathogenesis, and Treatment
6.3. Interdisciplinary Solutions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
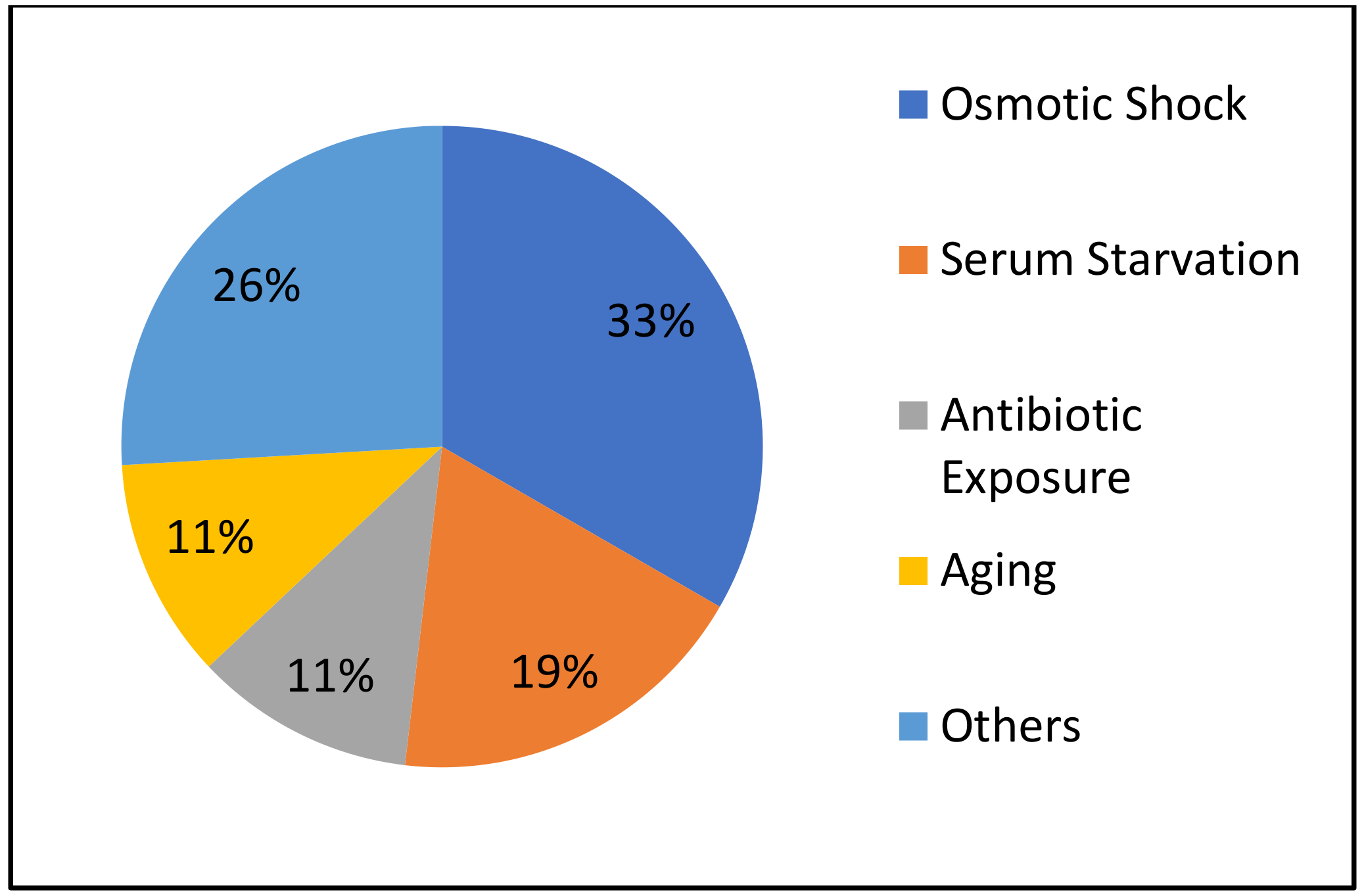
| Induction Method | Species/Strain | Conversion Kinetics | Morphology | Viability | Reference |
|---|---|---|---|---|---|
| Distilled Water | B. burgdorferi ACA-1 | > 95% converted within 1 min 100% RBs by 4 h; all globular. Aggregates of round bodies formed at 1 week. | Round; spirochete whirled into itself after expansion of outer membrane. | Assessed viability by reversion and propagation of motile spirochetes from round bodies. Also confirmed the presence of RNA in round bodies using an acridine orange stain. | [328] |
| B. burgdorferi B31 and ADB1 | Majority converted within 5 min. | Tangled into a general spherical shape approximately 3–5 μm in diameter. | Viability was indicated by reversion into helical spirochetes following immersion in BSK-II. Could not exclude possibility of unconverted spirochetes contributing to the observed “reversion”. | [313] | |
| B. burgdorferi B31 and FlaB mutant (MC-1) | > 95% converted within the first 2–3 h. | Globular (approximately 1 μm in diameter) with the end of the cylinder protruding outwards. | Mobile spirochetes were recovered after incubation in BSK-II from 4-day old round bodies. RNA was also isolated even after 16 days in distilled water. | [329] | |
| B. burgdorferi B31 | 85% converted by 10 min. | Spherical with a mean diameter of 2.8 μm. TEMs depicted intact inner and outer membranes. | 10 min and 2 h round bodies reverted back to helical spirochetes and reached a concentration of 4.0 × 107/mL after incubation in BSK-II for 6 and 8 days respectively. | [314] | |
| Cerebral Spinal Fluid | B. burgdorferi ACA-1 | 100% conversion within 24 h (37C incubation). Cyst formation was different depending on concentration of spinal fluid protein (higher = slower conversion). | Spherical, approximately 2 μm in diameter. | Reversion to motile spirochetes of logarithmic phase density when reintroduced into BSK medium. | [319] |
| RPMI-1640 | B. burgdorferi B31 (high-passage) and T15 (low-passage) | 90% converted into “cysts” by 48 h. | Twisted and knotted into a rounded form with an outer envelope. Approximately 2–3 μm in diameter. | Immediate reversion to non-motile, intact spirochetes upon the addition of rabbit serum or BSK. Cysts did not open when 20% sucrose was added to the culture. Cells began to regain motility 12–15 h after emerging from the cysts. | [353] |
| B. burgdorferi B31 | 17% converted into round bodies by day 4. | Spherical with folded spirochetal cylinder within a membrane. TEMs depict intact inner and outer membranes. Approximately 2.8 μm in diameter. | Did not specify. | [314] | |
| B. burgdorferi 297 | Quantitative data not provided. ΔrelB mutants converted faster than their wild type counterparts in the RPMI medium. | Spherical with either smooth surfaces or rough/blebbing surface. Spirochete appears to fold and curl within a round membrane with part of cylinder protruding out. Approximately 1.2–2 μm in diameter. | A small proportion of the round bodies did not stain with propidium iodide, indicating intact membranes and subsequent viability. | [225] | |
| Serum free BSKII (BSKII–S) or BSK-H (-rabbit serum) | B. burgdorferi ACA-1 | By one week, round “cystic” structures were seen at an unspecified proportion. By six weeks, only cystic structures were seen. | A combination of round and irregular shapes with a mean diameter ranging from 0.5–2.0 µm. | When re-introduced to BSK-H with rabbit serum, mobile spirochetes were seen after six weeks. Likewise, re-introduction of filtrate (un-transformed spirochetes) to regular BSK-H failed to yield mobile spirochetes or growth for at least three months | [354] |
| B. burgdorferi B31 | > 95% converted in 7–10 days. | Morphologically similar to the distilled water induction within the same experiment. | Did not specify. | [329] | |
| B. burgdorferi B31 | No different than control. | NA | NA | [314] | |
| Oxidative Stress | B. garinii BITS | 6% converted over 24 h (in MEM and H2O2). Rapid conversion of majority within 10 min (in H2O and H2O2). | Round expansion of outer membrane with folding of cylinder within. Size undetermined/not specified. | Reversion back to helical spirochetes took over 2 months. | [321] |
| Changes in pH | B. garinii BITS | Conversion of small fraction at pH 2 and 11 after 30 min. | Not specified. | Not specified. | [321] |
| B. burgdorferi B31 and ADB1 | Not specified. | Not specified | Not specified. | [313] | |
| Changes in Temperature | B. garinii BITS | “Cystic” forms increase as temperature increases when incubated in MEM. | Not specified. | Reversion to helical spirochetes within 30 days of incubation under regular growth conditions. | [321] |
| B. burgdorferi B31 and ADB1 | Not specified. | Not specified. | Not specified. | [313] | |
| Human Serum | B. burgdorferi B31 | 22% converted to round bodies after 4 days. | Spherical with a mean diameter of 2.8 μm. TEMs depicted intact inner and outer membranes. | Not specified. | [314] |
| Antibiotics | B. burgdorferi B31 | 96% converted into round bodies following a 3 day incubation in 50 μg/mL of amoxicillin. | Not specified. | Reversion to normal spirochetes within 5 days of subculture in BSK-H. | [334] |
| B. burgdorferi B31, H1, and H8 | “Spherical bodies” were present after 18 h in cultures with penicillin, ceftriaxone, and doxycycline. | “Spherical bodies” were described as being 0.8–1.4 μm in diameter. TEM depicts an electron dense sphere surrounded by an outer membrane. | Subculture from all samples failed to yield morphologically typical Borrelia by two weeks. | [333] | |
| Eukaryotic Cell Co-Culture/Infection | B. burgdorferi B31 and ADB1 | 1 week of incubation with chicken primary sympathetic neurons and rat astrocytes seem to yield “ring shaped cystic forms” that can aggregate. | The average “ring shaped cystic form” appears to be anywhere from 1–2 μm in diameter. | Not specified. | [313] |
References
- Weber, K.; Pfister, H.-W. History of Lyme Borreliosis in Europe. In Aspects of Lyme Borreliosis; Springer Berlin Heidelberg: Berlin, Heidelberg, 1993; pp. 1–20. [Google Scholar]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme Disease—A Tick-Borne Spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Benach, J.L. Discovery of the Lyme Disease Agent. MBio 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hinckley, A.F.; Connally, N.P.; Meek, J.I.; Johnson, B.J.; Kemperman, M.M.; Feldman, K.A.; White, J.L.; Mead, P.S. Lyme Disease Testing by Large Commercial Laboratories in the United States. Clin. Infect. Dis. 2014, 59, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Saha, S.; Kugeler, K.J.; Delorey, M.J.; Shankar, M.B.; Hinckley, A.F.; Mead, P.S. Incidence of Clinician-Diagnosed Lyme Disease, United States, 2005–2010. Emerg. Infect. Dis. 2015, 21, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Schotthoefer, A.M.; Frost, H.M. Ecology and epidemiology of Lyme borreliosis. Clin. Lab. Med. 2015, 35, 723–743. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital signs: Trends in reported vectorborne disease cases—United States and Territories, 2004–2016. Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Benach, J.L. Malaria and Lyme disease—The largest vector-borne US epidemics in the last 100 years: Success and failure of public health. BMC Public Health 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Franke, J.; Hildebrandt, A.; Dorn, W. Exploring gaps in our knowledge on Lyme borreliosis spirochaetes—Updates on complex heterogeneity, ecology, and pathogenicity. Ticks Tick. Borne. Dis. 2013, 4, 11–25. [Google Scholar] [CrossRef]
- Adeolu, M.; Gupta, R.S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: The emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the Lyme disease Borrelia (Borrelia burgdorferi sensu lato complex). Antonie Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar] [CrossRef]
- Margos, G.; Gofton, A.; Wibberg, D.; Dangel, A.; Marosevic, D.; Loh, S.M.; Oskam, C.; Fingerle, V. The genus Borrelia reloaded. PLoS ONE 2018, 13, e0208432. [Google Scholar] [CrossRef]
- Margos, G.; Hepner, S.; Mang, C.; Marosevic, D.; Reynolds, S.E.; Krebs, S.; Sing, A.; Derdakova, M.; Reiter, M.A.; Fingerle, V. Lost in plasmids: Next generation sequencing and the complex genome of the tick-borne pathogen Borrelia burgdorferi. BMC Genom. 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R.; Di, L.; Akther, S.; Mongodin, E.F.; Luft, B.J.; Schutzer, S.E.; Fraser, C.M.; Qiu, W.-G. Primordial origin and diversification of plasmids in Lyme disease agent bacteria. BMC Genom. 2018, 19, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Corona, A.; Schwartz, I. Borrelia burgdorferi: Carbon Metabolism and the Tick-Mammal Enzootic Cycle. Microbiol. Spectr. 2015, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Radolf, J.D.; Caimano, M.J.; Stevenson, B.; Hu, L.T. Of ticks, mice and men: Understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat. Rev. Microbiol. 2012, 10, 87–99. [Google Scholar] [CrossRef]
- Mead, P.; Hook, S.; Niesobecki, S.; Ray, J.; Meek, J.; Delorey, M.; Prue, C.; Hinckley, A. Risk factors for tick exposure in suburban settings in the Northeastern United States. Ticks Tick. Borne. Dis. 2018, 9, 319–324. [Google Scholar] [CrossRef]
- Kowalec, M.; Szewczyk, T.; Welc-Falęciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—Are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasit. Vectors 2017, 10, 1–19. [Google Scholar] [CrossRef]
- Morshed, M.G.; Scott, J.D.; Fernando, K.; Beati, L.; Mazerolle, D.F.; Geddes, G.; Durden, L.A. Migratory songbirds disperse ticks across Canada, and first isolation of the Lyme disease spirochete, Borrelia burgdorferi, from the avian tick, Ixodes auritulus. J. Parasitol. 2005, 91, 780–790. [Google Scholar] [CrossRef]
- Scott, J.D.; Scott, C.M.; Anderson, J.F. The Establishment of a Blacklegged Tick Population by Migratory Songbirds in Ontario, Canada. J. Veter Sci. Med. 2014, 2, 1–5. [Google Scholar] [CrossRef]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl. Environ. Microbiol. 2008, 74, 1780–1790. [Google Scholar] [CrossRef]
- Wills, M.K.B.; Kirby, A.M.; Lloyd, V.K. Detecting the Lyme Disease Spirochete, Borrelia burgdorferi, in Ticks Using Nested PCR. J. Vis. Exp. 2018, 56471. [Google Scholar] [CrossRef]
- Pachner, A.R. Neurologic manifestations of Lyme disease, the new “great imitator”. Rev. Infect. Dis. 1989, 11 (Suppl. 6), S1482–S1486. [Google Scholar] [CrossRef]
- Stanek, G.; Strle, F. Lyme Disease—European Perspective. Infect. Dis. Clin. N. Am. 2008, 22, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Strle, F. Lyme borreliosis: A European perspective on diagnosis and clinical management. Curr. Opin. Infect. Dis. 2009, 22, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Aucott, J.N.; Rebman, A.W.; Crowder, L.A.; Kortte, K.B. Post-treatment Lyme disease syndrome symptomatology and the impact on life functioning: Is there something here? Qual. Life Res. 2013, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J. Consequences of treatment delay in Lyme disease. J. Eval. Clin. Pract. 2007, 13, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.; Nelson, C.; Molins, C.; Mead, P.; Schriefer, M. Current Guidelines, Common Clinical Pitfalls, and Future Directions for Laboratory Diagnosis of Lyme Disease, United States. Emerg. Infect. Dis. 2016, 22, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.L. PCR in laboratory diagnosis of human Borrelia burgdorferi infections. Clin. Microbiol. Rev. 1997, 10, 185–201. [Google Scholar] [CrossRef]
- Doern, G. V Detection of selected fastidious bacteria. Clin. Infect. Dis. 2000, 30, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Eldin, C.; Raffetin, A.; Bouiller, K.; Hansmann, Y.; Roblot, F.; Raoult, D.; Parola, P. Review of European and American guidelines for the diagnosis of Lyme borreliosis. Med. Mal. Infect. 2019, 49, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Leeflang, M.M.G.; Ang, C.W.; Berkhout, J.; Bijlmer, H.A.; Van Bortel, W.; Brandenburg, A.H.; Van Burgel, N.D.; Van Dam, A.P.; Dessau, R.B.; Fingerle, V.; et al. The diagnostic accuracy of serological tests for Lyme borreliosis in Europe: A systematic review and meta-analysis. BMC Infect. Dis. 2016, 16, 140. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.B. Laboratory diagnostic testing for Borrelia burgdorferi infection. Lyme Dis. An Evidence-Based Approach 2011, 73–88. [Google Scholar]
- Waddell, L.A.; Greig, J.; Mascarenhas, M.; Harding, S.; Lindsay, R.; Ogden, N. The Accuracy of Diagnostic Tests for Lyme Disease in Humans, A Systematic Review and Meta-Analysis of North American Research. PLoS ONE 2016, 11, e0168613. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.; Petersen, J.; Hinckley, A. Updated CDC Recommendation for Serologic Diagnosis of Lyme Disease. MMWR. Morb. Mortal. Wkly. Rep. 2019, 68, 703. [Google Scholar] [CrossRef] [PubMed]
- Tracy, K.E.; Baumgarth, N. Borrelia burgdorferi Manipulates Innate and Adaptive Immunity to Establish Persistence in Rodent Reservoir Hosts. Front. Immunol. 2017, 8, 11–16. [Google Scholar] [CrossRef]
- Liang, F.T.; Yan, J.; Mbow, M.L.; Sviat, S.L.; Gilmore, R.D.; Mamula, M.; Fikrig, E. Borrelia burgdorferi changes its surface antigenic expression in response to host immune responses. Infect. Immun. 2004, 72, 5759–5767. [Google Scholar] [CrossRef]
- Chandra, A.; Wormser, G.P.; Marques, A.R.; Latov, N.; Alaedini, A. Anti-Borrelia burgdorferi antibody profile in post-lyme disease syndrome. Clin. Vaccine Immunol. 2011, 18, 767–771. [Google Scholar] [CrossRef]
- Hilton, E.; Tramontano, A.; Voti, J.D.E. Temporal Study of Immunoglobulin M Seroreactivity to Borrelia burgdorferi in Patients Treated for Lyme Borreliosis. J. Clin. Microbiol. 1996, 35, 774–776. [Google Scholar]
- Peltomaa, M.; McHugh, G.; Steere, A.C. Persistence of the Antibody Response to the VlsE Sixth Invariant Region (IR 6) Peptide of Borrelia burgdorferi after Successful Antibiotic Treatment of Lyme Disease. J. Infect. Dis. 2003, 187, 1178–1186. [Google Scholar] [CrossRef]
- Glatz, M.; Golestani, M.; Kerl, H.; Mu, R.R. Clinical Relevance of Different IgG and IgM Serum Antibody Responses to Borrelia burgdorferi After Antibiotic Therapy for Erythema Migrans. Arch. Dermatol. 2006, 142, 862–868. [Google Scholar] [CrossRef]
- Rebman, A.W.; Crowder, L.A.; Kirkpatrick, A.; Aucott, J.N. Characteristics of seroconversion and implications for diagnosis of post-treatment Lyme disease syndrome: Acute and convalescent serology among a prospective cohort of early Lyme disease patients. Clin. Rheumatol. 2014, 34, 585–589. [Google Scholar] [CrossRef]
- Blum, L.K.; Adamska, J.Z.; Martin, D.S.; Rebman, A.W.; Elliott, S.E.; Cao, R.R.L.; Embers, M.E.; Aucott, J.N.; Soloski, M.J.; Robinson, W.H. Robust B Cell Responses Predict Rapid Resolution of Lyme Disease. Front. Immunol. 2018, 9, 113–135. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Roen, D.R.; Lehmann, P.V.; Kellermann, G.H. An Enhanced ELISPOT Assay for Sensitive Detection of Antigen-Specific T Cell Responses to Borrelia burgdorferi. Cells 2013, 2, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Magni, R.; Espina, B.H.; Shah, K.; Lepene, B.; Mayuga, C.; Douglas, T.A.; Espina, V.; Rucker, S.; Dunlap, R.; Petricoin, E.F.; et al. Application of Nanotrap technology for high sensitivity measurement of urinary outer surface protein A carboxyl-terminus domain in early stage Lyme borreliosis. J. Transl. Med. 2015, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Pícha, D.; Moravcová, L.; Vaňousová, D.; Hercogová, J.; Blechová, Z. DNA persistence after treatment of Lyme borreliosis. Folia Microbiol. (Praha.) 2014, 59, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Vigliotti, J.; Vigliotti, V.; Jones, W.; Shearer, D. Detection of Borreliae in Archived Sera from Patients with Clinically Suspect Lyme Disease. Int. J. Mol. Sci. 2014, 15, 4284–4298. [Google Scholar] [CrossRef]
- Coulter, P.; Lema, C.; Flayhart, D.; Linhardt, A.S.; Aucott, J.N.; Auwaerter, P.G.; Dumler, J.S. Two-Year Evaluation of Borrelia burgdorferi Culture and Supplemental Tests for Definitive Diagnosis of Lyme Disease. J. Clin. Microbiol. 2005, 43, 5080–5084. [Google Scholar] [CrossRef]
- Middelveen, M.; Sapi, E.; Burke, J.; Filush, K.; Franco, A.; Fesler, M.; Stricker, R. Persistent Borrelia Infection in Patients with Ongoing Symptoms of Lyme Disease. Healthcare 2018, 6, 33. [Google Scholar] [CrossRef]
- Puri, B.K.; Segal, D.R.; Monro, J.A. Diagnostic use of the lymphocyte transformation test-memory lymphocyte immunostimulation assay in confirming active lyme borreliosis in clinically and serologically ambiguous cases. Int. J. Clin. Exp. Med. 2014, 7, 5890–5892. [Google Scholar]
- Lahey, L.J.; Panas, M.W.; Mao, R.; Delanoy, M.; Flanagan, J.J.; Binder, S.R.; Rebman, A.W.; Montoya, J.G.; Soloski, M.J.; Steere, A.C.; et al. Development of a Multiantigen Panel for Improved Detection of Borrelia burgdorferi Infection in Early Lyme Disease. J. Clin. Microbiol. 2015, 53, 3834–3841. [Google Scholar] [CrossRef]
- Wormser, G.P.; McKenna, D.; Carlin, J.; Nadelman, R.B.; Cavaliere, F.; Holmgren, D.; Byrne, D.W.; Nowakowski, J. Article Brief Communication: Hematogenous Dissemination in Early. Ann. Intern. Med. 2005, 142, 751–755. [Google Scholar] [CrossRef]
- Steere, A.C. Lyme Disease. N. Engl. J. Med. 2001, 345, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Gibofsky, A.; Patarroyo, M.E.; Winchester, R.J.; Hardin, J.A.; Malawista, S.E. Chronic lyme arthritis. Clinical and immunogenetic differentiation from rheumatoid arthritis. Ann. Intern. Med. 1979, 90, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Aucott, J.N.; Crowder, L.A.; Kortte, K.B. Development of a foundation for a case definition of post-treatment Lyme disease syndrome. Int. J. Infect. Dis. 2013, 17, e443–e449. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Hauffe, H.C.; Carpi, G.; Vourc’h, G.I.; Neteler, M.; Rosà, R. Lyme borreliosis in Europe. Eurosurveillance 2011, 16, 1–8. [Google Scholar]
- Wu, X.B.; Na, R.H.; Wei, S.S.; Zhu, J.S.; Peng, H.J. Distribution of tick-borne diseases in China. Parasit. Vectors 2013, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Borchers, A.T.; Keen, C.L.; Huntley, A.C.; Gershwin, M.E. Lyme disease: A rigorous review of diagnostic criteria and treatment. J. Autoimmun. 2015, 57, 82–115. [Google Scholar] [CrossRef]
- Prevention, C. for D.C. and Case definitions for public health surveillance. MMWR 1990, 39. [Google Scholar]
- Lipsker, D.; Lieber-Mbomeyo, A.; Hedelin, G. How Accurate Is a Clinical Diagnosis of Erythema Chronicum Migrans? Prospective Study Comparing the Diagnostic Accuracy of General Practitioners and Dermatologists in an Area Where Lyme Borreliosis Is Endemic. Arch. Dermatol. 2004, 140, 620–621. [Google Scholar] [CrossRef]
- Stonehouse, A.; Studdiford, J.S.; Henry, C.A. An update on the diagnosis and treatment of early lyme disease: “Focusing on the bull’s eye, you may miss the mark”. J. Emerg. Med. 2010, 39, e147–e151. [Google Scholar] [CrossRef]
- Schutzer, S.E.; Berger, B.W.; Krueger, J.G.; Eshoo, M.W.; Ecker, D.J.; Aucott, J.N. Atypical erythema migrans in patients with PCR-positive Lyme disease. Emerg. Infect. Dis. 2013, 19, 815–817. [Google Scholar] [CrossRef]
- Fix, A.D.; Peña, C.A.; Strickland, G.T. Racial differences in reported lyme disease incidence. Am. J. Epidemiol. 2000, 152, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, V.; Hawkins, R. Under-Detection of Lyme Disease in Canada. Healthcare 2018, 6, 125. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J.; Johnson, L.B.; Maloney, E.L. Evidence assessments and guideline recommendations in Lyme disease: The clinical management of known tick bites, erythema migransrashes and persistent disease. Expert Rev. Anti. Infect. Ther. 2014, 12, 1103–1135. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, E.D. Borrelia burgdorferi (Lyme disease). Pediatr. Rev. 2014, 35, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Wormser, G.P.; Dattwyler, R.J.; Shapiro, E.D.; Halperin, J.J.; Steere, A.C.; Klempner, M.S.; Krause, P.J.; Bakken, J.S.; Strle, F.; Stanek, G.; et al. The Clinical Assessment, Treatment, and Prevention of Lyme Disease, Human Granulocytic Anaplasmosis, and Babesiosis: Clinical Practice Guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2006, 43, 1089–1134. [Google Scholar] [CrossRef]
- Donta, S.T. Late and chronic Lyme disease. Med. Clin. N. Am. 2002, 86, 341–349. [Google Scholar] [CrossRef]
- Stanek, G.; Fingerle, V.; Hunfeld, K.P.; Jaulhac, B.; Kaiser, R.; Krause, A.; Kristoferitsch, W.; O’Connell, S.; Ornstein, K.; Strle, F.; et al. Lyme borreliosis: Clinical case definitions for diagnosis and management in Europe. Clin. Microbiol. Infect. 2011, 17, 69–79. [Google Scholar] [CrossRef]
- Gerstenblith, T.A.; Stern, T.A. Lyme Disease: A Review of Its Epidemiology, Evaluation, and Treatment. Psychosomatics 2014, 55, 421–429. [Google Scholar] [CrossRef]
- Feder, H.M.; Johnson, B.J.B.; O’Connell, S.; Shapiro, E.D.; Steere, A.C.; Wormser, G.P. A Critical Appraisal of “Chronic Lyme Disease”. N. Engl. J. Med. 2007, 357, 1422–1430. [Google Scholar] [CrossRef]
- Hays, R.D.; Sherbourne, C.D.; Mazel, R.M. The rand 36-item health survey 1.0. Health Econ. 1993, 2, 217–227. [Google Scholar] [CrossRef]
- Patrick, D.M.; Miller, R.R.; Gardy, J.L.; Parker, S.M.; Morshed, M.G.; Steiner, T.S.; Singer, J.; Shojania, K.; Tang, P.; Group, C.C.D.S. Lyme Disease Diagnosed by Alternative Methods: A Phenotype Similar to That of Chronic Fatigue Syndrome. Clin. Infect. Dis. 2015, 61, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Stricker, R.; MC, F. Chronic Diseases—International Chronic Lyme Disease: A Working Case Definition. Chron. Diesases-Int. 2017, 4, 1–10. [Google Scholar]
- Citera, M.; Freeman, P.R.; Horowitz, R.I. Empirical validation of the Horowitz Multiple Systemic Infectious Disease Syndrome Questionnaire for suspected Lyme disease. Int. J. Gen. Med. 2017, 10, 249–273. [Google Scholar] [CrossRef] [PubMed]
- Fallon, B.A.; Zubcevik, N.; Bennett, C.; Doshi, S.; Rebman, A.W.; Kishon, R.; Moeller, J.R.; Octavien, N.R.; Aucott, J.N. The General Symptom Questionnaire-30 (GSQ-30): A Brief Measure of Multi-System Symptom Burden in Lyme Disease. Front. Med. 2019, 6, 676–711. [Google Scholar] [CrossRef]
- Beck, D.S.; Barthold, S.W.; Moody, K.D. Susceptibility of Laboratory Rats to Isolates of Borrelia burgdorferi from Different Geographic Areas. Am. J. Trop. Med. Hyg. 1990, 42, 596–600. [Google Scholar]
- Barthold, S.W.; Terwilliger, G.A.; Moody, K.D. Lyme Borreliosis in Laboratory Animals: Effect of Host Species and in Vitro Passage of Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1990, 43, 87–92. [Google Scholar]
- Ramesh, G.; Didier, P.J.; England, J.D.; Santana-Gould, L.; Doyle-Meyers, L.A.; Martin, D.S.; Jacobs, M.B.; Philipp, M.T. Inflammation in the pathogenesis of lyme neuroborreliosis. Am. J. Pathol. 2015, 185, 1344–1360. [Google Scholar] [CrossRef]
- Embers, M.E.; Hasenkampf, N.R.; Jacobs, M.B.; Tardo, A.C.; Doyle-Meyers, L.A.; Philipp, M.T.; Hodzic, E. Variable manifestations, diverse seroreactivity and post-treatment persistence in non-human primates exposed to Borrelia burgdorferi by tick feeding. PLoS ONE 2017, 12, e0189071. [Google Scholar] [CrossRef]
- Barthold, S.W.; Persing, D.H.; Armstrong, A.L.; Peeples, R.A. Kinetics of Borrelia burgdorferi dissemination and evolution of disease after intradermal inoculation of mice. Am. J. Pathol. 1991, 139, 263–273. [Google Scholar]
- Schaible, U.E.; Kramer, M.D.; Museteanu, C.; Zimmer, G.; Mossmann, H.; Simon, M.M. The severe combined immunodeficiency (scid) mouse. A laboratory model for the analysis of Lyme arthritis and carditis. J. Exp. Med. 1989, 170, 1427–1432. [Google Scholar] [CrossRef]
- Schaible, U.E.; Gay, S.; Museteanu, C.; Kramer, M.D.; Zimmer, G.; Eichmann, K.; Museteanu, U.; Simon, M.M. Lyme borreliosis in the severe combined immunodeficiency (scid) mouse manifests predominantly in the joints, heart, and liver. Am. J. Pathol. 1990, 137, 811–820. [Google Scholar] [PubMed]
- Priem, S.; Munkelt, K.; Franz, J.K.; Schneider, U.; Werner, T.; Burmester, G.R.; Krause, A. Epidemiologie und Therapie der Lyme-Arthritis und anderer Manifestationen der Lyme-Borreliose in Deutschland. Zeitschrift für Rheumatol. 2003, 62, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulou, B.P.; Alroy, J.; Huber, B.T. CD28 deficiency exacerbates joint inflammation upon Borrelia burgdorferi infection, resulting in the development of chronic Lyme arthritis. J. Immunol. 2007, 179, 8076–8082. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S.; McInnes, I.B. Immunopathogenesis of Rheumatoid Arthritis. Immunity 2017, 46, 183–196. [Google Scholar] [CrossRef]
- Steere, A.C.; Dwyer, E.; Winchester, R. Association of Chronic Lyme Arthritis with HLA-DR4 and HLA-DR2 Alleles. N. Engl. J. Med. 1990, 323, 219–223. [Google Scholar] [CrossRef]
- Gross, D.M.; Forsthuber, T.; Tary-Lehmann, M.; Etling, C.; Ito, K.; Nagy, Z.A.; Field, J.A.; Steere, A.C.; Huber, B.T. Identification of LFA-1 as a candidate autoantigen in treatment-resistant Lyme arthritis. Science 1998, 281, 703–706. [Google Scholar] [CrossRef]
- Kalish, R.A.; Leong, J.M.; Steere, A.C. Association of treatment-resistant chronic Lyme arthritis with HLA-DR4 and antibody reactivity to OspA and OspB of Borrelia burgdorferi. Infect. Immun. 1993, 61, 2774–2779. [Google Scholar]
- Trollmo, C.; Meyer, A.L.; Steere, A.C.; Hafler, D.A.; Huber, B.T. Molecular Mimicry in Lyme Arthritis Demonstrated at the Single Cell Level: LFA-1α L Is a Partial Agonist for Outer Surface Protein A-Reactive T Cells. J. Immunol. 2001, 166, 5286–5291. [Google Scholar] [CrossRef]
- Benvenga, S.; Santarpia, L.; Trimarchi, F.; Guarneri, F. Human Thyroid Autoantigens and Proteins of Yersinia and Borrelia Share Amino Acid Sequence Homology That Includes Binding Motifs to HLA-DR Molecules and T-Cell Receptor. Thyroid 2006, 16, 225–236. [Google Scholar] [CrossRef]
- Benvenga, S.; Guarneri, F. Molecular mimicry and autoimmune thyroid disease. Rev. Endocr. Metab. Disord. 2016, 17, 485–498. [Google Scholar] [CrossRef]
- Iliopoulou, B.P.; Alroy, J.; Huber, B.T. Persistent arthritis in Borrelia burgdorferi- infected HLA-DR4-positive CD28-negative mice post-antibiotic treatment. Arthritis Rheum. 2008, 58, 3892–3901. [Google Scholar] [CrossRef] [PubMed]
- Ball, R.; Shadomy, S.V.; Meyer, A.; Huber, B.T.; Leffell, M.S.; Zachary, A.; Belotto, M.; Hilton, E.; Bryant-Genevier, M.; Schriefer, M.E.; et al. HLA Type and Immune Response to Borrelia burgdorferi Outer Surface Protein A in People in Whom Arthritis Developed After Lyme Disease Vaccination. Arthritis Rheum. 2009, 60, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Kalish, R.S.; Wood, J.A.; Golde, W.; Bernard, R.; Davis, L.E.; Grimson, R.C.; Coyle, P.K.; Luft, B.J. Human T Lymphocyte Response to Borrelia burgdorferi Infection: No Correlation between Human Leukocyte Function Antigen Type 1 Peptide Response and Clinical Status. J. Infect. Dis. 2003, 187, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Schröder, N.W.J.; Schombel, U.; Heine, H.; Göbel, U.B.; Zähringer, U.; Schumann, R.R. Acylated Cholesteryl Galactoside as a Novel Immunogenic Motif in Borrelia burgdorferi Sensu Stricto. J. Biol. Chem. 2003, 278, 33645–33653. [Google Scholar] [CrossRef] [PubMed]
- Ben-Menachem, G.; Kubler-Kielb, J.; Coxon, B.; Yergey, A.; Schneerson, R. A newly discovered cholesteryl galactoside from Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2003, 100, 7913–7918. [Google Scholar] [CrossRef]
- Crowley, J.T.; Toledo, A.M.; LaRocca, T.J.; Coleman, J.L.; London, E.; Benach, J.L. Lipid Exchange between Borrelia burgdorferi and Host Cells. PLoS Pathog. 2013, 9, e1003109. [Google Scholar] [CrossRef]
- Toledo, A.; Monzón, J.D.; Coleman, J.L.; Garcia-Monco, J.C.; Benach, J.L. Hypercholesterolemia and ApoE deficiency result in severe infection with Lyme disease and relapsing-fever Borrelia. Proc. Natl. Acad. Sci. USA 2015, 112, 5491–5496. [Google Scholar] [CrossRef]
- Straube, R.; Voit-Bak, K.; Gor, A.; Steinmeier, T.; Chrousos, G.P.; Boehm, B.O.; Birkenfeld, A.L.; Barbir, M.; Balanzew, W.; Bornstein, S.R. Lipid Profiles in Lyme Borreliosis: A Potential Role for Apheresis? Horm. Metab. Res. 2019, 51, 326–329. [Google Scholar] [CrossRef]
- Pratt, C.L.; Brown, C.R. The role of eicosanoids in experimental Lyme arthritis. Front. Cell. Infect. Microbiol. 2014, 4, 69. [Google Scholar] [CrossRef]
- Anguita, J.; Samanta, S.; Ananthanarayanan, S.K.; Revilla, B.; Geba, G.P.; Barthold, S.W.; Fikrig, E. Cyclooxygenase 2 activity modulates the severity of murine Lyme arthritis. FEMS Immunol. Med. Microbiol. 2002, 34, 187–191. [Google Scholar] [CrossRef]
- Blaho, V.A.; Mitchell, W.J.; Brown, C.R. Arthritis develops but fails to resolve during inhibition of cyclooxygenase 2 in a murine model of lyme disease. Arthritis Rheum. 2008, 58, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Blaho, V.A.; Buczynski, M.W.; Brown, C.R.; Dennis, E.A. Lipidomic analysis of dynamic eicosanoid responses during the induction and resolution of lyme arthritis. J. Biol. Chem. 2009, 284, 21599–21612. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Desnoyers, M.; Gilbert, K.; Rousseau, G. Cardioprotective Effects of Omega-3 Polyunsaturated Fatty Acids: Dichotomy between Experimental and Clinical Studies. Mar. Drugs 2018, 16, 234. [Google Scholar] [CrossRef] [PubMed]
- Dumlao, D.S.; Cunningham, A.M.; Wax, L.E.; Norris, P.C.; Hanks, J.H.; Halpin, R.; Lett, K.M.; Blaho, V.A.; Mitchell, W.J.; Fritsche, K.L.; et al. Dietary fish oil substitution alters the eicosanoid profile in ankle joints of mice during Lyme infection. J. Nutr. 2012, 142, 1582–1589. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L. Host-pathogen interactions: The attributes of virulence. J. Infect. Dis. 2001, 184, 337–344. [Google Scholar] [CrossRef]
- Adusumilli, S.; Booth, C.J.; Anguita, J.; Fikrig, E. Passage through Ixodes scapularis Ticks Enhances the Virulence of a Weakly Pathogenic Isolate of Borrelia burgdorferi. Infect. Immun. 2009, 78, 138–144. [Google Scholar] [CrossRef]
- Kasumba, I.N.; Bestor, A.; Tilly, K.; Rosa, P.A. Virulence of the Lyme disease spirochete before and after the tick bloodmeal: A quantitative assessment. Parasit. Vectors 2016, 9, 1–8. [Google Scholar] [CrossRef]
- Steere, A.C.; Sikand, V.K.; Schoen, R.T.; Nowakowski, J. Asymptomatic infection with Borrelia burgdorferi. Clin. Infect. Dis. 2003, 37, 528–532. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Vancova, M.; Clark, K.; Grubhoffer, L.; Oliver, J.H., Jr. Isolation of live Borrelia burgdorferi sensu lato spirochaetes from patients with undefined disorders and symptoms not typical for Lyme borreliosis. Clin. Microbiol. Infect. 2016, 22, 267.e9–267.e15. [Google Scholar] [CrossRef]
- Cutler, S.J.; Ružić-Sabljić, E.; Potkonjak, A. Emerging borreliae—Expanding beyond Lyme borreliosis. Mol. Cell. Probes 2017, 31, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Piesman, J.; Lane, R.S.; Ogden, N.H.; Sing, A.; Straubinger, R.K.; Fingerle, V. Borrelia kurtenbachii sp. nov., a widely distributed member of the Borrelia burgdorferi sensu lato species complex in North America. Int. J. Syst. Evol. Microbiol. 2014, 64, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Fedorova, N.; Lane, R.S. Genetic diversity of Borrelia burgdorferi and detection of B. bissettii-like DNA in serum of north-coastal California residents. J. Clin. Microbiol. 2011, 49, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.S.; Mead, P.S.; Johnson, D.K.H.; Neitzel, D.F.; Respicio-Kingry, L.B.; Davis, J.P.; Schiffman, E.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; et al. Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: A descriptive study. Lancet Infect. Dis. 2016, 16, 556–564. [Google Scholar] [CrossRef]
- Golovchenko, M.; Vancová, M.; Clark, K.; Oliver, J.H.; Grubhoffer, L.; Rudenko, N. A divergent spirochete strain isolated from a resident of the southeastern United States was identified by multilocus sequence typing as Borrelia bissettii. Parasit. Vectors 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Baranton, G.; Postic, D.; Saint Girons, I.; Boerlin, P.; Piffaretti, J.C.; Assous, M.; Grimont, P.A. Delineation of Borrelia burgdorferi sensu stricto, Borrelia garinii sp. nov., and group VS461 associated with Lyme borreliosis. Int. J. Syst. Bacteriol. 1992, 42, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, A.P.; Kuiper, H.; Vos, K.; Widjojokusumo, A.; de Jongh, B.M.; Spanjaard, L.; Ramselaar, A.C.; Kramer, M.D.; Dankert, J. Different genospecies of Borrelia burgdorferi are associated with distinct clinical manifestations of Lyme borreliosis. Clin. Infect. Dis. 1993, 17, 708–717. [Google Scholar] [CrossRef]
- Picken, R.N.; Strle, F.; Picken, M.M.; Ružić-Sabljić, E.; Maraspin, V.; Lotrič-Furlan, S.; Cimperman, J. Identification of Three Species of Borrelia burgdorferi Sensu Lato (B. burgdorferi Sensu Stricto, B. garinii, and B. afzelii) Among Isolates from Acrodermatitis Chronica Atrophicans Lesions. J. Investig. Dermatol. 1998, 110, 211–214. [Google Scholar] [CrossRef]
- Assous, M.V.; Postic, D.; Paul, G.; Névot, P.; Baranton, G. Western blot analysis of sera from Lyme borreliosis patients according to the genomic species of the Borrelia strains used as antigens. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12, 261–268. [Google Scholar] [CrossRef]
- Coipan, E.C.; Jahfari, S.; Fonville, M.; Oei, G.A.; Spanjaard, L.; Takumi, K.; Hovius, J.W.R.; Sprong, H. Imbalanced presence of Borrelia burgdorferi s.l. multilocus sequence types in clinical manifestations of Lyme borreliosis. Infect. Genet. Evol. 2016, 42, 66–76. [Google Scholar] [CrossRef]
- Jahfari, S.; Krawczyk, A.; Coipan, E.C.; Fonville, M.; Hovius, J.W.; Sprong, H.; Takumi, K. Enzootic origins for clinical manifestations of Lyme borreliosis. Infect. Genet. Evol. 2017, 49, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Tijsse-Klasen, E.; Pandak, N.; Hengeveld, P.; Takumi, K.; Koopmans, M.P.G.; Sprong, H. Ability to cause erythema migrans differs between Borrelia burgdorferi sensu lato isolates. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Mechai, S.; Margos, G.; Feil, E.J.; Barairo, N.; Lindsay, L.R.; Michel, P.; Ogden, N.H. Evidence for Host-Genotype Associations of Borrelia burgdorferi Sensu Stricto. PLoS ONE 2016, 11, e0149345. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Muellegger, R.R.; Means, T.K.; Lee, M.; Glickstein, L.J.; Damle, N.; Sikand, V.K.; Luster, A.D.; Steere, A.C. Higher mRNA Levels of Chemokines and Cytokines Associated with Macrophage Activation in Erythema Migrans Skin Lesions in Patients from the United States than in Patients from Austria with Lyme Borreliosis. Clin. Infect. Dis. 2008, 46, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Cerar, T.; Strle, F.; Stupica, D.; Ružić-Sabljić, E.; McHugh, G.; Steere, A.C.; Strle, K. Differences in Genotype, Clinical Features, and Inflammatory Potential of Borrelia burgdorferi sensu stricto Strains from Europe and the United States. Emerg. Infect. Dis. 2016, 22, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Baranton, G.; Seinost, G.; Theodore, G.; Postic, D.; Dykhuizen, D. Distinct levels of genetic diversity of Borrelia burgdorferi are associated with different aspects of pathogenicity. Res. Microbiol. 2001, 152, 149–156. [Google Scholar] [CrossRef]
- Wang, G.; Liveris, D.; Mukherjee, P.; Jungnick, S.; Margos, G.; Schwartz, I. Molecular Typing of Borrelia burgdorferi. Curr. Protoc. Microbiol. 2014, 34, 12C.5.1–12C.5.31. [Google Scholar]
- Wang, I.N.; Dykhuizen, D.E.; Qiu, W.; Dunn, J.J.; Bosler, E.M.; Luft, B.J. Genetic diversity of ospC in a local population of Borrelia burgdorferi sensu stricto. Genetics 1999, 151, 15–30. [Google Scholar]
- Izac, J.R.; Marconi, R.T. Diversity of the Lyme Disease Spirochetes and its Influence on Immune Responses to Infection and Vaccination. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 671–686. [Google Scholar] [CrossRef]
- Kenedy, M.R.; Lenhart, T.R.; Akins, D.R. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunol. Med. Microbiol. 2012, 66, 1–19. [Google Scholar] [CrossRef]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; Vanraden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. 2006, 74, 3554–3564. [Google Scholar] [CrossRef] [PubMed]
- Skare, J.T.; Shaw, D.K.; Trzeciakowski, J.P.; Hyde, J.A. In Vivo Imaging Demonstrates That Borrelia burgdorferi ospC Is Uniquely Expressed Temporally and Spatially throughout Experimental Infection. PLoS ONE 2016, 11, e0162501. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, S.E.; Troxell, B.; Yang, Y.; Brandt, S.L.; Li, H.; Sandusky, G.E.; Condon, K.W.; Serezani, C.H.; Yang, X.F. Outer surface protein OspC is an antiphagocytic factor that protects Borrelia burgdorferi from phagocytosis by macrophages. Infect. Immun. 2015, 83, 4848–4860. [Google Scholar] [CrossRef] [PubMed]
- Caine, J.A.; Lin, Y.-P.; Kessler, J.R.; Sato, H.; Leong, J.M.; Coburn, J. Borrelia burgdorferi outer surface protein C (OspC) binds complement component C4b and confers bloodstream survival. Cell. Microbiol. 2017, 19, e12786. [Google Scholar] [CrossRef]
- Tilly, K.; Casjens, S.; Stevenson, B.; Bono, J.L.; Samuels, D.S.; Hogan, D.; Rosa, P. The Borrelia burgdorferi circular plasmid cp26: Conservation of plasmid structure and targeted inactivation of the ospC gene. Mol. Microbiol. 1997, 25, 361–373. [Google Scholar] [CrossRef]
- Brisson, D.; Dykhuizen, D.E. ospC diversity in Borrelia burgdorferi: Different hosts are different niches. Genetics 2004, 168, 713–722. [Google Scholar] [CrossRef]
- Earnhart, C.G.; Buckles, E.L.; Dumler, J.S.; Marconi, R.T. Demonstration of OspC Type Diversity in Invasive Human Lyme Disease Isolates and Identification of Previously Uncharacterized Epitopes That Define the Specificity of the OspC Murine Antibody Response. Infect. Immun. 2005, 73, 7869–7877. [Google Scholar] [CrossRef]
- Barbour, A.G.; Travinsky, B. Evolution and Distribution of the ospC Gene, a Transferable Serotype Determinant of Borrelia burgdorferi. MBio 2010, 1, 111–176. [Google Scholar] [CrossRef]
- Seinost, G.; Dykhuizen, D.E.; Dattwyler, R.J.; Golde, W.T.; Dunn, J.J.; Wang, I.N.; Wormser, G.P.; Schriefer, M.E.; Luft, B.J. Four clones of Borrelia burgdorferi sensu stricto cause invasive infection in humans. Infect. Immun. 1999, 67, 3518–3524. [Google Scholar]
- Wormser, G.P.; Brisson, D.; Liveris, D.; Hanincova, K.; Sandigursky, S.; Nowakowski, J.; Nadelman, R.B.; Ludin, S.; Schwartz, I. Borrelia burgdorferi genotype predicts the capacity for hematogenous dissemination during early Lyme disease. J. Infect. Dis. 2008, 198, 1358–1364. [Google Scholar] [CrossRef]
- Lagal, V.; Postic, D.; Ružić-Sabljić, E.; Baranton, G. Genetic diversity among Borrelia strains determined by single-strand conformation polymorphism analysis of the ospC gene and its association with invasiveness. J. Clin. Microbiol. 2003, 41, 5059–5065. [Google Scholar] [CrossRef] [PubMed]
- Brisson, D.; Baxamusa, N.; Schwartz, I.; Wormser, G.P. Biodiversity of Borrelia burgdorferi Strains in Tissues of Lyme Disease Patients. PLoS ONE 2011, 6, e22926. [Google Scholar] [CrossRef] [PubMed]
- Dykhuizen, D.E.; Brisson, D.; Sandigursky, S.; Wormser, G.P.; Nowakowski, J.; Nadelman, R.B.; Schwartz, I. The propensity of different Borrelia burgdorferi sensu stricto genotypes to cause disseminated infections in humans. Am. J. Trop. Med. Hyg. 2008, 78, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Liveris, D.; Varde, S.; Iyer, R.; Koenig, S.; Bittker, S.; Cooper, D.; McKenna, D.; Nowakowski, J.; Nadelman, R.B.; Wormser, G.P.; et al. Genetic diversity of Borrelia burgdorferi in lyme disease patients as determined by culture versus direct PCR with clinical specimens. J. Clin. Microbiol. 1999, 37, 565–569. [Google Scholar] [PubMed]
- Eshoo, M.W.; Crowder, C.C.; Rebman, A.W.; Rounds, M.A.; Matthews, H.E.; Picuri, J.M.; Soloski, M.J.; Ecker, D.J.; Schutzer, S.E.; Aucott, J.N. Direct molecular detection and genotyping of Borrelia burgdorferi from whole blood of patients with early Lyme disease. PLoS ONE 2012, 7, e36825. [Google Scholar] [CrossRef] [PubMed]
- Mosel, M.R.; Carolan, H.E.; Rebman, A.W.; Castro, S.; Massire, C.; Ecker, D.J.; Soloski, M.J.; Aucott, J.N.; Eshoo, M.W. Molecular Testing of Serial Blood Specimens from Patients with Early Lyme Disease during Treatment Reveals Changing Coinfection with Mixtures of Borrelia burgdorferi Genotypes. Antimicrob. Agents Chemother. 2019, 63, 1625–1626. [Google Scholar] [CrossRef]
- Wormser, G.P.; Liveris, D.; Hanincova, K.; Brisson, D.; Ludin, S.; Stracuzzi, V.J.; Embers, M.E.; Philipp, M.T.; Levin, A.; Aguero Rosenfeld, M.; et al. Effect of Borrelia burgdorferi genotype on the sensitivity of C6 and 2-tier testing in North American patients with culture-confirmed Lyme disease. Clin. Infect. Dis. 2008, 47, 910–914. [Google Scholar] [CrossRef]
- Jones, K.L.; McHugh, G.A.; Glickstein, L.J.; Steere, A.C. Analysis of Borrelia burgdorferi genotypes in patients with Lyme arthritis: High frequency of ribosomal RNA intergenic spacer type 1 strains in antibiotic-refractory arthritis. Arthritis Rheum. 2009, 60, 2174–2182. [Google Scholar] [CrossRef]
- Strle, K.; Jones, K.L.; Drouin, E.E.; Li, X.; Steere, A.C. Borrelia burgdorferi RST1 (OspC Type A) Genotype Is Associated with Greater Inflammation and More Severe Lyme Disease. AJPA 2011, 178, 2726–2739. [Google Scholar] [CrossRef]
- Lagal, V.; Portnoï, D.; Faure, G.; Postic, D.; Baranton, G. Borrelia burgdorferi sensu stricto invasiveness is correlated with OspC–plasminogen affinity. Microbes Infect. 2006, 8, 645–652. [Google Scholar] [CrossRef]
- Qiu, W.-G. Wide Distribution of a High-Virulence Borrelia burgdorferi Clone in Europe and North America. Emerg. Infect. Dis. 2008, 14, 1097–1104. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barbour, A.G. Infection resistance and tolerance in Peromyscus spp., natural reservoirs of microbes that are virulent for humans. Semin. Cell Dev. Biol. 2017, 61, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Barthold, S.W.; BECK, D.S.; HANSEN, G.M.; Terwilliger, G.A.; Moody, K.D. Lyme Borreliosis in Selected Strains and Ages of Laboratory Mice. J. Infect. Dis. 1990, 162, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Aucott, J.N.; Seifter, A.; Rebman, A.W. Probable late lyme disease: A variant manifestation of untreated Borrelia burgdorferi infection. BMC Infect. Dis. 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Kraiczy, P.; Skerka, C.; Kirschfink, M.; Zipfel, P.F.; Brade, V. Mechanism of complement resistance of pathogenic Borrelia burgdorferi isolates. Int. Immunopharmacol. 2001, 1, 393–401. [Google Scholar] [CrossRef]
- Kraiczy, P.; Skerka, C.; Kirschfink, M.; Zipfel, P.F.; Brade, V. Immune evasion of Borrelia burgdorferi: Insufficient killing of the pathogens by complement and antibody. Int. J. Med. Microbiol. 2002, 291, 141–146. [Google Scholar] [CrossRef]
- Embers, M.E.; Ramamoorthy, R.; Philipp, M.T. Survival strategies of Borrelia burgdorferi, the etiologic agent of Lyme disease. Microbes Infect. 2004, 6, 312–318. [Google Scholar] [CrossRef]
- Berndtson, K. Review of evidence for immune evasion and persistent infection in Lyme disease. Int. J. Gen. Med. 2013, 6, 216–291. [Google Scholar] [CrossRef]
- Petzke, M.; Schwartz, I. Borrelia burgdorferi Pathogenesis and the Immune Response. Clin. Lab. Med. 2015, 35, 745–764. [Google Scholar] [CrossRef]
- Kraiczy, P. Hide and Seek: How Lyme Disease Spirochetes Overcome Complement Attack. Front. Immunol. 2016, 7, 385. [Google Scholar] [CrossRef]
- Aslam, B.; Nisar, M.A.; Khurshid, M.; Farooq Salamat, M.K. Immune escape strategies of Borrelia burgdorferi. Future Microbiol. 2017, 12, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.W. Complement Evasion in Borrelia spirochetes: Mechanisms and Opportunities for Intervention. Antibiotics 2019, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Hovius, J.W.R. Spitting Image: Tick Saliva Assists the Causative Agent of Lyme Disease in Evading Host Skin’s Innate Immune Response. J. Investig. Dermatol. 2009, 129, 2337–2339. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; Hovius, J.W.R.; van Burgel, N.D.; Ramamoorthi, N.; Fikrig, E.; van Dam, A.P. The tick salivary protein Salp15 inhibits the killing of serum-sensitive Borrelia burgdorferi sensu lato isolates. Infect. Immun. 2008, 76, 2888–2894. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Narasimhan, S.; Neelakanta, G.; Kumar, M.; Pal, U.; Fikrig, E.; Norgard, M.V. Activation of the RpoN-RpoS regulatory pathway during the enzootic life cycle of Borrelia burgdorferi. BMC Microbiol. 2012, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- de Taeye, S.W.; Kreuk, L.; van Dam, A.P.; Hovius, J.W.; Schuijt, T.J. Complement evasion by Borrelia burgdorferi: It takes three to tango. Trends Parasitol. 2013, 29, 119–128. [Google Scholar] [CrossRef]
- Garcia, B.L.; Zhi, H.; Wager, B.; Höök, M.; Skare, J.T. Borrelia burgdorferi BBK32 Inhibits the Classical Pathway by Blocking Activation of the C1 Complement Complex. PLoS Pathog. 2016, 12, e1005404–e1005428. [Google Scholar] [CrossRef]
- Pausa, M.; Pellis, V.; Cinco, M.; Giulianini, P.G.; Presani, G.; Perticarari, S.; Murgia, R.; Tedesco, F. Serum-Resistant Strains of Borrelia burgdorferi Evade Complement-Mediated Killing by Expressing a CD59-Like Complement Inhibitory Molecule. J. Immunol. 2003, 170, 3214–3222. [Google Scholar] [CrossRef]
- Lazarus, J.J.; Kay, M.A.; McCarter, A.L.; Wooten, R.M. Viable Borrelia burgdorferi enhances interleukin-10 production and suppresses activation of murine macrophages. Infect. Immun. 2008, 76, 1153–1162. [Google Scholar] [CrossRef]
- Chung, Y.; Zhang, N.; Wooten, R.M. Borrelia burgdorferi elicited-IL-10 suppresses the production of inflammatory mediators, phagocytosis, and expression of co-stimulatory receptors by murine macrophages and/or dendritic cells. PLoS ONE 2013, 8, e84980. [Google Scholar] [CrossRef]
- Dattwyler, R.J.; Volkman, D.J.; Halperin, J.J.; Luft, B.J.; Thomas, J.; Golightly, M.G. Specific Immune Response in Lyme Borreliosis. Ann. N. Y. Acad. Sci. 1988, 539, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, R.; Araneo, B.; Ma, Y.; Yang, L.; Weis, J.J. Demonstration of a B-lymphocyte mitogen produced by the lyme disease pathogen, Borrelia burgdorferi. Infect. Immun. 1991, 60, 455–464. [Google Scholar]
- De Souza Mark, S.; Fikrig, E.; Smith, A.L.; Flavell, R.A.; Barthold, S.W. Nonspecific Proliferative Responses of Murine Lymphocytes to Borrelia burgdorferi Antigens. J. Infect. Dis. 1992, 165, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Weis, J.J. Borrelia burgdorferi outer surface lipoproteins OspA and OspB possess B- cell mitogenic and cytokine-stimulatory properties. Infect. Immun. 1993, 61, 3843–3853. [Google Scholar] [PubMed]
- Tai, K.F.; Ma, Y.; Weis, J.J. Normal human B lymphocytes and mononuclear cells respond to the mitogenic and cytokine-stimulatory activities of Borrelia burgdorferi and its lipoprotein OspA. Infect. Immun. 1994, 62, 520–528. [Google Scholar]
- Chiao, J.W.; Pavia, C.; Riley, M.; Altmann-Lasekan, W.; Abolhassani, M.; Liegner, K.; Mittelman, A. Antigens of Lyme disease of spirochaete Borrelia burgdorferi inhibits antigen or mitogen-induced lymphocyte proliferation. FEMS Immunol. Med. Microbiol. 1994, 8, 151–155. [Google Scholar] [CrossRef]
- De Souza, M.S.; Smith, A.L.; Beck, D.S.; Terwilliger, G.A.; Fikrig, E.; Barthold, S.W. Long-term study of cell-mediated responses to Borrelia burgdorferi in the laboratory mouse. Infect. Immun. 1993, 61, 1814–1822. [Google Scholar]
- Chiao, J.W.; Villalon, P.; Schwartz, I.; Wormser, G.P. Modulation of lymphocyte proliferative responses by a canine Lyme disease vaccine of recombinant outer surface protein A (OspA). FEMS Immunol. Med. Microbiol. 2000, 28, 193–196. [Google Scholar] [CrossRef]
- Dorward, D.W.; Fischer, E.R.; Brooks, D.M. Invasion and Cytopathic Killing of Human Lymphocytes by Spirochetes Causing Lyme Disease. Clin. Infect. Dis. 1997, 25, S2–S8. [Google Scholar] [CrossRef]
- Hastey, C.J.; Ochoa, J.; Olsen, K.J.; Barthold, S.W.; Baumgarth, N. MyD88- and TRIF-independent induction of Type I interferon drives naive B cell accumulation but not loss of lymph node architecture in lyme disease. Infect. Immun. 2014, 82, 1548–1558. [Google Scholar] [CrossRef]
- Tunev, S.S.; Hastey, C.J.; Hodzic, E.; Feng, S.; Barthold, S.W.; Baumgarth, N. Lymphoadenopathy during Lyme Borreliosis Is Caused by Spirochete Migration-Induced Specific B Cell Activation. PLoS Pathog. 2011, 7, e1002066–e1002114. [Google Scholar] [CrossRef] [PubMed]
- Hastey, C.J.; Elsner, R.A.; Barthold, S.W.; Baumgarth, N. Delays and Diversions Mark the Development of B Cell Responses to Borrelia burgdorferi Infection. J. Immunol. 2012, 188, 5612–5622. [Google Scholar] [CrossRef] [PubMed]
- Elsner, R.A.; Hastey, C.J.; Olsen, K.J.; Baumgarth, N. Suppression of Long-Lived Humoral Immunity Following Borrelia burgdorferi Infection. PLoS Pathog. 2015, 11, e1004976. [Google Scholar] [CrossRef] [PubMed]
- Schutzer, S.E.; Coyle, P.K.; Belman, A.L.; Golightly, M.G.; Drulle, J. Sequestration of antibody to Borrelia burgdorferi in immune complexes in seronegative Lyme disease. Lancet (London, England) 1990, 335, 312–315. [Google Scholar] [CrossRef]
- Zhong, W.; Oschmann, P.; Wellensiek, H.J. Detection and preliminary characterization of circulating immune complexes in patients with Lyme disease. Med. Microbiol. Immunol. 1997, 186, 153–158. [Google Scholar] [CrossRef]
- Brunner, M.; Sigal, L.H. Immune complexes from serum of patients with lyme disease contain Borrelia burgdorferi antigen and antigen-specific antibodies: Potential use for improved testing. J. Infect. Dis. 2000, 182, 534–539. [Google Scholar] [CrossRef]
- Schutzer, S.E.; Luan, J. Early OspA immune complex formation in animal models of Lyme disease. J. Mol. Microbiol. Biotechnol. 2003, 5, 167–171. [Google Scholar] [CrossRef]
- Brunner, M. New method for detection of Borrelia burgdorferi antigen complexed to antibody in seronegative Lyme disease. J. Immunol. Methods 2001, 249, 185–190. [Google Scholar] [CrossRef]
- Brunner, M.; Sigal, L.H. Use of serum immune complexes in a new test that accurately confirms early Lyme disease and active infection with Borrelia burgdorferi. J. Clin. Microbiol. 2001, 39, 3213–3221. [Google Scholar] [CrossRef]
- Marques, A.R.; Hornung, R.L.; Dally, L.; Philipp, M.T. Detection of immune complexes is not independent of detection of antibodies in lyme disease patients and does not confirm active infection with Borrelia burgdorferi. Clin. Diagn. Lab. Immunol. 2005, 12, 1036–1040. [Google Scholar] [CrossRef]
- Dorward, D.W.; Schwan, T.G.; Garon, C.F. Immune capture and detection of Borrelia burgdorferi antigens in urine, blood, or tissues from infected ticks, mice, dogs, and humans. J. Clin. Microbiol. 1991, 29, 1162–1170. [Google Scholar] [PubMed]
- Benitez, K.Y.V.; Soloski, M.J.; Aucott, J.N. Quantification of Borrelia burgdorferi Membrane Proteins in Human Serum: A New Concept for Detection of Bacterial Infection. Anal. Chem. 2015, 87, 11383–11388. [Google Scholar]
- Norris, S.J. vls Antigenic Variation Systems of Lyme Disease Borrelia: Eluding Host Immunity through both Random, Segmental Gene Conversion and Framework Heterogeneity. Microbiol. Spectr. 2014, 2, 1–18. [Google Scholar]
- Coburn, J.; Leong, J.; Chaconas, G. Illuminating the roles of the Borrelia burgdorferi adhesins. Trends Microbiol. 2013, 21, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.P.; Bugrysheva, J.V.; Newman, S.A. Sleeper cells: The stringent response and persistence in the Borreliella (Borrelia) burgdorferi enzootic cycle. Environ. Microbiol. 2017, 19, 3846–3862. [Google Scholar] [CrossRef] [PubMed]
- Coburn, J.; Fischer, J.R.; Leong, J.M. Solving a sticky problem: New genetic approaches to host cell adhesion by the Lyme disease spirochete. Mol. Microbiol. 2005, 57, 1182–1195. [Google Scholar] [CrossRef]
- Wolgemuth, C.W. Flagellar motility of the pathogenic spirochetes. Semin. Cell Dev. Biol. 2015, 46, 104–112. [Google Scholar] [CrossRef]
- Vig, D.K.; Wolgemuth, C.W. Spatiotemporal evolution of erythema migrans, the hallmark rash of lyme disease. Biophys. J. 2014, 106, 763–768. [Google Scholar] [CrossRef][Green Version]
- Hyde, J.A. Borrelia burgdorferi Keeps Moving and Carries on: A Review of Borrelial Dissemination and Invasion. Front. Immunol. 2017, 8, 1016–1093. [Google Scholar] [CrossRef]
- Ebady, R.; Niddam, A.F.; Boczula, A.E.; Kim, Y.R.; Gupta, N.; Tang, T.T.; Odisho, T.; Zhi, H.; Simmons, C.A.; Skare, J.T.; et al. Biomechanics of Borrelia burgdorferi Vascular Interactions. Cell Rep. 2016, 16, 2593–2604. [Google Scholar] [CrossRef]
- Moriarty, T.J.; Norman, M.U.; Colarusso, P.; Bankhead, T.; Kubes, P.; Chaconas, G. Real-time high resolution 3D imaging of the lyme disease spirochete adhering to and escaping from the vasculature of a living host. PLoS Pathog. 2008, 4, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, M.C.; Beklemisheva, A.A.; Bryksin, A.V.; Newman, S.A.; Cabello, F.C. Borrelia burgdorferi binds to, invades, and colonizes native type I collagen lattices. Infect. Immun. 2004, 72, 3138–3146. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.L.; Roemer, E.J.; Benach, J.L. Plasmin-coated Borrelia burgdorferi degrades soluble and insoluble components of the mammalian extracellular matrix. Infect. Immun. 1999, 67, 3929–3936. [Google Scholar] [PubMed]
- Grab, D.J.; Perides, G.; Dumler, J.S.; Kim, K.J.; Park, J.; Kim, Y.V.; Nikolskaia, O.; Choi, K.S.; Stins, M.F.; Kim, K.S. Borrelia burgdorferi, host-derived proteases, and the blood-brain barrier. Infect. Immun. 2005, 73, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Behera, A.K.; Durand, E.; Cugini, C.; Antonara, S.; Bourassa, L.; Hildebrand, E.; Hu, L.T.; Coburn, J. Borrelia burgdorferi BBB07 interaction with integrin alpha3beta1 stimulates production of pro-inflammatory mediators in primary human chondrocytes. Cell. Microbiol. 2008, 10, 320–331. [Google Scholar] [PubMed]
- Guo, B.P.; Norris, S.J.; Rosenberg, L.C. Adherence of Borrelia burgdorferi to the proteoglycan decorin. These include: Adherence of Borrelia burgdorferi to the Proteoglycan Decorin. Infect. Immun. 1995, 63, 3467–3472. [Google Scholar] [PubMed]
- Guo, B.P.; Brown, E.L.; Dorward, D.W.; Rosenberg, L.C.; Höök, M. Decorin-binding adhesins from Borrelia burgdorferi. Mol. Microbiol. 1998, 30, 711–723. [Google Scholar] [CrossRef]
- Suhonen, J.; Komi, J.; Soukka, J.; Lassila, O.; Viljanen, M.K. Interaction Between Borrelia burgdorferi and Immature Human Dendritic Cells. Scand. J. Immunol. 2003, 58, 67–75. [Google Scholar] [CrossRef]
- Ma, Y.; Sturrock, A.; Weis, J.J. Intracellular Localization of Borrelia burgdorferi within Human Endothelial Cells. Infect. Immun. 1991, 59, 671–678. [Google Scholar]
- Wu, J.; Weening, E.H.; Faske, J.B.; Höök, M.; Skare, J.T. Invasion of eukaryotic cells by Borrelia burgdorferi requires β1 integrins and Src kinase activity. Infect. Immun. 2011, 79, 1338. [Google Scholar] [CrossRef]
- Klempner, M.S.; Noring, R.; Rogers, R.A. Invasion of human skin fibroblasts by the Lyme disease spirochete, Borrelia burgdorferi. J. Infect. Dis. 1993, 167, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Grab, D.J.; Lanners, H.; Martin, L.N.; Chesney, J.; Cai, C.; Adkisson, H.D.; Bucala, R. Interaction of Borrelia burgdorferi with peripheral blood fibrocytes, antigen-presenting cells with the potential for connective tissue targeting. Mol. Med. 1999, 5, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Brissette, C.A.; Bykowski, T.; Cooley, A.E.; Bowman, A.; Stevenson, B. Borrelia burgdorferi RevA Antigen Binds Host Fibronectin. Infect. Immun. 2009, 77, 2802–2812. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Chen, Q.; Ritchie, J.A.; Dufour, N.P.; Fischer, J.R.; Coburn, J.; Leong, J.M. Glycosaminoglycan binding by Borrelia burgdorferi adhesin BBK32 specifically and uniquely promotes joint colonization. Cell. Microbiol. 2015, 17, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Caimano, M.; Radolf, J.D.; Leong, J.M. Adaptation of the Lyme disease spirochaete to the mammalian host environment results in enhanced glycosaminoglycan and host cell binding. Mol. Microbiol. 2003, 47, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.R.; Malawista, S.E. Entry of Borrelia burgdorferi into Macrophages Is End-On and Leads to Degradation in Lysosomes. Infect Immun. 1996, 64, 2867–2872. [Google Scholar]
- Salazar, J.C.; Duhnam-Ems, S.; La Vake, C.; Cruz, A.R.; Moore, M.W.; Caimano, M.J.; Velez-Climent, L.; Shupe, J.; Krueger, W.; Radolf, J.D. Activation of Human Monocytes by Live Borrelia burgdorferi Generates TLR2-Dependent and -Independent Responses Which Include Induction of IFN-$β$. PLoS Pathog. 2009, 5, e1000444–e1000521. [Google Scholar] [CrossRef]
- Livengood, J.A.; Gilmore, R.D., Jr. Invasion of human neuronal and glial cells by an infectious strain of Borrelia burgdorferi. Microbes Infect. 2006, 8, 2832–2840. [Google Scholar] [CrossRef]
- Williams, S.K.; Weiner, Z.P.; Gilmore, R.D. Human neuroglial cells internalize Borrelia burgdorferi by coiling phagocytosis mediated by Daam1. PLoS ONE 2018, 13, e0197413. [Google Scholar] [CrossRef]
- Coburn, J.; Barthold, S.W.; Leong, J.M. Diverse Lyme disease spirochetes bind integrin alpha IIb beta 3 on human platelets. Infect. Immun. 1994, 62, 5559–5567. [Google Scholar]
- Coburn, J.; Leong, J.M.; Erban, J.K. Integrin alpha IIb beta 3 mediates binding of the Lyme disease agent Borrelia burgdorferi to human platelets. Proc. Natl. Acad. Sci. USA 1993, 90, 7059–7063. [Google Scholar] [CrossRef] [PubMed]
- Girschick, H.J.; Huppertz, H.I.; Russmann, H.; Krenn, V.; Karch, H. Intracellular persistence of Borrelia burgdorferi in human synovial cells. Rheumatol Int. 1996, 16, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Franz, J.K.; Fritze, O.; Rittig, M.; Keyßer, G.; Priem, S.; Zacher, J.; Burmester, G.; Krause, A. Insights from a Novel Three-Dimensional In Vitro Model of Lyme Arthritis Standardized Analysis of Cellular and Molecular Interactions Between. Arthritis Rheum. 2001, 44, 151–162. [Google Scholar] [CrossRef]
- Drecktrah, D.; Lybecker, M.; Popitsch, N.; Rescheneder, P.; Hall, L.S.; Samuels, D.S. The Borrelia burgdorferi RelA/SpoT Homolog and Stringent Response Regulate Survival in the Tick Vector and Global Gene Expression during Starvation. PLoS ONE 2015, 11, e1005160. [Google Scholar] [CrossRef] [PubMed]
- Bugrysheva, J.V.; Pappas, C.J.; Terekhova, D.A.; Iyer, R.; Godfrey, H.P.; Schwartz, I.; Cabello, F.C. Characterization of the RelBbu regulon in Borrelia burgdorferi reveals modulation of glycerol metabolism by (p)ppGpp. PLoS ONE 2015, 10, e0118063. [Google Scholar] [CrossRef]
- Bugrysheva, J.V.; Bryksin, A.V.; Godfrey, H.P.; Cabello, F.C. Borrelia burgdorferi rel is responsible for generation of guanosine-3′-diphosphate-5′-triphosphate and growth control. Infect. Immun. 2005, 73, 4972–4981. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Stevenson, B.; von Lackum, K.; Wattier, R.L.; McAlister, J.D.; Miller, J.C.; Babb, K. Quorum sensing by the Lyme disease spirochete. Microbes Infect. 2003, 5, 991–997. [Google Scholar] [CrossRef]
- Babb, K.; von Lackum, K.; Wattier, R.L.; Riley, S.P.; Stevenson, B. Synthesis of Autoinducer 2 by the Lyme Disease Spirochete, Borrelia burgdorferi. J. Bacteriol. 2005, 187, 3079–3087. [Google Scholar] [CrossRef]
- Stevenson, B.; Babb, K. LuxS-Mediated Quorum Sensing in Borrelia burgdorferi, the Lyme Disease Spirochete. Infect. Immun. 2002, 70, 4099–4105. [Google Scholar] [CrossRef]
- Hübner, A.; Revel, A.T.; Nolen, D.M.; Hagman, K.E.; Norgard, M.V. Expression of a luxS gene is not required for Borrelia burgdorferi infection of mice via needle inoculation. Infect. Immun. 2003, 71, 2892–2896. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blevins, J.S.; Revel, A.T.; Caimano, M.J.; Yang, X.F.; Richardson, J.A.; Hagman, K.E.; Norgard, M.V. The luxS gene is not required for Borrelia burgdorferi tick colonization, transmission to a mammalian host, or induction of disease. Infect. Immun. 2004, 72, 4864–4867. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Arnold, W.K.; Savage, C.R.; Antonicello, A.D.; Stevenson, B. Apparent Role for Borrelia burgdorferi LuxS during Mammalian Infection. Infect. Immun. 2015, 83, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef]
- Steere, A.C.; Green, J.; Schoen, R.T.; Taylor, E.; Hutchinson, G.J.; Rahn, W.D.; Malawista, S.E. Successful parenteral penicillin therapy of established Lyme arthritis. N. Engl. J. Med. 1985, 312, 869–874. [Google Scholar] [CrossRef]
- Hollstrom, E. Successful treatment of erythema migrans Afzelius. Acta Derm. Venereol. 1951, 31, 235–243. [Google Scholar]
- Steere, A.C.; Malawista, S.E.; Newman, J.H.; Spieler, P.N.; Bartenhagen, N.H. Antibiotic therapy in Lyme disease. Ann. Intern. Med. 1980, 93, 1–8. [Google Scholar] [CrossRef]
- Johnson, S.E.; Klein, G.C.; Schmid, G.P.; Feeley, J.C. Susceptibility of the Lyme disease spirochete to seven antimicrobial agents. Yale J. Biol. Med. 1984, 57, 549–553. [Google Scholar]
- Johnson, R.C.; Kodner, C.; Russell, M. In vitro and in vivo susceptibility of the Lyme disease spirochete, Borrelia burgdorferi, to four antimicrobial agents. Antimicrob. Agents Chemother. 1987, 31, 164–167. [Google Scholar] [CrossRef]
- Mursic, V.P.; Wilske, B.; Schierz, G. In vitro and in vivo susceptibility of Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1987, 6, 424–426. [Google Scholar]
- Dever, L.L.; Jorgensen, J.H.; Barbour, A.G. In vitro antimicrobial susceptibility testing of Borrelia burgdorferi: A microdilution MIC method and time-kill studies. J. Clin. Microbiol. 1992, 30, 2692–2697. [Google Scholar] [PubMed]
- Hunfeld, K.P.; Kraiczy, P.; Kekoukh, E.; Schäfer, V.; Brade, V. Standardised in vitro susceptibility testing of Borrelia burgdorferi against well-known and newly developed antimicrobial agents—Possible implications for new therapeutic approaches to Lyme disease. Int. J. Med. Microbiol. 2002, 291, 125–137. [Google Scholar] [CrossRef]
- Feng, J.; Wang, T.; Zhang, S.; Shi, W.; Zhang, Y. An Optimized SYBR Green I/PI Assay for Rapid Viability Assessment and Antibiotic Susceptibility Testing for Borrelia burgdorferi. PLoS ONE 2014, 9, e111809. [Google Scholar] [CrossRef] [PubMed]
- Pothineni, V.R.; Wagh, D.; Babar, M.M.; Inayathullah, M.; Solow-Cordero, D.; Kim, K.M.; Samineni, A.V.; Parekh, M.B.; Tayebi, L.; Rajadas, J. Identification of new drug candidates against Borrelia burgdorferi using high-throughput screening. Drug Des. Devel. Ther. 2016, 10, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.-P.; Brade, V. Antimicrobial susceptibility of Borrelia burgdorferi sensu lato: What we know, what we don’t know, and what we need to know. Wien. Klin. Wochenschr. 2006, 118, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Terekhova, D.; Sartakova, M.L.; Wormser, G.P.; Schwartz, I.; Cabello, F.C. Erythromycin resistance in Borrelia burgdorferi. Antimicrob. Agents Chemother. 2002, 46, 3637–3640. [Google Scholar] [CrossRef] [PubMed]
- Caskey, J.R.; Embers, M.E. Persister Development by Borrelia burgdorferi Populations In Vitro. Antimicrob. Agents Chemother. 2015, 59, 6288–6295. [Google Scholar] [CrossRef]
- Trott, D.J.; Abraham, S.; Adler, B. Antimicrobial Resistance in Leptospira, Brucella, and Other Rarely Investigated Veterinary and Zoonotic Pathogens. Microbiol. Spectr. 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Jiang, S.; Gilpin, M.E.; Attia, M.; Ting, Y.L.; Berti, P.J. Lyme disease enolpyruvyl-UDP-GlcNAc synthase: Fosfomycin-resistant MurA from Borrelia burgdorferi, a fosfomycin-sensitive mutant, and the catalytic role of the active site Asp. Biochemistry 2011, 50, 2205–2212. [Google Scholar] [CrossRef]
- Bunikis, I.; Denker, K.; Östberg, Y.; Andersen, C.; Benz, R.; Bergström, S. An RND-Type Efflux System in Borrelia burgdorferi Is Involved in Virulence and Resistance to Antimicrobial Compounds. PLoS Pathog. 2008, 4, e1000009. [Google Scholar] [CrossRef]
- Samuels, D.S.; Marconi, R.T.; Huang, W.M.; Garon, C.F. gyrB mutations in coumermycin A1-resistant Borrelia burgdorferi. J. Bacteriol. 1994, 176, 3072–3075. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Samuels, D.S.; Mach, K.E.; Garon, C.F. Genetic transformation of the Lyme disease agent Borrelia burgdorferi with coumarin-resistant gyrB. J. Bacteriol. 1994, 176, 6045–6049. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.R.; Boylan, J.A.; Frye, J.G.; Gherardini, F.C. Evidence of a conjugal erythromycin resistance element in the Lyme disease spirochete Borrelia burgdorferi. Int. J. Antimicrob. Agents 2007, 30, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G. Antibiotics and Lyme disease. In Lyme Disease: Why It’s Spreading, How It Makes You Sick, and What to Do About It; Johns Hopkins University Press: Baltimore, MD, USA, 2015; pp. 140–158. ISBN 9781421417219. [Google Scholar]
- Waddell, L.A.; Greig, J.; Lindsay, L.R.; Hinckley, A.F.; Ogden, N.H. A systematic review on the impact of gestational Lyme disease in humans on the fetus and newborn. PLoS ONE 2018, 13, e0207067. [Google Scholar] [CrossRef] [PubMed]
- Dolan, M.C.; Schulze, T.L.; Jordan, R.A.; Schulze, C.J.; Ullmann, A.J.; Hojgaard, A.; Williams, M.A.; Piesman, J. Evaluation of Doxycycline-Laden Oral Bait and Topical Fipronil Delivered in a Single Bait Box to Control Ixodes scapularis (Acari: Ixodidae) and Reduce Borrelia burgdorferi and Anaplasma phagocytophilum Infection in Small Mammal Reservoirs and Host-Seekin. J. Med. Entomol. 2016, 54, tjw194. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef]
- Dewachter, L.; Fauvart, M.; Michiels, J. Bacterial Heterogeneity and Antibiotic Survival: Understanding and Combatting Persistence and Heteroresistance. Mol. Cell 2019, 76, 255–267. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef]
- Balaban, N.Q. Bacterial Persistence as a Phenotypic Switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef]
- Sharma, B.; Brown, A.V.; Matluck, N.E.; Hu, L.T.; Lewis, K. Borrelia burgdorferi, the Causative Agent of Lyme Disease, Forms Drug-Tolerant Persister Cells. Antimicrob. Agents Chemother. 2015, 59, 4616–4624. [Google Scholar] [CrossRef]
- Feng, J.; Shi, W.; Zhang, S.; Zhang, Y. Persister mechanisms in Borrelia burgdorferi: Implications for improved intervention. Emerg. Microbes Infect. 2015, 4, 1–3. [Google Scholar]
- Hodzic, E.; Feng, S.; Holden, K.; Freet, K.J.; Barthold, S.W. Persistence of Borrelia burgdorferi following antibiotic treatment in mice. Antimicrob. Agents Chemother. 2008, 52, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia burgdorferi in Rhesus Macaques following Antibiotic Treatment of Disseminated Infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Sapi, E.; Kasliwala, R.S.; Ismail, H.; Torres, J.P.; Oldakowski, M.; Markland, S.; Gaur, G.; Melillo, A.; Eisendle, K.; Liegner, K.B.; et al. The Long-Term Persistence of Borrelia burgdorferi Antigens and DNA in the Tissues of a Patient with Lyme Disease. Antibiotics 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed]
- Caskey, J.R.; Hasenkampf, N.R.; Martin, D.S.; Chouljenko, V.N.; Subramanian, R.; Cheslock, M.A.; Embers, M.E. The functional and molecular effects of doxycycline treatment on Borrelia burgdorferi phenotype. Front. Microbiol. 2019, 10, 690. [Google Scholar] [CrossRef]
- Hunfeld, K.-P.; Ruzic-Sabljic, E.; Norris, D.E.; Kraiczy, P.; Strle, F. In vitro susceptibility testing of Borrelia burgdorferi sensu lato isolates cultured from patients with erythema migrans before and after antimicrobial chemotherapy. Antimicrob. Agents Chemother. 2005, 49, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Auwaerter, P.G.; Zhang, Y. Drug combinations against Borrelia burgdorferi persisters in vitro: Eradication achieved by using daptomycin, cefoperazone and doxycycline. PLoS ONE 2015, 10, e0117207. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Kybicova, K.; Vancová, M. Metamorphoses of Lyme disease spirochetes: Phenomenon of Borrelia persisters. Parasit. Vectors 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Sapi, E.; Theophilus, P.A.S.; Pham, T.V.; Burugu, D.; Luecke, D.F. Effect of RpoN, RpoS and LuxS Pathways on the Biofilm Formation and Antibiotic Sensitivity of Borrelia burgdorferi. Eur. J. Microbiol. Immunol. (Bp.) 2016, 6, 272–286. [Google Scholar] [CrossRef]
- Díaz-Salazar, C.; Calero, P.; Espinosa-Portero, R.; Jiménez-Fernández, A.; Wirebrand, L.; Velasco-Domínguez, M.G.; López-Sánchez, A.; Shingler, V.; Govantes, F. The stringent response promotes biofilm dispersal in Pseudomonas putida. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Feng, J.; Wang, T.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. Identification of novel activity against Borrelia burgdorferi persisters using an FDA approved drug library. Emerg. Microbes Infect. 2014, 3, e49. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, S.; Shi, W.; Zhang, Y. Ceftriaxone pulse dosing fails to eradicate biofilm-like microcolony B. burgdorferi persisters which are sterilized by daptomycin/doxycycline/cefuroxime without pulse dosing. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bockenstedt, L.K.; Mao, J.; Hodzic, E.; Barthold, S.W.; Fish, D. Detection of attenuated, noninfectious spirochetes in Borrelia burgdorferi-infected mice, after antibiotic treatment. J. Infect. Dis. 2002, 186, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Kohlhepp, W.; Oschmann, P.; Mertens, H.G. Treatment of Lyme borreliosis—Randomized comparison of doxycycline and penicillin G. J. Neurol. 1989, 236, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.W. Treatment of Erythema Chronicum Migrans of Lyme disease. Ann. N. Y. Acad. Sci. 1988, 539, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Nadelman, R.B.; Luger, S.W.; Frank, E.; Wisniewski, M.; Collins, J.J.; Wormser, G.P. Comparison of cefuroxime axetil and doxycycline in the treatment of early lyme disease. Ann. Intern. Med. 1992, 117, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Kodner, C.B.; Jurkovich, P.J.; Collins, J.J. Comparative in vitro and in vivo susceptibilities of the Lyme disease spirochete Borrelia burgdorferi to cefuroxime and other antimicrobial agents. Antimicrob. Agents Chemother. 1990, 34, 2133–2136. [Google Scholar] [CrossRef]
- Eliassen, K.E.; Reiso, H.; Berild, D.; Lindbæk, M. Comparison of phenoxymethylpenicillin, amoxicillin, and doxycycline for erythema migrans in general practice. A randomized controlled trial with a 1-year follow-up. Clin. Microbiol. Infect. 2018, 24, 1290–1296. [Google Scholar] [CrossRef]
- Torbahn, G.; Hofmann, H.; Rücker, G.; Bischoff, K.; Freitag, M.H.; Dersch, R.; Fingerle, V.; Motschall, E.; Meerpohl, J.J.; Schmucker, C. Efficacy and safety of antibiotic therapy in early cutaneous Lyme borreliosis: A Network Meta-analysis. JAMA Dermatol. 2018, 154, 1292–1303. [Google Scholar] [CrossRef]
- Cerar, D.; Cerar, T.; Ružić-Sabljić, E.; Wormser, G.P.; Strle, F. Subjective Symptoms after Treatment of Early Lyme Disease. Am. J. Med. 2010, 123, 79–86. [Google Scholar] [CrossRef]
- Klempner, M.S.; Hu, L.T.; Evans, J.; Schmid, C.H.; Johnson, G.M.; Trevino, R.P.; Norton, D.; Levy, L.; Wall, D.; McCall, J.; et al. Two controlled trials of antibiotic treatment in patients with persistent symptoms and a history of Lyme disease. N. Engl. J. Med. 2001, 345, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Krupp, L.B.; Hyman, L.G.; Grimson, R.; Coyle, P.K.; Melville, P.; Ahnn, S.; Dattwyler, R.; Chandler, B. Study and treatment of post Lyme disease (STOP-LD) A randomized double masked clinical trial. Neurology 2003, 60, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.F.; Trevino, R.P.; Johnson, G.M.; Levy, L.; Dornbush, R.; Hu, L.T.; Evans, J.; Weinstein, A.; Schmid, C.H.; Klempner, M.S. Cognitive function in post-treatment Lyme disease: Do additional antibiotics help? Neurology 2003, 60, 1916–1922. [Google Scholar] [CrossRef] [PubMed]
- Fallon, B.A.; Keilp, J.G.; Corbera, K.M.; Petkova, E.; Britton, C.B.; Dwyer, E.; Slavov, I.; Cheng, J.; Dobkin, J.; Nelson, D.R.; et al. A randomized, placebo-controlled trial of repeated IV antibiotic therapy for Lyme encephalopathy. Neurology 2008, 70, 992–1003. [Google Scholar] [CrossRef]
- Fallon, B.A.; Petkova, E.; Keilp, J.G.; Britton, C.B. A reappraisal of the u.s. Clinical trials of post-treatment lyme disease syndrome. Open Neurol. J. 2012, 6, 79–87. [Google Scholar] [CrossRef]
- Johnson, L.; Shapiro, M.; Mankoff, J. Removing the Mask of Average Treatment Effects in Chronic Lyme Disease Research Using Big Data and Subgroup Analysis. Healthcare 2018, 6, 124. [Google Scholar] [CrossRef]
- Horowitz, R.I.; Freeman, P. The Use of Dapsone as a Novel “Persister” Drug in the Treatment of Chronic Lyme Disease/Post Treatment Lyme Disease Syndrome. J. Clin. Exp. Dermatology Res. 2016, 07, 1000345. [Google Scholar] [CrossRef]
- Liegner, K.B. Disulfiram (Tetraethylthiuram disulfide) in the treatment of lyme disease and babesiosis: Report of experience in three cases. Antibiotics 2019, 8, 72. [Google Scholar] [CrossRef]
- Butler, T. The Jarisch-Herxheimer reaction after antibiotic treatment of spirochetal infections: A review of recent cases and our understanding of pathogenesis. Am. J. Trop. Med. Hyg. 2017, 96, 46–52. [Google Scholar] [CrossRef]
- Negussie, Y.; Remick, D.G.; Deforge, L.E.; Kunkel, S.L.; Eynon, A.; Griffin, G.E. Detection of plasma tumor necrosis factor, interleukins 6, and 8 during the Jarisch-Herxheimer Reaction of relapsing fever. J. Exp. Med. 1992, 175, 1207–1212. [Google Scholar] [CrossRef]
- Fekade, D.; Knox, K.; Hussein, K.; Melka, A.; Lalloo, D.G.; Coxon, R.E.; Warrell, D.A. Prevention of Jarisch-Herxheimer reactions by treatment with antibodies against tumor necrosis factor alpha. N. Engl. J. Med. 1996, 335, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B.; Munford, R.S. Tumor necrosis factor and the Jarisch-Herxheimer reaction. N. Engl. J. Med. 1996, 335, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Bockenstedt, L.K.; Haberman, A.M.; Alexia, A.; Bockenstedt, L.K.; Gonzalez, D.G.; Haberman, A.M.; Belperron, A.A. Spirochete antigens persist near cartilage after murine Lyme borreliosis therapy. J. Clin. Investig. 2012, 122, 2652–2660. [Google Scholar] [CrossRef] [PubMed]
- Beck, G.; Benach, J.L.; Habicht, G.S. Isolation, preliminary chemical characterization, and biological activity of Borrelia burgdorferi peptidoglycan. Biochem. Biophys. Res. Commun. 1990, 167, 89–95. [Google Scholar] [CrossRef]
- Jutras, B.L.; Lochhead, R.B.; Kloos, Z.A.; Biboy, J.; Strle, K.; Booth, C.J.; Govers, S.K.; Gray, J.; Schumann, P.; Vollmer, W.; et al. Borrelia burgdorferi peptidoglycan is a persistent antigen in patients with Lyme arthritis. Proc. Natl. Acad. Sci. USA 2019, 116, 13498–13507. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes tick-Borne pathogens: Ecological, epidemiological, and clinical consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef]
- Nieto, N.C.; Foley, J.E. Meta-analysis of coinfection and coexposure with Borrelia burgdorferi and Anaplasma phagocytophilum in humans, domestic animals, wildlife, and Ixodes ricinus-complex ticks. Vector-Borne Zoonotic Dis. 2009, 9, 93–101. [Google Scholar] [CrossRef]
- Feng, J.; Shi, W.; Miklossy, J.; Tauxe, G.; McMeniman, C.; Zhang, Y. Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi. Antibiotics 2018, 7, 89. [Google Scholar] [CrossRef]
- Goc, A.; Niedzwiecki, A.; Rath, M. In vitro evaluation of antibacterial activity of phytochemicals and micronutrients against Borrelia burgdorferi and Borrelia garinii. J. Appl. Microbiol. 2015, 119, 1561–1572. [Google Scholar] [CrossRef]
- Socarras, K.M.; Theophilus, P.A.S.; Torres, J.P.; Gupta, K.; Sapi, E. Antimicrobial activity of bee venom and melittin against Borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef]
- Lin, Y.P.; Li, L.; Zhang, F.; Linhardt, R.J. Borrelia burgdorferi glycosaminoglycan-binding proteins: A potential target for new therapeutics against Lyme disease. Microbiology 2017, 163, 1759–1766. [Google Scholar] [CrossRef]
- Almer, J.; Gesslbauer, B.; Kungl, A.J. Therapeutic strategies to target microbial protein–glycosaminoglycan interactions. Biochem. Soc. Trans. 2018, 46, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.U.; Moriarty, T.J.; Dresser, A.R.; Millen, B.; Kubes, P.; Chaconas, G. Molecular mechanisms involved in vascular interactions of the Lyme disease pathogen in a living host. PLoS Pathog. 2008, 4, e1000169. [Google Scholar] [CrossRef] [PubMed]
- Cornell, K.A.; Primus, S.; Martinez, J.A.; Parveen, N. Assessment of methylthioadenosine/S-adenosylhomocysteine nucleosidases of Borrelia burgdorferi as targets for novel antimicrobials using a novel high-throughput method. J. Antimicrob. Chemother. 2009, 63, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Caccammo, P.D.; Brun, Y.V. The Molecular Basis of Noncanonical Bacterial Morphology. Trends Microbiol. 2018, 26, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Justice, S.S.; Hunstad, D.A.; Cegelski, L.; Hultgren, S.J. Morphological plasticity as a bacterial survival strategy. Nat. Rev. Microbiol. 2008, 6, 162–168. [Google Scholar] [CrossRef]
- Domingue, G.J.; Woody, H.B. Bacterial persistence and expression of disease. Clin. Microbiol. Rev. 1997, 10, 320–344. [Google Scholar] [CrossRef]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef]
- Umemoto, T.; Namikawa, I.; Yamamoto, M. Colonial Morphology of Treponemes Observed by Electron Microscopy. Microbiol. Immunol. 1984, 28, 11–22. [Google Scholar] [CrossRef]
- Umemoto, T.; Namikawa, I.; Yoshii, Z.; Konishi, H. An Internal View of the Spherical Body of Treponema macrodentium as Revealed by Scanning Electron Microscopy. Microbiol. Immunol. 1982, 26, 191–198. [Google Scholar] [CrossRef]
- Miklossy, J.; Kasas, S.; Zurn, A.D.; McCall, S.; Yu, S.; McGeer, P.L. Persisting atypical and cystic forms of Borrelia burgdorferi and local inflammation in Lyme neuroborreliosis. J. Neuroinflamm. 2008, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Meriläinen, L.; Herranen, A.; Schwarzbach, A.; Gilbert, L. Morphological and biochemical features of Borrelia burgdorferi pleomorphic forms. Microbiol. (United Kingdom) 2015, 161, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Vancová, M.; Rudenko, N.; Vaněček, J.; Golovchenko, M.; Strnad, M.; Rego, R.O.M.; Tichá, L.; Grubhoffer, L.; Nebesářová, J. Pleomorphism and viability of the Lyme disease pathogen Borrelia burgdorferi exposed to physiological stress conditions: A correlative cryo-fluorescence and cryo-scanning electron microscopy study. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mursic, V.P.; Wanner, G.; Reinhardt, S.; Busch, U.; Maget, W. Formation and cultivation of Borrelia burgdorferi spheroblast-L-form variants. Infection 1996, 24, 218–226. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.B. Spirochetal cyst forms in neurodegenerative disorders,... hiding in plain sight. Med. Hypotheses 2006, 67, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Margulis, L.; Maniotis, A.; MacAllister, J.; Scythes, J.; Brorson, O.; Hall, J.; Krumbein, W.E.; Chanman, M.J. Spirochete round bodies syphilis, lyme disease & AIDS: Resurgence of “the great imitator”? Symbiosis 2009, 47, 51–58. [Google Scholar]
- Brorson, O.; Brorson, S.H. In vitro conversion of Borrelia burgdorferi to cystic forms in spinal fluid, and transformation to mobile spirochetes by incubation in BSK-H medium. Infection 1998, 26, 144–150. [Google Scholar] [CrossRef]
- Brorson, O.; Brorson, S.H. An in vitro study of the susceptibility of mobile and cystic forms of Borrelia burgdorferi to hydroxychloroquine. Int. Microbiol. 2002, 5, 25–31. [Google Scholar] [CrossRef]
- Murgia, R.; Cinco, M. Induction of cystic forms by different stress conditions in Borrelia burgdorferi. APMIS 2004, 112, 57–62. [Google Scholar] [CrossRef]
- Aberer, E.; Kersten, A.; Klade, H.; Poitschek, C.; Jurecka, W. Heterogeneity of Borrelia burgdorferi in the Skin. Am. J. Dermatopathol. 1996, 18, 571–579. [Google Scholar] [CrossRef]
- Sapi, E.; Balasubramanian, K.; Poruri, A.; Maghsoudlou, J.S.; Socarras, K.M.; Timmaraju, A.V.; Filush, K.R.; Gupta, K.; Shaikh, S.; Theophilus, P.A.S.; et al. Evidence of in vivo existence of Borrelia biofilm in borrelial lymphocytomas. Eur. J. Microbiol. Immunol. 2016, 6, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Bastian, S.L.; Mpoy, C.M.; Scott, S.; Rattelle, A.; Pabbati, N.; Poruri, A.; Burugu, D.; Theophilus, P.A.S.; Pham, T.V.; et al. Characterization of Biofilm Formation by Borrelia burgdorferi In Vitro. PLoS ONE 2012, 7, e48277. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, T.; Yee, R.; Yuan, Y.; Bai, C.; Cai, M.; Shi, W.; Embers, M.; Brayton, C.; Saeki, H.; et al. Stationary Phase Persister/Biofilm Microcolony of Borrelia burgdorferi Causes More Severe Disease in a Mouse Model of Lyme Arthritis: Implications for Understanding Persistence, Post-Treatment Lyme Disease Syndrome (PTLDS), and Treatment Failure. Discov. Med. 2019, 1–11. [Google Scholar]
- Gruntar, I.; Malovrh, T.; Murgia, R.; Cinco, M. Conversion of Borrelia garinii cystic forms to motile spirochetes in vivo. Apmis 2001, 109, 383–388. [Google Scholar] [CrossRef]
- Brorson, Ø.; Brorson, S.-H. A rapid method for generating cystic forms of Borrelia burgdorferi, and their reversal to mobile spirochetes. Apmis 1998, 106, 1131–1141. [Google Scholar] [CrossRef]
- Al-Robaiy, S.; Dihazi, H.; Kacza, J.; Seeger, J.; Schiller, J.; Huster, D.; Knauer, J.; Straubinger, R.K. Metamorphosis of Borrelia burgdorferi organisms—RNA, lipid and protein composition in context with the spirochetes’ shape. J. Basic Microbiol. 2010, 50, 5–17. [Google Scholar] [CrossRef]
- Groshong, A.M.; Dey, A.; Bezsonova, I.; Caimano, M.J.; Radolf, J.D. Peptide Uptake Is Essential for Borrelia burgdorferi Viability and Involves Structural and Regulatory Complexity of its Oligopeptide Transporter. MBio 2017, 8, e02047–e02117. [Google Scholar] [CrossRef]
- Fraser, C.M.; Casjens, S.; Huang, W.M.; Sutton, G.G.; Clayton, R.; Lathigra, R.; White, O.; Ketchum, K.A.; Dodson, R.; Hickey, E.K.; et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 1997, 390, 580–586. [Google Scholar] [CrossRef]
- Boyle, W.K.; Groshong, A.M.; Drecktrah, D.; Boylan, J.A.; Gherardini, F.C.; Blevins, J.S.; Scott Samuels, D.; Bourret, T.J. DksA controls the response of the lyme disease spirochete Borrelia burgdorferi to starvation. J. Bacteriol. 2019, 201, 1–20. [Google Scholar] [CrossRef]
- Kersten, A.; Poitschek, C.; Rauch, S.; Aberer, E. Effects of penicillin, ceftriaxone, and doxycycline on morphology of Borrelia burgdorferi. Antimicrob. Agents Chemother. 1995, 39, 1127–1133. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Feng, J.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. A drug combination screen identifies drugs active against amoxicillin-induced round bodies of in vitro Borrelia burgdorferi persisters from an FDA drug library. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Levin, J.M.; Nelson, J.A.; Segreti, J.; Harrison, B.; Benson, C.A.; Strle, F. In Vitro Susceptibility of Borrelia burgdorferi to 11 Antimicrobial Agents. Antimicrob. Agents Chemother. 1993, 37, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Dunham-Ems, S.M.; Caimano, M.J.; Eggers, C.H.; Radolf, J.D. Borrelia burgdorferi Requires the Alternative Sigma Factor RpoS for Dissemination within the Vector during Tick-to-Mammal Transmission. PLoS Pathog. 2012, 8, e1002532. [Google Scholar] [CrossRef] [PubMed]
- Brorson, O.; Brorson, S.-H.; Scythes, J.; MacAllister, J.; Wier, A.; Margulis, L. Destruction of spirochete Borrelia burgdorferi round-body propagules (RBs) by the antibiotic Tigecycline. Proc. Natl. Acad. Sci. USA 2009, 106, 18656–18661. [Google Scholar] [CrossRef]
- Lantos, P.M.; Auwaerter, P.G.; Wormser, G.P. A systematic review of Borrelia burgdorferi morphologic variants does not support a role in chronic lyme disease. Clin. Infect. Dis. 2014, 58, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Weitner, M.; Shi, W.; Zhang, S.; Zhang, Y. Eradication of biofilm-like microcolony structures of Borrelia burgdorferi by daunomycin and daptomycin but not mitomycin C in combination with doxycycline and cefuroxime. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Timmaraju, V.A.; Theophilus, P.A.S.S.; Balasubramanian, K.; Shakih, S.; Luecke, D.F.; Sapi, E. Biofilm formation by Borrelia burgdorferi sensu lato. FEMS Microbiol. Lett. 2015, 362, 1–11. [Google Scholar] [CrossRef][Green Version]
- Sapi, E.; Gupta, K.; Wawrzeniak, K.; Gaur, G.; Torres, J.; Filush, K.; Melillo, A.; Zelger, B. Borrelia and Chlamydia can form mixed biofilms in infected human skin tissues. Eur. J. Microbiol. Immunol. 2019, 9, 46–55. [Google Scholar] [CrossRef]
- Flemming, H.C.; Neu, T.R.; Wozniak, D.J. The EPS matrix: The “House of Biofilm Cells”. J. Bacteriol. 2007, 189, 7945–7947. [Google Scholar] [CrossRef]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS 2013, 121, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chinese Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Kaur, N.; Anyanwu, S.; Luecke, D.F.; Datar, A.; Patel, S.; Rossi, M.; Stricker, R.B. Evaluation of in-vitro antibiotic susceptibility of different morphological forms of Borrelia burgdorferi. Infect. Drug Resist. 2011, 4, 97–113. [Google Scholar] [PubMed]
- Mickiewicz, K.M.; Kawai, Y.; Drage, L.; Gomes, M.C.; Davison, F.; Pickard, R.; Hall, J.; Mostowy, S.; Aldridge, P.D.; Errington, J. Possible role of L-form switching in recurrent urinary tract infection. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Schutzer, S.E.; Body, B.A.; Boyle, J.; Branson, B.M.; Dattwyler, R.J.; Fikrig, E.; Gerald, N.J.; Gomes-Solecki, M.; Kintrup, M.; Ledizet, M.; et al. Direct Diagnostic Tests for Lyme Disease. Clin. Infect. Dis. 2019, 68, 1052–1057. [Google Scholar] [CrossRef]
- Auwaerter, P.G.; Bakken, J.S.; Dattwyler, R.J.; Dumler, J.S.; Halperin, J.J.; McSweegan, E.; Nadelman, R.B.; O’Connell, S.; Shapiro, E.D.; Sood, S.K.; et al. Antiscience and ethical concerns associated with advocacy of Lyme disease. Lancet. Infect. Dis. 2011, 11, 713–719. [Google Scholar] [CrossRef]
- Arvikar, S.L.; Steere, A.C. Diagnosis and Treatment of Lyme Arthritis. Infect. Dis. Clin. N. Am. 2015, 29, 269–280. [Google Scholar] [CrossRef]
- Johnson, L.; Wilcox, S.; Mankoff, J.; Stricker, R.B. Severity of chronic Lyme disease compared to other chronic conditions: A quality of life survey. PeerJ 2014, 2, e322. [Google Scholar] [CrossRef]
- Chan, K.; Awan, M.; Barthold, S.W.; Parveen, N. Comparative molecular analyses of Borrelia burgdorferi sensu stricto strains B31 and N40D10/E9 and determination of their pathogenicity. BMC Microbiol. 2012, 12, 157. [Google Scholar] [CrossRef]
- Carpi, G.; Walter, K.S.; Bent, S.J.; Hoen, A.G.; Diuk-Wasser, M.; Caccone, A. Whole genome capture of vector-borne pathogens from mixed DNA samples: A case study of Borrelia burgdorferi. BMC Genom. 2015, 16, 1–11. [Google Scholar] [CrossRef][Green Version]
- Alban, P.S.; Johnson, P.W.; Nelson, D.R. Serum-starvation-induced changes in protein synthesis and morphology of Borrelia burgdorferi. Microbiology 2000, 146, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Brorson, O.; Brorson, S.H. Transformation of cystic forms of Borrelia burgdorferi to normal, mobile spirochetes. Infection 1997, 25, 240–246. [Google Scholar] [CrossRef] [PubMed]






| Disease Classification | CDC Surveillance Definition | |
|---|---|---|
| Early | Confirmed | EM + exposure in high incidence region (if exposure occurs in low incidence state or is unknown, laboratory evidence * is required) |
| Probable | Physician diagnosed Lyme disease with laboratory evidence * | |
| Suspected | Case of EM with no known exposure & no laboratory evidence * | |
| Late | Objective signs of specific organ damage + concurrent laboratory evidence * | |
| Stage | Description | Microbiology | References |
|---|---|---|---|
| Early Localized Lyme Disease 7–14 days after inoculation | - Can present with a single EM lesion (~70% of patients) - Can present with mild flu-like symptoms (fatigue, headache, myalgias, arthralgias and fever) | Localized Borrelia infection | [64,65,66] |
| Early Disseminated Lyme Disease Days to months | - Can present as multiple EM lesions, acute neurologic disease, Lyme carditis, borrelial lymphocytoma, and/or systemic symptoms | Borrelia enters the bloodstream and disseminates systemically | [64,65,66,67] |
| Late Disseminated Lyme Disease Months to years (untreated) | - Intermittent or ongoing objective signs of specific organ damage to joints (arthritis), heart (Lyme carditis), nervous system (encephalitis, polyneuropathy), and/or skin (acrodermatitis chronica atrophicans and lymphocytoma) | Ongoing Borrelia infection of tertiary organ sites | [66,68,69] |
| Post-Treatment Lyme Disease Syndrome (PTLDS) (previously referred to as Post Lyme Disease Syndrome/PLDS) | - Previous objective evidence of infection - Treated with antibiotics followed by resolution of objective signs - Onset of subjective symptoms (fatigue, widespread musculoskeletal pain and cognitive difficulties—see below for more information) within 6 months of treatment, that persists for at least 6 months | Borrelia infection that has been treated with antibiotics. There are several hypotheses for the cause of ongoing symptoms | [54,66] |
| Chronic Lyme Disease (CLD) | - Ongoing symptoms consistent with Lyme disease (fatigue, arthralgias, myalgias, nervous system dysfunction) not meeting strict definitions of another category | Topic of controversy | [67,70] |
| Host Material | Host–Borrelia Interaction | Methods of Analysis | Citations |
|---|---|---|---|
| Chondrocytes | Adherence | Protein interaction analyses | (Behera et al., 2008) [206] |
| Collagen Matrix | Adherence and Sequestered | Adherence assay, BacLight | (Zambrano et al., 2004) [203] |
| Decorin (ECM proteoglycan) | Adherence | ELISA, IF, Protein interaction analyses | (Guo et al., 1995) (Guo et al., 1998) [207,208] |
| Dendritic Cells | Internalized | Darkfield microscopy, CLSM, BacLight | (Suhonen et al., 2003) [209] |
| Endothelial Cells | Adherence and Internalized | IF | (Ma et al., 1991) (Wu et al., 2011) [210,211] |
| Fibroblasts | Internalized | IF, Antibiotic Challenge & Re-Growth, RT-PCR | (Wu et al., 2011) (Klempner et al., 1993) [211,212] |
| Fibrocytes | Held in membrane invaginations | Light and electron microscopy | (Grab et al., 1999) [213] |
| Fibronectin Matrix | Adherence | Darkfield microscopy and ELISA | (Brissette et al., 2009) [214] |
| ECM Glycosaminoglycans | Adherence | Protein interaction analyses and liquid scintillation, IF | (Lin et al., 2015) (Parveen et al., 2003) [215,216] |
| Laminin Matrix | Adherence | ELISA | (Brissette et al., 2009) [214] |
| Lymphocytes | Internalized | Light microscopy, TEM, SEM | (Dorward et al., 1997) [180] |
| Macrophages | Internalized | CLSM, Re-Culture | (Montgomery et al., 1996) [217] |
| Monocytes | Internalized | Cytokine response analyses, CLSM | (Salazar et al., 2009) [218] |
| Neuronal and Glial Cells | Internalized | IF, Antibiotic Challenge | (Livengood et al., 2006) (Williams et al., 2018) [219,220] |
| Platelets | Adherence | IF | (Coburn et al., 1994) (Coburn et al., 1993) [221,222] |
| Synovial Cells | Internalized | CLSM and TEM, IHC on 3D model, Antibiotic challenge | (Girschick et al., 1996) (Franz et al., 2001) [223,224] |
| Term | Defining Features | Documentation | Verdict |
|---|---|---|---|
| Cyst | Encasement of a vegetative spirochete within a defined outer membrane. | (Miklossy et al., 2008) [313] | Encasement within an outer membrane reported to occur. Defining features of outer envelope/membrane unclear whether it meets criteria for cyst. |
| Cell Wall Deficient (CWD) Form/L-form/Spheroplast/Protoplast | Absence, or partial absence, of the cell wall. Absence of murein. | (Mursic et al., 1996) [316] | Does not seem to be the case as studies show the presence of peptidoglycan in the outer envelope/membrane of the round body. |
| Biofilm | Structurally rearranging aggregates within a matrix composed of extracellular polymeric substances (EPS) that attaches to biotic and abiotic surfaces. | (Sapi et al., 2012) [325] | Clusters and aggregates can be seen. Growth on collagen, fibronectin, and agarose confirmed. Alginate, calcium, and eDNA present in matrix. |
| Microcolony/Aggregate | Assemblies of spirochetes that are either adherent to a solid surface or free-floating within liquid. May include bundles or radial clusters. | (Feng et al., 2019) [326] | Evident in culture. Evidence in animal and clinical models unclear. |
| Spherule/Coccoid Form/Round Body | Spirochetes that have adopted a rounded, spherical morphology while retaining an intact outer membrane. | (Meriläinen et al., 2015) [314] | Evident in culture. PI staining indicates that the outer envelope/membrane is not intact. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamm, V.V.; Ko, J.T.; Mainprize, I.L.; Sanderson, V.P.; Wills, M.K.B. Lyme Disease Frontiers: Reconciling Borrelia Biology and Clinical Conundrums. Pathogens 2019, 8, 299. https://doi.org/10.3390/pathogens8040299
Bamm VV, Ko JT, Mainprize IL, Sanderson VP, Wills MKB. Lyme Disease Frontiers: Reconciling Borrelia Biology and Clinical Conundrums. Pathogens. 2019; 8(4):299. https://doi.org/10.3390/pathogens8040299
Chicago/Turabian StyleBamm, Vladimir V., Jordan T. Ko, Iain L. Mainprize, Victoria P. Sanderson, and Melanie K. B. Wills. 2019. "Lyme Disease Frontiers: Reconciling Borrelia Biology and Clinical Conundrums" Pathogens 8, no. 4: 299. https://doi.org/10.3390/pathogens8040299
APA StyleBamm, V. V., Ko, J. T., Mainprize, I. L., Sanderson, V. P., & Wills, M. K. B. (2019). Lyme Disease Frontiers: Reconciling Borrelia Biology and Clinical Conundrums. Pathogens, 8(4), 299. https://doi.org/10.3390/pathogens8040299