
Development and Validation of a Loop-Mediated Isothermal Amplification Diagnostic Method to Detect the Quarantine Potato Pale Cyst Nematode, Globodera pallida




Abstract
1. Introduction
2. Results
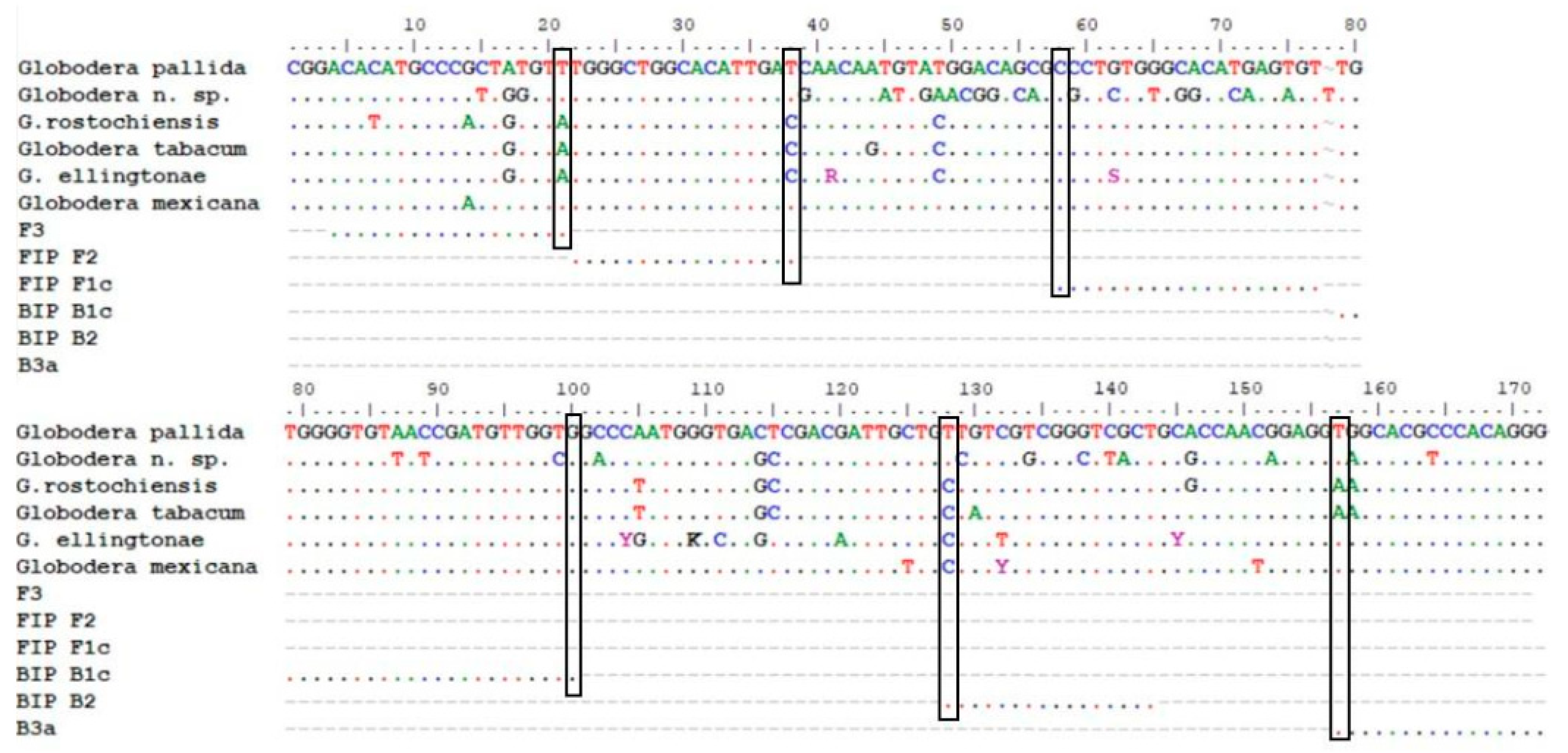
2.1. Sequencing and Primer Design
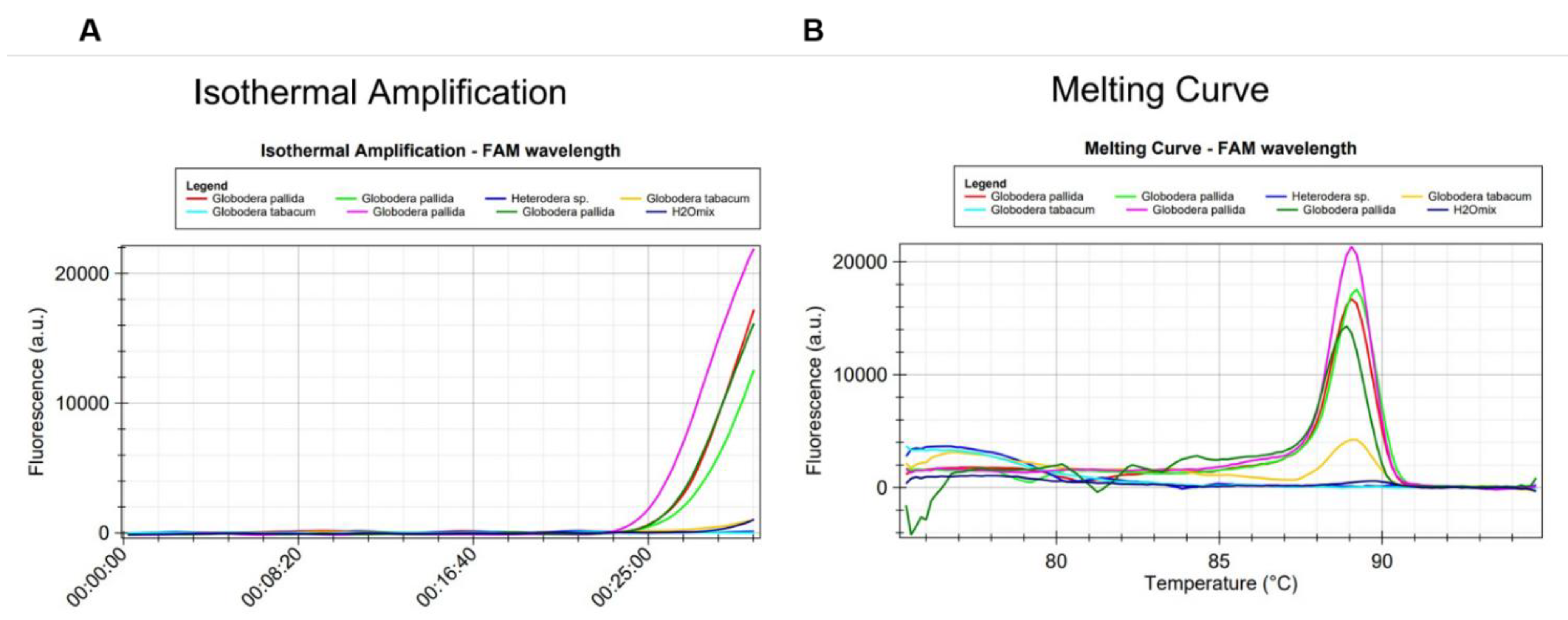
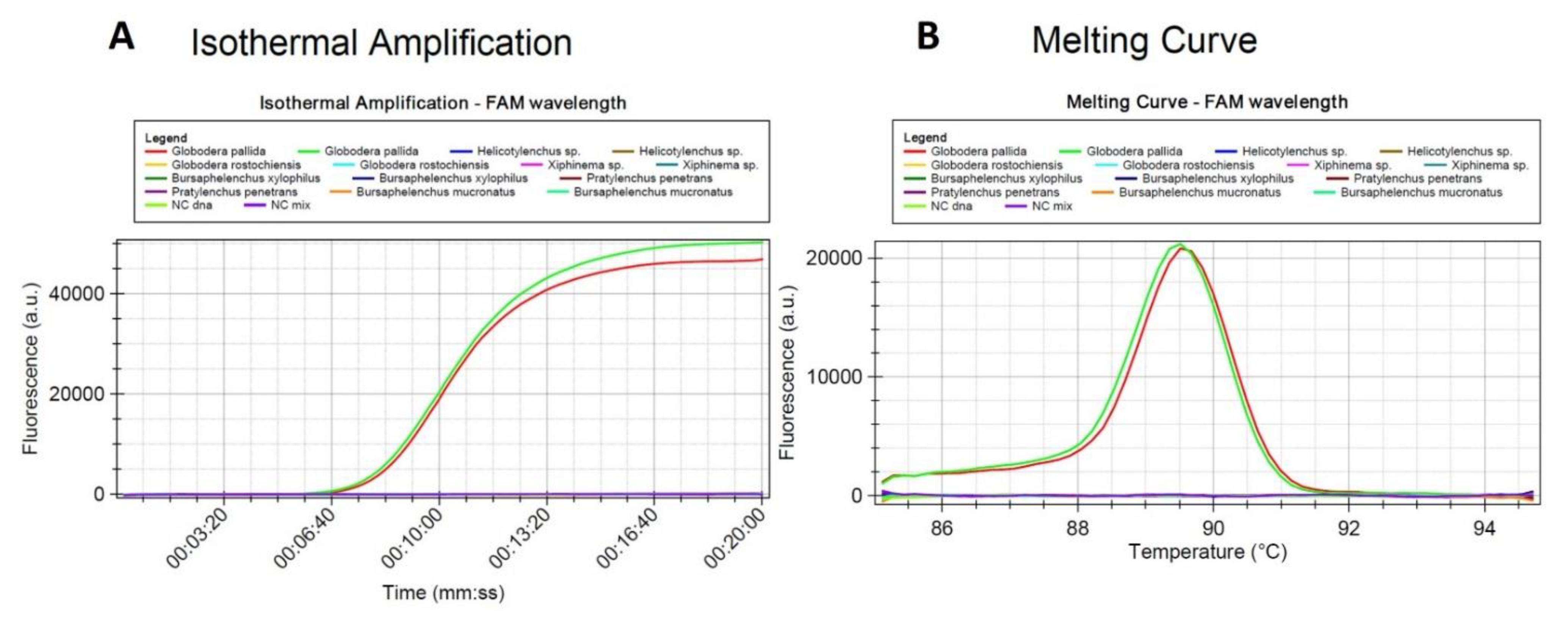
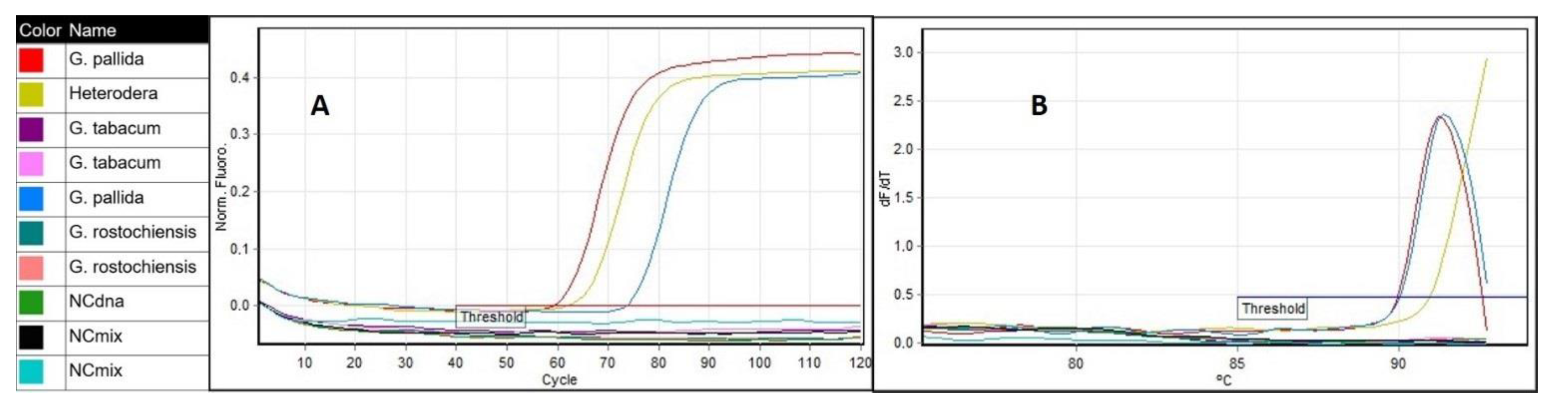
2.2. Optimization of the LAMP Assay Protocol and Specificity
2.3. LAMP Sensitivity Assay for Globodera pallida
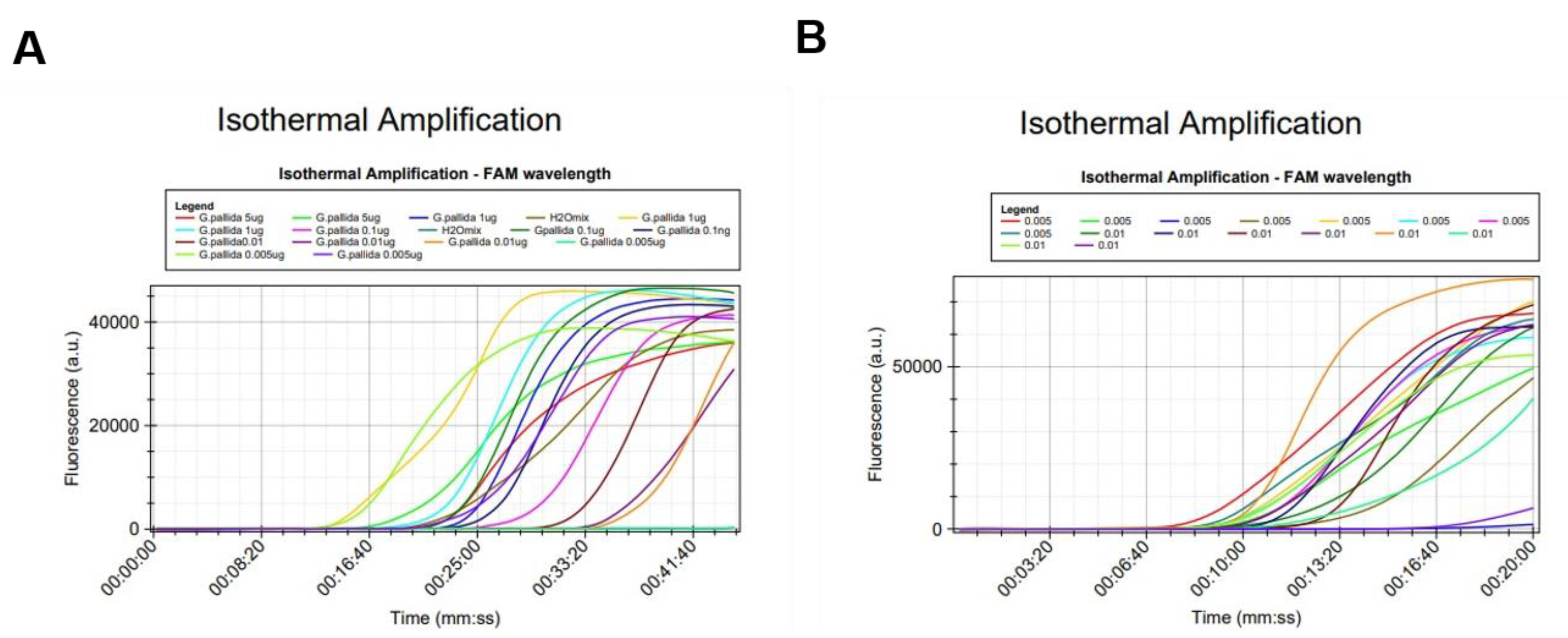
2.3.1. Analytical Sensitivity
2.3.2. Diagnostic Sensitivity
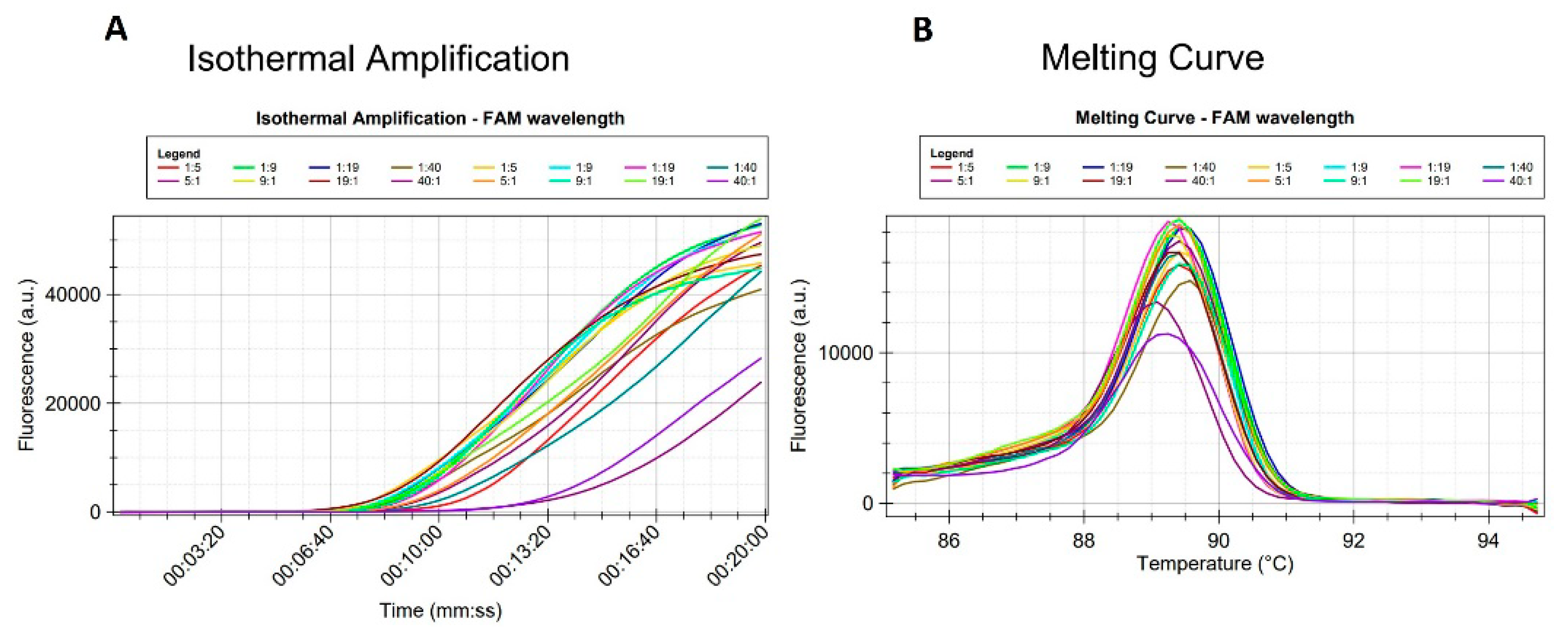
2.3.3. LAMP Reproducibility
3. Discussion
4. Materials and Methods
4.1. Samples, Chemicals and Standard Techniques
4.2. Globodera sp. Sequences and Primer Design
4.3. LAMP Assay
4.4. LAMP Specificity
4.5. LAMP Sensitivity
4.6. LAMP Reproducibility
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wollenweber, H.W. Krankheiten und Beschsädigungen der Kartoffel; Verlagsbuchhandlung Paul Parey: Berlin, Germany, 1923. [Google Scholar]
- Skarbilovich, T.S. On the structure and systematics of the nematode order Tylenchida Thorne, 1949. Acta Parasitol. Polonica 1959, 7, 117–132. [Google Scholar]
- Stone, A. Heterodera Pallida N. Sp. (Nematoda: Heteroderidae), a Second Species of Potato Cyst Nematode 1. Nematologica 1972, 18, 591–606. [Google Scholar] [CrossRef]
- Grenier, E.; Fournet, S.; Petit, E.; Anthoine, G. A cyst nematode ’species factory’ called the Andes. Nematoogy 2010, 12, 163–169. [Google Scholar] [CrossRef]
- EPPO Global Database. Globodera pallida. Available online: https://gd.eppo.int/taxon/HETDRO/distribution (accessed on 30 October 2020).
- Lavrova, V.V.; Matveeva, E.M.; Zinovieva, S.V. Expression of genes, encoded defense proteins, in potato plants infected with the cyst-forming nematode Globodera rostochiensis (Wollenweber 1923) Behrens, 1975 and modulation of their activity during short-term exposure to low temperatures. Biol. Bull. 2017, 44, 128–136. [Google Scholar] [CrossRef]
- EPPO. Globodera rostochiensis and Globodera pallida PM 7/40 (4). EPPO Bull. 2017, 47, 174–197. [Google Scholar] [CrossRef]
- Camacho, M.J.; Nóbrega, F.; Lima, A.; Mota, M.; Inácio, M.L. Morphological and molecular identification of the potato cyst nematodes Globodera rostochiensis and G. pallida in Portuguese potato fields. Nematology 2017, 19, 883–889. [Google Scholar] [CrossRef]
- Bulman, S.R.; Marshall, J.W. Differentiation of Australasian potato cyst nematode (PCN) populations using the polymerase chain reaction (PCR). N. Z. J. Crop. Hortic. Sci. 1997, 25, 123–129. [Google Scholar] [CrossRef]
- Kogovšek, P.; Hodgetts, J.; Hall, J.; Prezelj, N.; Nikolic, P.; Mehle, N.; Lenarčič, R.; Rotter, A.; Dickinson, M.; Boonham, N.; et al. LAMP assay and rapid sample preparation method for on-site detection of flavescence dorée phytoplasma in grapevine. Plant Pathol. 2015, 64, 286–296. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchai, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Subbotin, S.; Waeyenberge, L.; Moens, M. Molecular Systematics. In Plant Nematology; Perry, R., Moens, M., Eds.; CAB International: Wallingford, UK, 2013. [Google Scholar]
- Freitas, P.P.; Martins, V.C.; Cardoso, F.A.; Fernandes, E.; Sobrino, T.; Castillo, J.; Chicharo, A.; Abal, M.; Lopez-Lopez, R.; Dias, T.S.; et al. Spintronic biochips: From the laboratory to pre-clinical applications. In Nanomagnetism: Applications and Perspectives; Fermon, C., Van De Worde, M., Eds.; John Wiley and Sons: New York, NY, USA, 2016. [Google Scholar]
- Martins, V.; Cardoso, F.; Germano, J.; Cardoso, S.; Sousa, L.; Piedade, M.; Freitas, P.; Fonseca, L.P. Femtomolar limit of detection with a magnetoresistive biochip. Biosens. Bioelectron. 2009, 24, 2690–2695. [Google Scholar] [CrossRef]
- Dias, T.M.; Cardoso, F.A.; Martins, S.A.M.; Martins, V.C.; Cardoso, S.; Gaspar, J.F.; Monteiro, G.; De Freitas, S.C. Implementing a strategy for on-chip detection of cell-free DNA fragments using GMR sensors: A translational application in cancer diagnostics using ALU elements. Anal. Methods 2015, 8, 119–128. [Google Scholar] [CrossRef]
- Sarkes, A.; Fu, H.; Feindel, D.; Harding, M.; Feng, J. Development and evaluation of a loop-mediated isothermal amplification (LAMP) assay for the detection of Tomato brown rugose fruit virus (ToBRFV). PLoS ONE 2020, 15, e0230403. [Google Scholar] [CrossRef]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, A.S. Loop Mediated Isothermal Amplification: Principles and Applications in Plant Virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M. Loop-Mediated Isothermal Amplification (LAMP) for Detection of Phytoplasmas in the Field. In Plant Pathology; Humana Press: New York, NY, USA, 2015; pp. 99–111. [Google Scholar] [CrossRef]
- Blaser, S.; Diem, H.; Von Felten, A.; Gueuning, M.; Andreou, M.; Boonham, N.; Tomlinson, J.; Müller, P.; Utzinger, J.; Frey, B.; et al. A Loop-mediated Isothermal Amplification (LAMP) Assay for Rapid Identification of Bemisia tabaci. J. Vis. Exp. 2018, 140, e58502. [Google Scholar] [CrossRef]
- Duan, Y.; Ge, C.; Zhang, X.; Wang, J.; Zhou, M. A rapid detection method for the plant pathogen Sclerotinia sclerotiorum based on loop-mediated isothermal amplification (LAMP). Australas. Plant Pathol. 2013, 43, 61–66. [Google Scholar] [CrossRef]
- Tian, Q.; Lu, C.; Wang, S.; Xiong, Q.; Zhang, H.; Wang, Y.; Zheng, X. Rapid diagnosis of soybean anthracnose caused by Colletotrichum truncatum using a loop-mediated isothermal amplification (LAMP) assay. Eur. J. Plant Pathol. 2016, 148, 785–793. [Google Scholar] [CrossRef]
- Khan, M.; Wang, R.; Li, B.; Liu, P.; Weng, Q.; Chen, Q. Comparative Evaluation of the LAMP Assay and PCR-Based Assays for the Rapid Detection of Alternaria solani. Front. Microbiol. 2018, 9, 2089. [Google Scholar] [CrossRef] [PubMed]
- Aglietti, C.; Luchi, N.; Pepori, A.L.; Bartolini, P.; Pecori, F.; Raio, A.; Capretti, P.; Santini, A. Real-time loop-mediated isothermal amplification: An early-warning tool for quarantine plant pathogen detection. AMB Express 2019, 9, 50. [Google Scholar] [CrossRef]
- King, K.M.; Hawkins, N.J.; Atkins, S.; Dyer, P.S.; West, J.S.; Fraaije, B.A. First application of loop-mediated isothermal amplification (LAMP) assays for rapid identification of mating type in the heterothallic fungus Aspergillus fumigatus. Mycoses 2019, 62, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, S.Y.; Zhang, C.-Q. Establishment of a Rapid Detection Method for Rice Blast Fungus Based on One-Step Loop-Mediated Isothermal Amplification (LAMP). Plant Dis. 2019, 103, 1967–1973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Dai, D.J.; Wang, H.D.; Zhang, C.Q. One-step loop-mediated isothermal amplification (LAMP) for the rapid and sensitive detection of Fusarium fujikuroi in bakanae disease through NRPS31, an important gene in the gibberellic acid bio-synthesis. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Chen, Z.-D.; Kang, H.-J.; Chai, A.-L.; Shi, Y.-X.; Xie, X.-W.; Li, L.; Li, B.-J. Development of a loop-mediated isothermal amplification (LAMP) assay for rapid detection of Pseudomonas syringae pv. tomato in planta. Eur. J. Plant Pathol. 2020, 156, 739–750. [Google Scholar] [CrossRef]
- Ahuja, A.; Somvanshi, V.S. Diagnosis of plant-parasitic nematodes using loop-mediated isothermal amplification (LAMP): A review. Crop. Prot. 2020, 105459, 105459. [Google Scholar] [CrossRef]
- Kikuchi, T.; Aikawa, T.; Oeda, Y.; Karim, N.; Kanzaki, N. A Rapid and Precise Diagnostic Method for Detecting the Pinewood Nematode Bursaphelenchus xylophilus by Loop-Mediated Isothermal Amplification. Phytopathology 2009, 99, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Kanetani, S.; Kikuchi, T.; Akiba, M.; Nakamura, K.; Ikegame, H.; Tetsuka, K. Detection of Bursaphelenchus xylophilus from old discs of dead Pinus armandii var. amamiana trees using a new detection kit. For. Pathol. 2010, 41, 387–391. [Google Scholar] [CrossRef]
- Kang, J.S.; Kim, A.-Y.; Han, H.R.; Moon, Y.S.; Koh, Y.H. Development of two alternative Loop-mediated isothermal amplification tools for detecting pathogenic pine wood nematodes. For. Pathol. 2014, 45, 127–133. [Google Scholar] [CrossRef]
- Leal, I.; A Allen, E.; Foord, B.; Anema, J.; Reisle, C.; Uzunović, A.; Varga, A.; James, D. Detection of living Bursaphelenchus xylophilusin wood, using reverse transcriptase loop-mediated isothermal amplification (RT-LAMP). For. Pathol. 2015, 45, 134–148. [Google Scholar] [CrossRef]
- Meng, F.; Wang, X.; Wang, L.; Gou, D.; Liu, H.; Wang, Y.; Piao, C. A loop-mediated isothermal amplification-based method for detecting Bursaphelenchus xylophilus from Monochamus alternatus. For. Pathol. 2018, 48, e12404. [Google Scholar] [CrossRef]
- Nakajima, G.; Iki, T.; Yamanobe, T.; Nakamura, K.; Aikawa, T. Spatial and temporal distribution of Bursaphelenchus xylophilus inoculated in grafts of a resistant clone of Pinus thunbergii. J. For. Res. 2019, 24, 93–99. [Google Scholar] [CrossRef]
- Ide, T.; Kanzaki, N.; Giraldo, P.P.P.; Giblin-Davis, R.M. Loop-mediated isothermal amplification (LAMP) for detection of the red ring nematode, Bursaphelenchus cocophilus. Nematology 2017, 19, 559–565. [Google Scholar] [CrossRef]
- Niu, J.-H.; Guo, Q.-X.; Jian, H.; Chen, C.-L.; Yang, D.; Liu, Q.; Guo, Y.-D. Rapid detection of Meloidogyne spp. by LAMP assay in soil and roots. Crop. Prot. 2011, 30, 1063–1069. [Google Scholar] [CrossRef]
- Niu, J.H.; Jian, H.; Guo, Q.X.; Chen, C.L.; Wang, X.Y.; Liu, Q.; Guo, Y.D. Evaluation of loop-mediated isothermal amplification (LAMP) assays based on 5S rDNA-IGS2 regions for detecting Meloidogyne enterolobii. Plant Pathol. 2012, 61, 809–819. [Google Scholar] [CrossRef]
- Peng, H.; Long, H.; Huang, W.; Liu, J.; Cui, J.; Kong, L.; Hu, X.; Gu, J.; Peng, D. Rapid, simple and direct detection of Meloidogyne hapla from infected root galls using loop-mediated isothermal amplification combined with FTA technology. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Zhou, Q.-J.; Cai, Y.; Gu, J.-F.; Wang, X.; Chen, J. Rapid and sensitive detection of Meloidogyne mali by loop-mediated isothermal amplification combined with a lateral flow dipstick. Eur. J. Plant Pathol. 2016, 148, 755–769. [Google Scholar] [CrossRef]
- Zhang, L.; Gleason, C. Loop-Mediated Isothermal Amplification for the Diagnostic Detection of Meloidogyne chitwoodi and M. fallax. Plant Dis. 2019, 103, 12–18. [Google Scholar] [CrossRef]
- Waliullah, S.; Bell, J.; Jagdale, G.; Stackhouse, T.; Hajihassani, A.; Brenneman, T.; Ali, E. Rapid detection of pecan root-knot nematode, Meloidogyne partityla, in laboratory and field conditions using loop-mediated isothermal amplification. PLoS ONE 2020, 15, e0228123. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-I.; Yu, G.-Y. A loop-mediated isothermal amplification assay for the plant-parasitic nematode Aphelenchoides besseyi in rice seedlings. J. Nematol. 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Wang, D.-W.; Xu, C.-L.; Bai, Z.-S.; Li, J.-Y.; Han, Y.-C.; Zhao, L.-R.; Xie, H. Development of a loop-mediated isothermal amplification for rapid diagnosis of Aphelenchoides ritzemabosi. Eur. J. Plant Pathol. 2019, 155, 173–179. [Google Scholar] [CrossRef]
- Yu, L.-Z.; Song, S.-Y.; Yu, C.; Jiao, B.-B.; Tian, Y.-M.; Li, Y.-J. Rapid detection of Anguina agrostis by loop-mediated isothermal amplification. Eur. J. Plant Pathol. 2020, 156, 819–825. [Google Scholar] [CrossRef]
- Yu, L.-Z.; Song, S.-Y.; Yu, C.; Qi, L.-J.; Yu, Z.-X.; Jiao, B.-B.; Yang, J. A loop mediated isothermal amplification (LAMP) assay for rapid and reliable detection of Anguina wevelli, a grass parasitic nematode. Eur. J. Plant Pathol. 2018, 150, 725–734. [Google Scholar] [CrossRef]
- Peng, H.; Peng, D.; Hu, X.; He, X.; Wang, Q.; Huang, W.; He, W. Loop-mediated isothermal amplification for rapid and precise detection of the burrowing nematode, Radopholus similis, directly from diseased plant tissues. Nematology 2012, 14, 977–986. [Google Scholar] [CrossRef]
- Deng, M.-H.; Zhong, L.-Y.; Kamolnetr, O.; Limpanont, Y.; Lv, Z.-Y. Detection of helminths by loop-mediated isothermal amplification assay: A review of updated technology and future outlook. Infect. Dis. Poverty 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Wang, H.; Zhuo, K.; Liao, J. Loop-Mediated Isothermal Amplification for the Detection of Tylenchulus semipenetrans in Soil. Plant Dis. 2016, 100, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.-Q.; Cheng, J.-E.; Cheng, F.-X.; Zhang, D.-Y.; Liu, Y. Development and Evaluation of Loop-Mediated Isothermal Amplification Assay for Rapid Detection of Tylenchulus semipenetrans Using DNA Extracted from Soil. Plant Pathol. J. 2017, 33, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Shiqi, P.H. Globodera Rostochiensis SCAR Mark as well as LAMP Fast Detection Method and Application of Method. CN 104059909A, 24 September 2014. [Google Scholar]
- Peng, D.; Shiqi, P.H. Globodera Rostochiensis SCAR Mark and LAMP Method for Quick and Application. CN 104059909B, 4 May 2016. [Google Scholar]
- Camacho, M.J.; de Andrade, E.; Mota, M.; Nobrega, F.; Vicente, C.; Rusinque, L.; Inácio, M.L. Potato Cyst Nematodes: Geographical Distribution, Phylogenetic Relationships and Integrated Pest Management Outcomes in Portugal. Front. Plant Sci. 2020, 11, 606178. [Google Scholar] [CrossRef]
- Grenier, E.; Blok, V.C.; Jones, J.T.; Fouville, D.; Mugniéry, D. Identification of gene expression differences between Globodera pallida and G. ‘mexicana’ by suppression subtractive hybridization. Mol. Plant Pathol. 2002, 3, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Sabeh, M.; Lord, E.; Grenier, É.; St-Arnaud, M.; Mimee, B. What determines host specificity in hyperspecialized plant parasitic nematodes? BMC Genom. 2019, 20, 457. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A.; Franco, J.; Knoetze, R.; Roubtsova, T.V.; Bostock, R.M.; Vera, I.C.D.P. DNA barcoding, phylogeny and phylogeography of the cyst nematode species from the genus Globodera (Tylenchida: Heteroderidae). Nematology 2020, 22, 269–297. [Google Scholar] [CrossRef]
- Djebroune, A.; Chakali, G.; de Andrade, E.; Camacho, M.J.; Rusinque, L.; Inácio, M.L. Integrative Morphometric and Molecular Approach to Update the Impact and Distribution of Potato Cyst Nematodes Globodera rostochiensis and Globodera pallida (Tylenchida: Heteroderidae) in Algeria. Pathogens 2021, 10, 216. [Google Scholar] [CrossRef]
- Huang, B.; Montgomery, B.L.; Adamczyk, R.; Ehlers, G.; Hurk, A.F.V.D.; Warrilow, D. A LAMP-based colorimetric assay to expedite field surveillance of the invasive mosquito species Aedes aegypti and Aedes albopictus. PLoS Negl. Trop. Dis. 2020, 14, e0008130. [Google Scholar] [CrossRef] [PubMed]
- Martinez, B.; Reaser, J.K.; Dehgan, A.; Zamft, B.; Baisch, D.; McCormick, C.; Giordano, A.J.; Aicher, R.; Selbe, S. Technology innovation: Advancing capacities for the early detection of and rapid response to invasive species. Biol. Invasions 2019, 22, 75–100. [Google Scholar] [CrossRef]
- Tomlinson, J. Potential of LAMP for detection of plant pathogens. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2008, 3, 1–7. [Google Scholar] [CrossRef]
- Reis, L.G. Some morphological characters of a remarkable Globodera species (Nematoda: Heteroderidae) occurring in Portugal. Acta Parasitol. Port. 1997, 4, 126–127. [Google Scholar]
- BioEdit. Biological Sequence Alignment Editor for Win95/98/NT/2K/XP. Available online: http://www.mbio.ncsu.edu/BioEdit/BioEdit.html (accessed on 23 February 2013).
- Higgins, D.; Thompson, J.D.; Gibson, T.J. [22] Using CLUSTAL for multiple sequence alignments. Methods Enzym. 1996, 266, 383–402. [Google Scholar] [CrossRef]
- EPPO. PM 7/98 (4) Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity. EPPO Bull. 2019, 49, 530–563. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Set 1 |
| FIP (F1c + F2) | CAC GGC CAC GGA CGT AGC ACA TGT CGT ACG TGC CGT ACC C |
| BIP (B1c + B2) | GAG ACG ACG TGT TAG GAC CCA CTC ATC AAG TCT TAA ACC G |
| F3 | CAT GGA GTG TAG GCT GCT AT |
| B3 | TTA TAA AAA TGA GAA AAA G |
| Primers | Set 2a |
| FIP (F1c + F2) | ACA CTC ATG TGC CCA CAG GGT GGG CTG GCA CAT TGA T |
| BIP (B1c + B2) | TGG GGT GTA ACC GAT GTT GGT GAG CGA CCC GAC GAC AA |
| F3 | ACA CAT GCC CGC TAT GTT |
| B3(a) | CCC TGT GGG CGT GCC A |
| Species | Isolate | Origin | ng/µL | |
|---|---|---|---|---|
| I | G. pallida | MK791521 | Portugal | 5.2 |
| G. pallida | NPPO-NL Pa3 HLB | Netherlands | 1.4 | |
| G. rostochiensis | MK791264 | Portugal | 28.2 | |
| G. rostochiensis | NPPO-NL Ro1 HLB | Netherlands | 2.9 | |
| G. tabacum | NPPO-NL C6876 | Netherlands | 39.4 | |
| Globodera n. sp. | MT256387 | Portugal | 11.6 | |
| Heterodera sp. | SV-18-10003 * | Portugal | 18.1 | |
| G. rostochiensis | 058 | Samples from a interlaboratory test (European origin) | 16.8 | |
| II | G. pallida | 094 | Samples from a interlaboratory test (European origin) | 1.5 |
| G. pallida | 138 | 13.1 | ||
| G. tabacum | 185 | 3.1 | ||
| Heterodera sp. | 414 | 2.3 | ||
| G. tabacum | 447 | 1.4 | ||
| G. rostochiensis | 471 | 3.1 | ||
| G. pallida | 546 | 2.2 | ||
| G. pallida | 580 | 2.5 | ||
| G. rostochiensis | 629 | 3.0 |
| Protocol | Master Mix | Primer Volume (µL) | Amplification Temp. (°C), Time (s) | TMelting Heat-Cooling (°C) | |
|---|---|---|---|---|---|
| F3, B3 (Initial Conc. 50 µM) | FIP, BIP (Initial Conc. 50 µM) | ||||
| A | ISO-004 ([Mg2+] = 5 mM) | 0.10 | 0.80 | 65 °C, 60 min | 95 °C–75 °C |
| B | 0.60 | 65 °C, 20 min | |||
| C | 0.15 | 0.80 | |||
| D | 0.40 | ||||
| E | 0.60 | ||||
| F | 0.80 | 64 °C, 20 min | |||
| G | 0.12 | 0.70 | 66 °C, 20 min | ||
| H | 0.15 | 0.90 | 64 °C, 20 min | ||
| I | 0.12 | ||||
| J | ISO-001 ([Mg2+] = 3 mM) | 0.15 | 0.80 | 64 °C, 30 min | |
| K | 64 °C, 60 min | ||||
| L | 95 °C–85 °C | ||||
| M | ISO-004 | 64 °C, 20 min | |||
| Component | Initial Concentration | Vol/Reaction (μL) |
|---|---|---|
| ISO-004 (or 001) master mix | - | 15 |
| Primers FIP and BIP | 50 µM | 0.80 |
| F3 and B3a | 50 µM | 0.15 |
| Molecular grade water | - | 3.1 |
| DNA template | ≥5 pg | 5 |
| Samples | Samples Ratio (J2 G. rostochiensis: J2 G. pallida) | ng/µL |
|---|---|---|
| 1 | 1:5 | 3.2 |
| 2 | 1:9 | 1.7 |
| 3 | 1:19 | 2.3 |
| 4 | 1:40 | 2.6 |
| 5 | 5:1 | 2.4 |
| 6 | 9:1 | 2.0 |
| 7 | 19:1 | 4.3 |
| 8 | 40:1 | 2.0 |
| Species | Isolate | Origin |
|---|---|---|
| Pratylenchus penetrans | A44L4 * | Portugal |
| Xiphinema sp. | SV-21-00826 * | Portugal |
| Helicotylenchus sp. | SV-20-0967-01 * | Portugal |
| Bursaphelencus xylophilus | SV-21-0502-02 * | Portugal |
| Bursaphelencus mucronatus | BmCh3 * | Portugal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, M.J.; Inácio, M.L.; Mota, M.; de Andrade, E. Development and Validation of a Loop-Mediated Isothermal Amplification Diagnostic Method to Detect the Quarantine Potato Pale Cyst Nematode, Globodera pallida. Pathogens 2021, 10, 744. https://doi.org/10.3390/pathogens10060744
Camacho MJ, Inácio ML, Mota M, de Andrade E. Development and Validation of a Loop-Mediated Isothermal Amplification Diagnostic Method to Detect the Quarantine Potato Pale Cyst Nematode, Globodera pallida. Pathogens. 2021; 10(6):744. https://doi.org/10.3390/pathogens10060744
Chicago/Turabian StyleCamacho, Maria João, Maria L. Inácio, Manuel Mota, and Eugénia de Andrade. 2021. "Development and Validation of a Loop-Mediated Isothermal Amplification Diagnostic Method to Detect the Quarantine Potato Pale Cyst Nematode, Globodera pallida" Pathogens 10, no. 6: 744. https://doi.org/10.3390/pathogens10060744
APA StyleCamacho, M. J., Inácio, M. L., Mota, M., & de Andrade, E. (2021). Development and Validation of a Loop-Mediated Isothermal Amplification Diagnostic Method to Detect the Quarantine Potato Pale Cyst Nematode, Globodera pallida. Pathogens, 10(6), 744. https://doi.org/10.3390/pathogens10060744