New Insight into the Systematics of European Lepidocyrtus (Collembola: Entomobryidae) Using Molecular and Morphological Data



Abstract
1. Introduction
2. Materials and Methods
2.1. Species Sampling and Processing
2.2. Molecular Methods
2.2.1. DNA Extraction and Sequencing
2.2.2. Data Analyses
2.2.3. Phylogenetic Analyses
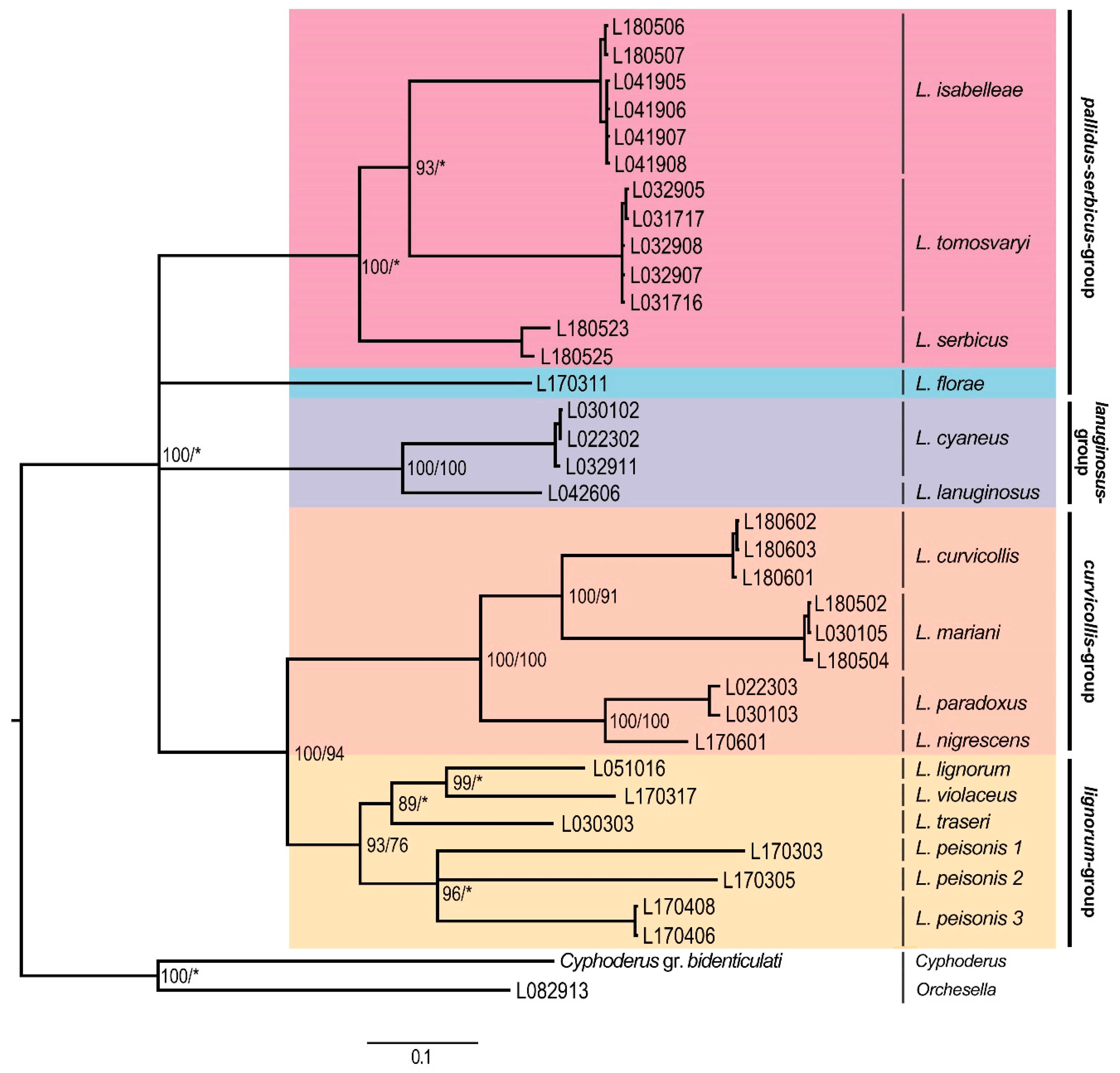
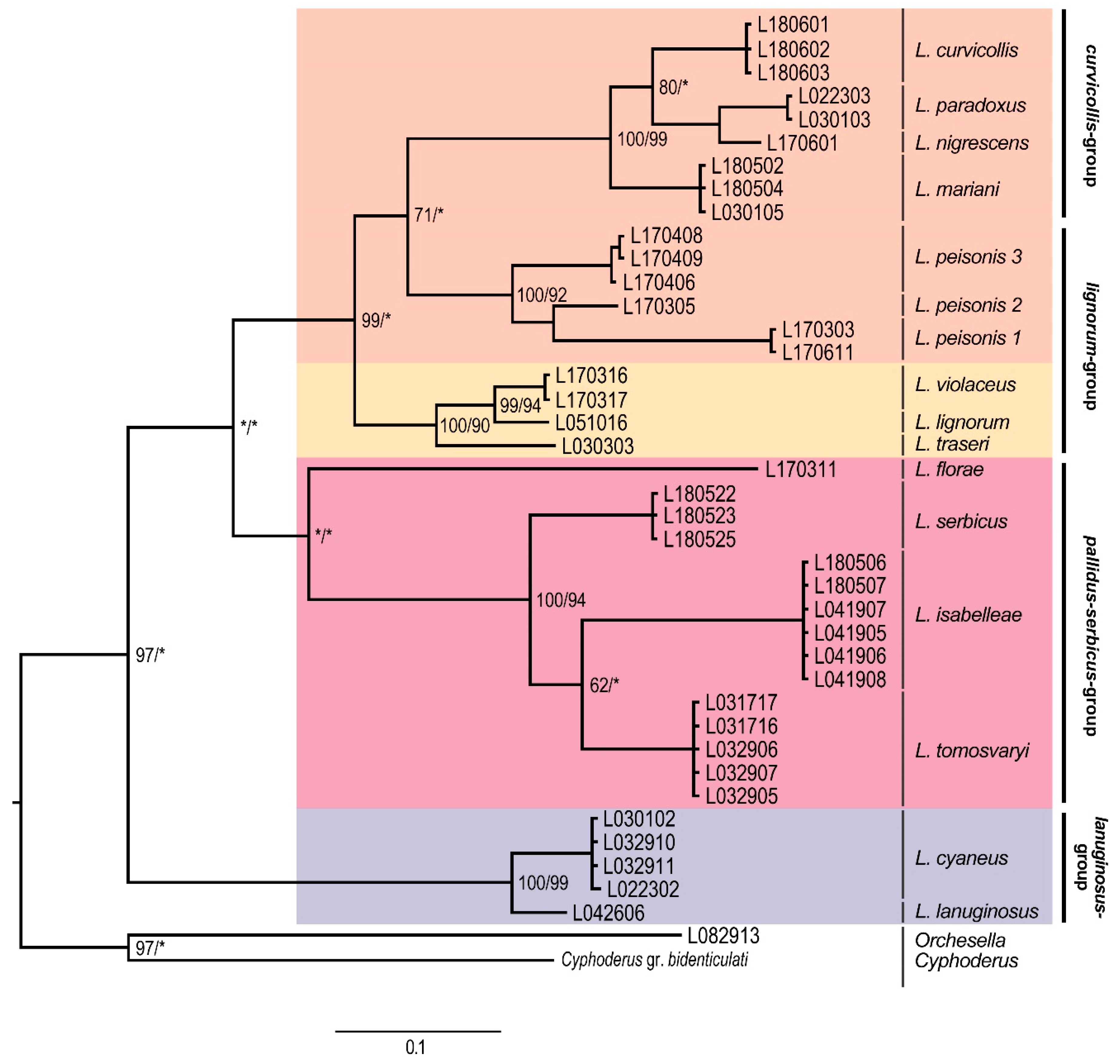
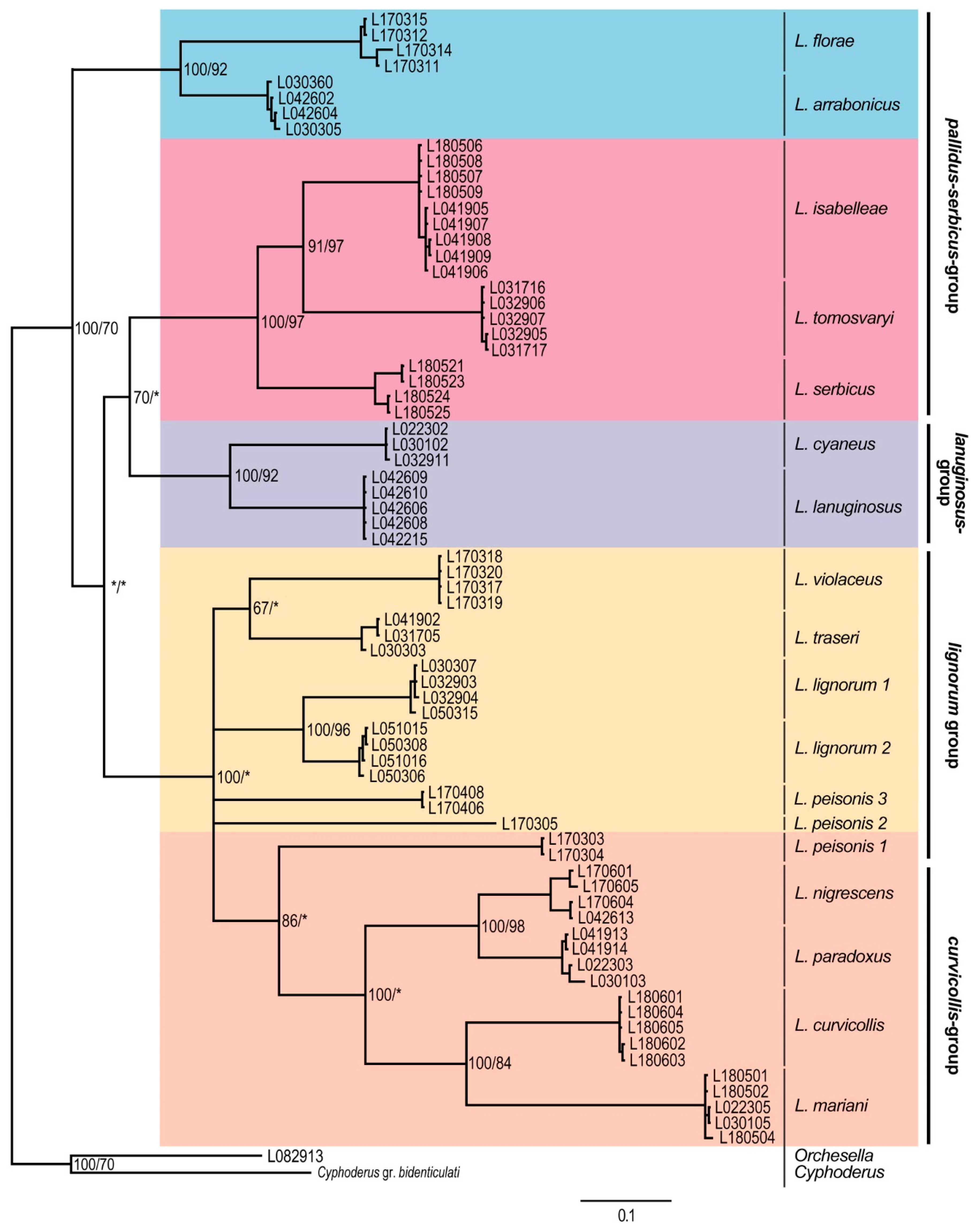
3. Results
Molecular Analyses
4. Discussion
4.1. Subgeneric Aspects of the European Lepidocyrtus
4.2. Lepidocyrtus Species Groups
4.3. Cryptic Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yoshii, R. Lepidocyrtid Collembola of Sabah. Entomol. Rep. Sabah For. Res. 1982, 5, 1–47. [Google Scholar]
- Deharveng, L. Recent advances in Collembola systematics. Pedobiologia 2004, 48, 415–433. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 14 February 2020).
- Winkler, D. A new species of Lepidocyrtus (Collembola, Entomobryidae) from the Börzsöny Mountains, Hungary. Zootaxa 2016, 4150, 388–400. [Google Scholar] [CrossRef]
- Querner, P.; Milasowszky, N.; Zulka, K.P.; Abensperg-Traun, M.; Willner, W.; Sauberer, N.; Jakomini, C.; Wrbka, T.; Schmitzberger, I.; Zechmeister, H.G. Habitat Structure, Quality and Landscape Predict Species Richness and Communities of Collembola in Dry Grasslands in Austria. Insects 2018, 9, 81. [Google Scholar] [CrossRef]
- Dányi, L. Cave dwelling springtails (Collembola) of Hungary: A review. Soil Org. 2011, 83, 419–432. [Google Scholar]
- Fiera, C.; Habel, J.C.; Ulrich, W. Neutral colonisations drive high beta-diversity in cavernicole springtails (Collembola). PLoS ONE 2018, 13, e0189638. [Google Scholar] [CrossRef]
- Arbea, J.I.; Jordana, R. New species of Pseudosinella and Lepidocyrtus from Navarra (Northern Iberian Peninsula). Spixiana 1989, 13, 25–31. [Google Scholar]
- Cipola, N.G.; de Morais, J.W.; Bellini, B.C. New subgenus and four species of Lepidocyrtus Bourlet (Collembola, Entomobryidae, Lepidocyrtinae) from Amazon. Insect Syst. Evol. 2018, 4027, 189–234. [Google Scholar] [CrossRef]
- Traser, G.; Christian, E. Lepidocyrtus peisonis sp. n., ein neuer Springschwanz aus dem Neusiedlersee-Gebiet (Collembola: Entomobryidae). Folia Entomol. Hung. 1992, 52, 119–121. [Google Scholar]
- Cicconardi, F.; Nardi, F.; Emerson, B.C.; Frati, F.; Fanciulli, P.P. Deep phylogeographic divisions and long-term persistence of forest invertebrates (Hexapoda: Collembola) in the North-Western Mediterranean basin. Mol. Ecol. 2010, 19, 386–400. [Google Scholar] [CrossRef]
- Cicconardi, F.; Fanciulli, P.P.; Emerson, B.C. Collembola, the biological species concept and the underestimation of global species richness. Mol. Ecol. 2013, 22, 5382–5396. [Google Scholar] [CrossRef] [PubMed]
- Mateos, E.; Escuer, P.; Busmachiu, G.; Riutort, M.; Álvarez-Presas, M. Untangling Lepidocyrtus (Collembola, Entomobryidae): New molecular data shed light on the relationships of the European groups. Invertebr. Syst. 2018, 32, 639–651. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, T.W.; Mateos, E.; Scheu, S.; Schaefer, I. Cryptic species in Lepidocyrtus lanuginosus (Collembola: Entomobryidae) are sorted by habitat type. Pedobiologia 2018, 68, 12–19. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, T.W.; Mateos, E.; Scheu, S.; Schaefer, I. DNA-based approaches uncover cryptic diversity in the European Lepidocyrtus lanuginosus species group (Collembola: Entomobryidae). Invertebr. Syst. 2019, 33, 661–670. [Google Scholar] [CrossRef]
- Gisin, H. Collembolenfauna Europas; Museum d’Histoire Naturelle: Genève, Switzerland, 1960; pp. 1–312. [Google Scholar]
- Gisin, H. Collemboles d’Europe. VI. Rev. Suisse Zool. 1964, 71, 383–400. [Google Scholar] [CrossRef]
- Gisin, H. Collemboles d’Europe VII. Rev. Suisse Zool. 1964, 71, 649–678. [Google Scholar]
- Gisin, H. Nouvelles notes taxonomiques sur les Lepidocyrtus. Rev. Ecol. Biol. Sol 1965, 2, 519–524. [Google Scholar]
- Gisin, H. La systématique idéale. J. Zool. Syst. Evol. Res. 1967, 5, 111–128. [Google Scholar] [CrossRef]
- Szeptycki, A. Morpho-systematic studies on Collembola III. Body chaetotaxy in the first instars of several genera of the Entomobryomorpha. Acta Zool. Cracov. 1972, 17, 341–372. [Google Scholar]
- Szeptycki, A. Chaetotaxy of the Entomobryidae and its Phylogenetical Significance. Morpho-Systematic Studies of Collembola, IV; Polska Akademia Nauk, Zakład Zoologii Systematycznej i Doświadczalnej: Kraków, Poland, 1979; pp. 1–219. [Google Scholar]
- Mateos, E. Definition of Lepidocyrtus lusitanicus Gama, 1964 species-complex (Collembola, Entomobryidae), with description of new species and color forms from the Iberian Peninsula. Zootaxa 2008, 1917, 38–54. [Google Scholar] [CrossRef]
- Mateos, E. New Lepidocyrtus Bourlet, 1839 taxa from Greece (Collembola: Entomobryidae). Zootaxa 2011, 3108, 25–40. [Google Scholar] [CrossRef]
- Mateos, E. The European Lepidocyrtus lanuginosus group (Collembola: Entomobryidae), definition and description of a new species from Spain. Zootaxa 2012, 3570, 69–81. [Google Scholar] [CrossRef]
- Mateos, E.; Petersen, H. Definition of the European Lepidocyrtus curvicollis group (Collembola: Entomobryidae) with description of a new species from Sardinia (Italy). Zootaxa 2012, 3273, 51–62. [Google Scholar] [CrossRef]
- Winkler, D.; Traser, G. Explanation of the European Lepidocyrtus pallidus–serbicus group (Collembola, Entomobryidae), with description of new species from Hungary. Zootaxa 2012, 3394, 35–47. [Google Scholar] [CrossRef]
- Mateos, E.; Winkler, D. New data clarifying the taxonomy of European members of the Lepidocyrtus pallidus–serbicus group (Collembola, Entomobryidae). Zootaxa 2018, 4429, 548–568. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J.T. Keys and Bibliography to the Collembola. Zool. Pub. Victoria Univ. College 1951, 8, 1–81. [Google Scholar]
- Yosii, R. Studies on the Collembolan fauna of Malay and Singapore. Contrib. Biol. Lab. Kyoto Univ. 1959, 10, 1–65. [Google Scholar]
- Yosii, R. On some Collembola of Hindukush with notes on Isotoma and its allies. Res. Kyoto Univ. Sci. Exp. Karakoram Hindukush 1963, 4, 3–42. [Google Scholar]
- Yoshii, R.; Suhardjono, Y.R. Notes on the Collembolan Fauna of Indonesia and its vicinities. I. Miscellaneous Notes, with special references to Seirini and Lepidocyrtini. AZAO 1989, 1, 23–90. [Google Scholar]
- Ma, Y. Two new species of Lepidocyrtus Bourlet s. lat. (Collembola: Entomobryidae) from China. Eur. J. Taxon. 2019, 565, 1–121. [Google Scholar] [CrossRef]
- Christiansen, K.A.; Bellinger, P.F. Phylogeny of the Hawaiian Species of the Genus Lepidocyrtus (Collembola, Entomobryidae). Rev. Ecol. Biol. Sol. 1991, 28, 207–215. [Google Scholar]
- Soto-Adames, F.N. Phylogeny of Neotropical Lepidocyrtus (Collembola: Entomobryidae): First assessment of patterns of speciation in Puerto Rico and phylogenetic relevance of some subgeneric diagnostic characters. Syst. Entomol. 2000, 25, 485–502. [Google Scholar] [CrossRef]
- Wang, F.; Chen, J.-X.; Christiansen, K. Taxonomy of the genus Lepidocyrtus s.l. (Collembola: Entomobryidae) in East and Southeast Asia and Malaysia, with description of a new species from the People’s Republic of China. Can. Entomol. 2003, 135, 823–837. [Google Scholar] [CrossRef]
- Mateos, E. The European Lepidocyrtus Bourlet, 1839 (Collembola: Entomobryidae). Zootaxa 2008, 1769, 35–59. [Google Scholar] [CrossRef]
- Yoshii, R.; Suhardjono, Y.R. Collembolan fauna of Indonesia and its affinities. II: Collembola of Irian Jaya and Maluku Island. Acta Zool. Asiae Orient. 1992, 2, 1–52. [Google Scholar]
- Mateos, E.; Greenslade, P. Towards understanding Lepidocyrtus Bourlet, 1839 (Collembola, Entomobryidae) I: Diagnosis of the subgenus Setogaster, new records and redescriptions of species. Zootaxa 2015, 4044, 105–129. [Google Scholar] [CrossRef]
- Winkler, D.; Mateos, E. Redescription of Lepidocyrtus peisonis Traser & Christian, 1992 with notes on Lepidocyrtus mariani Traser & Dányi, 2008 (Collembola: Entomobryidae). Zootaxa 2018, 4375, 392–408. [Google Scholar] [CrossRef]
- Arbea, I.; Jordana, R. Colémbolos de las Islas Baleares (Insecta, Collembola). Redia 1990, 73, 187–200. [Google Scholar]
- Bonnet, L.; Cassagnau, P.; Deharveng, L. Recherche d’une méthodologie dans l’analyse de la rupture des équilibres biocénotiques: Application aux Collemboles édaphiques des Pyrénées. Rev. Ecol. Biol. Sol. 1979, 16, 373–401. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X Windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Delamare-Deboutteville, C. Recherches sur les Collemboles termitophiles et myrmécophiles (Ecologie, Ethologie, Systématique). Archs. Zool. Exp. Gen. 1948, 85, 261–425. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree, ver. 1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 December 2019).
- Varga, Z. Extra-Mediterranean Refugia, Post-Glacial Vegetation History and Area Dynamics in Eastern Central Europe. In Relict Species–Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Heidelberg, Germany, 2010; pp. 57–87. [Google Scholar]
- Dányi, L.; Traser, G. Springtails of Hungary. In The Origin of the Fauna of the Carpathian Basin; Forró, L., Ed.; Hungarian Natural History Museum: Budapest, Hungary, 2007; pp. 21–28. [Google Scholar]
- Winkler, D. New Lepidocyrtus Bourlet, 1839 from riverine woodland in Hungary (Collembola, Entomobryidae). Zootaxa 2017, 4250, 529–540. [Google Scholar] [CrossRef]
- Mari Mutt, J.A. Puerto Rican species of Lepidocyrtus and Pseudosinella (Collembola: Entomobryidae). Caribb. J. Sci. 1986, 22, 1–48. [Google Scholar]
- Xu, G.-L.; Pan, Z.-H.; Zhang, F. First record of Acrocyrtus Yosii, 1959 (Collemobla, Entomobryidae) from Chinese mainland. ZooKeys 2013, 260, 1–16. [Google Scholar] [CrossRef][Green Version]
- Handschin, E. Hygrophile Collembolen aus Niederländische-Indian. Arch. Hydrobiol. 1932, 9, 472–490. [Google Scholar]
- Deharveng, L.; Jantarit, S.; Bedos, A. Revisiting Lepidonella Yosii (Collembola: Paronellidae): Character overview, checklist of world species and reassessment of Pseudoparonella doveri Carpenter. Ann. Soc. Entomol. Fr. 2018, 54, 381–400. [Google Scholar] [CrossRef]
- Soto-Adames, F.N. Molecular phylogeny of the Puerto Rican Lepidocyrtus and Pseudosinella (Hexapoda: Collembola), a validation of Yoshii’s ‘colour pattern species’. Mol. Phylogenet. Evol. 2002, 25, 27–42. [Google Scholar] [CrossRef]
- Zhang, F.; Deharveng, L. Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 2015, 44, 298–311. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Nr of Spec. | Municipality | Habitat | Position (WGS 84) | Date of Collection |
|---|---|---|---|---|---|---|
| Lepidocyrtus | arrabonicus | 4 | Fertőújlak (loc. typ.) | meadow | N47°40′58″ E16°50′37″ | 20.xi.2015 |
| Lepidocyrtus | curvicollis | 5 | Bajót, Gerecse | cave | N47°43′22″ E18°34′31″ | 01.vi.2018 |
| Lepidocyrtus | cyaneus | 4 | Tüskevár | grassland | N47°07′07″ E17°19′51″ | 03.xii.2015 |
| Lepidocyrtus | florae | 4 | Hanság (loc. typ.) | alder forest | N47°40′47″ E17°07′53″ | 11.iv.2016 |
| Lepidocyrtus | isabelleae | 5 | Nagykapornak | beech forest | N46°50′30″ E16°58′32″ | 15.vii.2017 |
| Lepidocyrtus | isabelleae | 4 | Rum (loc. typ.) | riverine forest | N47°05′40″ E16°49′41″ | 09.viii.2016 |
| Lepidocyrtus | lanuginosus | 5 | Sopron | urban park | N47°40′53″ E16°34′29″ | 02.iv.2016 |
| Lepidocyrtus | lignorum | 3 | Sopron | grassland | N47°41′54″ E16°38′22″ | 24.x.2015 |
| Lepidocyrtus | lignorum | 1 | Darvastó | oak forest | N46°58′56″ E17°22′12″ | 04.xii.2015 |
| Lepidocyrtus | lignorum | 4 | Kerecsend | oak forest | N47°47′23″ E20°19′27″ | 01.x.2014 |
| Lepidocyrtus | mariani | 2 | Nagykapornak | beech forest | N46°50′31″ E16°58′32″ | 01.vii.2014 |
| Lepidocyrtus | mariani | 3 | Porva (loc. typ.) | alder forest | N47°18′42″ E17°47′30″ | 25.ix.2014 |
| Lepidocyrtus | nigrescens | 3 | Kerecsend | grassland | N47°47′23″ E20°19′27″ | 01.x.2014 |
| Lepidocyrtus | nigrescens | 1 | Darány | sedge | N45°58′03″ E17°32′18″ | 03.xi.2016 |
| Lepidocyrtus | paradoxus | 2 | Fertőújlak | meadow | N47°40′58″ E16°50′37″ | 20.xi.2015 |
| Lepidocyrtus | paradoxus | 2 | Bakonygyepes | we meadow | N47°08′29″ E17°33′37″ | 02.xii.2015 |
| Lepidocyrtus | peisonis | 4 | Fertőrákos (loc. typ.) | reed-bed | N47°42′54″ E16°40′14″ | 25.iii.2017 |
| Lepidocyrtus | peisonis | 3 | Darány | wet meadow | N45°59′14″ E17°33′23″ | 03.xi.2016 |
| Lepidocyrtus | serbicus | 5 | Mátrafüred | beech | N47°53′26″ E19°56′37″ | 27.10.2016 |
| Lepidocyrtus | tomosvaryi | 5 | Tüskevár (loc. typ.) | ash forest | N47°07′13″ E17°20′04″ | 02.xii.2015 |
| Lepidocyrtus | traseri | 3 | Börzsöny (loc. typ.) | grassland | N47°53′55″ E18°51′08″ | 08.x.2014 |
| Lepidocyrtus | violaceus | 5 | Csákvár | oak forest | N47°25′33″ E18°25′51″ | 13.ix.2016 |
| Orchesella | cincta | 1 | Somlóvásárhely | grassland | N47° 8′43″ E17°22′35″ | 26.vi.2017 |
| Cyphoderus | gr. bidenticulati | 1 | – | – | – | – |
| Species Group/Species | Head Mac | Body Mac | Scales | Th. II | Abd. IV s | Abd. IV pse (dl-l) | Lateral Tuft | Dental Tubercle | Unguiculus |
|---|---|---|---|---|---|---|---|---|---|
| L. lanuginosus-group | |||||||||
| L. cyaneus | A0 A2 A3 M2 S3 Pa5 | 10/0101+2 | – | n | – | – | – | – | A |
| L. lanuginosus | A0 A2 A3 M2 S3 Pa5 | 10/0101+2 | – | n | – | – | – | – | A |
| L. pallidus–serbicus group | |||||||||
| L. arrabonicus | A0 A2 A3 Pa5 | 00/0101+2 | – | n | – | – | – | – | A |
| L. florae | A0 A2 A3 S3 Pa5 | 00/0101+2 | – | n | – | – | – | – | A |
| L. isabelleae | A0 A2 A3 M2 S3 Pa5 | 00/0101+2 | – | n | – | – | – | – | A |
| L. serbicus | A0 A2 A3 M2 S3 Pa5 | 00/0101+2 | – | n | – | – | – | – | A |
| L. tomosvaryi | A0 A2 A3 M2 S3 Pa5 | 00/0101+2 | – | n | – | – | – | + | A |
| L. lignorum group | |||||||||
| L. lignorum | A0 A2 A3 Pa5 | 00/0101+3 | + | p | – | – | – | – | A |
| L. peisonis | A0 A2 A3 Pa5 | 00/0101+3 | + | p | – | – | + | – | T |
| L. traseri | A0 A2 A3 Pa5 | 00/0101+3 | + | p | – | – | – | – | A |
| L. violaceus | A0 A2 A3 Pa5 | 00/0101+3 | + | p | – | – | – | – | A |
| L. curvicollis group | |||||||||
| L. curvicollis | A0 A2 Pa5 | 00/0101+3 | + | P | + | + | – | – | A |
| L. mariani | A0 A2 Pa5 | 00/0101+3 | + | P | + | + | – | – | A |
| L. nigrescens | A0 A2 Pa5 | 00/0101+3 | + | P | + | – | – | – | A |
| L. paradoxus | A0 A2 Pa5 | 00/0101+3 | + | P | + | – | – | – | A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winkler, D.; Mateos, E.; Traser, G.; Lakatos, F.; Tóth, V. New Insight into the Systematics of European Lepidocyrtus (Collembola: Entomobryidae) Using Molecular and Morphological Data. Insects 2020, 11, 302. https://doi.org/10.3390/insects11050302
Winkler D, Mateos E, Traser G, Lakatos F, Tóth V. New Insight into the Systematics of European Lepidocyrtus (Collembola: Entomobryidae) Using Molecular and Morphological Data. Insects. 2020; 11(5):302. https://doi.org/10.3390/insects11050302
Chicago/Turabian StyleWinkler, Daniel, Eduardo Mateos, György Traser, Ferenc Lakatos, and Viktória Tóth. 2020. "New Insight into the Systematics of European Lepidocyrtus (Collembola: Entomobryidae) Using Molecular and Morphological Data" Insects 11, no. 5: 302. https://doi.org/10.3390/insects11050302
APA StyleWinkler, D., Mateos, E., Traser, G., Lakatos, F., & Tóth, V. (2020). New Insight into the Systematics of European Lepidocyrtus (Collembola: Entomobryidae) Using Molecular and Morphological Data. Insects, 11(5), 302. https://doi.org/10.3390/insects11050302