A GntR Family Transcription Factor (VPA1701) for Swarming Motility and Colonization of Vibrio parahaemolyticus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
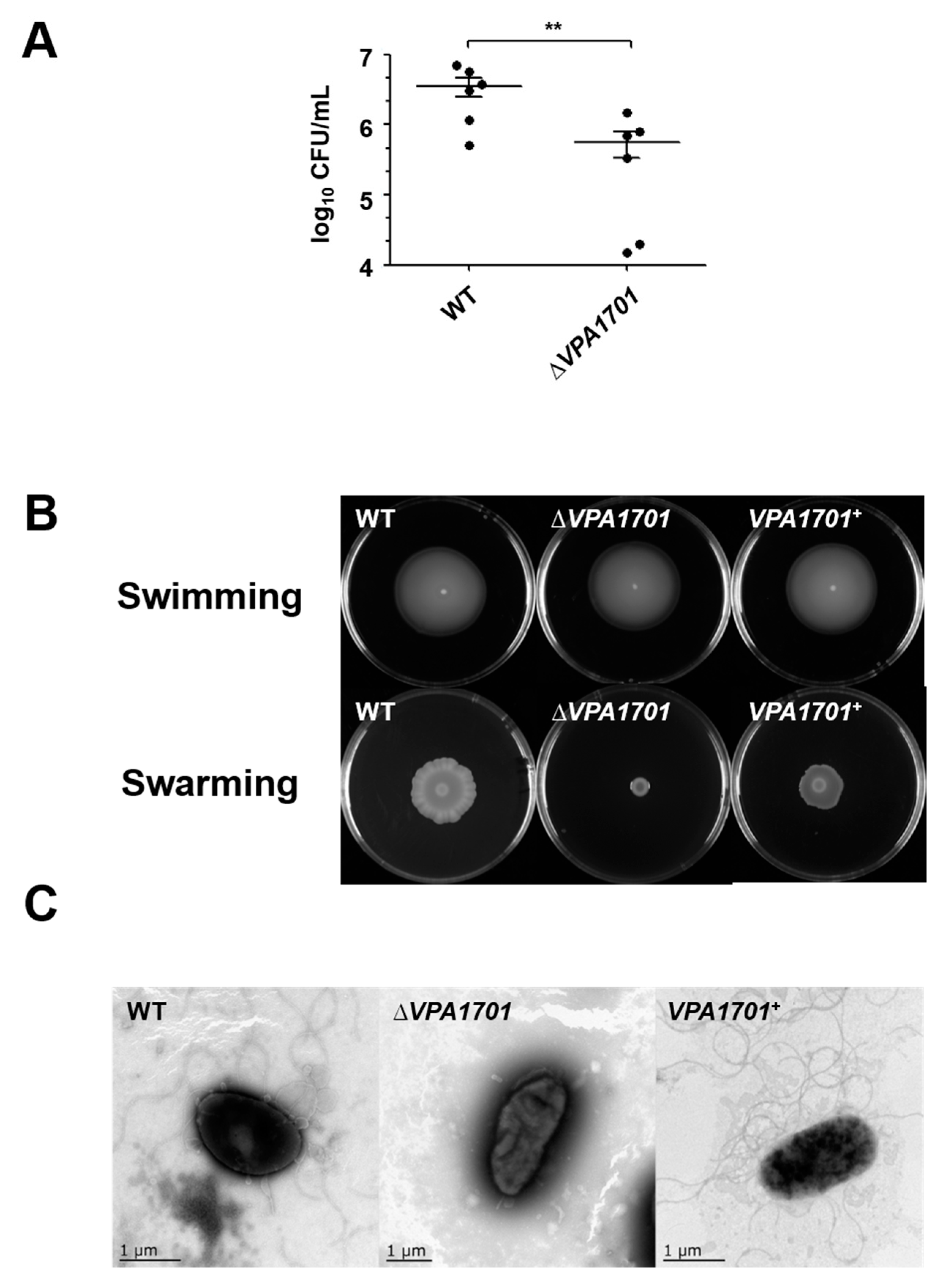
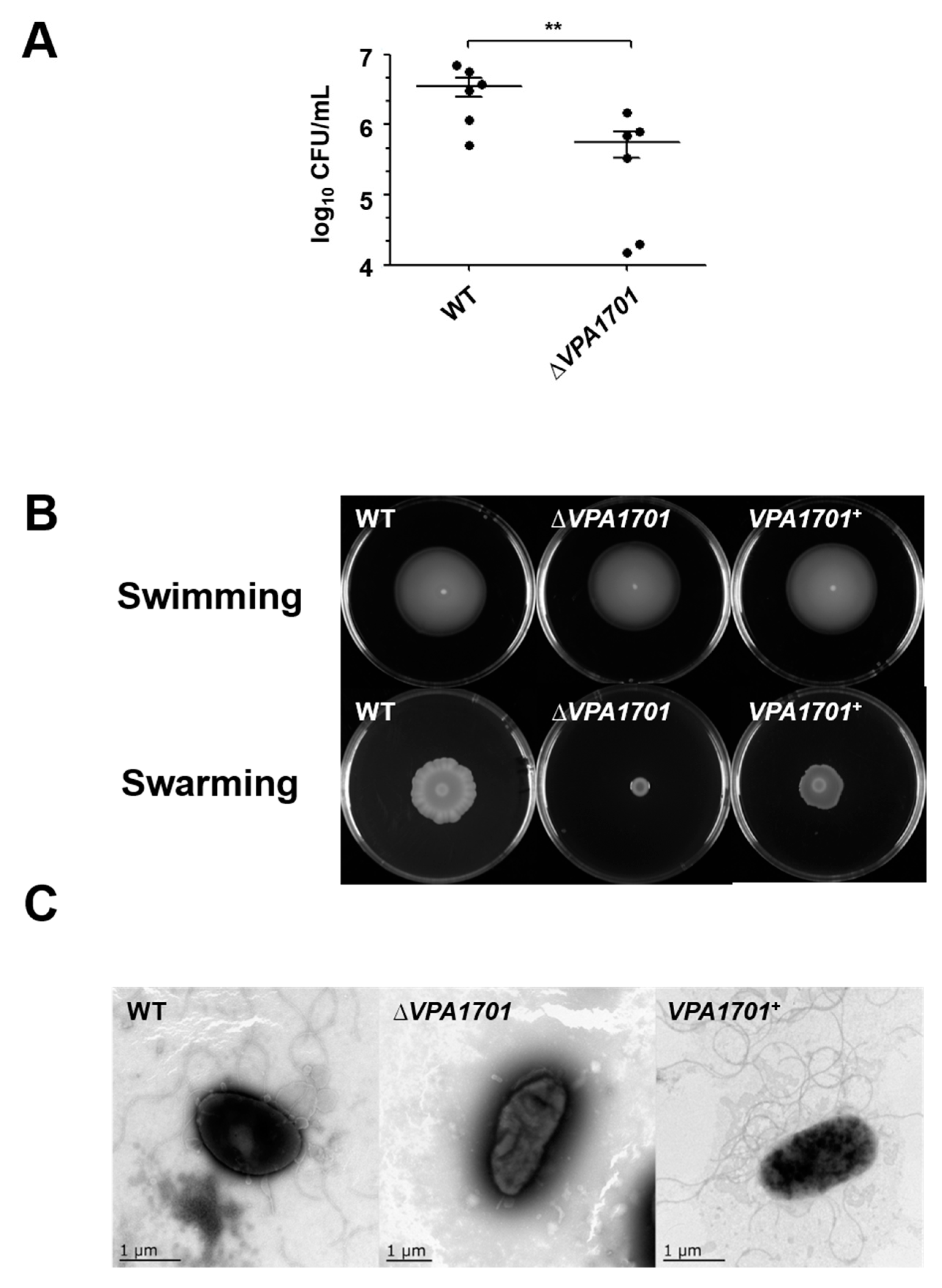
2.1. Essential Roles of VPA1701 in the Colonization and Swarming Motility of V. parahaemolyticus
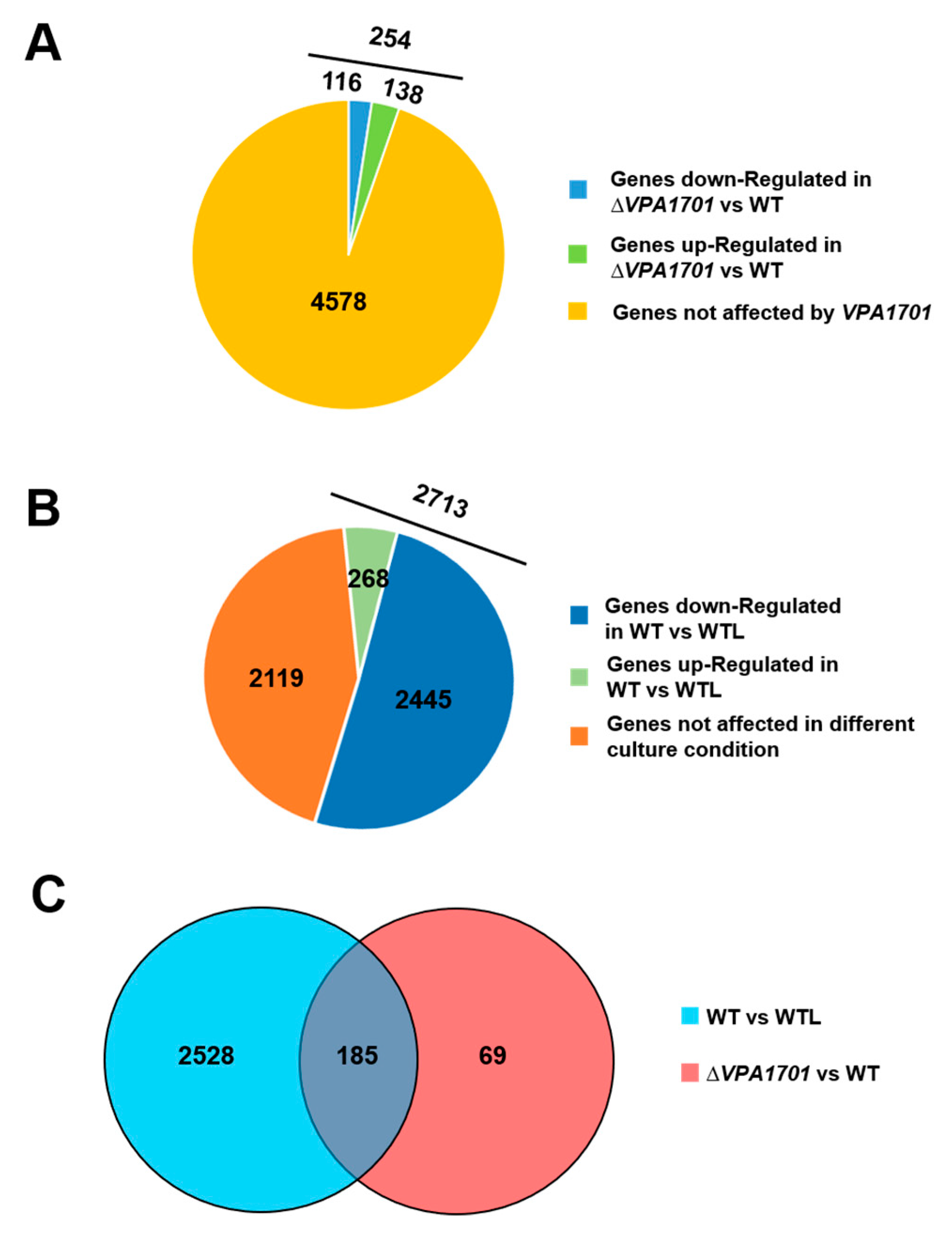
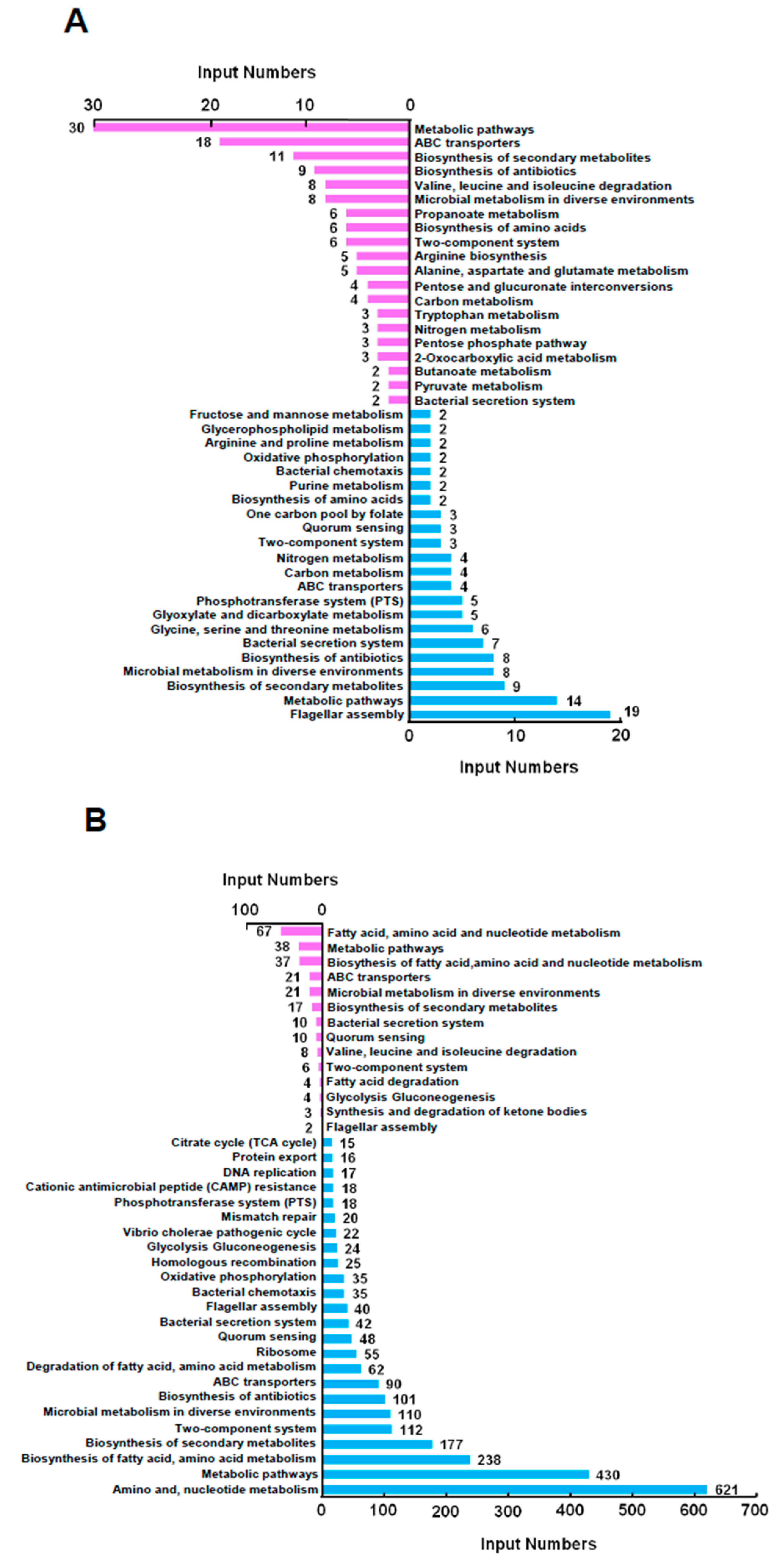
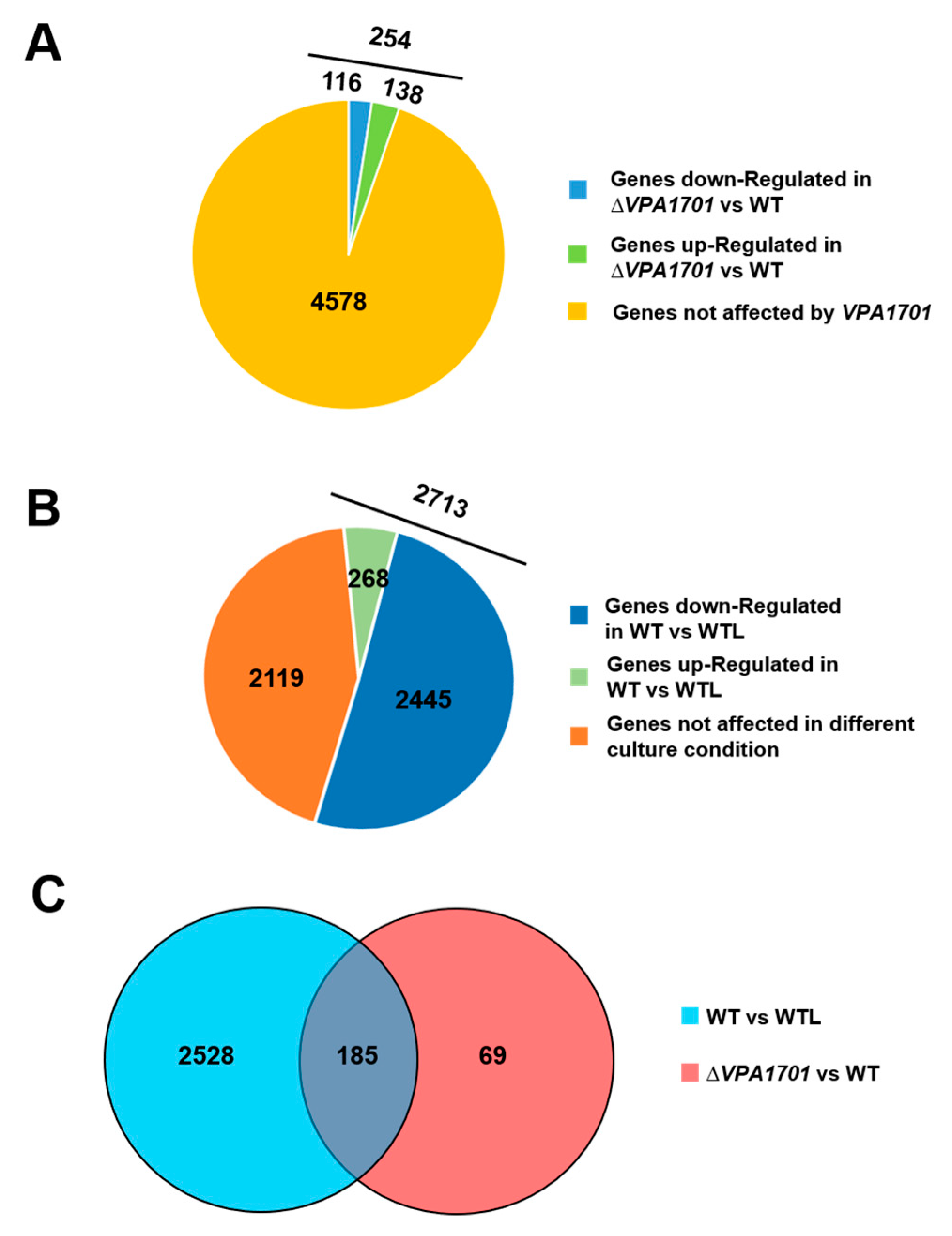
2.2. The Global Transcriptional Analysis of VPA1701 in V. parahaemolyticus
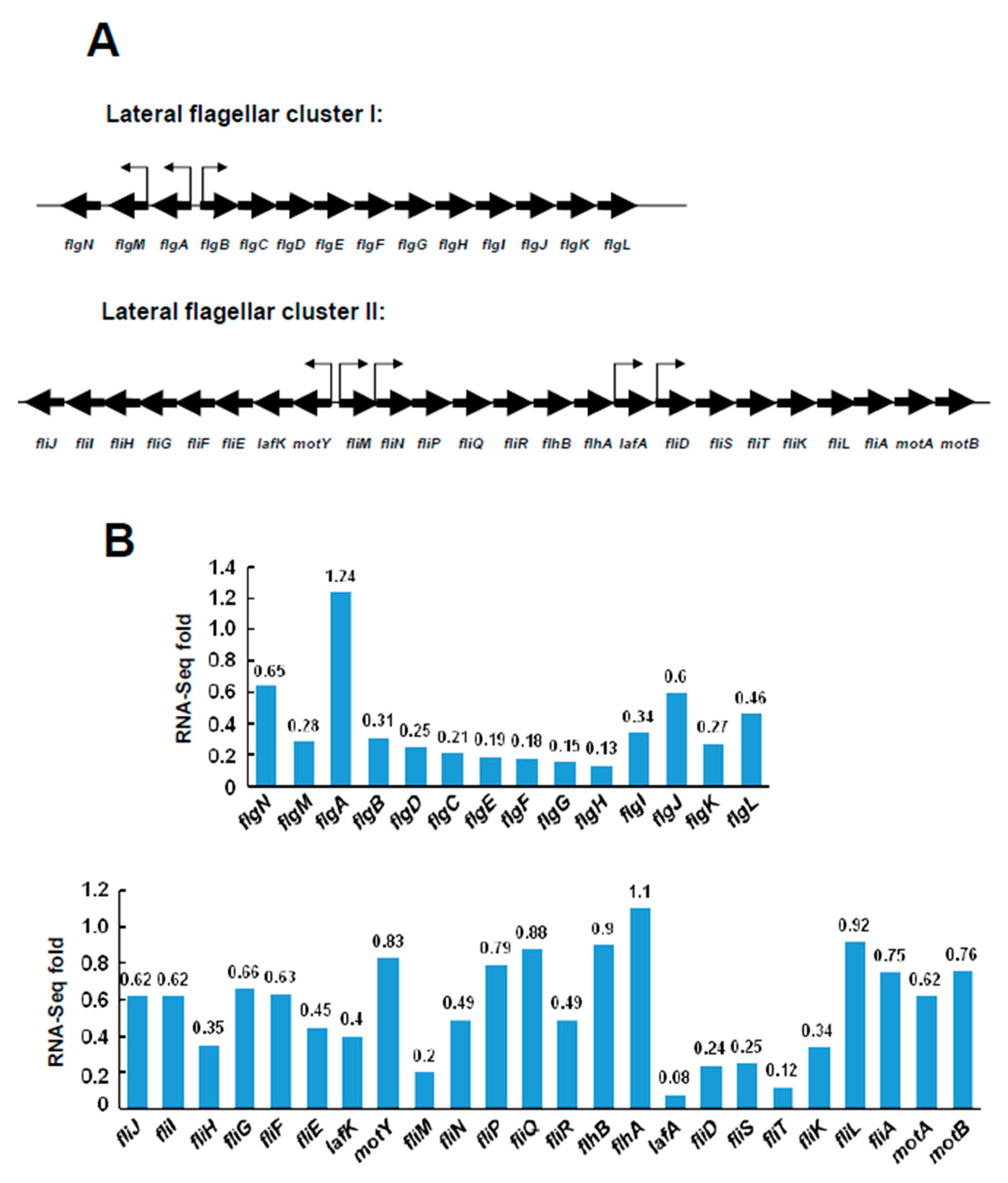
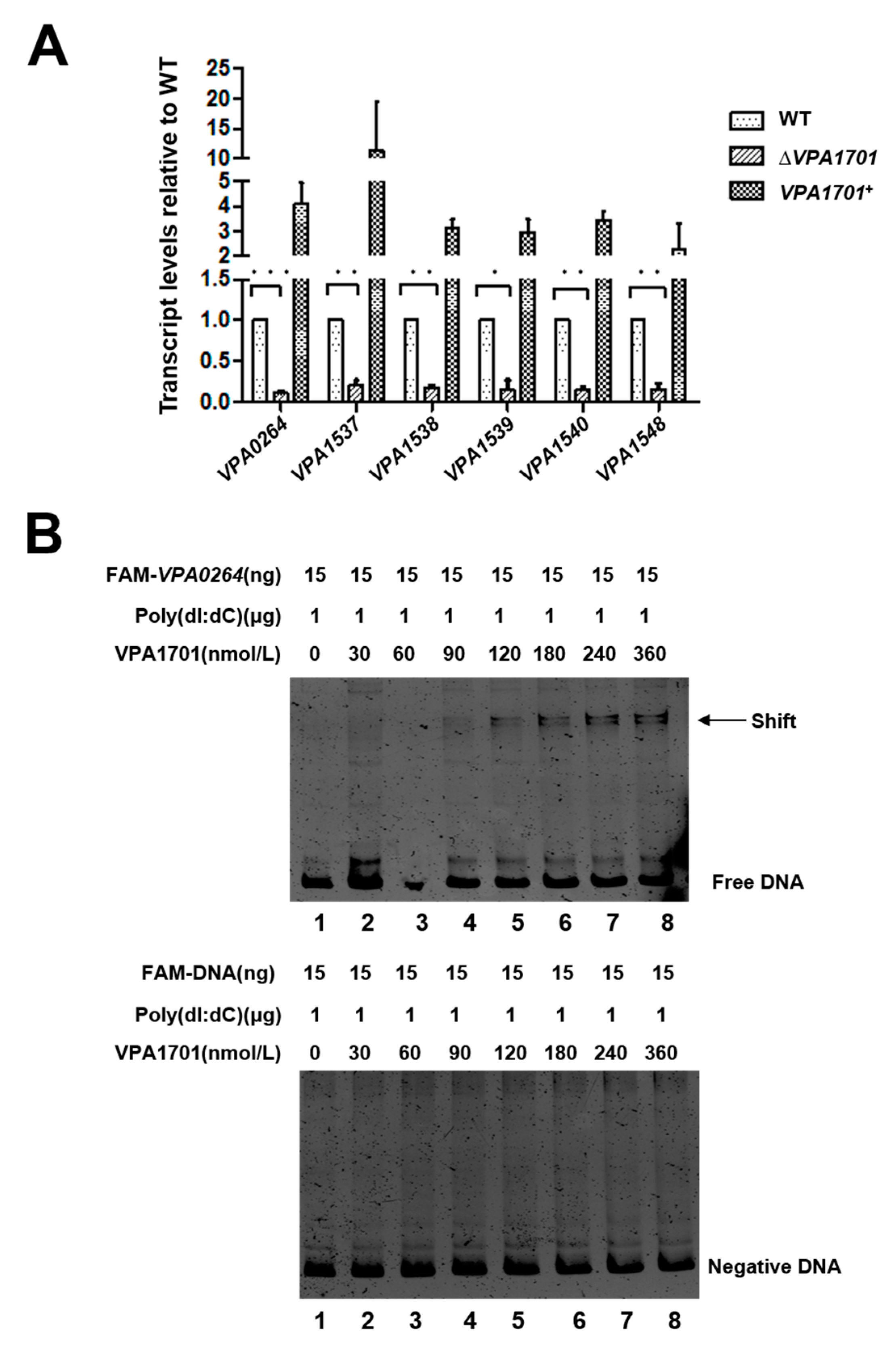
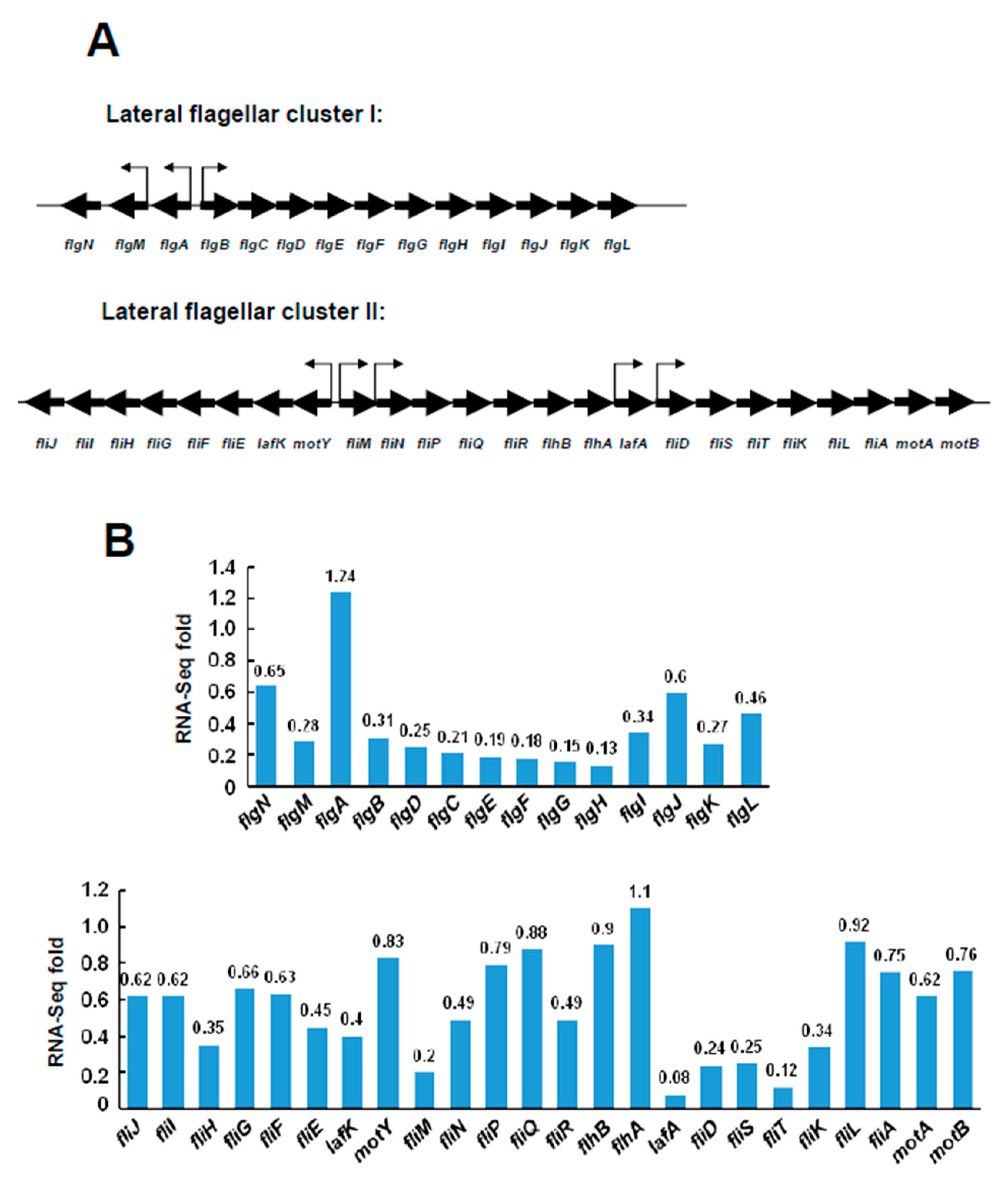
2.3. Transcriptional Levels of the Lateral Flagellar Genes in ΔVPA1701
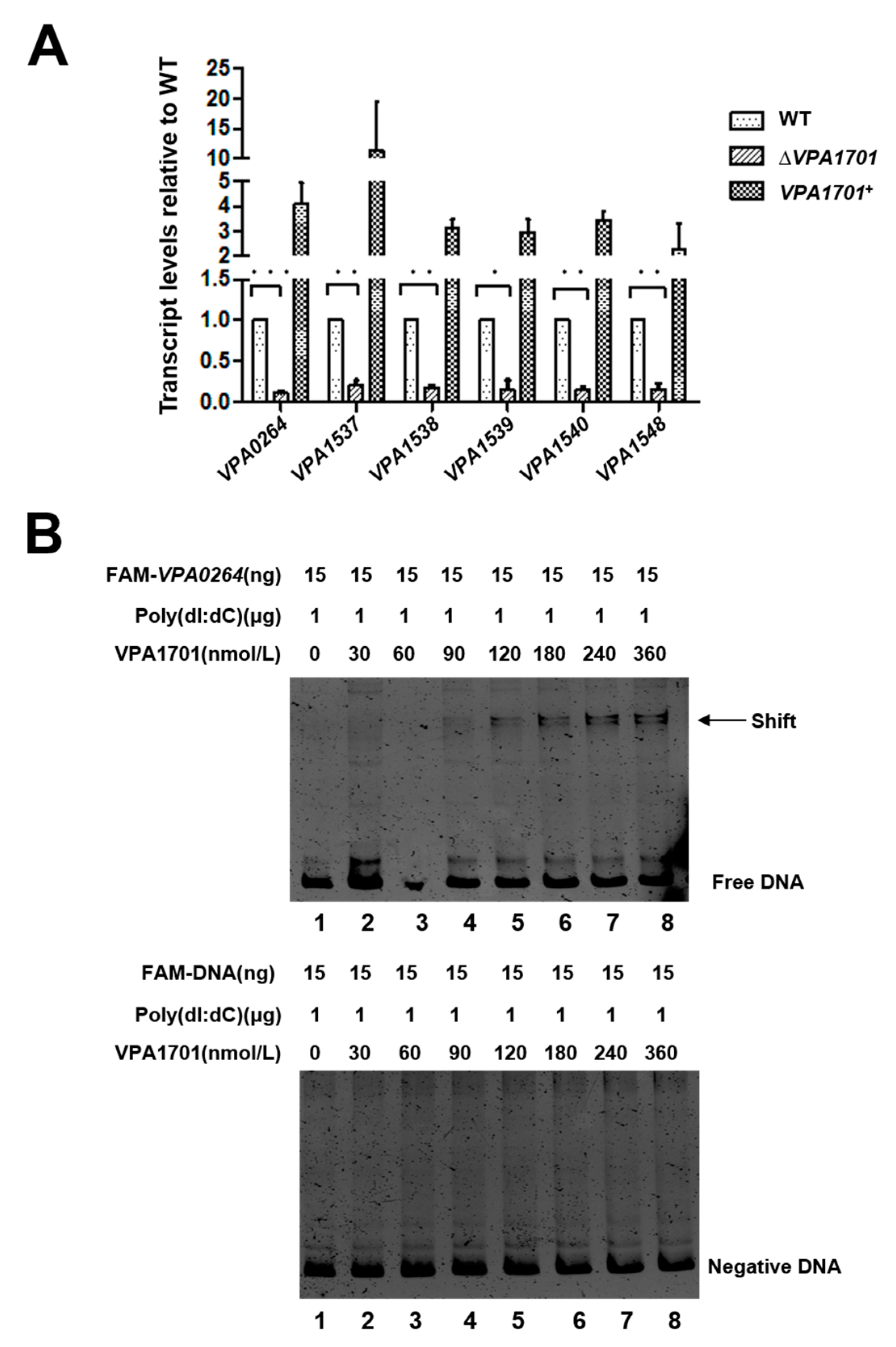
2.4. VPA1701 Directly Binds the Promoter of VPA0264 to Activate the Expression of Lateral Flagellar Genes
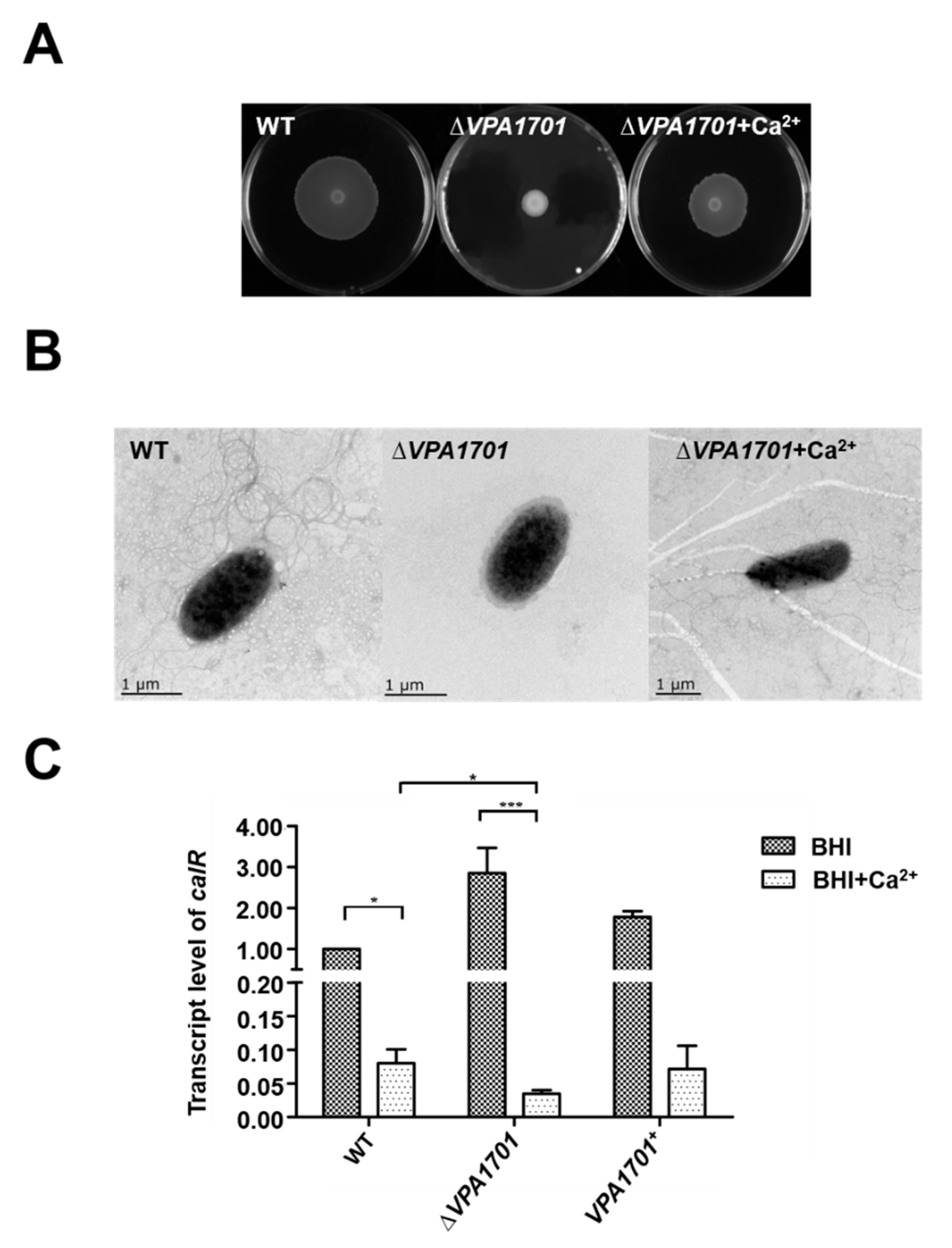
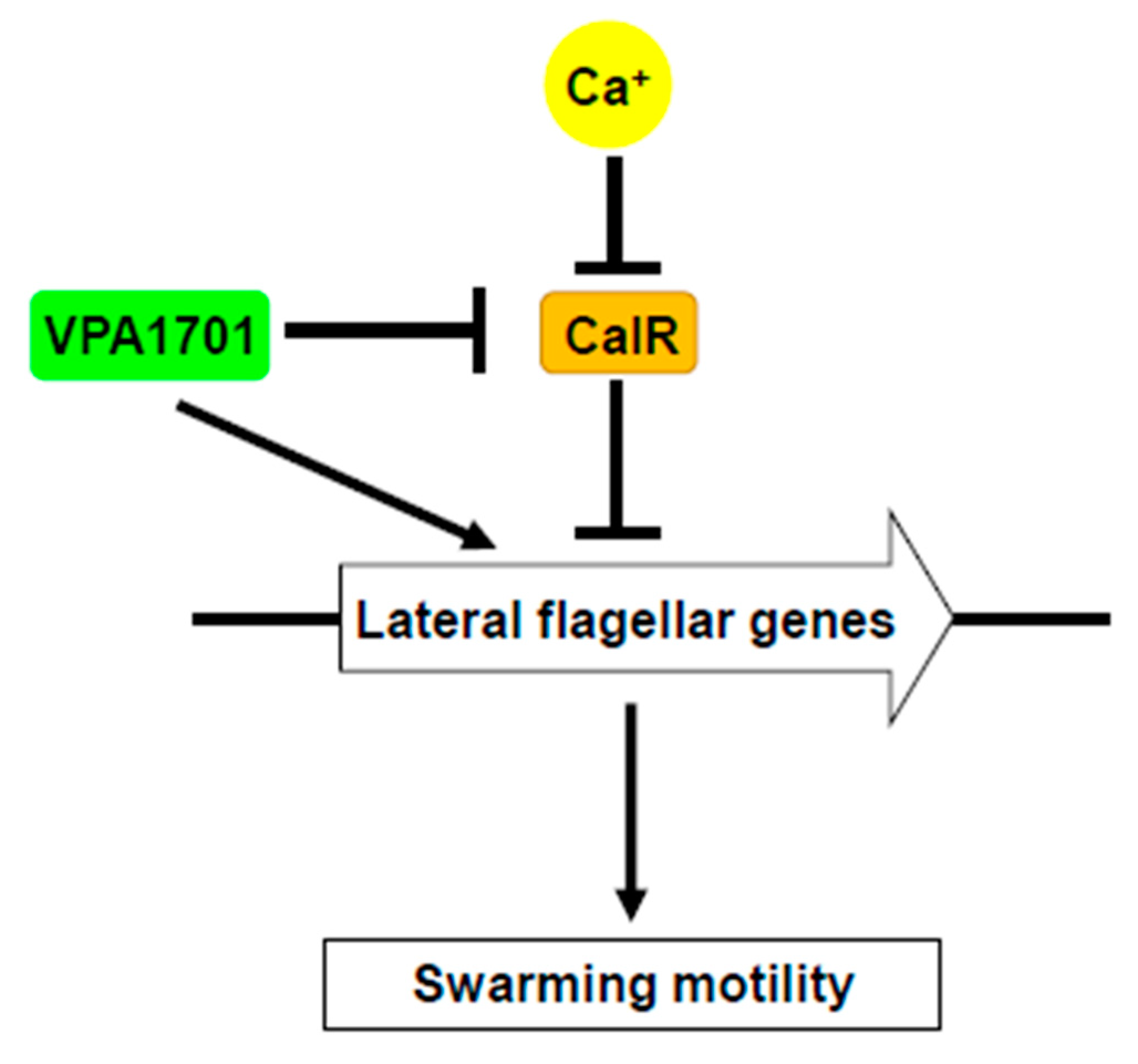
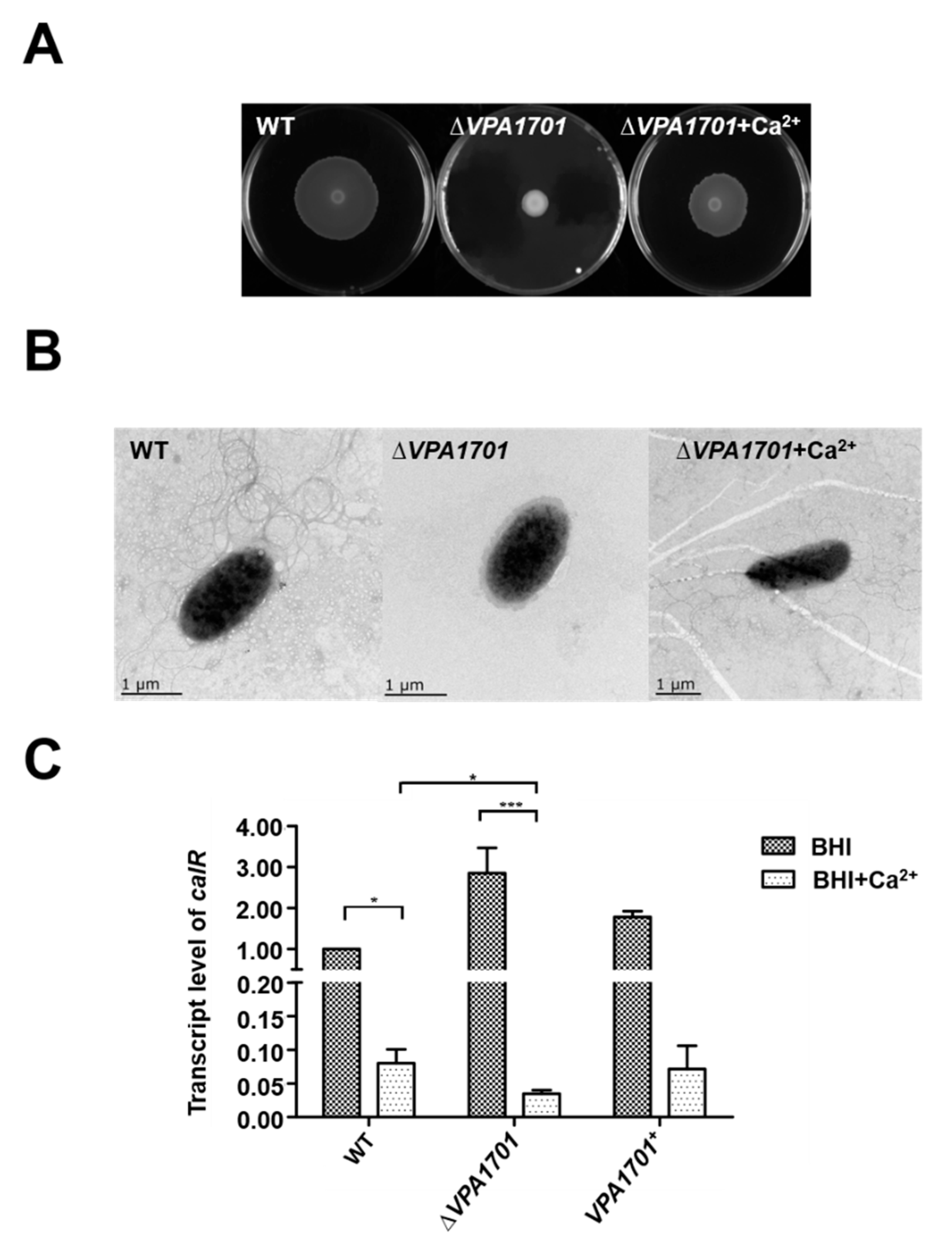
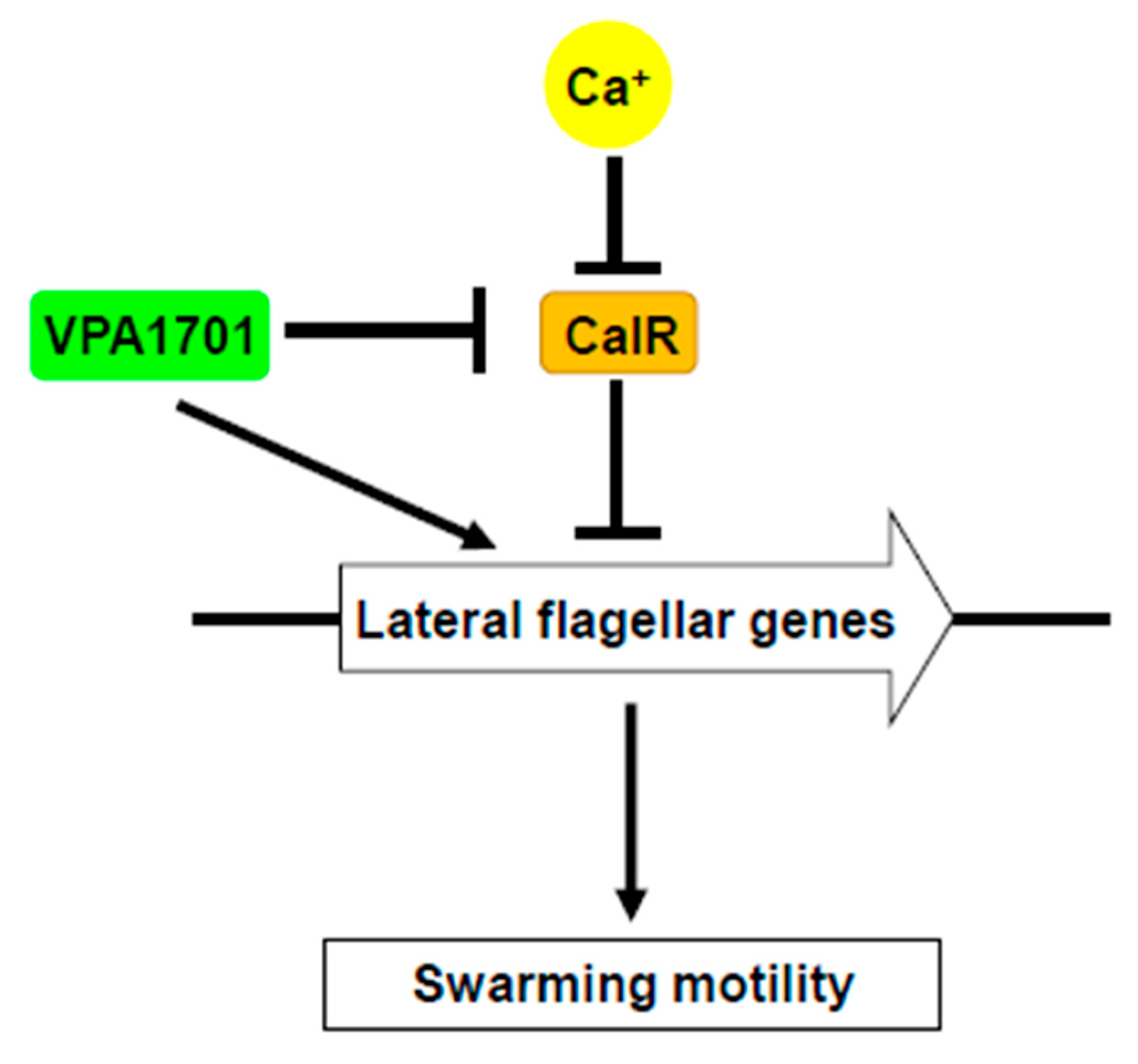
2.5. Calcium Restored the Swarming Ability of ∆VPA1701
3. Materials and Methods
3.1. Bacterial Strains, Plasmids and Culture Conditions
3.2. Construction of the VPA1701 Mutant and Complemented Strains
3.3. Motility Analysis
3.4. Transmission Electron Microscope (TEM) of the Lateral Flagellar
3.5. Quantification Real-Time Reverse Transcription PCR (qRT-PCR)
3.6. RNA-Seq Analysis
3.7. Overexpression and Purification of VPA1701 Protein
3.8. Electrophoretic Mobility Shift Assay (EMSA)
3.9. Infant Rabbit Model of V. parahaemolyticus Infection
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nair, G.B.; Ramamurthy, T.; Bhattacharya, S.K.; Dutta, B.; Takeda, Y.; Sack, D.A. Global dissemination of Vibrio parahaemolyticus serotype O3:K6 and its serovariants. Clin. Microbiol. Rev. 2007, 20, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Austin, B. Vibrios as causal agents of zoonoses. Vet. Microbiol. 2010, 140, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of Vibrio cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Salomon, D.; Gonzalez, H.; Updegraff, B.L.; Orth, K. Vibrio parahaemolyticus type VI secretion system 1 is activated in marine conditions to target bacteria, and is differentially regulated from system 2. PLoS ONE 2013, 8, e61086. [Google Scholar] [CrossRef]
- Han, N.; Mizan, M.F.R.; Jahid, I.K.; Ha, S.D. Biofilm formation by Vibrio parahaemolyticus on food and food contact surfaces increases with rise in temperature. Food Control 2016, 70, 161–166. [Google Scholar] [CrossRef]
- Li, L.; Meng, H.; Gu, D.; Li, Y.; Jia, M. Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol. Res. 2019, 222, 43–51. [Google Scholar] [CrossRef]
- Salamone, M.; Nicosia, A.; Bennici, C.; Quatrini, P.; Catania, V.; Mazzola, S.; Ghersi, G.; Cuttitta, A. Comprehensive Analysis of a Vibrio parahaemolyticus Strain Extracellular Serine Protease VpSP37. PLoS ONE 2015, 10, e0126349. [Google Scholar] [CrossRef]
- Salamone, M.; Nicosia, A.; Ghersi, G.; Tagliavia, M. Vibrio Proteases for Biomedical Applications: Modulating the Proteolytic Secretome of V. alginolyticus and V. parahaemolyticus for Improved Enzymes Production. Microorganisms 2019, 7, 387. [Google Scholar] [CrossRef]
- Merrell, B.R.; Walker, R.I.; Joseph, S.W. In vitro and in vivo pathologic effects of Vibrio parahaemolyticus on human epithelial cells. Can. J. Microbiol. 1984, 30, 381–388. [Google Scholar] [CrossRef]
- O’Boyle, N.; Houeix, B.; Kilcoyne, M.; Joshi, L.; Boyd, A. The MSHA pilus of Vibrio parahaemolyticus has lectin functionality and enables TTSS-mediated pathogenicity. Int. J. Med. Microbiol. 2013, 303, 563–573. [Google Scholar] [CrossRef]
- McCarter, L.; Silverman, M. Surface-induced swarmer cell differentiation of Vibrio parahaemolyticus. Mol. Microbiol. 1990, 4, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.S.; Ledizet, M.; Kazmierczak, B. Swarming motility, secretion of type 3 effectors and biofilm formation phenotypes exhibited within a large cohort of Pseudomonas aeruginosa clinical isolates. J. Med. Microbiol. 2010, 59, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Merino, S.; Shaw, J.G.; Tomás, J.M. Bacterial lateral flagella: An inducible flagellar system. FEMS Microbiol. Lett. 2006, 263, 127–135. [Google Scholar] [CrossRef]
- Inaba, S.; Nishigaki, T.; Takekawa, N.; Kojima, S.; Homma, M. Localization and domain characterization of the SflA regulator of flagellar formation in Vibrio alginolyticus. Genes Cells 2017, 22, 619–627. [Google Scholar] [CrossRef]
- McCarter, L.L. Polar Flagellar Motility of the Vibrionaceae. Microbiol. Mol. Biol. Rev. 2001, 65, 445–462. [Google Scholar] [CrossRef]
- Boles, B.R.; McCarter, L.L. Insertional Inactivation of Genes Encoding Components of the Sodium-Type Flagellar Motor and Switch of Vibrio parahaemolyticus. J. Bacteriol. 2000, 182, 1035–1045. [Google Scholar] [CrossRef]
- Kim, Y.K.; McCarter, L.L. Cross-Regulation in Vibrio parahaemolyticus: Compensatory Activation of Polar Flagellar Genes by the Lateral Flagellar Regulator LafK. J. Bacteriol. 2004, 186, 4014–4018. [Google Scholar] [CrossRef]
- Kim, Y.K.; McCarter, L.L. Analysis of the polar flagellar gene system of Vibrio parahaemolyticus. J. Bacteriol. 2000, 182, 3693–3704. [Google Scholar] [CrossRef]
- Stewart, B.J.; McCarter, L.L. Lateral flagellar gene system of Vibrio parahaemolyticus. J. Bacteriol. 2003, 185, 4508–4518. [Google Scholar] [CrossRef]
- Shinoda, S.; Okamoto, K. Formation and function of Vibrio parahaemolyticus lateral flagella. J. Bacteriol. 1977, 129, 1266–1271. [Google Scholar] [PubMed]
- Tian, Y.; Wang, Q.; Liu, Q.; Ma, Y.; Cao, X.; Guan, L.; Zhang, Y. Involvement of LuxS in the regulation of motility and flagella biogenesis in Vibrio alginolyticus. Biosci. Biotechnol. Biochem. 2008, 72, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Henares, B.M.; Xu, Y.; Boon, E.M. A Nitric Oxide-Responsive Quorum Sensing Circuit in Vibrio harveyi Regulates Flagella Production and Biofilm Formation. Int. J. Mol. Sci. 2013, 14, 16473–16484. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Tang, H.; Qiu, Y.; Yang, W.; Yang, H.; Zhou, D.; Huang, X.; Hu, L.; Zhang, Y. Quorum sensing regulates the transcription of lateral flagellar genes in Vibrio parahaemolyticus. Future Microbiol. 2019, 14, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Gode-Potratz, C.J.; Chodur, D.M.; McCarter, L.L. Calcium and Iron Regulate Swarming and Type III Secretion in Vibrio parahaemolyticus. J. Bacteriol. 2010, 192, 6025–6038. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; Hsieh, M.L.; Khataokar, A.; Neiditch, M.B.; Waters, C.M. Cyclic di-GMP inhibits Vibrio cholerae motility by repressing induction of transcription and inducing extracellular polysaccharide production. Mol. Microbiol. 2013, 90, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Park, Y.H.; Lee, C.R.; Kim, Y.R.; Seok, Y.J. Glucose induces delocalization of a flagellar biosynthesis protein from the flagellated pole. Mol. Microbiol. 2016, 101, 795–808. [Google Scholar] [CrossRef]
- Chen, L.; Qiu, Y.; Tang, H.; Hu, L.F.; Yang, W.H.; Zhu, X.J.; Huang, X.X.; Wang, T.; Zhang, Y.Q. ToxR Is Required for Biofilm Formation and Motility of Vibrio Parahaemolyticus. Biomed. Environ. Sci. 2018, 31, 848–850. [Google Scholar]
- Hubbard, T.P.; Chao, M.C.; Abel, S.; Blondel, C.J.; Abel Zur Wiesch, P.; Zhou, X.; Davis, B.M.; Waldor, M.K. Genetic analysis of Vibrio parahaemolyticus intestinal colonization. Proc. Natl. Acad. Sci. USA 2016, 113, 6283–6288. [Google Scholar] [CrossRef]
- Haydon, D.J.; Guest, J.R. A new family of bacterial regulatory proteins. FEMS Microbiol. Lett. 1991, 63, 291–295. [Google Scholar] [CrossRef]
- Ushijima, B.; Häse, C.C. Influence of Chemotaxis and Swimming Patterns on the Virulence of the Coral Pathogen Vibrio coralliilyticus. J. Bacteriol. 2018, 200, e00791-17. [Google Scholar] [CrossRef]
- Jaques, S.; McCarter, L.L. Three new regulators of swarming in Vibrio parahaemolyticus. J. Bacteriol. 2006, 88, 2625–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Osei-Adjei, G.; Zhang, Y.; Gao, H.; Yang, W.; Zhou, D.; Huang, X.; Yang, H.; Zhang, Y. CalR is required for the expression of T6SS2 and the adhesion of Vibrio parahaemolyticus to HeLa cells. Arch. Microbiol. 2017, 199, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, L.; Osei-Adjei, G.; Yang, W.; Zhou, D.; Huang, X.; Yang, H.; Yin, Z.; Zhang, Y. Transcriptional regulation of cpsQ-mfpABC and mfpABC by CalR in Vibrio parahaemolyticus. Microbiologyopen 2017, 6, e470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Gao, H.; Zhang, L.; Yin, Z.; Huang, X.; Zhou, D.; Yang, H.; Yang, W.; Wang, L. Vibrio parahaemolyticus CalR down regulates the thermostable direct hemolysin (TDH) gene transcription and thereby inhibits hemolytic activity. Gene 2017, 613, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Konkel, M.E.; Call, D.R. Vp1659 is a Vibrio parahaemolyticus type III secretion system 1 protein that contributes to translocation of effector proteins needed to induce cytolysis, autophagy, and disruption of actin structure in HeLa cells. J. Bacteriol. 2010, 192, 3491–3502. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, W.B.; Richards, G.P.; Boyd, E.F. Loss of Sigma Factor RpoN Increases Intestinal Colonization of Vibrio parahaemolyticus in an Adult Mouse Model. Infect. Immun. 2014, 82, 544–556. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; Guo, M.; Yang, M.; Zhang, Y.; Zhou, X.; Wang, Q. A σE-Mediated Temperature Gauge Controls a Switch from LuxR-Mediated Virulence Gene Expression to Thermal Stress Adaptation in Vibrio alginolyticus. PLoS Pathog. 2016, 12, e1005645. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, L.; Yang, M.; Yin, K.; Zhou, X.; Leung, K.Y.; Liu, Q.; Zhang, Y.; Wang, Q. Transcriptomic dissection of the horizontally acquired response regulator EsrB reveals its global regulatory roles in the physiological adaptation and activation of T3SS and the cognate effector repertoire in Edwardsiella piscicida during infection toward turbot. Virulence 2017, 8, 1355–1377. [Google Scholar]
- Abel, S.; Waldor, M.K. Infant Rabbit Model for Diarrheal Diseases. Curr. Protoc. Microbiol. 2015, 38, 6A.6.1-15. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, J.M.; Rui, H.; Zhou, X.; Iida, T.; Kodoma, T.; Ito, S.; Davis, B.M.; Bronson, R.T.; Waldor, M.K. Inflammation and Disintegration of Intestinal Villi in an Experimental Model for Vibrio parahaemolyticus-Induced Diarrhea. PLoS Pathog. 2012, 8, e1002593. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, D.; Meng, H.; Li, Y.; Ge, H.; Jiao, X. A GntR Family Transcription Factor (VPA1701) for Swarming Motility and Colonization of Vibrio parahaemolyticus. Pathogens 2019, 8, 235. https://doi.org/10.3390/pathogens8040235
Gu D, Meng H, Li Y, Ge H, Jiao X. A GntR Family Transcription Factor (VPA1701) for Swarming Motility and Colonization of Vibrio parahaemolyticus. Pathogens. 2019; 8(4):235. https://doi.org/10.3390/pathogens8040235
Chicago/Turabian StyleGu, Dan, Hongmei Meng, Yang Li, Haojie Ge, and Xinan Jiao. 2019. "A GntR Family Transcription Factor (VPA1701) for Swarming Motility and Colonization of Vibrio parahaemolyticus" Pathogens 8, no. 4: 235. https://doi.org/10.3390/pathogens8040235
APA StyleGu, D., Meng, H., Li, Y., Ge, H., & Jiao, X. (2019). A GntR Family Transcription Factor (VPA1701) for Swarming Motility and Colonization of Vibrio parahaemolyticus. Pathogens, 8(4), 235. https://doi.org/10.3390/pathogens8040235