Airway Epithelial Derived Cytokines and Chemokines and Their Role in the Immune Response to Respiratory Syncytial Virus Infection


 and
and
Abstract
1. Introduction
2. Methods of Methodological Review
3. Airway Epithelial Immune Response to RSV Infection
3.1. Epithelial Barrier Functions and Virus Infection
3.2. Sensory Function of Epithelial Cells
3.3. Epithelium-Derived Inflammatory Mediators during RSV Infection
3.4. Secretion of Inflammatory Mediators
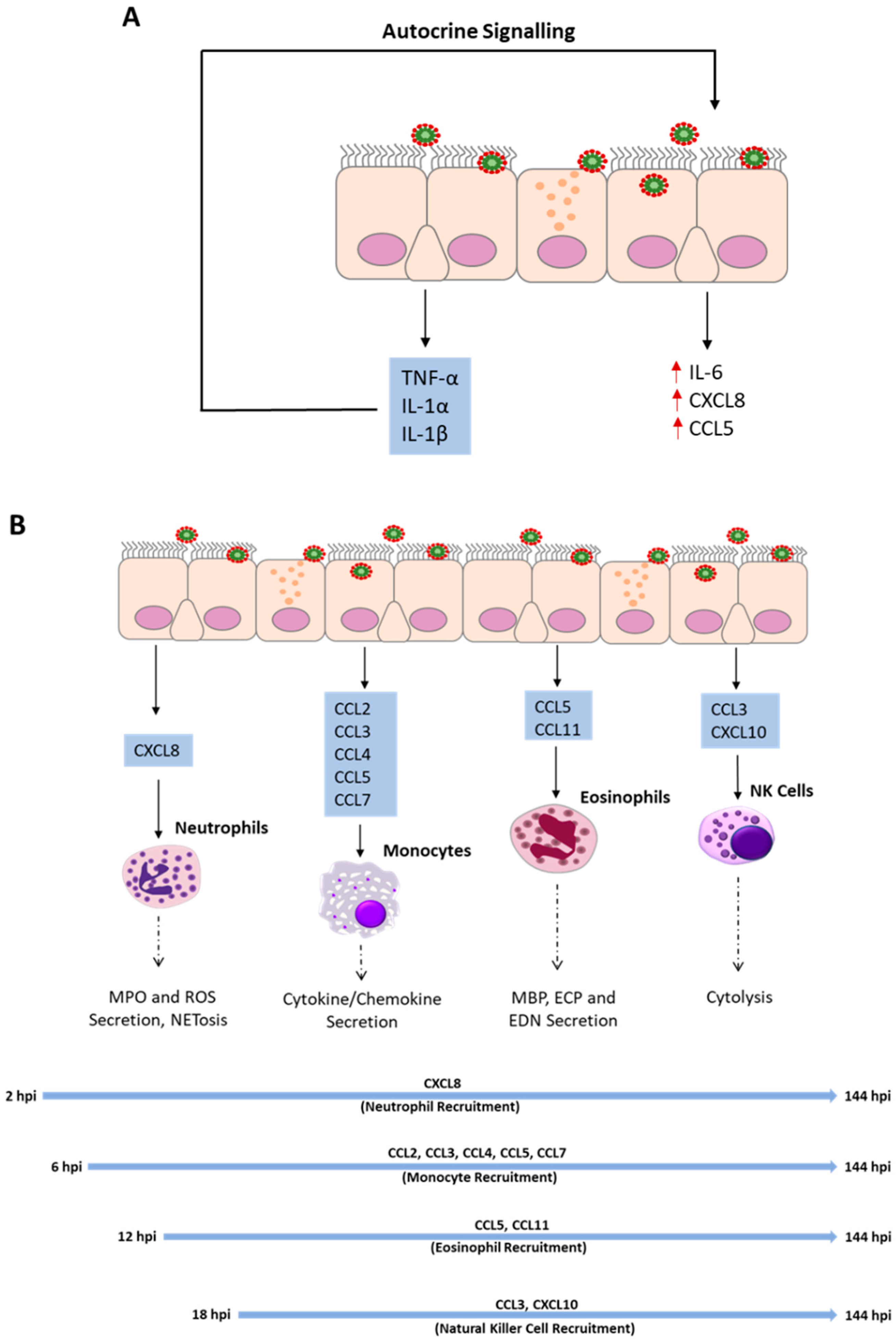
3.5. Autocrine Signaling Amplifies Inflammatory Mediator Release
3.6. IFNs Amplify Cytokine and Chemokine Release from Epithelial Cells
3.7. Peripheral Blood Cell and Epithelial Cell Interactions
3.8. Differential Secretion of Cytokines/Chemokines Following Infection with Common Respiratory Viruses
4. Immune Cell Recruitment to the Airway Epithelium
4.1. Neutrophils
4.2. Monocytes
4.3. Eosinophils
4.4. Natural Killer Cells
5. Models of RSV Infection
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef]
- McNamara, P.S.; Smyth, R.L. The pathogenesis of respiratory syncytial virus disease in childhood. Br. Med. Bull. 2002, 61, 13–28. Available online: https://academic.oup.com/bmb/article/61/1/13/286884 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Johnson, J.E.; Gonzales, R.A.; Olson, S.J.; Wright, P.F.; Graham, B.S. The histopathology of fatal untreated human respiratory syncytial virus infection. Mod. Pathol. 2007, 20, 108–119. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17143259 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Touzelet, O.; Power, U.F. Cellular and molecular characteristics of RSV-induced disease in humans. Hum. Respir. Syncytial Virus Infect. 2011, 97–122. Available online: http://www.intechopen.com/books/human-respiratory-syncytial-virus-infection/cellular-and-molecular-characteristics-of-rsv-induced-disease-in-humans (accessed on 16 July 2019).
- Dou, Y.; Zhao, Y.; Zhang, Z.; Mao, H.; Tu, W.; Zhao, X. Respiratory Syncytial Virus Infection Induces Higher Toll-Like Receptor-3 Expression and TNF-α Production Than Human Metapneumovirus Infection. PLoS ONE 2013, 8, 1–10. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3767791/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Guo Parke, H.; Canning, P.; Douglas, I.; Villenave, R.; Heaney, L.G.; Coyle, P.V.; Lyons, J.D.; Shields, M.D.; Power, U.F. Relative respiratory syncytial virus cytopathogenesis in upper and lower respiratory tract epithelium. Am. J. Respir. Crit. Care Med. 2013, 188, 842–851. Available online: https://www.atsjournals.org/doi/full/10.1164/rccm.201304-0750OC (accessed on 16 July 2019). [CrossRef] [PubMed]
- Villenave, R.; Thavagnanam, S.; Sarlang, S.; Parker, J.; Douglas, I.; Skibinski, G.; Heaney, L.G.; McKaigue, J.P.; Coyle, P.V.; Shields, M.D.; et al. In vitro modeling of respiratory syncytial virus infection of pediatric bronchial epithelium, the primary target of infection in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 5040–5045. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Peeples, M.E.; Boucher, R.C.; Collins, P.L.; Pickles, R.J. Respiratory syncytial virus infection of human airway epithelial cells is polarized, specific to ciliated cells, and without obvious cytopathology. J. Virol. 2002, 76, 5654–5666. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11991994 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Ioannidis, I.; McNally, B.; Willette, M.; Peeples, M.E.; Chaussabel, D.; Durbin, J.E.; Ramilo, O.; Mejias, A.; Flaño, E. Plasticity and Virus Specificity of the Airway Epithelial Cell Immune Response during Respiratory Virus Infection. J. Virol. 2012, 86, 5422–5436. [Google Scholar] [CrossRef]
- Korell, J.; Green, B.; DeVincenzo, J.; Huntjens, D. A human challenge model for respiratory syncytial virus kinetics, the pharmacological effect of a novel fusion inhibitor, and the modelling of symptoms scores. Eur. J. Pharm. Sci. 2017, 109, S154–S160. [Google Scholar] [CrossRef]
- Russell, C.D.; Unger, S.A.; Walton, M.; Schwarze, J. The Human Immune Response to Respiratory Syncytial Virus Infection. Clin. Microbiol. Rev. 2017, 30, 481–502. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5355638/ (accessed on 16 July 2019). [CrossRef]
- Vareille, M.; Kieninger, E.; Edwards, M.R.; Regamey, N. The airway epithelium: Soldier in the fight against respiratory viruses. Clin. Microbiol. Rev. 2011, 24, 210–229. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3021210/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Hillyer, P.; Shepard, R.; Uehling, M.; Krenz, M.; Sheikh, F.; Thayer, K.R.; Huang, L.; Yan, L.; Panda, D.; Luongo, C.; et al. Differential responses by human respiratory epithelial cell lines to respiratory syncytial virus reflect distinct patterns of infection control. J. Virol. 2018, 92, e02202–e02217. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.A.; Kunimoto, M.; Sim, T.C.; Garofalo, R.; Eliott, T.; Baron, S.; Ruuskanen, O.; Chonmaitree, T.; Ogra, P.L.; Schmalstieg, F. Interleukin-1 alpha mediates the enhanced expression of intercellular adhesion molecule-1 in pulmonary epithelial cells infected with respiratory syncytial virus. Am. J. Respir. Cell Mol. Biol. 1995, 13, 602–609. Available online: http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=emed6&NEWS=N&AN=125137762 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Patel, J.A.; Jiang, Z.; Nakajima, N.; Kunimoto, M. Autocrine regulation of interleukin-8 by interleukin-1α in respiratory syncytial virus-infected pulmonary epithelial cells in vitro. Immunology 1998, 95, 501–506. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1364344/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Oshansky, C.M.; Barber, J.P.; Crabtree, J.; Tripp, R.A. Respiratory syncytial virus F and G proteins induce interleukin 1alpha, CC, and CXC chemokine responses by normal human bronchoepithelial cells. J. Infect. Dis. 2010, 201, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Foronjy, R.F.; Dabo, A.J.; Cummins, N.; Geraghty, P. Leukemia inhibitory factor protects the lung during respiratory syncytial viral infection. BMC Immunol. 2014, 15, 1–15. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4189665/ (accessed on 16 July 2019). [CrossRef]
- Di Paolo, N.C.; Shayakhmetov, D.M. Immunol Author manuscript N. Interleukin 1α and the inflammatory process. Nat. Immunol. 2016, 17, 906–913. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5152572/ (accessed on 16 July 2019). [CrossRef]
- Dinarello, C.A. Immunological and Inflammatory Functions of the Interleukin-1 Family. Annu. Rev. Immunol. 2009, 27, 519–550. Available online: https://www.annualreviews.org/doi/abs/10.1146/annurev.immunol.021908.132612?rfr_dat=cr_pub%3Dpubmed&url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&journalCode=immunol (accessed on 16 July 2019). [CrossRef]
- Noah, T.L.; Becker, S. Respiratory syncytial virus-induced cytokine production by a human bronchial epithelial cell line. Am. J. Physiol. Cell Mol. Physiol. 1993, 265, L472–L478. Available online: http://www.physiology.org/doi/10.1152/ajplung.1993.265.5.L472 (accessed on 16 July 2019). [CrossRef]
- Villenave, R.; O'Donoghue, D.; Thavagnanam, S.; Touzelet, O.; Skibinski, G.; Heaney, L.G.; McKaigue, J.P.; Coyle, P.V.; Shields, M.D.; Power, U.F. Differential cytopathogenesis of respiratory syncytial virus prototypic and clinical isolates in primary pediatric bronchial epithelial cells. Virol. J. 2011, 8, 43. Available online: http://www.virologyj.com/content/8/1/43 (accessed on 16 July 2019). [CrossRef]
- Hackett, T.L.; Singhera, G.K.; Shaheen, F.; Hayden, P.; Jackson, G.R.; Hegele, R.G.; Van Eeden, S.; Bai, T.R.; Dorscheid, D.R.; Knight, D.A. Intrinsic phenotypic differences of asthmatic epithelium and its inflammatory responses to respiratory syncytial virus and air pollution. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1090–1100. Available online: https://www.atsjournals.org/doi/full/10.1165/rcmb.2011-0031OC?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub%3Dpubmed (accessed on 16 July 2019). [CrossRef] [PubMed]
- Fonceca, A.M.; Flanagan, B.F.; Trinick, R.; Smyth, R.L.; McNamara, P.S. Primary airway epithelial cultures from children are highly permissive to respiratory syncytial virus infection. Thorax 2012, 67, 42–48. Available online: https://thorax.bmj.com/content/67/1/42.long (accessed on 16 July 2019). [CrossRef] [PubMed][Green Version]
- Becker, S.; Reed, W.; Henderson, F.W.; Noah, T.L. RSV infection of human airway epithelial cells causes production of the beta-chemokine RANTES. Am. J. Physiol. Cell Mol. Physiol. 1997, 272, L512–L520. Available online: http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=emed4&NEWS=N&AN=1997102900 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Tian, B.; Yang, J.; Zhao, Y.; Ivanciuc, T.; Sun, H.; Wakamiya, M.; Garofalo, R.P.; Brasier, A.R. Central Role of the NF-κB Pathway in the Scgb1a1-Expressing Epithelium in Mediating Respiratory Syncytial Virus-Induced Airway Inflammation. J. Virol. 2018, 92, 1–19. Available online: http://www.ncbi.nlm.nih.gov/pubmed/29593031 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Jiang, Z.; Kunimoto, M. Autocrine regulation and experimental modulation of interleukin-6 expression by human pulmonary epithelial cells infected with respiratory syncytial virus. J. Virol. 1998, 72, 2496–2499. Available online: http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=emed6&NEWS=N&AN=28100812 (accessed on 16 July 2019). [PubMed]
- Schijf, M.A.; Lukens, M.V.; Kruijsen, D.; van Uden, N.O.; Garssen, J.; Coenjaerts, F.E.; van’t Land, B.; van Bleek, G.M. Respiratory syncytial virus induced type I IFN production by pDC is regulated by RSV-infected airway epithelial cells, RSV-exposed monocytes and virus specific antibodies. PLoS ONE 2013, 8, 1–18. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3841124/ (accessed on 16 July 2019). [CrossRef] [PubMed][Green Version]
- Arnold, R.; Humbert, B.; Werchau, H.; Gallati, H.; König, W. Interleukin-8, interleukin-6, and soluble tumour necrosis factor receptor type I release from a human pulmonary epithelial cell line (A549) exposed to respiratory syncytial virus. Immunology 1994, 82, 126–133. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7519169%0Ahttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC1414862 (accessed on 16 July 2019).
- Levitz, R.; Wattier, R.; Phillips, P.; Solomon, A.; Lawler, J.; Lazar, I.; Weibel, C.; Kahn, J.S. Induction of IL-6 and CCL5 (RANTES) in human respiratory epithelial (A549) cells by clinical isolates of respiratory syncytial virus is strain specific. Virol. J. 2012, 9, 190. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22962966 (accessed on 16 July 2019). [CrossRef]
- Das, S.; Palmer, O.P.; Leight, W.D.; Surowitz, J.B.; Pickles, R.J.; Randell, S.H.; Buchman, C.A. Cytokine amplification by respiratory syncytial virus infection in human nasal epithelial cells. Laryngoscope 2005, 115, 764–768. Available online: https://onlinelibrary.wiley.com/doi/full/10.1097/01.MLG.0000159527.76949.93 (accessed on 16 July 2019). [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro-and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 878–888. Available online: https://www.sciencedirect.com/science/article/pii/S0167488911000425 (accessed on 16 July 2019). [CrossRef]
- Lee, C.G.; Yoon, H.J.; Zhu, Z.; Link, H.; Wang, Z.; Gwaltney, J.M., Jr.; Landry, M.; Elias, J.A. Respiratory Syncytial Virus Stimulation of Vascular Endothelial Cell Growth Factor/Vascular Permeability Factor. Am. J. Respir. Cell Mol. Biol. 2000, 23, 662–669. Available online: http://www.atsjournals.org/doi/abs/10.1165/ajrcmb.23.5.4188 (accessed on 16 July 2019). [CrossRef] [PubMed][Green Version]
- Alkharsah, K.R. VEGF upregulation in viral infections and its possible therapeutic implications. Int. J. Mol. Sci. 2018, 19, 1642. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6032371/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Dosanjh, A.; Rednam, S.; Martin, M. Respiratory syncytial virus augments production of fibroblast growth factor basic in vitro: Implications for a possible mechanism of prolonged wheezing after infection. Pediatr. Allergy Immunol. 2003, 14, 437–440. Available online: https://onlinelibrary.wiley.com/doi/full/10.1046/j.0905-6157.2003.00090.x?sid=nlm%3Apubmed (accessed on 16 July 2019). [CrossRef] [PubMed]
- Yun, Y.R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.H.; Shin, U.S.; Kim, H.W. Fibroblast growth factors: Biology, function, and application for tissue regeneration. J. Tissue Eng. 2010, 1, 218142. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3042641/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Hosakote, Y.M.; Brasier, A.R.; Casola, A.; Garofalo, R.P.; Kurosky, A. Respiratory Syncytial Virus Infection Triggers Epithelial HMGB1 Release as a Damage-Associated Molecular Pattern Promoting a Monocytic Inflammatory Response. Lyles DS, editor. J. Virol. 2016, 90, 9618–9631. [Google Scholar] [CrossRef] [PubMed]
- Eyles, J.L.; Roberts, A.W.; Metcalf, D.; Wicks, I.P. Granulocyte colony-stimulating factor and neutrophils-Forgotten mediators of inflammatory disease. Nat. Clin. Pract. Rheumatol. 2006. Available online: https://www.nature.com/articles/ncprheum0291 (accessed on 16 July 2019).
- Ishioka, T.; Kimural, H.; Kita, H.; Obuchi, M.; Hoshino, H.; Noda, M.; Nishina, A.; Kozawa, K.; Kato, M. Effects of respiratory syncytial virus infection and major basic protein derived from eosinophils in pulmonary alveolar epithelial cells (A549). Cell Biol. Int. 2011, 35, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Becher, B.; Tugues, S.; Greter, M. GM-CSF: From Growth Factor to Central Mediator of Tissue Inflammation. Immunity 2016. Available online: https://www.sciencedirect.com/science/article/pii/S107476131630437X?via%3Dihub (accessed on 16 July 2019).
- Kotelkin, A.; Prikhod’ko, E.A.; Cohen, J.I.; Collins, P.L.; Bukreyev, A. Respiratory syncytial virus infection sensitizes cells to apoptosis mediated by tumor necrosis factor-related apoptosis-inducing ligand. J. Virol. 2003, 77, 9156–9172. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12915532%5Cnhttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC187410/pdf/0484.pdf (accessed on 16 July 2019). [CrossRef]
- Falschlehner, C.; Schaefer, U.; Walczak, H. Following TRAIL’s path in the immune system. Immunology 2009, 127, 145–154. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2691779/ (accessed on 16 July 2019). [CrossRef]
- Masaki, T.; Kojima, T.; Okabayashi, T.; Ogasawara, N.; Ohkuni, T.; Obata, K.; Takasawa, A.; Murata, M.; Tanaka, S.; Hirakawa, S.; et al. A nuclear factor-B signaling pathway via protein kinase C regulates replication of respiratory syncytial virus in polarized normal human nasal epithelial cells. Mol. Biol. Cell 2011, 22, 2144–2156. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.R. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/path.2287 (accessed on 16 July 2019). [CrossRef]
- Villenave, R.; Broadbent, L.; Douglas, I.; Lyons, J.D.; Coyle, P.V.; Teng, M.N.; Tripp, R.A.; Heaney, L.G.; Shields, M.D.; Power, U.F. Induction and Antagonism of Antiviral Responses in Respiratory Syncytial Virus-Infected Pediatric Airway Epithelium. Perlman, S, Ed.; J. Virol. 2015, 89, 12309–12318. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12915532%5Cnhttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC187410/pdf/0484.pdf (accessed on 16 July 2019). [CrossRef] [PubMed]
- Okabayashi, T.; Kojima, T.; Masaki, T.; Yokota, S.I.; Imaizumi, T.; Tsutsumi, H.; Himi, T.; Fujii, N.; Sawada, N. Type-III interferon, not type-I, is the predominant interferon induced by respiratory viruses in nasal epithelial cells. Virus. Res. 2011, 160, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and Functions of the IL-10 Family of Cytokines in Inflammation and Disease. Annu. Rev. Immunol. 2011, 29, 71–109. Available online: https://www.annualreviews.org/doi/abs/10.1146/annurev-immunol-031210-101312?rfr_dat=cr_pub%3Dpubmed&url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&journalCode=immunol (accessed on 16 July 2019).
- McNamara, P.S.; Fonceca, A.M.; Howarth, D.; Correia, J.B.; Slupsky, J.R.; Trinick, R.E.; Al Turaiki, W.; Smyth, R.L.; Flanagan, B.F. Respiratory syncytial virus infection of airway epithelial cells, in vivo and in vitro, supports pulmonary antibody responses by inducing expression of the B cell differentiation factor BAFF. Thorax 2012, 68, 76–81. Available online: https://thorax.bmj.com/content/68/1/76.long (accessed on 16 July 2019). [CrossRef] [PubMed]
- Sakai, J.; Akkoyunlu, M. The role of BAFF system molecules in host response to pathogens. Clin. Microbiol. Rev. 2017, 30, 991–1014. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5608883/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Lee, H.C.; Headley, M.B.; Loo, Y.M.; Berlin, A.; Gale, M., Jr.; Debley, J.S.; Lukacs, N.W.; Ziegler, S.F. Thymic stromal lymphopoietin is induced by respiratory syncytial virus-infected airway epithelial cells and promotes a type 2 response to infection. J. Allergy Clin. Immunol. 2012, 130, 1187–1196. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4284103/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Zhao, Y.; Jamaluddin, M.; Zhang, Y.; Sun, H.; Ivanciuc, T.; Garofalo, R.P.; Brasier, A.R. Systematic Analerysis of Cell-Type Differences in the Epithelial Secretome Reveals Insights into the Pathogenesis of Respiratory Syncytial Virus–Induced Lower Respiratory Tract Infections. J. Immunol. 2017, 198, 3345–3364. Available online: http://www.jimmunol.org/lookup/doi/10.4049/jimmunol.1601291 (accessed on 16 July 2019). [CrossRef]
- Roan, F.; Bell, B.D.; Stoklasek, T.A.; Kitajima, M.; Han, H.; Ziegler, S.F. The multiple facets of thymic stromal lymphopoietin (TSLP) during allergic inflammation and beyond. J. Leukoc. Biol. 2012, 91, 877–886. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3360473/ (accessed on 16 July 2019).
- Gibbs, J.D.; Ornoff, D.M.; Igo, H.A.; Zeng, J.Y.; Imani, F. Cell Cycle Arrest by Transforming Growth Factor 1 Enhances Replication of Respiratory Syncytial Virus in Lung Epithelial Cells. J. Virol. 2009, 83, 12424–12431. [Google Scholar] [CrossRef]
- Xia, Y.C.; Radwan, A.; Keenan, C.R.; Langenbach, S.Y.; Li, M.; Radojicic, D.; Londrigan, S.L.; Gualano, R.C.; Stewart, A.G. Glucocorticoid Insensitivity in Virally Infected Airway Epithelial Cells Is Dependent on Transforming Growth Factor-β Activity. PLoS Pathog. 2017, 13, e1006138. Available online: https://dx.plos.org/10.1371/journal.ppat.1006138 (accessed on 16 July 2019). [CrossRef]
- Sanjabi, S.; Zenewicz, L.A.; Kamanaka, M.; Flavell, R.A. Anti-inflammatory and pro-inflammatory roles of TGF-β, IL-10, and IL-22 in immunity and autoimmunity. Curr. Opin. Pharmacol. 2009, 9, 447–453. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2755239/ (accessed on 16 July 2019). [CrossRef]
- Olszewska-Pazdrak, B.; Casola, A.; Saito, T.; Alam, R.; Crowe, S.E.; Mei, F.; Ogra, P.L.; Garofalo, R.P. Cell-specific expression of RANTES, MCP-1, and MIP-1alpha by lower airway epithelial cells and eosinophils infected with respiratory syncytial virus. J. Virol. 1998, 72, 4756–4764. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9573240%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC110009 (accessed on 16 July 2019).
- Carpenter, L.R.; Moy, J.N.; Roebuck, K.A. Respiratory syncytial virus and TNFalpha induction of chemokine gene expression involves differential activation of Rel A and NF-kappaB1. BMC Infect. Dis. 2002, 2, 1–9. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC102322/ (accessed on 16 July 2019). [CrossRef]
- Zhang, Y.; Luxon, B.A.; Casola, A.; Garofalo, R.P.; Jamaluddin, M.; Brasier, A.R. Expression of respiratory syncytial virus-induced chemokine gene networks in lower airway epithelial cells revealed by cDNA microarrays. J. Virol. 2001, 75, 9044–9058. Available online: http://jvi.asm.org/cgi/content/abstract/75/19/9044%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=114473&tool=pmcentrez&rendertype=abstract (accessed on 16 July 2019). [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and Chemokine Receptors: Positioning Cells for Host Defense and Immunity. Annu. Rev. Immunol. 2014, 32, 659–702. Available online: https://www.annualreviews.org/doi/abs/10.1146/annurev-immunol-032713-120145?rfr_dat=cr_pub%3Dpubmed&url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&journalCode=immunol (accessed on 16 July 2019). [CrossRef] [PubMed]
- Becker, S.; Soukup, J.M. Airway epitheliol cell-induced activation of monocytes and eosinophils in respiratory syncytial viral infection. Immunobiology 1999, 201, 88–106. [Google Scholar] [CrossRef]
- Harrison, A.M.; Bonville, C.A.; Rosenberg, H.F.; Domachowske, J.B. Respiratory Syncytical Virus–induced Chemokine Expression in the Lower Airways. Am. J. Respir. Crit. Care Med. 1999, 159, 1918–1924. [Google Scholar] [CrossRef]
- Chun, Y.H.; Park, J.Y.; Lee, H.; Kim, H.S.; Won, S.; Joe, H.J.; Chung, W.J.; Yoon, J.S.; Kim, H.H.; Kim, J.T.; et al. Rhinovirus-infected epithelial cells produce more IL-8 and rantes compared with other respiratory viruses. Allergy Asthma Immunol. Res. 2013, 5, 216–223. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3695236/ (accessed on 16 July 2019). [CrossRef] [PubMed]
- Pazdrak, K.; Olszewska-Pazdrak, B.; Liu, T.; Takizawa, R.; Brasier, A.R.; Garofalo, R.P.; Casola, A. MAPK activation is involved in posttranscriptional regulation of RSV-induced RANTES gene expression. Available online: https://www.ncbi.nlm.nih.gov/pubmed/12114198 (accessed on 16 July 2019).
- Saito, T.; Deskin, R.W.; Casola, A.; Häeberle, H.; Olszewska, B.; Ernst, P.B.; Alam, R.; Ogra, P.L.; Garofalo, R. Respiratory syncytial virus induces selective production of the chemokine RANTES by upper airway epithelial cells. J. Infect. Dis. 1997, 175, 497–504. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9041319 (accessed on 16 July 2019). [CrossRef]
- Mellow, T.E.; Murphy, P.C.; Carson, J.L.; Noah, T.L.; Zhang, L.; Pickles, R.J. The effect of respiratory synctial virus on chemokine release by differentiated airway epithelium. Exp. Lung Res. 2004, 30, 43–57. [Google Scholar] [CrossRef]
- Jung, S.Y.; Shin, S.Y.; Eun, Y.G.; Kim, S.W.; Cho, J.S. Changes of histamine receptors and CC chemokines in nasal epithelial cells and fibroblasts after respiratory syncytial virus infection. Am. J. Rhinol. Allergy 2013, 27, 17–21. [Google Scholar] [CrossRef]
- Fiedler, M.A.; Wernke-Dollries, K.; Stark, J.M. Respiratory syncytial virus increases IL-8 gene expression and protein release in A549 cells. Am. J. Physiol. Cell Mol. Physiol. 1995, 269, L865–L872. [Google Scholar] [CrossRef]
- Becker, S.; Koren, H.S.; Henke, D.C. Interleukin-8 expression in normal nasal epithelium and its modulation by infection with respiratory syncytial virus and cytokines tumor necrosis factor, interleukin-1, and interleukin-6. Am. J. Respir. Cell Mol. Biol. 1993, 8, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Monick, M.M.; Powers, L.S.; Hassan, I.; Groskreutz, D.; Yarovinsky, T.O.; Barrett, C.W.; Castilow, E.M.; Tifrea, D.; Varga, S.M.; Hunninghake, G.W. Respiratory syncytial virus synergizes with Th2 cytokines to induce optimal levels of TARC/CCL17. J. Immunol. 2007, 179, 1648–1658. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17641031 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Andersson, U.; Tracey, K.J. HMGB1 Is a Therapeutic Target for Sterile Inflammation and Infection. Annu. Rev. Immunol. 2011, 29, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Bustamante-Marin, X.M.; Ostrowski, L.E. Cilia and Mucociliary Clearance. Cold Spring Harb. Perspect. Biol. 2017. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27864314 (accessed on 16 July 2019).
- Smith, C.M.; Kulkarni, H.; Radhakrishnan, P.; Rutman, A.; Bankart, M.J.; Williams, G.; Hirst, R.A.; Easton, A.J.; Andrew, P.W.; O’Callaghan, C. Ciliary dyskinesia is an early feature of respiratory syncytial virus infection. Eur. Respir J. 2014, 43, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Loxham, M.; Davies, D.E. Phenotypic and genetic aspects of epithelial barrier function in asthmatic patients. J. Allergy Clin. Immunol. 2017, 139, 1736–1751. Available online: http://dx.doi.org/10.1016/j.jaci.2017.04.005 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Zhang, N.; Van Crombruggen, K.; Gevaert, E.; Bachert, C. Barrier function of the nasal mucosa in health and type-2 biased airway diseases. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 295–307. [Google Scholar] [CrossRef]
- Zhang, L.; Collins, P.L.; Lamb, R.A.; Pickles, R.J. Comparison of differing cytopathic effects in human airway epithelium of parainfluenza virus 5 (W3A), parainfluenza virus type 3, and respiratory syncytial virus. Virology 2011, 421, 67–77. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21986028 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Wong, J.Y.W.; Rutman, A.; O’Callaghan, C. Recovery of the ciliated epithelium following acute bronchiolitis in infancy. Thorax 2005, 60, 582–587. [Google Scholar] [CrossRef]
- Sun, Y.; López, C.B. The innate immune response to RSV: Advances in our understanding of critical viral and host factors. Vaccine 2017, 35, 481–488. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27686836 (accessed on 16 July 2019). [CrossRef]
- Weitnauer, M.; Mijošek, V.; Dalpke, A.H. Control of local immunity by airway epithelial cells. Mucosal. Immunol. 2016, 9, 287–298. Available online: http://www.ncbi.nlm.nih.gov/pubmed/26627458 (accessed on 16 July 2019). [CrossRef]
- Raz, E. Organ-specific regulation of innate immunity. Nat. Immunol. 2007, 8, 3–4. Available online: http://www.nature.com/articles/ni0107-3 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Ioannidis, I.; Ye, F.; McNally, B.; Willette, M.; Flaño, E. Toll-like receptor expression and induction of type I and type III interferons in primary airway epithelial cells. J. Virol. 2013, 87, 3261–3270. Available online: http://www.ncbi.nlm.nih.gov/pubmed/23302870 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Monick, M.M.; Yarovinsky, T.O.; Powers, L.S.; Butler, N.S.; Carter, A.B.; Gudmundsson, G.; Hunninghake, G.W. Respiratory syncytial virus up-regulates TLR4 and sensitizes airway epithelial cells to endotoxin. J. Biol. Chem. 2003, 278, 53035–53044. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14565959 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Kurt-Jones, E.A.; Popova, L.; Kwinn, L.; Haynes, L.M.; Jones, L.P.; Tripp, R.A.; Walsh, E.E.; Freeman, M.W.; Golenbock, D.T.; Anderson, L.J.; et al. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000, 1, 398–401. Available online: http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=emed8&NEWS=N&AN=33455132 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Groskreutz, D.J.; Monick, M.M.; Powers, L.S.; Yarovinsky, T.O.; Look, D.C.; Hunninghake, G.W. Respiratory Syncytial Virus Induces TLR3 Protein and Protein Kinase R, Leading to Increased Double-Stranded RNA Responsiveness in Airway Epithelial Cells. J. Immunol. 2006, 176, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Rudd, B.D.; Burstein, E.; Duckett, C.S.; Li, X.; Lukacs, N.W. Differential Role for TLR3 in Respiratory Syncytial Virus-Induced Chemokine Expression. J. Virol. 2005, 79, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Martin, J.F.; Garcia-Arevalo, M.C.; De Lejarazu, R.O.; Ardura, J.; Eiros, J.M.; Alonso, A.; Matías, V.; Pino, M.; Bernardo, D.; Arranz, E.; et al. Predominance of Th2 cytokines, CXC chemokines and innate immunity mediators at the mucosal level during severe respiratory syncytial virus infection in children. Eur. Cytokine Netw. 2007, 18, 162–167. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17823085 (accessed on 16 July 2019).
- Murai, H.; Terada, A.; Mizuno, M.; Asai, M.; Hirabayashi, Y.; Shimizu, S.; Morishita, T.; Kakita, H.; Hussein, M.H.; Ito, T.; et al. IL-10 and RANTES are Elevated in Nasopharyngeal Secretions of Children with Respiratory Syncytial Virus Infection. Allergol. Int. 2007, 56, 157–163. [Google Scholar] [CrossRef]
- Hornsleth, A.; Loland, L.; Larsen, L.B. Cytokines and chemokines in respiratory secretion and severity of disease in infants with respiratory syncytial virus (RSV) infection. J. Clin. Virol. 2001, 21, 163–170. Available online: http://www.sciencedirect.com/science/article/pii/S1386653201001597 (accessed on 16 July 2019). [CrossRef]
- Garofalo, R.P.; Olszewska-Pazdrak, B.; Ogra, P.L.; Welliver, R.C. Beta-Chemokines in Nasal Secretions of Infants with Respiratory Syncytial Virus-Induced Respiratory Infections. Pediatr. Asthma Allergy Immunol. 2002, 15, 89–96. [Google Scholar] [CrossRef]
- Kerrin, A.; Fitch, P.; Errington, C.; Kerr, D.; Waxman, L.; Riding, K.; Mehendele, F.; McSorley, H.; MacKenzie, K.; Wronski, S. Differential lower airway dendritic cell patterns May reveal distinct endotypes of RSV bronchiolitis. Thorax 2017, 72, 620–627. Available online: https://thorax.bmj.com/content/72/7/620.long (accessed on 16 July 2019). [CrossRef] [PubMed]
- Hull, J.; Thomson, A.; Kwiatkowski, D. Association of respiratory syncytial virus bronchiolitis with the interleukin 8 gene region in UK families. Thorax 2000, 55, 1023–1027. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11083887 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Gentile, D.A.; Doyle, W.J.; Zeevi, A.; Howe-Adams, J.; Kapadia, S.; Trecki, J.; Skoner, D.P. Cytokine gene polymorphisms moderate illness severity in infants with respiratory syncytial virus infection. Hum. Immunol. 2003, 64, 338–344. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12590978 (accessed on 16 July 2019). [CrossRef]
- Hattori, S.; Shimojo, N.; Mashimo, T.; Inoue, Y.; Ono, Y.; Kohno, Y.; Okamoto, Y.; Hata, A.; Suzuki, Y. Relationship between RANTES polymorphisms and respiratory syncytial virus bronchiolitis in a Japanese infant population. Jpn. J. Infect. Dis. 2011, 64, 242–245. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21617311 (accessed on 16 July 2019). [PubMed]
- Goritzka, M.; Makris, S.; Kausar, F.; Durant, L.R.; Pereira, C.; Kumagai, Y.; Culley, F.J.; Mack, M.; Akira, S.; Johansson, C. Alveolar macrophage-derived type I interferons orchestrate innate immunity to RSV through recruitment of antiviral monocytes. J. Exp. Med. 2015, 212, 699–714. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25897172 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Goritzka, M.; Durant, L.R.; Pereira, C.; Salek-Ardakani, S.; Openshaw, P.J.M.; Johansson, C. Alpha/beta interferon receptor signaling amplifies early proinflammatory cytokine production in the lung during respiratory syncytial virus infection. J. Virol. 2014, 88, 6128–6136. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24648449 (accessed on 16 July 2019). [CrossRef]
- Schwarze, J. Lung Dendritic cells in respiratory syncytial virus bronchiolitis. Pediatric Infect. Dis. J. 2008, 10, S89–S91. Available online: https://www.ncbi.nlm.nih.gov/pubmed/18820586 (accessed on 16 July 2019). [CrossRef] [PubMed]
- McDermott, D.S.; Weiss, K.A.; Knudson, C.J.; Varga, S.M. Central role of dendritic cells in shaping the adaptive immune response during respiratory syncytial virus infection. Future Virol. 2011, 6, 963–973. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21887154 (accessed on 16 July 2019). [CrossRef]
- Openshaw, P.J.M.; Tregoning, J.S. Immune responses and disease enhancement during respiratory syncytial virus infection. Clin Microbiol. Rev. 2005, 18, 541–555. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16020689 (accessed on 16 July 2019). [CrossRef]
- Tregoning, J.S.; Pribul, P.K.; Pennycook, A.M.J.; Hussell, T.; Wang, B.; Lukacs, N.; Schwarze, J.; Culley, F.J.; Openshaw, P.J. The chemokine MIP1alpha/CCL3 determines pathology in primary RSV infection by regulating the balance of T cell populations in the murine lung. PLoS ONE 2010, 5, e9381. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20195359 (accessed on 16 July 2019). [CrossRef]
- McNamara, P.S.; Ritson, P.; Selby, A.; Hart, C.A.; Smyth, R.L. Bronchoalveolar lavage cellularity in infants with severe respiratory syncytial virus bronchiolitis. Arch. Dis. Child. 2003, 88, 922–926. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14500316 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Herold, S.; von Wulffen, W.; Steinmueller, M.; Pleschka, S.; Kuziel, W.A.; Mack, M.; Srivastava, M.; Seeger, W.; Maus, U.A.; Lohmeyer, J. Alveolar epithelial cells direct monocyte transepithelial migration upon influenza virus infection: Impact of chemokines and adhesion molecules. J. Immunol. 2006, 177, 1817–1824. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16849492 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Wang, S.Z.; Hallsworth, P.G.; Dowling, K.D.; Alpers, J.H.; Bowden, J.J.; Forsyth, K.D. Adhesion molecule expression on epithelial cells infected with respiratory syncytial virus. Eur. Respir. J. 2000, 15, 358–366. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10706505 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Bao, X.; Sinha, M.; Liu, T.; Hong, C.; Luxon, B.A.; Garofalo, R.P.; Casola, A. Identification of human metapneumovirus-induced gene networks in airway epithelial cells by microarray analysis. Virology 2008, 374, 114–127. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18234263 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Laham, F.R.; Israele, V.; Casellas, J.M.; Garcia, A.M.; Lac Prugent, C.M.; Hoffman, S.J.; Hauer, D.; Thumar, B.; Name, M.I.; Pascual, A.; et al. Differential production of inflammatory cytokines in primary infection with human metapneumovirus and with other common respiratory viruses of infancy. J. Infect. Dis. 2004, 189, 2047–2056. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15143472 (accessed on 16 July 2019). [CrossRef]
- Guerrero-Plata, A.; Casola, A.; Garofalo, R.P. Human Metapneumovirus Induces a Profile of Lung Cytokines Distinct from That of Respiratory Syncytial Virus. J. Virol. 2005, 79, 14992–14997. [Google Scholar] [CrossRef]
- Malmo, J.; Moe, N.; Krokstad, S.; Ryan, L.; Loevenich, S.; Johnsen, I.B.; Espevik, T.; Nordbø, S.A.; Døllner, H.; Anthonsen, M.W. Cytokine profiles in human metapneumovirus infected children: Identification of genes involved in the antiviral response and pathogenesis. PLoS ONE. 2016, 11, 1–15. [Google Scholar] [CrossRef]
- Rajan, D.; McCracken, C.E.; Kopleman, H.B.; Kyu, S.Y.; Lee, F.E.-H.; Lu, X.; Anderson, L.J. Human rhinovirus induced cytokine/chemokine responses in human airway epithelial and immune cells. PLoS ONE 2014, 9, e114322. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25500821%0Ahttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC4264758 (accessed on 16 July 2019). [CrossRef]
- Schroth, M.K.; Grimm, E.; Frindt, P.; Galagan, D.M.; Konno, S.I.; Love, R.; Gern, J. ERhinovirus replication causes RANTES production in primary bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1999, 20, 1220–1228. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7615827 (accessed on 16 July 2019). [CrossRef]
- Griego, S.D.; Weston, C.B.; Adams, J.L.; Tal-Singer, R.; Dillon, S.B. Role of p38 mitogen-activated protein kinase in rhinovirus-induced cytokine production by bronchial epithelial cells. J. Immunol. 2000, 165, 5211–5220. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11046054 (accessed on 16 July 2019). [CrossRef]
- Everard, M.L.; Swarbrick, A.; Wrightham, M.; McIntyre, J.; Dunkley, C.; James, P.D.; Sewell, H.F.; Milner, A.D. Analysis of cells obtained by bronchial lavage of infants with respiratory syncytial virus infection. Arch. Dis. Child. 1994, 71, 428–432. Available online: http://www.ncbi.nlm.nih.gov/pubmed/7826113 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Openshaw, P.J.M.; Chiu, C.; Culley, F.J.; Johansson, C. Protective and Harmful Immunity to RSV Infection. Annu. Rev. Immunol. 2017, 35, 501–532. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28226227 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Geerdink, R.J.; Pillay, J.; Meyaard, L.; Bont, L. Neutrophils in respiratory syncytial virus infection: A target for asthma prevention. J. Allergy Clin. Immunol. 2015, 136, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Rajarathnam, K.; Schnoor, M.; Richardson, R.M.; Rajagopal, S. How do chemokines navigate neutrophils to the target site: Dissecting the structural mechanisms and signaling pathways. Cell Signal. 2019, 54, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.M.; Godding, V.; Sedgwick, J.B.; Busse, W.W. Respiratory syncytial virus infection enhances neutrophil and eosinophil adhesion to cultured respiratory epithelial cells. Roles of CD18 and intercellular adhesion molecule-1. J. Immunol. 1996, 156, 4774–4782. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8648124 (accessed on 16 July 2019). [PubMed]
- Deng, Y.; Herbert, J.A.; Smith, C.M.; Smyth, R.L. An in vitro transepithelial migration assay to evaluate the role of neutrophils in Respiratory Syncytial Virus (RSV) induced epithelial damage. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Bataki, E.L.; Evans, G.S.; Everard, M.L. Respiratory syncytial virus and neutrophil activation. Clin. Exp. Immunol. 2005, 140, 470–477. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15932508 (accessed on 16 July 2019). [CrossRef]
- Aherne, W.; Bird, T.; Court, S.D.; Gardner, P.S.; McQuillin, J. Pathological changes in virus infections of the lower respiratory tract in children. J. Clin. Pathol. 1970, 23, 7–18. Available online: http://www.ncbi.nlm.nih.gov/pubmed/4909103 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Funchal, G.A.; Jaeger, N.; Czepielewski, R.S.; Machado, M.S.; Muraro, S.P.; Stein, R.T.; Bonorino, C.B.; Porto, B.N. Respiratory syncytial virus fusion protein promotes TLR-4-dependent neutrophil extracellular trap formation by human neutrophils. PLoS ONE 2015, 10, 1–14. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0124082 (accessed on 16 July 2019). [CrossRef]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host Microbe 2012, 12, 109–116. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22817992 (accessed on 16 July 2019). [CrossRef]
- Abu-Harb, M.; Bell, F.; Finn, A.; Rao, W.H.; Nixon, L.; Shale, D.; Everard, M.L. IL-8 and neutrophil elastase levels in the respiratory tract of infants with RSV bronchiolitis. Eur. Respir. J. 1999, 14, 139–143. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10489841 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Jones, A.; Qui, J.M.; Bataki, E.; Elphick, H.; Ritson, S.; Evans, G.S.; Everard, M.L. Neutrophil survival is prolonged in the airways of healthy infants and infants with RSV bronchiolitis. Eur. Respir. J. 2002, 20, 651–657. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12358343 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Drescher, B.; Bai, F. Neutrophil in viral infections, friend or foe? Virus. Res. 2013, 171, 1–7. Available online: http://www.ncbi.nlm.nih.gov/pubmed/23178588 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Lindemans, C.A.; Coffer, P.J.; Schellens, I.M.M.; de Graaff, P.M.A.; Kimpen, J.L.L.; Koenderman, L. Respiratory Syncytial Virus Inhibits Granulocyte Apoptosis through a Phosphatidylinositol 3-Kinase and NF- B-Dependent Mechanism. J. Immunol. 2006, 176, 5529–5537. Available online: http://www.jimmunol.org/cgi/doi/10.4049/jimmunol.176.9.5529 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Coleman, M.C.; Plant, K.; Newton, S.; Hobson, L.; Whyte, K.B.M.; Everard, L.M. The Anti-Apoptotic Effect of Respiratory Syncytial Virus on Human Peripheral Blood Neutrophils is Mediated by a Monocyte Derived Soluble Factor. Open Virol. J. 2011, 5, 114–123. Available online: http://benthamopen.com/ABSTRACT/TOVJ-5-114 (accessed on 16 July 2019). [CrossRef]
- Bohmwald, K.; Espinoza, J.A.; Pulgar, R.A.; Jara, E.L.; Kalergis, A.M. Functional Impairment of Mononuclear Phagocyte System by the Human Respiratory Syncytial Virus. Front. Immunol. 2017, 8, 1643. Available online: http://www.ncbi.nlm.nih.gov/pubmed/29230219 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Ahout, I.M.L.; Jans, J.; Haroutiounian, L.; Simonetti, E.R.; van der Gaast-de Jongh, C.; Diavatopoulos, D.A.; de Jonge, M.I.; de Groot, R.; Ferwerda, G. Reduced Expression of HLA-DR on Monocytes During Severe Respiratory Syncytial Virus Infections. Pediatr. Infect. Dis. J. 2016, 35, e89–e96. Available online: http://www.ncbi.nlm.nih.gov/pubmed/26658377 (accessed on 16 July 2019). [CrossRef]
- Morrison, P.T.; Thomas, L.H.; Sharland, M.; Friedland, J.S. RSV-infected airway epithelial cells cause biphasic up-regulation of CCR1 expression on human monocytes. J. Leukoc. Biol. 2007, 81, 1487–1495. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17389578 (accessed on 16 July 2019). [CrossRef]
- Garofalo, R.P.; Patti, J.; Hintz, K.A.; Hill, V.; Ogra, P.L.; Welliver, R.C. Macrophage Inflammatory Protein–1α (Not T Helper Type 2 Cytokines) Is Associated with Severe Forms of Respiratory Syncytial Virus Bronchiolitis. J. Infect. Dis. 2001, 184, 393–399. [Google Scholar] [CrossRef]
- McNamara, P.S.; Flanagan, B.F.; Hart, C.A.; Smyth, R.L. Production of chemokines in the lungs of infants with severe respiratory syncytial virus bronchiolitis. J. Infect Dis. 2005, 191, 1225–1232. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15776367 (accessed on 16 July 2019). [CrossRef]
- Soukup, J.M.; Becker, S. Role of monocytes and eosinophils in human respiratory syncytial virus infection in vitro. Clin. Immunol. 2003, 107, 178–185. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12804531 (accessed on 16 July 2019). [CrossRef]
- Garofalo, R.; Kimpen, J.L.; Welliver, R.C.; Ogra, P.L. Eosinophil degranulation in the respiratory tract during naturally acquired respiratory syncytial virus infection. J. Pediatr. 1992, 120, 28–32. Available online: http://www.ncbi.nlm.nih.gov/pubmed/1731020 (accessed on 16 July 2019). [CrossRef]
- Dimova-Yaneva, D.; Russell, D.; Main, M.; Brooker, R.J.; Helms, P.J. Eosinophil activation and cysteinyl leukotriene production in infants with respiratory syncytial virus bronchiolitis. Clin. Exp. Allergy 2004, 34, 555–558. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15080807 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Kimpen, J.L.; Garofalo, R.; Welliver, R.C.; Ogra, P.L. Activation of human eosinophils in vitro by respiratory syncytial virus. Pediatr. Res. 1992, 32, 160–164. Available online: http://www.ncbi.nlm.nih.gov/pubmed/1324464 (accessed on 16 July 2019). [CrossRef] [PubMed][Green Version]
- Domachowske, J.B.; Dyer, K.D.; Bonville, C.A.; Rosenberg, H.F. Recombinant human eosinophil-derived neurotoxin/RNase 2 functions as an effective antiviral agent against respiratory syncytial virus. J. Infect. Dis. 1998, 177, 1458–1464. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9607820 (accessed on 16 July 2019). [CrossRef]
- Domachowske, J.B.; Dyer, K.D.; Adams, A.G.; Leto, T.L.; Rosenberg, H.F. Eosinophil cationic protein/RNase 3 is another RNase A-family ribonuclease with direct antiviral activity. Nucleic Acids Res. 1998, 26, 3358–3363. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9649619 (accessed on 16 July 2019). [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Dyer, K.D.; Rosenberg, H.F. Evolution of antiviral activity in the ribonuclease A gene superfamily: Evidence for a specific interaction between eosinophil-derived neurotoxin (EDN/RNase 2) and respiratory syncytial virus. Nucleic Acids Res. 1998, 26, 5327–5332. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9826755 (accessed on 16 July 2019). [CrossRef]
- Welliver, R.C.; Duffy, L. The relationship of RSV-specific immunoglobulin E antibody responses in infancy, recurrent wheezing, and pulmonary function at age 7–8 years. Pediatr. Pulmonol. 1993, 15, 19–27. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8419894 (accessed on 16 July 2019). [CrossRef]
- Ye, Q.; Shao, W.-X.; Shang, S.-Q.; Pan, Y.-X.; Shen, H.-Q.; Chen, X.-J. Epidemiological characteristics and immune status of children with Respiratory Syncytial Virus. J. Med. Virol. 2015, 87, 323–329. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25123681 (accessed on 16 July 2019). [CrossRef]
- Kneyber, M.C.J.; Steyerberg, E.W.; de Groot, R.; Moll, H.A. Long-term effects of respiratory syncytial virus (RSV) bronchiolitis in infants and young children: A quantitative review. Acta Paediatr. 2000, 89, 654–660. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10914957 (accessed on 16 July 2019). [CrossRef]
- Trefny, P.; Stricker, T.; Baerlocher, C.; Sennhauser, F.H. Family history of atopy and clinical course of RSV infection in ambulatory and hospitalized infants. Pediatr. Pulmonol. 2000, 30, 302–306. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11015130 (accessed on 16 July 2019). [CrossRef]
- Comberiati, P.; Di Cicco, M.E.; D’Elios, S.; Peroni, D.G. How Much Asthma Is Atopic in Children? Front. Pediatr. 2017, 5, 122. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28603709 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Larrañaga, C.L.; Ampuero, S.L.; Luchsinger, V.F.; Carrión, F.A.; Aguilar, N.V.; Morales, P.R.; Palomino, M.A.M.; Tapia, L.F.; Avendaño, L.F. Impaired immune response in severe human lower tract respiratory infection by respiratory syncytial virus. Pediatr. Infect. Dis. J. 2009, 28, 867–873. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19738511 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Harker, J.A.; Godlee, A.; Wahlsten, J.L.; Lee, D.C.P.; Thorne, L.G.; Sawant, D.; Tregoning, J.S.; Caspi, R.R.; Bukreyev, A.; Collins, P.L.; et al. Interleukin 18 coexpression during respiratory syncytial virus infection results in enhanced disease mediated by natural killer cells. J. Virol. 2010, 84, 4073–4082. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhu, H.; Sun, R.; Wei, H.; Tian, Z. Natural Killer Cells Are Involved in Acute Lung Immune Injury Caused by Respiratory Syncytial Virus Infection. J. Virol. 2012, 86, 2251–2258. [Google Scholar] [CrossRef] [PubMed]
- Culley, F.J. Natural killer cells in infection and inflammation of the lung. Immunology. 2009, 128, 151–163. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19740372 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Zdrenghea, M.T.; Telcian, A.G.; Laza-Stanca, V.; Bellettato, C.M.; Edwards, M.R.; Nikonova, A.; Khaitov, M.R.; Azimi, N.; Groh, V.; Mallia, P.; et al. RSV infection modulates IL-15 production and MICA levels in respiratory epithelial cells. Eur. Respir. J. 2012, 39, 712–720. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21852331 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Bauer, S. Activation of natural killer cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science (80- ). 1999, 285, 727–730. [Google Scholar] [CrossRef]
- Groh, V.; Wu, J.; Yee, C.; Spies, T. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature 2002, 419, 734–738. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12384702 (accessed on 16 July 2019). [CrossRef]
- Bem, R.A.; Domachowske, J.B.; Rosenberg, H.F. Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Cell Mol. Physiol. 2011, 301, L148–L156. Available online: http://www.physiology.org/doi/10.1152/ajplung.00065.2011 (accessed on 16 July 2019). [CrossRef]
- Clarke, C.J.; Watt, N.J.; Meredith, A.; McIntyre, N.; Burns, S.M. Respiratory syncytial virus-associated bronchopneumonia in a young chimpanzee. J. Comp. Pathol. 1994, 110, 207–212. Available online: http://www.ncbi.nlm.nih.gov/pubmed/8040386 (accessed on 16 July 2019). [CrossRef]
- Unwin, S.; Chatterton, J.; Chantrey, J. Management of severe respiratory tract disease caused by human respiratory syncytial virus and Streptococcus pneumoniae in captive chimpanzees (Pan troglodytes). J. Zoo Wildl Med. 2013, 44, 105–115. Available online: http://www.ncbi.nlm.nih.gov/pubmed/23505710 (accessed on 16 July 2019). [CrossRef] [PubMed]
- DeVincenzo, J.P.; Wilkinson, T.; Vaishnaw, A.; Cehelsky, J.; Meyers, R.; Nochur, S.; Harrison, L.; Meeking, P.; Mann, A.; Moane, E.; et al. Viral load drives disease in humans experimentally infected with respiratory syncytial virus. Am. J. Respir Crit. Care Med. 2010, 182, 1305–1314. Available online: http://www.ncbi.nlm.nih.gov/pubmed/20622030 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Pickles, R.J. Human Airway Epithelial Cell Cultures for Modeling Respiratory Syncytial Virus Infection. Current Topics in Microbiology and Immunology. 2013, pp. 371–387. Available online: http://link.springer.com/10.1007/978-3-642-38919-1_19 (accessed on 16 July 2019).
- Thavagnanam, S.; Parker, J.C.; McBrien, M.E.; Skibinski, G.; Shields, M.D.; Heaney, L.G. Nasal epithelial cells can act as a physiological surrogate for paediatric asthma studies. PLoS ONE 2014, 9, e85802. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24475053 (accessed on 16 July 2019). [CrossRef] [PubMed]
- McDougall, C.M.; Blaylock, M.G.; Douglas, J.G.; Brooker, R.J.; Helms, P.J.; Walsh, G.M. Nasal epithelial cells as surrogates for bronchial epithelial cells in airway inflammation studies. Am. J. Respir. Cell Mol. Biol. 2008, 39, 560–568. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18483420 (accessed on 16 July 2019). [CrossRef] [PubMed]
- Nawroth, J.C.; Barrile, R.; Conegliano, D.; van Riet, S.; Hiemstra, P.S.; Villenave, R. Stem cell-based Lung-on-Chips: The best of both worlds? Adv. Drug Deliv. Rev. 2018. Available online: https://doi.org/10.1016/j.addr.2018.07.005 (accessed on 16 July 2019).


{kind=link}
| Name | Cell Culture Model | Known Functions in Inflammation | Interactions with Other Cells | Secretion Time Point (hpi) |
|---|---|---|---|---|
| IL-1α/FAF | WD-hAECs, A549, SAEs, A549, and BEAS-2B [9,13,14,15,16,17] | Pleiotropic cytokine; initiates self-perpetuating inflammatory responses; known pyrogen; induces hyperalgesia and vasoconstriction [18] | Induces the secretion of cytokines and chemokines from tissue cells and lymphocytes via IL-1R1 signaling [18] | 12 [16] 24 [17] 48 [9,13,14,15] |
| IL-1β | A549 and SAEs [14,15,17] | Pleiotropic cytokine, initiates self-perpetuating inflammatory responses; known pyrogen; induces hyperalgesia and vasodilation [19] | Induces the secretion of cytokines and chemokines from tissue cells and lymphocytes via IL-1R1 signaling [19] | 24 [17] 48 [14,15] |
| IL-6 | WD-HAECs, A549, SAEs, BEAS-2B, hAECs [6,7,9,13,17,20,21,22,23,24,25,26,27,28,29,30] | Pleiotropic cytokine; induces acute phase protein secretion, influences T- and B-cell growth and maturation [31] | Induces the secretion of inflammatory mediators from tissue and immune cells via IL6R signaling [31] | 6 [28] 12 [23] 24 [6,7,17,26,27,28,29] 48 [9,13,26,30] 72 [24] 96 [6,7,20,21,22] 120 [7] 144 [7] |
| VEGF | A549, SAEs, hAECs [17,32] | Angiogenesis and vascular remodelling [33] | Activates endothelial cells [33] | 6 [32] 24 [17,32] 48 [32] |
| FGF | A549 and SAEs [17,34] | Pleiotropic growth factor involved in tissue repair and regeneration; associated with cell proliferation and angiogenesis [35] | Activates endothelial cells and tissue cells expressing Fibroblast growth factor receptor (FGFR) to induce angiogenesis and proliferation [35] | 24 [17,34] |
| G-CSF | WD-hAECs, A549, and SAEs [9,17,36] | Immunostimulation of neutrophils; immunosuppression of myelomonocytic cells [37] | Regulates neutrophil development and survival; modifies cytokine production from neutrophils, monocytes, macrophages, and DCs [37] | 24 [17,36] 48 [9] |
| GM-CSF | WD-hAECs and A549 [17,20,22,38] | Immunostimulation of myelomonocytic cells [39] | Activates all myelomonocytic cells that express the GM-CSF receptor and promotes their survival and differentiation [39] | 24 [17] 48 [38] 96 [20,22] |
| TRAIL | WD-hAECs, A549, and SAEs [6,7,17,40] | Induces apoptosis in virus-infected cells and tumor cells; implicated in the regulation of T-cell homeostasis [41] | Interact with infected or tumorigenic tissue cells that express TRAIL receptors DR4 and DR5, influences the expansion and maturation of CD4+ and CD8+ T-cells via DR4 and DR5 signaling [41] | 24 [17] 96 [6,7] 120 [7] 144 [7] |
| TNF-α | WD-hAECs, A549, and SAEs [9,14,15,17,27,42] | Pleiotropic cytokine; promotes leukocyte extravasation; known pyrogen, promotes vasodilation, involved in the regulation of the coagulation cascade [43] | Promotes inflammation by interacting with TNFR1 expressing cells, especially known for the activation of endothelial cells [43] | 24 [17,27,42] 48 [9,14,15,42] 72 [42] |
| IFNλ/IL-29 | WD-hAECs, hAECs, A549 and BEAS-2B [6,13,44,45] | Stimulate innate antiviral mechanisms [46] | Thought to mostly act on epithelial cells in various organs [46] | 24 [45] 48 [13] 72 [13] 96 [6,13,44] |
| BAFF/TNFSF13B | WD-hAECs, hAECs [9,47] | Regulates B-cell homeostasis, promotes the induction of pathogen specific antibody production [48] | Interacts with B-cells via BAFF receptor (BAFF-R) or transmembrane activator and calcium-modulating cyclophilin ligand interactor (TACI) signalling [48] | 48 [9,47] |
| TSLP | hAECs [49,50] | Stimulates haematopoietic cells to induce Th2 responses; associated with blockage of Th1 and Th17 responses [51] | Interacts with Monocytes, DCs, CD4+ T-cells, B-cells and eosinophils via TSLP receptor (TSLP-R) [51] | 24 [49] |
| TGF-β | WD-hAECs, hAECs, A549 and BEAS-2B [52,53] | Pleiotropic cytokine; regulates peripheral tolerance; regulates T-cell homeostasis and survival; promotes Th17 cell differentiation; suppresses cells of the innate immune system [54] | Interacts with tissue cells and leukocytes that express the TGF-β receptors TGF-β I and II [54] | 24 [52] |
| CCL2/MCP-1 | WD-hAECs, hAECs A549, SAEs, BEAS-2B [9,13,16,17,55,56,57] | Mainly involved in monocyte trafficking [58] | Recruits monocytes via CCR2 and CCR4 signaling [58] | 6 [16] 24 [16,17,56] 48 [9,13,55] |
| CCL3/MIP-1α | WD-hAECs, hAECs A549, SAEs, BEAS-2B and Hep-2 [9,17,25,55,57,59,60] | Mainly promotes monocyte and NK cell trafficking [58] | Recruits monocytes, macrophages and NK cells via CCR1, CCR4 and CCR5 signaling [58] | 24 [17] 48 [9,55,59] 72 [60] 96 [60] 120 [60] |
| CCL4/MIP-1β | WD-hAECs, hAECs, A459 and SAEs [9,17,25,57] | Mainly promotes monocyte and NK cell trafficking [58] | Recruits monocytes, macrophages and NK cells via CCR1, CCR5, and CCR8 signaling [58] | 24 [17] 48 [9] |
| CCL5/RANTES | WD-hAECs, hAECs, A549 and SAEs, HEp-2 [6,7,9,13,16,17,21,24,25,29,30,36,45,55,56,57,59,60,61,62,63,64,65] | Mainly promotes monocyte and NK cell trafficking [58] | Recruits monocytes, NK cells, basophils, DCs and eosinophils via CCR1, CCR3, CCR4 and CCR5 signaling [58] | 12 [45,61] 24 [6,17,24,29,36,45,56,61,62,63] 48 [9,13,24,30,55,59,61,63,64] 72 [24,63] 96 [6,7,21,60] 120 [7] 144 [7] |
| CCL7/MCP-3 | WD-hAECs and SAEs [9,17] | Mainly involved in monocyte recruitment [58] | Recruits monocytes via CCR1, CCR2 and CCR3 signaling [58] | 24 [17] 48 [9] |
| CCL11/Eotaxin | WD-hAECs, hAECs [17,65] | Promotes eosinophil and basophil recruitment [58] | Recruits eosinophils, basophils via CCR3 signaling [58] | 24 [17] |
| CXCL8/IL-8 | WD-hAECs, A549, SAEs, BEAS-2B, Hep-2 [6,7,9,13,15,16,17,20,21,22,23,24,25,27,28,30,36,42,56,57,60,61,64,66,67] | Mainly involved in neutrophil trafficking [58] | Recruits neutrophils via CXCR1 and CXCR2 signaling [58] | 2 [16,28,66] 6 [16,28,64,66] 12 [16,23,61] 24 [15,16,17,20,24,27,28,36,42,58,61,66] 48 [9,13,15,22,23,24,30,42,61,64] 72 [24,42] 96 [6,7,20,21,22,60] 120 [7] 144 [7] |
| CXCL9/MIG | WD-hAECs, SAEs [9,17] | Promotes T-cell and NK cell trafficking; associated with Th1 responses [58] | Recruits T-cells and NK cells via CXCR3 signaling [58] | 24 [17] 48 [9] |
| CXCL10/IP-10 | WD-hAECs, A549, SAEs, BEAS-2B [6,7,9,13,16,17,21,68] | Promotes T-cell and NK cell trafficking; associated with Th1 responses [58] | Recruits T-cells and NK cells via CXCR3 signaling [58] | 18 [16] 24 [7,17,68] 48 [9,13,68] 72 [13,68] 96 [6,7,21] 120 [7] 144 [7] |
| CXCL11/I-TAC/IP-9 | WD-hAECs, A549 and SAEs [6,9,57] | Promotes T-cell and NK cell trafficking; associated with Th1 responses [58] | Recruits T-cells and NK cells via CXCR3 signaling [58] | 48 [9] 96 [6] |
| HMGB1 | hAECs and A549 [36,50] | Pleiotropic cytokine, initiates self-perpetuating inflammatory responses; known pyrogen [69] | Induces the secretion of cytokines and chemokines from tissue cells and lymphocytes via TLR signaling [69] | 24 [36] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaser, L.; Coulter, P.J.; Shields, M.; Touzelet, O.; Power, U.F.; Broadbent, L. Airway Epithelial Derived Cytokines and Chemokines and Their Role in the Immune Response to Respiratory Syncytial Virus Infection. Pathogens 2019, 8, 106. https://doi.org/10.3390/pathogens8030106
Glaser L, Coulter PJ, Shields M, Touzelet O, Power UF, Broadbent L. Airway Epithelial Derived Cytokines and Chemokines and Their Role in the Immune Response to Respiratory Syncytial Virus Infection. Pathogens. 2019; 8(3):106. https://doi.org/10.3390/pathogens8030106
Chicago/Turabian StyleGlaser, Lena, Patricia J. Coulter, Michael Shields, Olivier Touzelet, Ultan F. Power, and Lindsay Broadbent. 2019. "Airway Epithelial Derived Cytokines and Chemokines and Their Role in the Immune Response to Respiratory Syncytial Virus Infection" Pathogens 8, no. 3: 106. https://doi.org/10.3390/pathogens8030106
APA StyleGlaser, L., Coulter, P. J., Shields, M., Touzelet, O., Power, U. F., & Broadbent, L. (2019). Airway Epithelial Derived Cytokines and Chemokines and Their Role in the Immune Response to Respiratory Syncytial Virus Infection. Pathogens, 8(3), 106. https://doi.org/10.3390/pathogens8030106