Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt

 ,
,  ,
,  ,
, 
 and
and
Abstract
1. Introduction
2. Results
2.1. Resistance Phenotypes and Genotypes of Human, Chicken and Beef E. coli Isolates
2.2. Correlation and Principal Component Analyses of Resistance Phenotypes and Genotypes in E. coli Isolates
2.3. Phylogrouping of Human, Chicken and Beef E. coli Isolates
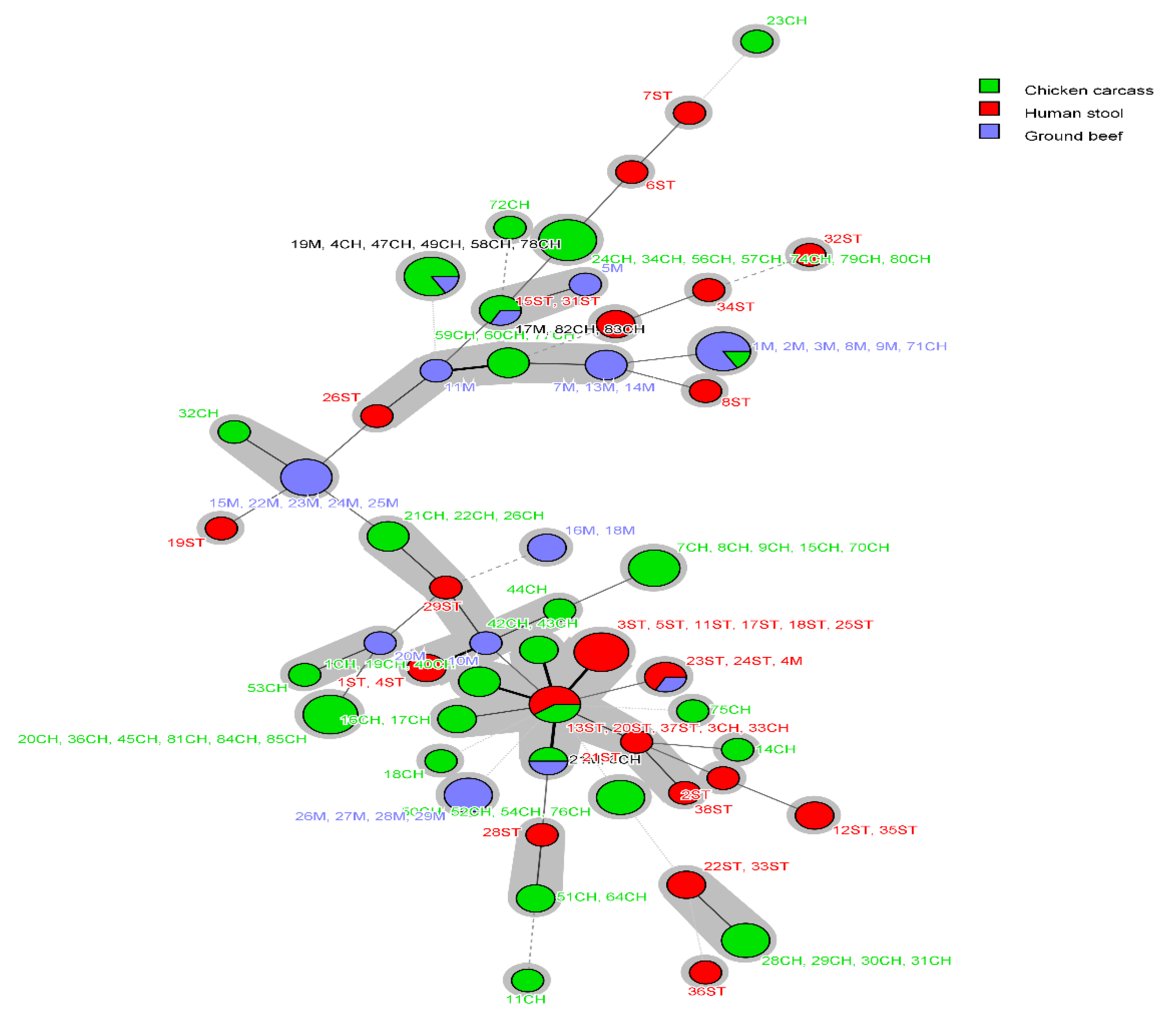
2.4. PFGE and MLST Typing of the Examined E. coli Isolates
3. Discussion
4. Materials and Methods
4.1. E. coli Isolates from Humans and Retail Chicken and Beef
4.2. Antimicrobial Susceptibility Testing of E. coli
4.3. Resistance Genotypes and E. coli Phylogrouping
4.4. Pulsed-Field Gel Electrophoresis (PFGE) Analysis
4.5. Multilocus Sequence Typing (MLST)
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clements, A.; Young, J.C.; Constantinou, N.; Frankel, G. Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes 2012, 3, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Padola, N.; Parma, A.; Lucchesi, P. Enteropathogenic Escherichia coli contamination at different stages of the chicken slaughtering process. Poult. Sci. 2011, 90, 2638–2641. [Google Scholar] [CrossRef] [PubMed]
- Projahn, M.; Pacholewicz, E.; Becker, E.; Correia-Carreira, G.; Bandick, N.; Kaesbohrer, A. Reviewing interventions against Enterobacteriaceae in broiler processing: Using old techniques for meeting the new challenges of ESBL E. coli? BioMed Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Frye, J.G.; Jackson, C.R. Genetic mechanisms of antimicrobial resistance identified in Salmonella enterica, Escherichia coli, and Enterococcus spp. isolated from US food animals. Front. Microbiol. 2013, 4, 135. [Google Scholar] [CrossRef]
- Adenipekun, E.O.; Jackson, C.R.; Ramadan, H.; Iwalokun, B.A.; Oyedeji, K.S.; Frye, J.G.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A.; Oluwadun, A. Prevalence and multidrug resistance of Escherichia coli from community-acquired infections in Lagos, Nigeria. J. Infect. Dev. Ctries. 2016, 10, 920–931. [Google Scholar] [CrossRef]
- Collignon, P.C.; Conly, J.M.; Andremont, A.; McEwen, S.A.; Aidara-Kane, A.; World Health Organization Advisory Group; Bogotá Meeting on Integrated Surveillance of Antimicrobial Resistance (WHO-AGISAR); Agerso, Y.; Andremont, A.; Collignon, P.; et al. World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar] [CrossRef]
- Dandachi, I.; Chabou, S.; Daoud, Z.; Rolain, J.-M. Prevalence and emergence of extended-spectrum cephalosporin-, carbapenem- and colistin-resistant Gram negative bacteria of animal origin in the Mediterranean Basin. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- World Health Organization. Critically Important Antimicrobials for Human Medicine: Ranking of Antimicrobial Agents for Risk Management of Antimicrobial Resistance due to Non-Human Use; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Castellanos, L.R.; Donado-Godoy, P.; Leon, M.; Clavijo, V.; Arevalo, A.; Bernal, J.F.; Timmerman, A.J.; Mevius, D.J.; Wagenaar, J.A.; Hordijk, J. High Heterogeneity of Escherichia coli sequence types harbouring ESBL/AmpC genes on IncI1 plasmids in the Colombian poultry chain. PLoS ONE 2017, 12, e0170777. [Google Scholar] [CrossRef]
- Yamaji, R.; Friedman, C.R.; Rubin, J.; Suh, J.; Thys, E.; McDermott, P.; Hung-Fan, M.; Riley, L.W. A population-based surveillance study of shared genotypes of Escherichia coli isolates from retail meat and suspected cases of urinary tract infections. mSphere 2018, 3, e00179-18. [Google Scholar] [CrossRef]
- Kluytmans, J.A.J.W.; Overdevest, I.T.M.A.; Willemsen, I.; Kluytmans-van den Bergh, M.F.Q.; van der Zwaluw, K.; Heck, M.; Rijnsburger, M.; Vandenbroucke-Grauls, C.M.J.E.; Savelkoul, P.H.M.; Johnston, B.D.; et al. Extended-Spectrum β-Lactamase–producing Escherichia coli from retail chicken meat and humans: Comparison of strains, plasmids, resistance genes, and virulence factors. Clin. Infect. Dis. 2012, 56, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Ribot, E.M.; Fair, M.; Gautom, R.; Cameron, D.; Hunter, S.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157: H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Reuland, E.A.; Wintermans, B.B.; Al Naiemi, N.; Koek, A.; Abdelwahab, A.M.; Ammar, A.M.; Mohamed, A.A.; Vandenbroucke-Grauls, C.M.J.E. Extended-Spectrum β-Lactamases and/or carbapenemases-producing Enterobacteriaceae isolated from retail chicken meat in Zagazig, Egypt. PLoS ONE 2015, 10, e0136052. [Google Scholar] [CrossRef]
- El-Shazly, D.A.; Nasef, S.A.; Mahmoud, F.F.; Jonas, D. Expanded spectrum β–lactamase producing Escherichia coli isolated from chickens with colibacillosis in Egypt. Poult. Sci. 2017, 96, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El-Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Enany, M.E.; Algammal, A.M.; Nasef, S.A.; Abo-Eillil, S.A.M.; Bin-Jumah, M.; Taha, A.E.; Allam, A.A. The occurrence of the multidrug resistance (MDR) and the prevalence of virulence genes and QACs resistance genes in E. coli isolated from environmental and avian sources. AMB Express 2019, 9, 192. [Google Scholar] [CrossRef]
- Wang, J.; Zhi, C.-P.; Chen, X.-J.; Guo, Z.-W.; Liu, W.-L.; Luo, J.; Huang, X.-Y.; Zeng, L.; Huang, J.-W.; Xia, Y.-B.; et al. Characterization of oqxAB in Escherichia coli isolates from animals, retail meat, and human patients in Guangzhou, China. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Adenipekun, E.O.; Jackson, C.R.; Ramadan, H.; Iwalokun, B.A.; Frye, J.G.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A.; House, S.L.; McMillan, E.A.; et al. Plasmid replicons and beta-lactamase-encoding genes of multidrug-resistant Escherichia coli isolated from humans and food animals in Lagos, Southwest Nigeria. Microb. Drug Resist. 2019, 25, 1410–1423. [Google Scholar] [CrossRef]
- Yassin, A.K.; Gong, J.; Kelly, P.; Lu, G.; Guardabassi, L.; Wei, L.; Han, X.; Qiu, H.; Price, S.; Cheng, D.; et al. Antimicrobial resistance in clinical Escherichia coli isolates from poultry and livestock, China. PLoS ONE 2017, 12, e0185326. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial resistance in bacterial poultry pathogens: A review. Front. Vet. Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.D.; Ahmed, M.F.; El-Adawy, H.; Hotzel, H.; Engelmann, I.; Weiß, D.; Monecke, S.; Ehricht, R. Surveillance of extended-spectrum beta-lactamase-producing Escherichia coli in dairy cattle farms in the Nile Delta, Egypt. Front. Microbiol. 2016, 7, 1020. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, H.; Gupta, S.K.; Sharma, P.; Ahmed, M.; Hiott, L.M.; Barrett, J.B.; Woodley, T.A.; Frye, J.G.; Jackson, C.R. Circulation of emerging NDM-5-producing Escherichia coli among humans and dogs in Egypt. Zoonoses Public Health 2019. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Shimamoto, T.; Shimamoto, T. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Int. J. Med. Microbiol. IJMM 2013, 303, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, H.; Awad, A. Phenotypic and genetic characterization of β-lactam resistance in Klebsiella from retail chicken meat in Mansoura, Egypt. Iran J. Microbiol. 2017, 9, 74–81. [Google Scholar]
- Cuong, N.V.; Padungtod, P.; Thwaites, G.; Carrique-Mas, J.J. Antimicrobial usage in animal production: A review of the literature with a focus on low- and middle-income countries. Antibiotics 2018, 7, 75. [Google Scholar] [CrossRef]
- Brown, E.M.; Nathwani, D. Antibiotic cycling or rotation: A systematic review of the evidence of efficacy. J. Antimicrob. Chemother. 2005, 55, 6–9. [Google Scholar] [CrossRef]
- Wales, A.D.; Davies, R.H. Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef]
- Pietsch, M.; Irrgang, A.; Roschanski, N.; Brenner Michael, G.; Hamprecht, A.; Rieber, H.; Käsbohrer, A.; Schwarz, S.; Rösler, U.; Kreienbrock, L.; et al. Whole genome analyses of CMY-2-producing Escherichia coli isolates from humans, animals and food in Germany. BMC Genom. 2018, 19, 601. [Google Scholar] [CrossRef]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and characterization of ESBL-producing Escherichia coli from humans and poultry in Ghana. Front. Microbiol. 2019, 9, 3358. [Google Scholar] [CrossRef]
- Fam, N.; Leflon-Guibout, V.; Fouad, S.; Aboul-Fadl, L.; Marcon, E.; Desouky, D.; El-Defrawy, I.; Abou-Aitta, A.; Klena, J.; Nicolas-Chanoine, M.H. CTX-M-15-producing Escherichia coli clinical isolates in Cairo (Egypt), including isolates of clonal complex ST10 and clones ST131, ST73, and ST405 in both community and hospital settings. Microb. Drug Resist. 2011, 17, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Elnahriry, S.S.; Khalifa, H.O.; Soliman, A.M.; Ahmed, A.M.; Hussein, A.M.; Shimamoto, T.; Shimamoto, T. Emergence of Plasmid-mediated colistin resistance gene mcr-1 in a clinical Escherichia coli isolate from Egypt. Antimicrob. Agents Chemother. 2016, 60, 3249. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.H.; Ghanem, I.A.; Eid, A.A.; Ali, M.A.; Sherwood, J.S.; Li, G.; Nolan, L.K.; Logue, C.M. Molecular and phenotypic characterization of Escherichia coli isolated from broiler chicken flocks in Egypt. Avian Dis. 2013, 57, 602–611. [Google Scholar] [CrossRef]
- Yu, F.; Chen, X.; Zheng, S.; Han, D.; Wang, Y.; Wang, R.; Wang, B.; Chen, Y. Prevalence and genetic diversity of human diarrheagenic Escherichia coli isolates by multilocus sequence typing. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2018, 67, 7–13. [Google Scholar] [CrossRef]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef]
- Nemoy, L.L.; Kotetishvili, M.; Tigno, J.; Keefer-Norris, A.; Harris, A.D.; Perencevich, E.N.; Johnson, J.A.; Torpey, D.; Sulakvelidze, A.; Morris, J.G., Jr.; et al. Multilocus sequence typing versus pulsed-field gel electrophoresis for characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolates. J. Clin. Microbiol. 2005, 43, 1776–1781. [Google Scholar] [CrossRef]
- Johnson, J.K.; Arduino, S.M.; Stine, O.C.; Johnson, J.A.; Harris, A.D. Multilocus sequence typing compared to pulsed-field gel electrophoresis for molecular typing of Pseudomonas aeruginosa. J. Clin. Microbiol. 2007, 45, 3707–3712. [Google Scholar] [CrossRef]
- Henri, C.; Félix, B.; Guillier, L.; Leekitcharoenphon, P.; Michelon, D.; Mariet, J.-F.; Aarestrup, F.M.; Mistou, M.-Y.; Hendriksen, R.S.; Roussel, S. Population Genetic structure of Listeria monocytogenes strains as determined by Pulsed-Field Gel Electrophoresis and Multilocus Sequence Typing. Appl. Environ. Microbiol. 2016, 82, 5720–5728. [Google Scholar] [CrossRef]
- Machado, G.E.; Matsumoto, C.K.; Chimara, E.; Duarte, R.d.S.; de Freitas, D.; Palaci, M.; Hadad, D.J.; Lima, K.V.B.; Lopes, M.L.; Ramos, J.P.; et al. Multilocus sequence typing scheme versus pulsed-field gel electrophoresis for typing Mycobacterium abscessus isolates. J. Clin. Microbiol. 2014, 52, 2881–2891. [Google Scholar] [CrossRef]
- Quinn, P.; Markey, B.; Carter, M.; Donnelly, W.; Leonard, F. Veterinary Microbiology and Microbial Disease; Blackwell Science Company: Hoboken, NJ, USA, 2002. [Google Scholar]
- Müller, D.; Greune, L.; Heusipp, G.; Karch, H.; Fruth, A.; Tschäpe, H.; Schmidt, M.A. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl. Environ. Microbiol. 2007, 73, 3380–3390. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Seventh Informational Supplement M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Ramadan, H.H.; Jackson, C.R.; Taha, S.A.; Moawad, A.A.; Barrett, J.B.; Woodley, T.A. Contribution of healthy chickens to antimicrobial-resistant Escherichia coli associated with human extraintestinal infections in Egypt. Vector Borne Zoonotic Dis. 2018, 18, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Nguyen, H.A.T.; McDonald, J.M.; Woodley, T.A.; Hiott, L.M.; Barrett, J.B.; Jackson, C.R.; Frye, J.G. Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA. Int. J. Environ. Res. Public Health 2019, 16, 3761. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Friendly, M. Corrgrams. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antimicrobial Agent | Antimicrobial Class | Total (n = 120) No. Resistant (%) | Chicken (n = 61) No. Resistant (%) | Beef (n = 27) No. Resistant (%) | Human (n = 32) No. Resistant (%) | p-Value |
|---|---|---|---|---|---|---|
| AM/CLA | β-Lactam/β-Lactamase inhibitor | 4 (3.3) | 2 (3.3) | 0 | 2 (6.3) | 0.540000 |
| AMP | Penicillins | 87 (72.5) | 60 (98.4) | 10 (37) | 17 (53.1) | <0.0001 |
| AXO | Cephalosporins | 51 (42.5) | 43 (70.5) | 2 (7.4) | 6 (18.8) | <0.0001 |
| FOX | 2 (1.7) | 2 (3.3) | 0 | 0 | 0.720000 | |
| NAL | Quinolones | 66 (55) | 59 (96.7) | 4 (14.8) | 3 (9.4) | <0.0001 |
| CIP | 52 (43.3) | 47 (77) | 3 (11.1) | 2 (6.3) | <0.0001 | |
| STR | Aminoglycosides | 79 (65.8) | 57 (93.4) | 11 (40.7) | 11 (34.4) | <0.0001 |
| GEN | 49 (40.8) | 45 (73.8) | 3 (11.1) | 1 (3.1) | <0.0001 | |
| SXT | Folate pathway inhibitors | 59 (49.2) | 39 (63.9) | 12 (44.4) | 8 (25) | 0.0014 |
| FIS | 82 (68.3) | 58 (95.1) | 13 (48.1) | 11 (34.4) | <0.0001 | |
| AZI | Macrolides | 37 (30.8) | 31 (50.8) | 3 (11.1) | 3 (9.4) | <0.0001 |
| TET | Tetracyclines | 82 (68.3) | 59 (96.7) | 12 (44.4) | 11 (34.4) | <0.0001 |
| CHL | Phenicols | 73 (60.8) | 57 (93.4) | 11 (40.7) | 5 (15.6) | <0.0001 |
| MERO | Carbapenems | 0 | 0 | 0 | 0 | NA |
| Multidrug resistance (MDR) No. of isolates (%) | 83 (69.1) | 60 (98.4) | 12 (44.4) | 11 (34.4) | <0.0001 | |
| Resistance Gene | Total (n = 120) No. (%) | Chicken (n = 61) No. (%) | Beef (n = 27) No. (%) | Human (n = 32) No. (%) | p-Value |
|---|---|---|---|---|---|
| blaTEM | 75 (62.5) | 51 (83.6) | 9 (33.3) | 15 (46.9) | <0.0001 |
| blaCTX | 47 (39.2) | 39 (63.9) | 2 (7.4) | 6 (18.8) | <0.0001 |
| blaCMY | 12 (10) | 9 (14.8) | 1 (3.7) | 2 (6.3) | 0.1998 |
| blaSHV | 8 (6.7) | 7 (11.5) | 1 (3.7) | 0 | 0.0849 |
| blaOXA | 2 (1.7) | 0 | 0 | 2 (6.3) | 0.061 |
| cat1 | 18 (15) | 15 (24.6) | 1 (3.7) | 2 (6.3) | 0.011 |
| cat2 | 17 (14.2) | 14 (23) | 1 (3.7) | 2 (6.3) | 0.0188 |
| floR | 64 (53.3) | 53 (86.9) | 9 (33.3) | 2 (6.3) | <0.0001 |
| tetA | 77 (64.2) | 57 (93.4) | 11 (40.7) | 9 (28.1) | <0.0001 |
| tetB | 8 (6.7) | 5 (8.2) | 1 (3.7) | 2 (6.3) | 0.7337 |
| sul1 | 68 (56.7) | 41 (67.2) | 14 (51.9) | 13 (40.6) | 0.0413 |
| sul2 | 60 (50) | 40 (65.6) | 10 (37) | 10 (31.3) | 0.0022 |
| strA | 57 (47.5) | 39 (63.9) | 9 (33.3) | 9 (28.1) | 0.0011 |
| strB | 58 (48.3) | 40 (65.6) | 9 (33.3) | 9 (28.1) | 0.0006 |
| dhfr1 | 8 (6.7) | 7 (11.5) | 0(0) | 1 (3.1) | 0.0889 |
| dhfr5 | 14 (11.7) | 4 (6.6) | 7 (25.9) | 3 (9.4) | 0.0297 |
| dhfr12 | 31 (25.8) | 24 (39.3) | 5 (18.5) | 2 (6.3) | 0.0015 |
| dhfr13 | 31 (25.8) | 25 (41) | 4 (14.8) | 2 (6.3) | 0.0004 |
| mphA | 37 (30.8) | 32 (52.5) | 2 (7.4) | 3 (9.4) | <0.0001 |
| Phylogroup | Total (n = 120) No. (%) | Chicken (n = 61) No. (%) | Beef (n = 27) No. (%) | Human (n = 32) No. (%) | p-Value |
|---|---|---|---|---|---|
| A | 43 (35.8) | 15 (24.6) | 8 (29.6) | 20 (62.5) | 0.0011 |
| B1 | 38 (31.7) | 16 (26.2) | 13 (48.1) | 9 (28.1) | 0.1103 |
| C | 13 (10.8) | 12 (19.7) | 1 (3.7) | 0 | 0.006 |
| B2 | 5 (4.2) | 1 (1.6) | 4 (14.8) | 0 | 0.01 |
| D | 2 (1.7) | 0 | 0 | 2 (6.3) | 0.1186 |
| E | 12 (10) | 10 (16.4) | 1 (3.7) | 1 (3.1) | 0.0596 |
| F | 6 (5) | 6 (9.8) | 0 | 0 | 0.05 |
| Clade I | 0 | 0 | 0 | 0 | NA |
| U * | 1 (0.83) | 1 (1.6) | 0 | 0 | 0.99 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramadan, H.; Jackson, C.R.; Frye, J.G.; Hiott, L.M.; Samir, M.; Awad, A.; Woodley, T.A. Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt. Pathogens 2020, 9, 357. https://doi.org/10.3390/pathogens9050357
Ramadan H, Jackson CR, Frye JG, Hiott LM, Samir M, Awad A, Woodley TA. Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt. Pathogens. 2020; 9(5):357. https://doi.org/10.3390/pathogens9050357
Chicago/Turabian StyleRamadan, Hazem, Charlene R. Jackson, Jonathan G. Frye, Lari M. Hiott, Mohamed Samir, Amal Awad, and Tiffanie A. Woodley. 2020. "Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt" Pathogens 9, no. 5: 357. https://doi.org/10.3390/pathogens9050357
APA StyleRamadan, H., Jackson, C. R., Frye, J. G., Hiott, L. M., Samir, M., Awad, A., & Woodley, T. A. (2020). Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt. Pathogens, 9(5), 357. https://doi.org/10.3390/pathogens9050357