
The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum

 ,
, 
 ,
, 
 ,
, 

Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stevanović, O.; Diakou, A.; Morelli, S.; Paraš, S.; Trbojević, I.; Nedić, D.; Sladojević, Ž.; Kasagić, D.; Di Cesare, A. Severe Verminous Pneumonia Caused by Natural Mixed Infection with Aelurostrongylus abstrusus and Angiostrongylus chabaudi in a European Wildcat from Western Balkan Area. Acta Parasitol. 2019, 64, 411–417. [Google Scholar] [CrossRef]
- Diakou, A.; Dimzas, D.; Astaras, C.; Savvas, I.; Di Cesare, A.; Morelli, S.; Neofitos, Κ.; Migli, D.; Traversa, D. Clinical investigations and treatment outcome in a European wildcat (Felis silvestris silvestris) infected by cardio-pulmonary nematodes. Vet. Parasitol. Reg. Stud. Rep. 2020, 19, 100357. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Morelli, S.; Di Cesare, A.; Diakou, A. Felid Cardiopulmonary Nematodes: Dilemmas Solved and New Questions Posed. Pathogens 2021, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, A.; Capelli, G.; Joachim, A.; Hinney, B.; Losson, B.; Kirkova, Z.; René-Martellet, M.; Papadopoulos, E.; Farkas, R.; Napoli, E.; et al. Lungworms and gastrointestinal parasites of domestic cats: A European perspective. Int. J. Parasitol. 2017, 47, 517–528. [Google Scholar] [CrossRef]
- Traversa, D.; Morelli, S.; Cassini, R.; Crisi, P.E.; Russi, I.; Grillotti, E.; Manzocchi, S.; Simonato, G.; Beraldo, P.; Viglietti, A.; et al. Occurrence of canine and feline extra-intestinal nematodes in key endemic regions of Italy. Acta Trop. 2019, 193, 227–235. [Google Scholar] [CrossRef]
- Morelli, S.; Diakou, A.; Di Cesare, A.; Schnyder, M.; Colombo, M.; Strube, C.; Latino, R.; Traversa, D. Feline lungworms in Greece: Copromicroscopic, molecular and serological study. Parasitol. Res. 2020, 119, 2877–2883. [Google Scholar] [CrossRef]
- Carruth, A.J.; Buch, J.S.; Braff, J.C.; Chandrashekar, R.; Bowman, D.D. Distribution of the feline lungworm Aelurostrongylus abstrusus in the USA based on fecal testing. J. Feline Med. Surg. Open Rep. 2019, 5, 2055116919869053. [Google Scholar] [CrossRef]
- da Silva Lima, W.; Ferreira Farago, E.C.; do Nascimento Mesquita, M.; Duarte Pacheco, A.; Fernandes Nunes da Silva Malavazi, P.; Salvador Oliveira, H.; Morelli, S.; Colombo, M.; Di Cesare, A.; Figueiredo de Souza, S. First Case of Clinical Cat Aelurostrongylosis in the Brazilian Amazon: Clinical and Molecular Insights. Pathogens 2021, 10, 595. [Google Scholar] [CrossRef]
- Di Cesare, A.; Laiacona, F.; Iorio, R.; Marangi, M.; Menegotto, A. Aelurostrongylus abstrusus in wild felids of South Africa. Parasitol. Res. 2016, 115, 3731–3735. [Google Scholar] [CrossRef]
- Morelli, S.; Diakou, A.; Colombo, M.; Di Cesare, A.; Barlaam, A.; Dimzas, D.; Traversa, D. Cat Respiratory Nematodes: Current Knowledge, Novel Data and Warranted Studies on Clinical Features, Treatment and Control. Pathogens 2021, 10, 454. [Google Scholar] [CrossRef] [PubMed]
- Gerichter, C.B. Studies on the nematodes parasitic in the lungs of Felidae in Palestine. Parasitology 1949, 39, 251–262. [Google Scholar] [CrossRef]
- López, C.; Panadero, R.; Paz, A.; Sánchez-Andrade, R.; Díaz, P.; Díez-Baños, P.; Morrondo, P. Larval development of Aelurostrongylus abstrusus (Nematoda, Angiostrongylidae) in experimentally infected Cernuella (Cernuella) virgata (Mollusca, Helicidae). Parasitol Res. 2005, 95, 13–16. [Google Scholar] [CrossRef]
- Giannelli, A.; Ramos, R.A.; Annoscia, G.; Di Cesare, A.; Colella, V.; Brianti, E.; Dantas-Torres, F.; Mutafchiev, Y.; Otranto, D. Development of the feline lungworms Aelurostrongylus abstrusus and Troglostrongylus brevior in Helix aspersa snails. Parasitology 2014, 14, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Di Cesare, A. Diagnosis and management of lungworm infections in cats: Cornerstones, dilemmas and new avenues. J. Feline Med. Surg. 2016, 18, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Veronesi, F.; Traversa, D. Felid Lungworms and Heartworms in Italy: More Questions than Answers? Trends Parasitol. 2015, 31, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Crisi, P.E.; Di Giulio, E.; Veronesi, F.; Frangipane di Regalbono, A.; Talone, T.; Traversa, D. Larval development of the feline lungworm Aelurostrongylus abstrusus in Helix aspersa. Parasitol. Res. 2013, 112, 3101–3108. [Google Scholar] [CrossRef]
- Cardillo, N.M.; Ercole, M.; Fariña, F.; Pasqualetti, M.; Loiza, Y.; Pérez, M.; Bonboni, A.; Ribicich, M. Larval development of Aelurostrongylus abstrusus in experimentally infected Rumina decollata snails. Vet. Parasitol. 2018, 251, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Zottler, E.; Schnyder, M. Larval development of the cat lungworm Aelurostrongylus abstrusus in the tropical freshwater snail Biomphalaria glabrata. Parasitol. Open 2016, 2, e8. [Google Scholar] [CrossRef] [Green Version]
- Jeżewski, W.; Buńkowska-Gawlik, K.; Hildebrand, J.; Perec-Matysiak, A.; Laskowski, Z. Intermediate and paratenic hosts in the life cycle of Aelurostrongylus abstrusus in natural environment. Vet. Parasitol. 2013, 198, 401–405. [Google Scholar] [CrossRef]
- Lange, M.K.; Penagos-Tabares, F.; Hirzmann, J.; Failing, K.; Schaper, R.; Van Bourgonie, Y.R.; Backeljau, T.; Hermosilla, C.; Taubert, A. Prevalence of Angiostrongylus vasorum, Aelurostrongylus abstrusus and Crenosoma vulpis larvae in native slug populations in Germany. Vet. Parasitol. 2018, 254, 120–130. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Lange, M.K.; Vélez, J.; Hirzmann, J.; Gutiérrez-Arboleda, J.; Taubert, A.; Hermosilla, C.; Chaparro Gutiérrez, J.J. The invasive giant African snail Lissachatina fulica as natural intermediate host of Aelurostrongylus abstrusus, Angiostrongylus vasorum, Troglostrongylus brevior, and Crenosoma vulpis in Colombia. PLoS Negl. Trop. Dis. 2019, 13, e0007277. [Google Scholar] [CrossRef]
- Penagos-Tabares, F.; Groß, K.M.; Hirzmann, J.; Hoos, C.; Lange, M.K.; Taubert, A.; Hermosilla, C. Occurrence of canine and feline lungworms in Arion vulgaris in a park of Vienna: First report of autochthonous Angiostrongylus vasorum, Aelurostrongylus abstrusus and Troglostrongylus brevior in Austria. Parasitol. Res. 2020, 119, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Fuehrer, H.P.; Morelli, S.; Bleicher, J.; Brauchart, T.; Edler, M.; Eisschiel, N.; Hering, T.; Lercher, S.; Mohab, K.; Reinelt, S.; et al. Detection of Crenosoma spp., Angiostrongylus vasorum and Aelurostrongylus abstrusus in Gastropods in Eastern Austria. Pathogens 2020, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Dimzas, D.; Morelli, S.; Traversa, D.; Di Cesare, A.; Van Bourgonie, Y.R.; Breugelmans, K.; Backeljau, T.; Frangipane di Regalbono, A.; Diakou, A.; Backeljau, T. Intermediate gastropod hosts of major feline cardiopulmonary nematodes in an area of wildcat and domestic cat sympatry in Greece. Parasites Vectors 2020, 13, 345. [Google Scholar] [CrossRef]
- Global Invasive Species Database. Available online: www.iucngisd.org (accessed on 14 April 2021).
- Guiller, A.; Martin, M.C.; Hiraux, C.; Madec, L. Tracing the invasion of the mediterranean land snail Cornu aspersum aspersum becoming an agricultural and garden pest in areas recently introduced. PLoS ONE 2012, 7, e49674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helm, J.; Roberts, L.; Jefferies, R.; Shaw, S.E.; Morgan, E.R. Epidemiological survey of Angiostrongylus vasorum in dogs and slugs around a new endemic focus in Scotland. Vet. Rec. 2015, 177, 46. [Google Scholar] [CrossRef]
- Schnyder, M.; Di Cesare, A.; Basso, W.; Guscetti, F.; Riond, B.; Glaus, T.; Crispi, P.; Deplazes, P. Clinical, laboratory and pathological findings in cats experimentally infected with Aelurostrongylus abstrusus. Parasitol. Res. 2014, 113, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Raue, K.; Rohdich, N.; Hauck, D.; Zschiesche, E.; Morelli, S.; Traversa, D.; Di Cesare, A.; Roepke, R.K.A.; Strube, C. Efficacy of Bravecto® Plus spot-on solution for cats (280 mg/ml fluralaner and 14 mg/ml moxidectin) for the prevention of aelurostrongylosis in experimentally infected cats. Parasites Vectors. 2021, 14, 110. [Google Scholar] [CrossRef]
- Gökpinar, S.; Yildiz, K. The effect of different temperatures on viability of Aelurostrongylus abstrusus first stage larvae in faeces of cats. Turk. Parazitol. Derg. 2010, 34, 102–105. [Google Scholar]
- Morelli, S.; Traversa, D.; Colombo, M.; Raue, K.; Strube, C.; Pollmeier, M.; Di Cesare, A. The effect of the hibernation on the larval development of Troglostrongylus brevior in the land snail Cornu aspersum. Vet. Parasitol. 2020, 282, 109123. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Arfuso, F.; Gaglio, G.; Abbate, J.M.; Giannetto, S.; Brianti, E. Effect of different temperatures on survival and development of Aelurostrongylus abstrusus (Railliet, 1898) larvae. J. Helminthol. 2020, 94, e113. [Google Scholar] [CrossRef]
- Mendonca, C.L.; Carvalho, O.S.; Mota, E.M.; Pelajo-Machado, M.; Caputo, L.F.; Lenzi, H.L. Penetration sites and migratory routes of Angiostrongylus costaricensis in the experimental intermediate host (Sarasinula marginata). Mem. Inst. Oswaldo Cruz 1999, 94, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Giannelli, A.; Colella, V.; Abramo, F.; do Nascimento Ramos, R.A.; Falsone, L.; Brianti, E.; Varcasia, A.; Dantas-Torres, F.; Knaus, M.; Fox, M.T.; et al. Release of lungworm larvae from snails in the environment: Potential for alternative transmission pathways. PLoS Negl. Trop. Dis. 2015, 9, e0003722. [Google Scholar] [CrossRef]
- Di Cesare, A.; Crisi, P.E.; Bartolini, R.; Iorio, R.; Talone, T.; Filippi, L.; Traversa, D. Larval development of Angiostrongylus vasorum in the land snail Helix aspersa. Parasitol. Res. 2015, 114, 3649–3655. [Google Scholar] [CrossRef]
- Colella, V.; Mutafchiev, Y.; Cavalera, M.A.; Giannelli, A.; Lia, R.P.; Dantas-Torres, F.; Otranto, D. Development of Crenosoma vulpis in the common garden snail Cornu aspersum: Implications for epidemiological studies. Parasites Vectors 2016, 9, 208. [Google Scholar] [CrossRef] [Green Version]
- Ansart, A.; Vernon, P.; Daguzan, J. Photoperiod is the main cue that triggers supercooling ability in the land snail, Helix aspersa (Gastropoda: Helicidae). Cryobiology 2001, 42, 266–273. [Google Scholar] [CrossRef]
- Ramos-Vasconcelos, G.R.; Hermes-Lima, M. Hypometabolism, antioxidant defenses and free radical metabolism in the pulmonate land snail Helix aspersa. J. Exp. Biol. 2003, 206, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Artacho, P.; Nespolo, R.F. Natural selection reduces energy metabolism in the garden snail, Helix aspersa (Cornu aspersum). Evolution 2009, 63, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, A.; Filser, J.; Briand, V.; Charrier, M. Seasonally contrasting life-history strategies in the land snail Cornu aspersum: Physiological and ecological implications. Can. J. Zool. 2010, 88, 995–1002. [Google Scholar] [CrossRef]
- Tunholi-Alves, V.M.; Tunholi, V.M.; Garcia, J.; Mota, E.M.; Castro, R.N.; Pontes, E.G.; Pinheiro, J. Unveiling the oxidative metabolism of Achatina fulica (Mollusca: Achatina fulica (Mollusca: Gastropoda) experimentally infected to Angiostrongylus cantonensis (Nematoda: Metastrongylidae). Parasitol. Res. 2018, 117, 1773–1781. [Google Scholar] [CrossRef]
- Rojas, M.; Li, L.Z.; Kanakidou, M.; Hatzianastassiou, N.; Seze, G.; Le Treut, H. Winter weather regimes over the Mediterranean region: Their role for the regional climate and projected changes in the twenty-first century. Clim. Dyn. 2013, 41, 551–571. [Google Scholar] [CrossRef]
- Diakou, A.; Di Cesare, A.; Barros, L.A.; Morelli, S.; Halos, L.; Beugnet, F.; Traversa, S. Occurrence of Aelurostrongylus abstrusus and Troglostrongylus brevior in domestic cats in Greece. Parasites Vectors 2015, 8, 590. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Di Francesco, G.; Frangipane di Regalbono, A.; Eleni, C.; De Liberato, C.; Marruchella, G.; Iorio, R.; Malatesta, D.; Romanucci, M.R.; Bongiovanni, L.; et al. Retrospective study on the occurrence of the feline lungworms Aelurostrongylus abstrusus and Troglostrongylus spp. in endemic areas of Italy. Vet. J. 2015, 203, 233–238. [Google Scholar] [CrossRef]
- Di Cesare, A.; Veronesi, F.; Grillotti, E.; Manzocchi, S.; Perrucci, S.; Beraldo, P.; Cazzin, S.; De Liberato, C.; Barros, L.A.; Simonato, G.; et al. Respiratory nematodes in cat populations of Italy. Parasitol. Res. 2015, 114, 4463–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdushy, T.; Kapel, C.M.; Webster, P.; Al-Sabi, M.N.; Grønvold, J.R. The effect of temperature and host age on the infectivity and development of Angiostrongylus vasorum in the slug Arion lusitanicus. Parasitol. Res. 2010, 107, 147–151. [Google Scholar] [CrossRef]
- Hodžić, A.; Alić, A.; Klebić, I.; Kadrić, M.; Brianti, E.; Duscher, G.G. Red fox (Vulpes vulpes) as a potential reservoir host of cardiorespiratory parasites in Bosnia and Herzegovina. Vet. Parasitol. 2016, 223, 63–70. [Google Scholar] [CrossRef]
- Grandi, G.; Comin, A.; Ibrahim, O.; Schaper, R.; Forshell, U.; Lind, E.O. Prevalence of helminth and coccidian parasites in Swedish outdoor cats and the first report of Aelurostrongylus abstrusus in Sweden: A coprological investigation. Acta Vet. Scand. 2017, 59, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueldner, E.K.; Gilli, U.; Strube, C.; Schnyder, M. Seroprevalence, biogeographic distribution and risk factors for Aelurostrongylus abstrusus infections in Swiss cats. Vet. Parasitol. 2019, 266, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.A.; Giannelli, A.; Dantas-Torres, F.; Brianti, E.; Otranto, D. Survival of first-stage larvae of the cat lungworm Troglostrongylus brevior (Strongylida: Crenosomatidae) under different conditions. Exp. Parasitol. 2013, 135, 570–572. [Google Scholar] [CrossRef]
- Traversa, D.; Faculty of Veterinary Medicine, University of Teramo, Piano d’Accio, Teramo, Italy. Personal communication, 2021.
- Dimzas, D.; Diakou, A.; Di Cesare, A.; Van Bourgornie, Y.R.; Backeliau, T.; Staikou, A.; Traversa, D. Gastropods as intermediate hosts of feline cardio-pulmonary parasites in Greece: Preliminary results. In Proceedings of the 14th International Congress on the Zoogeography and Ecology of Greece and Adjacent Regions, Thessaloniki, Greece, 27–30 June 2019. [Google Scholar]
- Di Cesare, A.; Frangipane di Regabono, A.; Tessarin, C.; Seghetti, M.; Iorio, R.; Simonato, G.; Traversa, D. Mixed infection by Aelurostrongylus abstrusus and Troglostrongylus brevior in kittens from the same litter in Italy. Parasitol. Res. 2014, 113, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Proćków, M.; Konowalik, K.; Proćków, J. Contrasting effects of climate change on the European and global potential distributions of two Mediterranean helicoid terrestrial gastropods. Reg. Environ. Chang. 2019, 19, 2637–2650. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Morelli, S.; Colombo, M.; Simonato, G.; Veronesi, F.; Marcer, F.; Diakou, A.; D’Angelosante, R.; Pantchev, N.; Psaralexi, E.; et al. Is Angiostrongylosis a Realistic Threat for Domestic Cats? Front. Vet. Sci. 2020, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Veronesi, F.; Frangipane di Regalbono, A.; Iorio, R.; Traversa, D. Novel Molecular Assay for Simultaneous Identification of Neglected Lungworms and Heartworms Affecting Cats. J. Clin. Microbiol. 2015, 53, 3009–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Euzeby, J. Helminthes para-sites de l’appareil respiratoire. In Diagnostic Expérimental des Helminthoses Animales; Informations Techniques des Services Vétérinaires”; Minitère de l’Agriculture: Paris, France, 1981; Volume 2. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
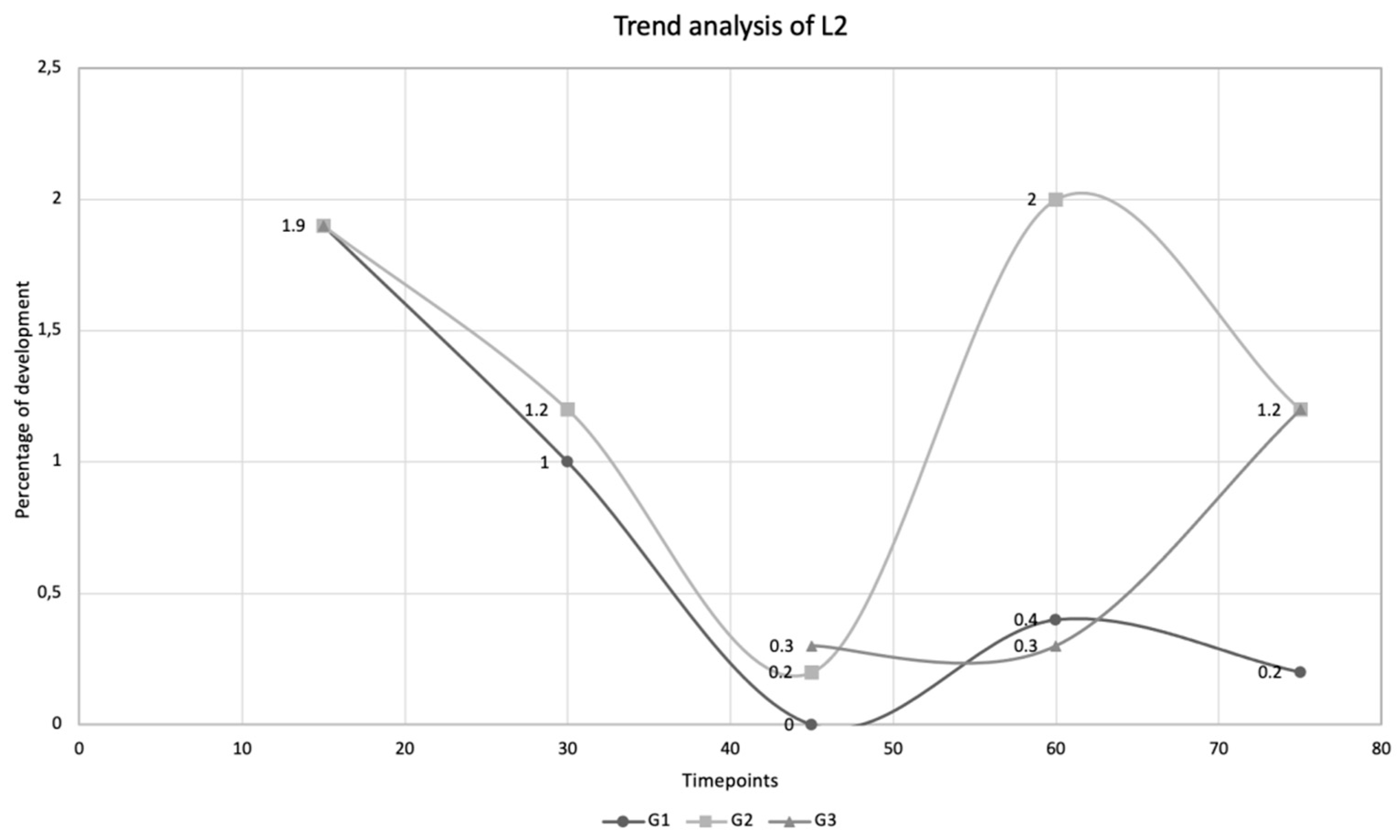
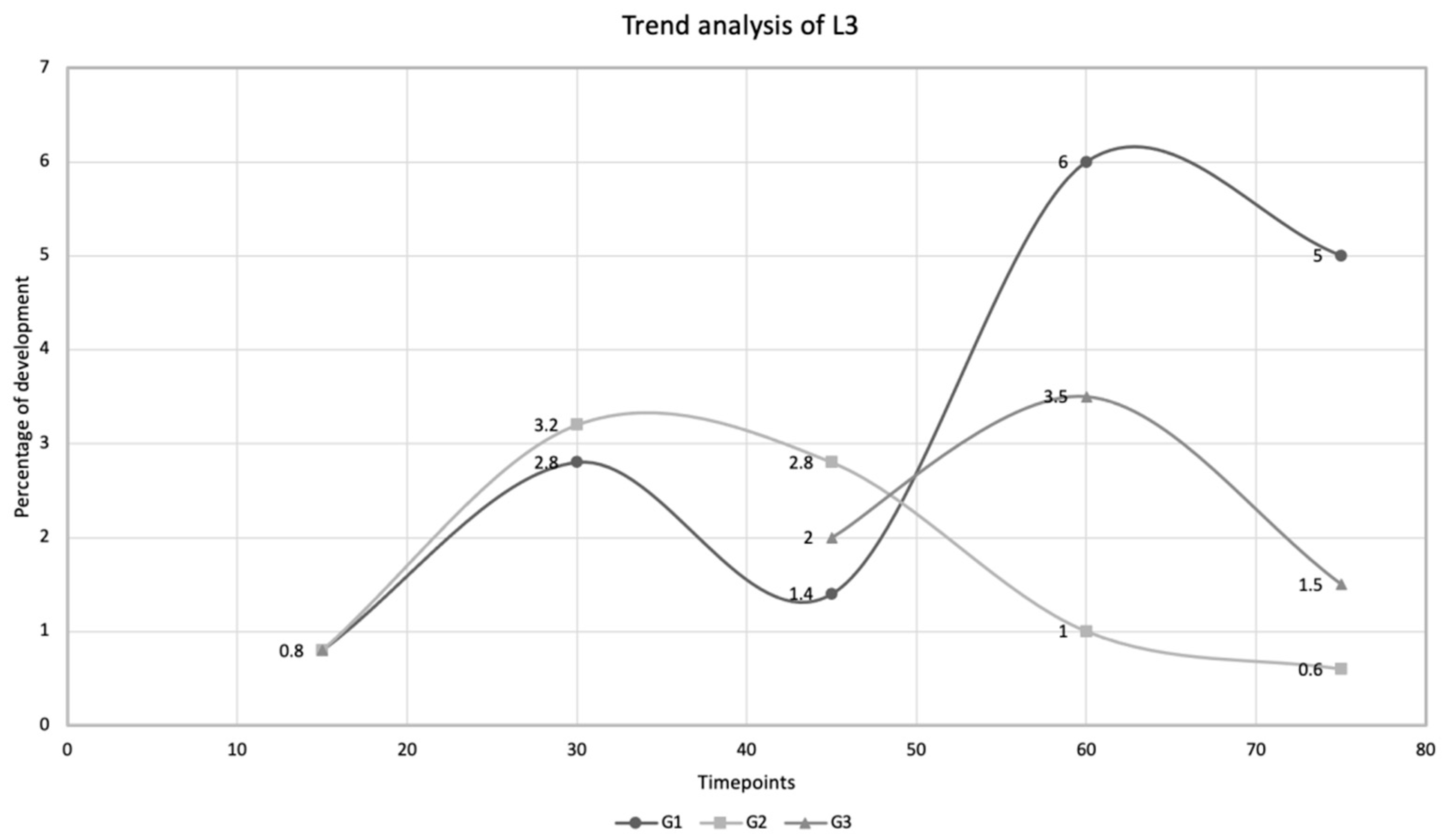
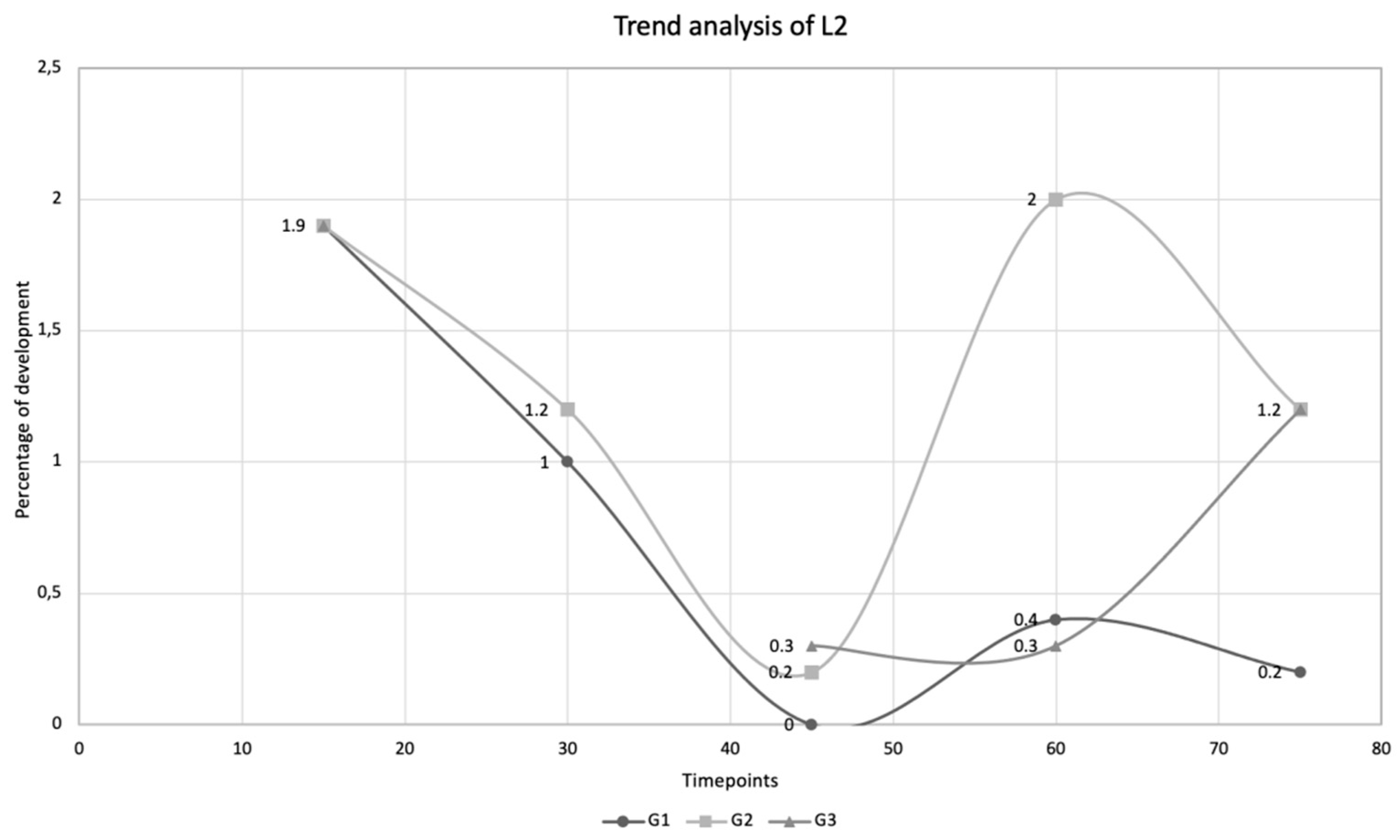
| Time Points | Larval Stages | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| L1 (%) | L2 (%) | L3 (%) | |||||||
| D15 | 2.8 | 1.9 | 0.8 | ||||||
| G1 | G2 | G3 | G1 | G2 | G3 | G1 | G2 | G3 | |
| D30 | 0.6 | 0.2 | 0 | 1.0 | 1.2 | 0 | 2.8 | 3.2 | 0 |
| D45 | 0 | 0 | 0 | 0 | 0.2 | 0.3 | 1.4 | 2.8 | 2.0 |
| D60 | 0 | 0 | 0 | 0.4 | 2.0 | 0.3 | 6.0 | 1.0 | 3.5 |
| D75 | 0 | 0 | 0 | 0.2 | 1.2 | 1.2 | 5.0 | 0.6 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morelli, S.; Colombo, M.; Diakou, A.; Traversa, D.; Grillini, M.; Frangipane di Regalbono, A.; Di Cesare, A. The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum. Pathogens 2021, 10, 960. https://doi.org/10.3390/pathogens10080960
Morelli S, Colombo M, Diakou A, Traversa D, Grillini M, Frangipane di Regalbono A, Di Cesare A. The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum. Pathogens. 2021; 10(8):960. https://doi.org/10.3390/pathogens10080960
Chicago/Turabian StyleMorelli, Simone, Mariasole Colombo, Anastasia Diakou, Donato Traversa, Marika Grillini, Antonio Frangipane di Regalbono, and Angela Di Cesare. 2021. "The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum" Pathogens 10, no. 8: 960. https://doi.org/10.3390/pathogens10080960
APA StyleMorelli, S., Colombo, M., Diakou, A., Traversa, D., Grillini, M., Frangipane di Regalbono, A., & Di Cesare, A. (2021). The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum. Pathogens, 10(8), 960. https://doi.org/10.3390/pathogens10080960