Genetic Diversity among Pseudorabies Viruses Isolated from Dogs in France from 2006 to 2018


Abstract
1. Introduction
2. Results
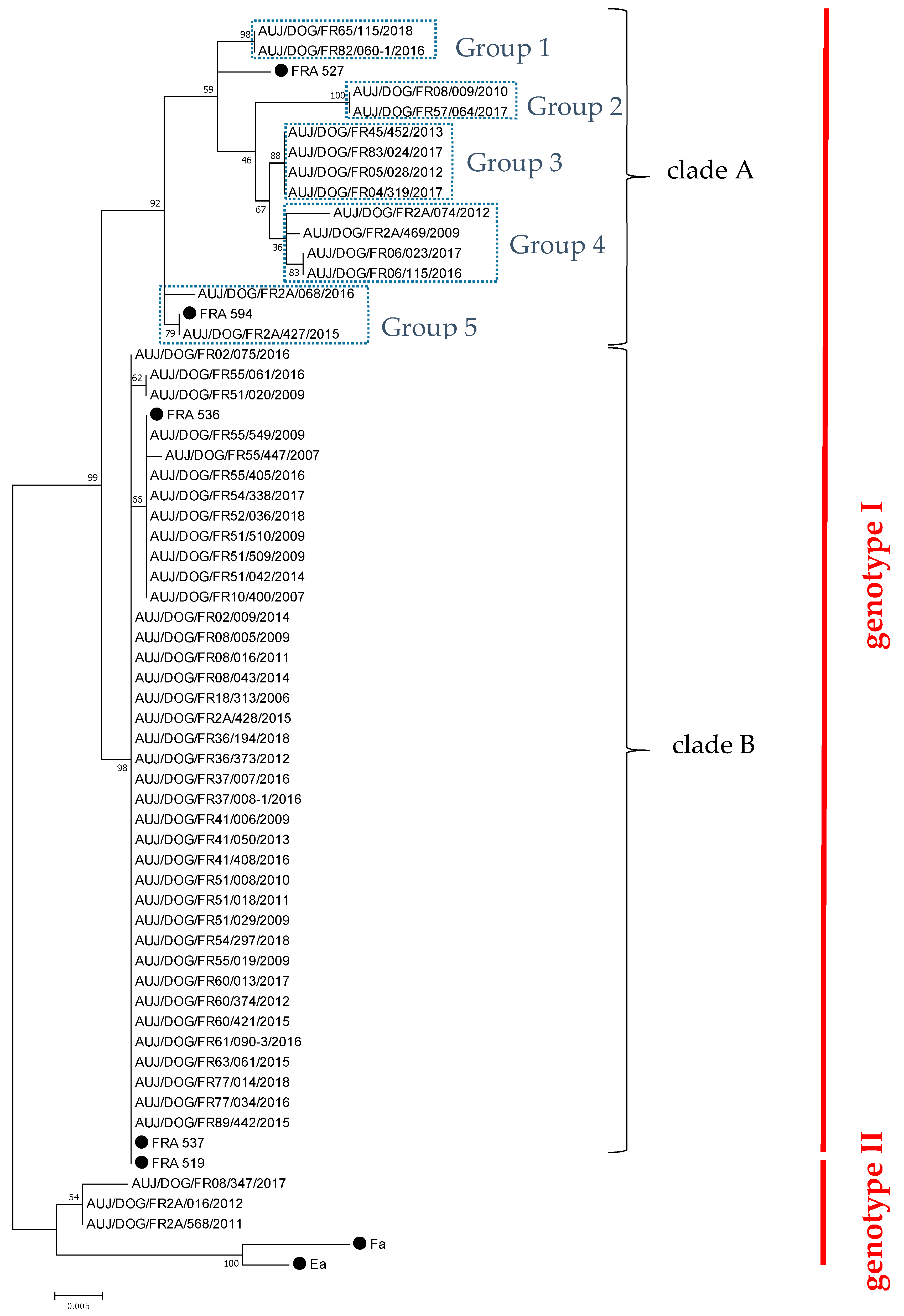
2.1. Characterisation of French Canine PRV Strains
2.2. Geographical Distribution of PRV Strains Isolated from Dogs in France from 2006 to 2018
2.3. Comparison of French Canine PRV Strains with PRV Strains Isolated in Other Countries
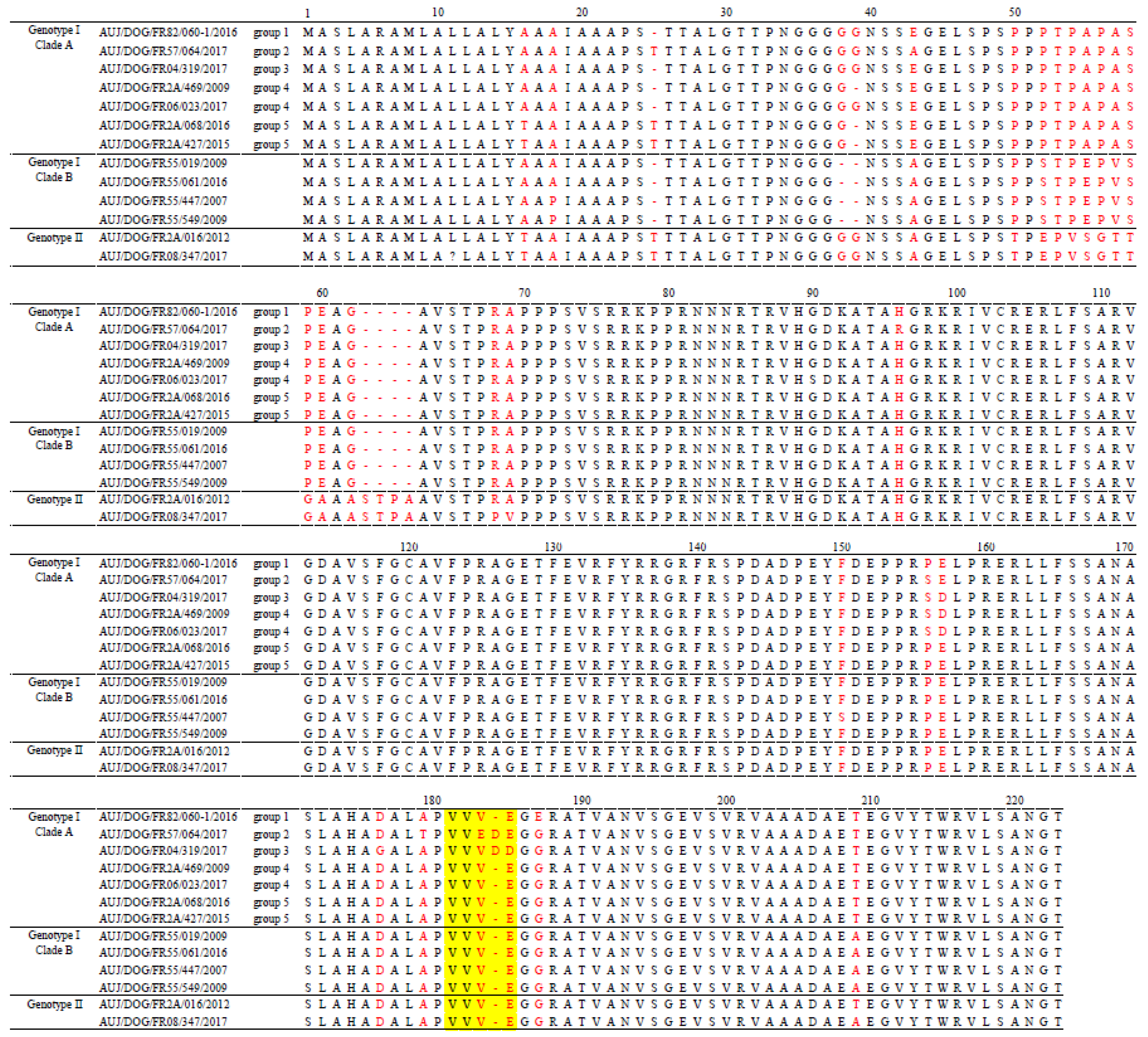
2.4. Analysis of the Translated Protein Sequences of French Canine PRV Strains
3. Discussion
4. Material and Methods
4.1. PRV Detection and Isolation from Dog Samples
4.2. Sequencing of the gC-Encoding Gene
4.3. Phylogenetic Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mettenleiter, T.C.; Ehlers, B.; Müller, T.; Yoon, K.-J.; Teifke, J.P. Herpesviruses. In Diseases of Swine; Wiley Blackwell: Hoboken, NJ, USA, 2019; pp. 548–575. [Google Scholar]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed]
- Klupp, B.G.; Hengartner, C.J.; Mettenleiter, T.C.; Enquist, L.W. Complete, Annotated Sequence of the Pseudorabies Virus Genome. J. Virol. 2004, 78, 424–440. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, T.L.; Weigel, R.M.; Hahn, E.C.; Scherba, G. Comparative utility of restriction fragment length polymorphism analysis and gene sequencing to the molecular epidemiological investigation of a viral outbreak. Epidemiol. Infect. 2001, 126, 415–424. [Google Scholar] [CrossRef] [PubMed][Green Version]
- He, W.; Auclert, L.Z.; Zhai, X.; Wong, G.; Zhang, C.; Zhu, H.; Xing, G.; Wang, S.; He, W.; Li, K.; et al. Interspecies Transmission, Genetic Diversity, and Evolutionary Dynamics of Pseudorabies Virus. J. Infect. Dis. 2019, 219, 1705–1715. [Google Scholar] [CrossRef]
- Ye, C.; Guo, J.C.; Gao, J.C.; Wang, T.Y.; Zhao, K.; Chang, X.B.; Wang, Q.; Peng, J.M.; Tian, Z.J.; Cai, X.H.; et al. Genomic analyses reveal that partial sequence of an earlier pseudorabies virus in China is originated from a Bartha-vaccine-like strain. Virology 2016, 491, 56–63. [Google Scholar] [CrossRef]
- Ye, C.; Zhang, Q.Z.; Tian, Z.J.; Zheng, H.; Zhao, K.; Liu, F.; Guo, J.C.; Tong, W.; Jiang, C.G.; Wang, S.J.; et al. Genomic characterization of emergent pseudorabies virus in China reveals marked sequence divergence: Evidence for the existence of two major genotypes. Virology 2015, 483, 32–43. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, W.; Liu, Q.; Zhao, T.; Zhu, H.; Hua, L.; Peng, Z.; Tang, X.; Stratton, C.W.; Zhou, D.; et al. Epidemiological and genetic characteristics of swine pseudorabies virus in mainland China between 2012 and 2017. PeerJ 2018, 6, e5785. [Google Scholar] [CrossRef]
- Muller, T.; Hahn, E.C.; Tottewitz, F.; Kramer, M.; Klupp, B.G.; Mettenleiter, T.C.; Freuling, C. Pseudorabies virus in wild swine: A global perspective. Arch. Virol. 2011, 156, 1691–1705. [Google Scholar] [CrossRef]
- Muller, T.; Klupp, B.G.; Freuling, C.; Hoffmann, B.; Mojcicz, M.; Capua, I.; Palfi, V.; Toma, B.; Lutz, W.; Ruiz-Fon, F.; et al. Characterization of pseudorabies virus of wild boar origin from Europe. Epidemiol. Infect. 2010, 138, 1590–1600. [Google Scholar] [CrossRef]
- Verpoest, S.; Cay, A.B.; De Regge, N. Molecular characterization of Belgian pseudorabies virus isolates from domestic swine and wild boar. Vet. Microbiol. 2014, 172, 72–77. [Google Scholar] [CrossRef]
- Fonseca, A.A., Jr.; Camargos, M.F.; Barbosa, A.A.; Goncalves, V.L.; Heinemann, M.B.; Reis, J.K. Evolutionary Diversity of Suid Herpesvirus 1 Based on Ul44 Partial Sequences. Intervirology 2016, 59, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Sozzi, E.; Grilli, G.; Gibelli, L.R.; Gelmetti, D.; Lelli, D.; Chiari, M.; Prati, P.; Alborali, G.L.; Boniotti, M.B.; et al. Detection and molecular analysis of Pseudorabies virus strains isolated from dogs and a wild boar in Italy. Vet. Microbiol. 2015, 177, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. 2008/185/EC: Commission Decision of 21 February 2008 on additional guarantees in intra-Community trade of pigs relating to Aujeszky’s disease and criteria to provide information on this disease. Off. J. Eur. Union 2008, L59, 19–30. [Google Scholar]
- Anonymous. 2008/269/EC: Commission Decision of 19 March 2008 amending Decision 2001/618/EC to include the departments of Côtes-d’Armor, Finistère, Ille-et-Vilaine, Morbihan and Nord, France, in the list of regions free of Aujeszky’s disease. Off. J. Eur. Union 2008, L85, 9–10. [Google Scholar]
- ONCFS. Wild Ungulates Hunting Tables 2017–2018; French Agency for Wildlife and Hunting (ONCFS): Paris, France, 2018; Available online: http://www.oncfs.gouv.fr/IMG/file/publications/revue%20faune%20sauvage/FS-320-ENCART-tableauxchasseongules.pdf (accessed on 25 November 2019).
- Albina, E.; Mesplède, A.; Chenut, G.; Le Potier, M.F.; Bourbao, G.; Le Gal, S.; Leforban, Y. A serological survey on classical swine fever (CSF), Aujeszky’s disease (AD) and porcine reproductive and respiratory syndrome (PRRS) virus infections in French wild boars from 1991 to 1998. Vet. Microbiol. 2000, 77, 43–57. [Google Scholar] [CrossRef]
- Rossi, S.; Hars, J.; Garin-Bastuji, B.; Le Potier, M.-F.; Boireau, P.; Aubry, P.; Hattenberger, A.-M.; Louguet, Y.; Toma, B.; Boué, F. Résultats de l’enquête nationale sérologique menée chez le sanglier sauvage (2000–2004). Bull. Epidémiol. 2008, 29, 5–7. [Google Scholar]
- Payne, A.; Rossi, S.; Lacour, S.; Vallée, I.; Garin-Bastuji, B.; Simon, G.; Hervé, S.; Pavio, N.; Richomme, C.; Dunoyer, C. Health report on wild boar focusing on trichinosis, Aujeszky’s disease, brucellosis, hepatitis Eandswine influenzaviruses in France. Bull. Épidémiol. Santé Anim. Aliment. 2011, 44, 2–8. [Google Scholar]
- Charrier, F.; Rossi, S.; Jori, F.; Maestrini, O.; Richomme, C.; Casabianca, F.; Ducrot, C.; Jouve, J.; Pavio, N.; Le Potier, M.F. Aujeszky’s Disease and Hepatitis E Viruses Transmission between Domestic Pigs and Wild Boars in Corsica: Evaluating the Importance of Wild/Domestic Interactions and the Efficacy of Management Measures. Front. Vet. Sci. 2018, 5, 1. [Google Scholar] [CrossRef]
- Marcé, C.; Bronner, A.; Fradin, N.; Rose, N.; Simon, G.; Pol, F.; Le Potier, M.-F. Report on surveillance ofAujeszky’s disease in France in 2010: Detection of outbreaks in outdoor units. Bull. Épidémiol. Santé Anim. Aliment. Anses-DGAl 2011, 46, 41–42. [Google Scholar]
- OIE. Event Report; World Animal Health Interface: Paris, France, 2018; Available online: https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=26292 (accessed on 25 November 2019).
- OIE. Event Report; World Animal Health Interface: Paris, France, 2019; Available online: http://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=30260 (accessed on 25 November 2019).
- Marcé, C.; Deblanc, C.; Oger, A.; Bourry, O.; Simon, G.; Rose, N.; Le Potier, M.-F. Upholding of Aujeszky’s disease-free status in 2014: Improvement of detection in high risk pig herds but decrease in field player vigilance. Bull. Épidémiol. Santé Anim. Aliment. 2015, 71, 50–53. [Google Scholar]
- Marcé, C.; Deblanc, C.; Oger, A.; Bourry, O.; Simon, G.; Rose, N.; Le Potier, M.-F. Review of surveillance of Aujszky’s disease in France in 2013: Mainland France continues to benefit from Aujeszky’s disease-free status. Bull. Épidémiol. Santé Anim. Aliment. 2014, 64, 45–48. [Google Scholar]
- Wendling, S.; Deblanc, C.; Oger, A.; Bourry, O.; Simon, G.; Rose, N.; Le Potier, M.-F. Review of surveillance of Aujeszky’s disease in France in 2015: Upholding of disease-free status for Aujeszky’s disease in mainland France and La Réunion Island. Bull. Epidémiol. Santé Anim. Aliment. 2018, 83, 1–4. [Google Scholar]
- Toma, B.; Dufour, B. Transmission de la maladie d’Aujeszky des sangliers sauvages aux suidés domestiques. Epidémiol. Santé Anim. 2004, 45, 115–119. [Google Scholar]
- Fonseca, A.A., Jr.; Camargos, M.F.; de Oliveira, A.M.; Ciacci-Zanella, J.R.; Patricio, M.A.; Braga, A.C.; Cunha, E.S.; D’Ambros, R.; Heinemann, M.B.; Leite, R.C.; et al. Molecular epidemiology of Brazilian pseudorabies viral isolates. Vet. Microbiol. 2010, 141, 238–245. [Google Scholar] [CrossRef]
- Fonseca, A.A., Jr.; Camargos, M.F.; Sales, M.L.; Heinemann, M.B.; Leite, R.C.; Reis, J.K. Pseudorabies virus can be classified into five genotypes using partial sequences of UL44. Braz. J. Microbiol. 2012, 43, 1632–1640. [Google Scholar] [CrossRef]
- Sozzi, E.; Moreno, A.; Lelli, D.; Cinotti, S.; Alborali, G.L.; Nigrelli, A.; Luppi, A.; Bresaola, M.; Catella, A.; Cordioli, P. Genomic characterization of pseudorabies virus strains isolated in Italy. Transbound. Emerg. Dis. 2014, 61, 334–340. [Google Scholar] [CrossRef]
- Muller, T.F.; Teuffert, J.; Zellmer, R.; Conraths, F.J. Experimental infection of European wild boars and domestic pigs with pseudorabies viruses with differing virulence. Am. J. Vet. Res. 2001, 62, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Hahn, E.C.; Page, G.R.; Hahn, P.S.; Gillis, K.D.; Romero, C.; Annelli, J.A.; Gibbs, E.P. Mechanisms of transmission of Aujeszky’s disease virus originating from feral swine in the USA. Vet. Microbiol. 1997, 55, 123–130. [Google Scholar] [CrossRef]
- Keros, T.; Brnic, D.; Prpic, J.; Dezdek, D.; Jemersic, L.; Roic, B.; Bedekovic, T. Characterisation of pseudorabies virus in domestic pigs and wild boars in Croatia. Acta Vet. Hung. 2014, 62, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Milicevic, V.; Radojicic, S.; Valcic, M.; Ivovic, V.; Radosavljevic, V. Evidence of Aujeszky’s disease in wild boar in Serbia. BMC Vet. Res. 2016, 12, 134. [Google Scholar] [CrossRef]
- Steinrigl, A.; Revilla-Fernandez, S.; Kolodziejek, J.; Wodak, E.; Bago, Z.; Nowotny, N.; Schmoll, F.; Kofer, J. Detection and molecular characterization of Suid herpesvirus type 1 in Austrian wild boar and hunting dogs. Vet. Microbiol. 2012, 157, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Verpoest, S.; Cay, A.B.; Van Campe, W.; Mostin, L.; Welby, S.; Favoreel, H.; De Regge, N. Age- and strain-dependent differences in the outcome of experimental infections of domestic pigs with wild boar pseudorabies virus isolates. J. Gen. Virol. 2016, 97, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Gu, Z.; Jin, L.; Lv, L.; Wang, J.; Sun, T.; Bai, J.; Sun, H.; Wang, X.; Jiang, P. Polymorphisms affecting the gE and gI proteins partly contribute to the virulence of a newly-emergent highly virulent Chinese pseudorabies virus. Virology 2018, 519, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Hars, J.; Rossi, S. Évaluation des risques sanitaires liés à l’augmentation des effectifs de sangliers en France. Revue ONCFS Faune Sauvag. 2010, 288, 23–28. [Google Scholar]
- OIE. Aujeszky’s Disease; Springer: Paris, France, 2018; pp. 321–337. [Google Scholar]
- Pol, F.; Deblanc, C.; Oger, A.; Le Dimna, M.; Simon, G.; Le Potier, M.F. Validation of a commercial real-time PCR kit for specific and sensitive detection of Pseudorabies. J. Virol. Methods 2013, 187, 421–423. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Isolate ID | Location (Name of French Administrative Département) | gC Genotype | Clade | GenBank Accession Number |
|---|---|---|---|---|---|
| 2006 | AUJ/DOG/FR18/313/2006 | Cher | I | B | MN590185 |
| 2007 | AUJ/DOG/FR10/400/2007 | Aube | I | B | MN590184 |
| AUJ/DOG/FR55/447/2007 | Meuse | I | B | MN590214 | |
| 2009 | AUJ/DOG/FR08/005/2009 | Ardennes | I | B | MN590179 |
| AUJ/DOG/FR41/006/2009 | Loir-et-Cher | I | B | MN590197 | |
| AUJ/DOG/FR55/019/2009 | Meuse | I | B | MN590211 | |
| AUJ/DOG/FR51/020/2009 | Marne | I | B | MN590203 | |
| AUJ/DOG/FR51/029/2009 | Marne | I | B | MN590204 | |
| AUJ/DOG/FR2A/469/2009 | Corse-du-Sud | I | A | MN590191 | |
| AUJ/DOG/FR51/509/2009 | Marne | I | B | MN590206 | |
| AUJ/DOG/FR51/510/2009 | Marne | I | B | MN590207 | |
| AUJ/DOG/FR55/549/2009 | Meuse | I | B | MN590215 | |
| 2010 | AUJ/DOG/FR51/008/2010 | Marne | I | B | MN590201 |
| AUJ/DOG/FR08/009/2010 * | Ardennes | I | A | MN590180 | |
| 2011 | AUJ/DOG/FR08/016/2011 | Ardennes | I | B | MN590181 |
| AUJ/DOG/FR51/018/2011 | Marne | I | B | MN590202 | |
| AUJ/DOG/FR2A/568/2011 | Corse-du-Sud | II | MN590192 | ||
| 2012 | AUJ/DOG/FR2A/016/2012 | Corse-du-Sud | II | MN590186 | |
| AUJ/DOG/FR05/028/2012 | Hautes-Alpes | I | A | MN590176 | |
| AUJ/DOG/FR2A/074/2012 | Corse-du-Sud | I | A | MN590188 | |
| AUJ/DOG/FR36/373/2012 | Indre | I | B | MN590194 | |
| AUJ/DOG/FR60/374/2012 | Oise | I | B | MN590218 | |
| 2013 | AUJ/DOG/FR41/050/2013 | Loir-et-Cher | I | B | MN590198 |
| AUJ/DOG/FR45/452/2013 | Loiret | I | A | MN590200 | |
| 2014 | AUJ/DOG/FR02/009/2014 | Aisne | I | B | MN590173 |
| AUJ/DOG/FR51/042/2014 | Marne | I | B | MN590205 | |
| AUJ/DOG/FR08/043/2014 | Ardennes | I | B | MN590182 | |
| 2015 | AUJ/DOG/FR63/061/2015 | Puy-de-Dôme | I | B | MN590221 |
| AUJ/DOG/FR60/421/2015 | Oise | I | B | MN590219 | |
| AUJ/DOG/FR2A/427/2015 | Corse-du-Sud | I | A | MN590189 | |
| AUJ/DOG/FR2A/428/2015 | Corse-du-Sud | I | B | MN590190 | |
| AUJ/DOG/FR89/442/2015 | Yonne | I | B | MN590227 | |
| 2016 | AUJ/DOG/FR37/007/2016 | Indre-et-Loire | I | B | MN590195 |
| AUJ/DOG/FR37/008-1/2016 | Indre-et-Loire | I | B | MN590196 | |
| AUJ/DOG/FR77/034/2016 | Seine-et-Marne | I | B | MN590224 | |
| AUJ/DOG/FR82/060-1/2016 | Tarn-et-Garonne | I | A | MN590225 | |
| AUJ/DOG/FR55/061/2016 | Meuse | I | B | MN590212 | |
| AUJ/DOG/FR2A/068/2016 | Corse-du-Sud | I | A | MN590187 | |
| AUJ/DOG/FR02/075/2016 | Aisne | I | B | MN590174 | |
| AUJ/DOG/FR61/090-3/2016 | Orne | I | B | MN590220 | |
| AUJ/DOG/FR06/115/2016 | Alpes-Maritimes | I | A | MN590178 | |
| AUJ/DOG/FR55/405/2016 | Meuse | I | B | MN590213 | |
| AUJ/DOG/FR41/408/2016 | Loir-et-Cher | I | B | MN590199 | |
| 2017 | AUJ/DOG/FR60/013/2017 | Oise | I | B | MN590217 |
| AUJ/DOG/FR06/023/2017 | Alpes-Maritimes | I | A | MN590177 | |
| AUJ/DOG/FR83/024/2017 | Var | I | A | MN590226 | |
| AUJ/DOG/FR57/064/2017 | Moselle | I | A | MN590216 | |
| AUJ/DOG/FR04/319/2017 | Alpes-de-Haute-Provence | I | A | MN590175 | |
| AUJ/DOG/FR54/338/2017 | Meurthe-et-Moselle | I | B | MN590210 | |
| AUJ/DOG/FR08/347/2017 | Ardennes | II | MN590183 | ||
| 2018 | AUJ/DOG/FR77/014/2018 | Seine-et-Marne | I | B | MN590223 |
| AUJ/DOG/FR52/036/2018 | Haute-Marne | I | B | MN590208 | |
| AUJ/DOG/FR65/115/2018 | Hautes-Pyrénées | I | A | MN590222 | |
| AUJ/DOG/FR36/194/2018 | Indre | I | B | MN590193 | |
| AUJ/DOG/FR54/297/2018 * | Meurthe-et-Moselle | I | B | MN590209 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deblanc, C.; Oger, A.; Simon, G.; Le Potier, M.-F. Genetic Diversity among Pseudorabies Viruses Isolated from Dogs in France from 2006 to 2018. Pathogens 2019, 8, 266. https://doi.org/10.3390/pathogens8040266
Deblanc C, Oger A, Simon G, Le Potier M-F. Genetic Diversity among Pseudorabies Viruses Isolated from Dogs in France from 2006 to 2018. Pathogens. 2019; 8(4):266. https://doi.org/10.3390/pathogens8040266
Chicago/Turabian StyleDeblanc, Céline, Aurélie Oger, Gaëlle Simon, and Marie-Frédérique Le Potier. 2019. "Genetic Diversity among Pseudorabies Viruses Isolated from Dogs in France from 2006 to 2018" Pathogens 8, no. 4: 266. https://doi.org/10.3390/pathogens8040266
APA StyleDeblanc, C., Oger, A., Simon, G., & Le Potier, M.-F. (2019). Genetic Diversity among Pseudorabies Viruses Isolated from Dogs in France from 2006 to 2018. Pathogens, 8(4), 266. https://doi.org/10.3390/pathogens8040266