Transcriptome Analysis of Gene Expression Profiles of Tomato Yellow Leaf Curl Virus-Infected Whiteflies over Different Viral Acquisition Access Periods

Abstract
1. Introduction
2. Materials and Methods
2.1. Whitefly Samples for RNA-Seq
2.2. RNA Isolation and cDNA Library Constructions for RNA-Seq
2.3. Transcriptiome Sequencing and Clean Reads Mapping
2.4. DEGs Analysis
2.5. GO and KEGG Analysis
2.6. qPCR Validation
3. Results
3.1. Sequencing and Annotation of Transcriptomes
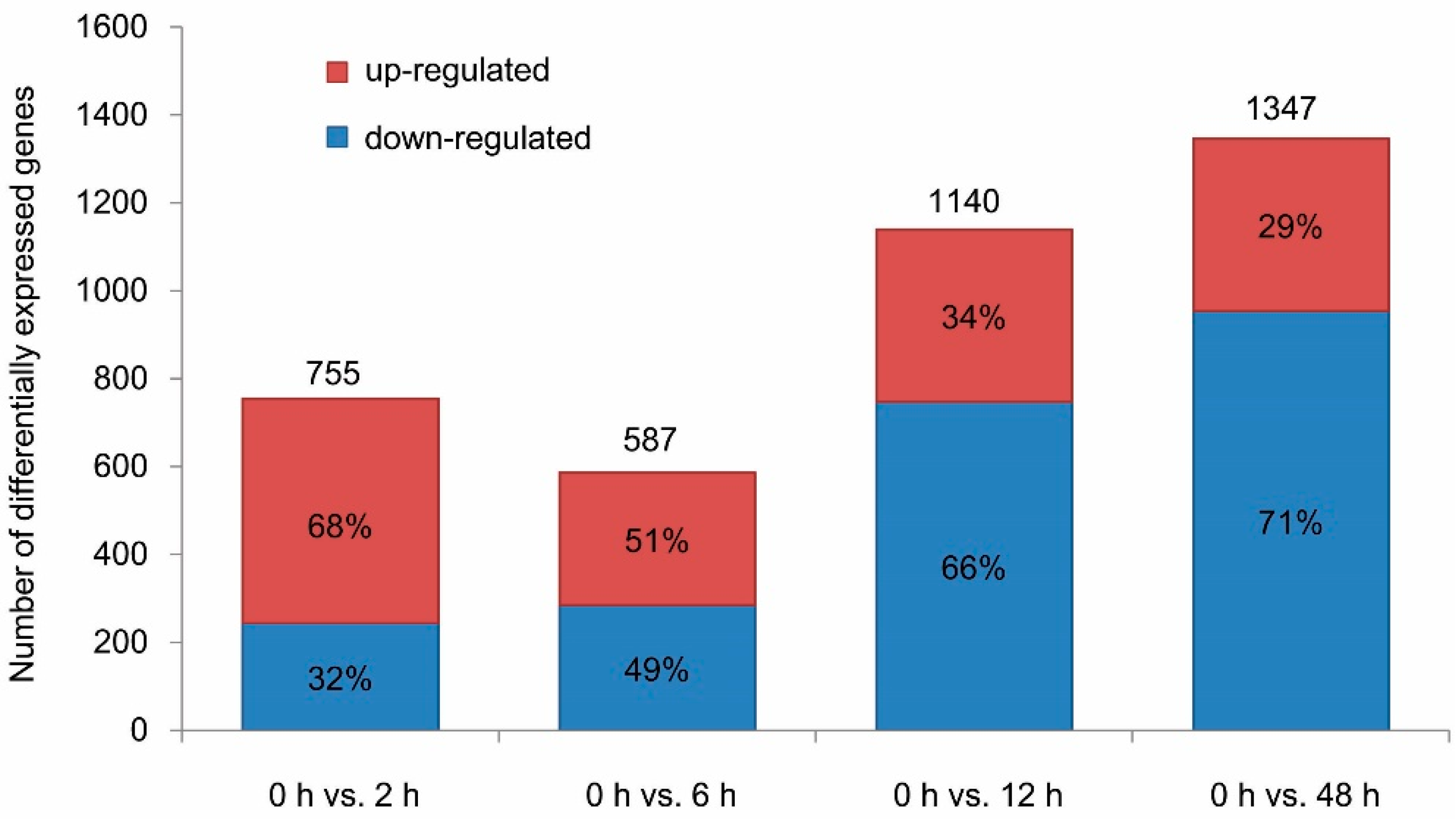
3.2. DEGs in Virus-Infected Whiteflies
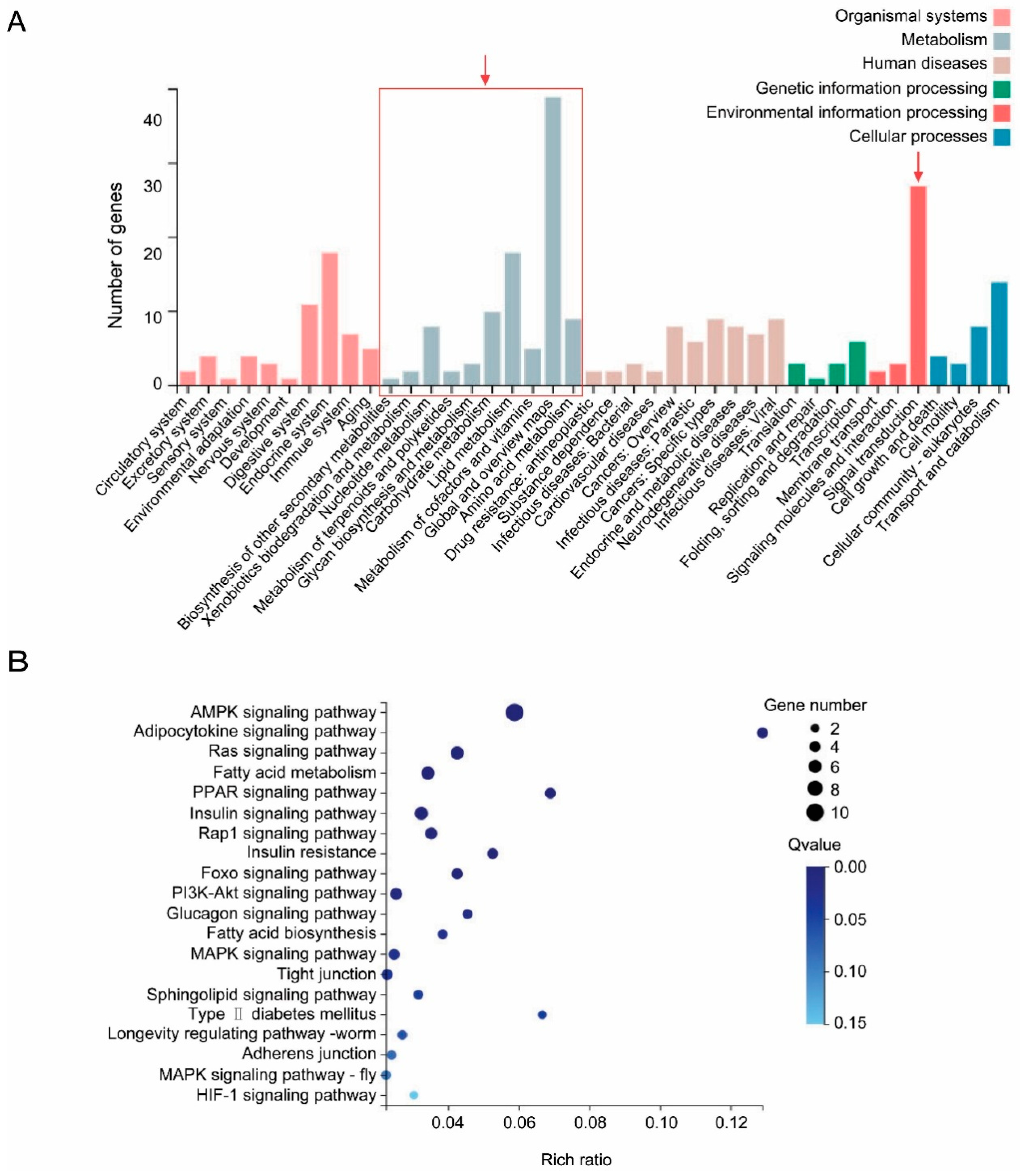
3.3. Common DEGs in Virus–Infected Whiteflies Indicated the Perturbance of Vector Metabolisms and Signal Transductions by Tylcv Infection
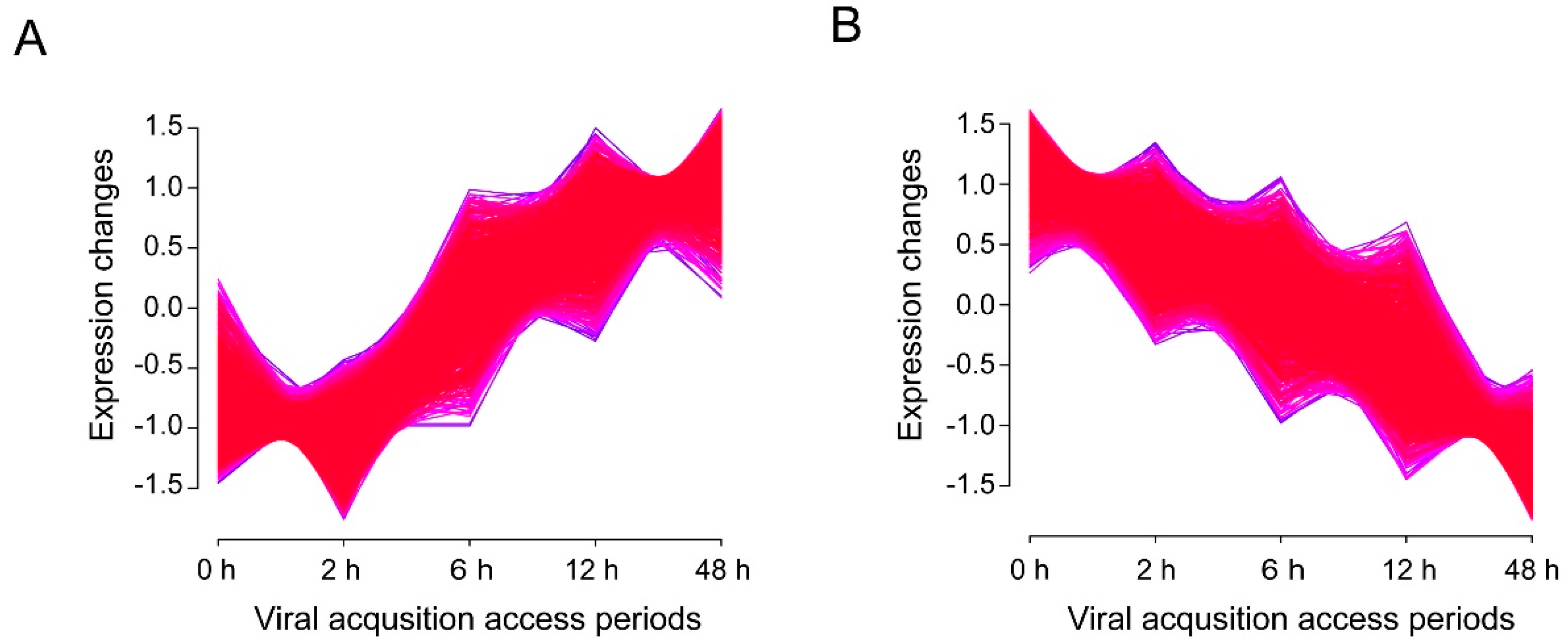
3.4. Time-Associated Gene Cluster Analysis Suggested Activated Transporters But Impaired Il-17 Signaling Pathway in Virus-Infected Whiteflies
3.5. qPCR Validation of DEGs Identified by RNA-Seq
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Sauder, S.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Elena, S.F.; Fraile, A.; García-Arenal, F. Evolution and emergence of plant viruses. Adv. Virus Res. 2014, 88, 161–191. [Google Scholar] [PubMed]
- Ng, J.C.; Falk, B.W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 2006, 44, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef]
- Morales, F.J. Bemisia: Bionomics and management of a global pest. In Distribution and Dissemination of Begomoviruses in Latin America and the Caribbean; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Heidelberg, Germany, 2010; pp. 283–318. [Google Scholar]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Harrison, B.; Robinson, D. Natural genomic and antigenic variation in whitefly-transmitted geminiviruses (begomoviruses). Annu. Rev. Phytophathol. 1999, 37, 369–398. [Google Scholar] [CrossRef]
- Rosen, R.; Kanakala, S.; Kliot, A.; Pakkianathan, B.C.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Legg, J.P.; Shirima, R.; Tajebe, L.S.; Guastella, D.; Boniface, S.; Jeremiah, S.; Nsami, E.; Chikoti, P.; Rapisarda, C. Biology and management of Bemisia whitefly vectors of cassava virus pandemics in Africa. Pest Manag. Sci. 2014, 70, 1446–1453. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Liu, S.S.; Colvin, J.; De Barro, P.J. Species concepts as applied to the whitefly Bemisia tabaci systematics: How many species are there? J. Integr. Agric. 2012, 11, 176–186. [Google Scholar] [CrossRef]
- Mugerwa, H.; Seal, S.; Wang, H.L.; Patel, M.V.; Kabaalu, R.; Omongo, C.A.; Alicai, T.; Tairo, F.; Ndunguru, J.; Sseruwagi, P.; et al. African ancestry of New World, Bemisia tabaci-whitefly species. Sci. Rep. 2018, 8, 2734. [Google Scholar] [CrossRef]
- Li, M.; Hu, J.; Xu, F.C.; Liu, S.S. Transmission of Tomato yellow leaf curl virus by two invasive biotypes and a Chinese indigenous biotype of the whitefly Bemisia tabaci. Int. J. Pest. Manag. 2010, 56, 275–280. [Google Scholar] [CrossRef]
- Li, J.M.; Ruan, Y.M.; Li, F.F.; Liu, S.S.; Wang, X.W. Gene expression profiling of the whitefly (Bemisia tabaci) Middle East-Asia Minor 1 feeding on healthy and Tomato yellow leaf curl China virus-infected tobacco. Insect Sci. 2011, 18, 11–22. [Google Scholar] [CrossRef]
- Luan, J.B.; Li, J.M.; Varela, N.; Wang, Y.L.; Li, F.F.; Bao, Y.Y.; Zhang, C.X.; Liu, S.S.; Wang, X.W. Global analysis of the transcriptional response of whitefly to Tomato yellow leaf curl China virus reveals the relationship of coevolved adaptations. J. Virol. 2011, 85, 3330–3340. [Google Scholar] [CrossRef]
- Hasegawa, D.K.; Chen, W.; Zheng, Y.; Kaur, N.; Wintermantel, W.M.; Simmons, A.M.; Fei, Z.; Ling, K. Comparative transcriptome analysis reveals networks of genes activated in the whitefly, Bemisia tabaci, when fed on tomato plants infected with Tomato yellow leaf curl virus. Virology 2018, 513, 52–64. [Google Scholar] [CrossRef]
- Geng, L.; Qian, L.X.; Shao, R.X.; Liu, Y.Q.; Liu, S.S.; Wang, X.W. Transcriptome profiling of whitefly guts in response to Tomato yellow leaf curl virus infection. Virol. J. 2018, 15, 14. [Google Scholar] [CrossRef]
- Zhao, J.; Chi, Y.; Zhang, X.J.; Lei, T.; Wang, X.W.; Liu, S.S. Comparative proteomic analysis provides new insight into differential transmission of two begomoviruses by a whitefly. Virol. J. 2019, 16, 32. [Google Scholar] [CrossRef]
- Ghanim, M.; Morin, S.; Czosnek, H. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology 2001, 91, 188–196. [Google Scholar] [CrossRef]
- Czosnek, H.; Ghanim, M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci-insights from studies with Tomato yellow leaf curl virus. Annu. Appl. Biol. 2002, 140, 215–231. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton–Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The incredible journey of begomoviruses in their whitefly vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.L.; Chen, Q.F.; Zhao, J.; Guo, T.; Wang, X.W.; Hariton-Shalev, A.; Czosnek, H.; Liu, S.S. Clathrin-mediated endocytosis is involved in Tomato yellow leaf curl virus transport across the midgut barrier of its whitefly vector. Virology 2017, 502, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Min, T.; Bao, Y.; Zhou, B.; Yi, Y.; Wang, L.; Hou, W.; Ai, Y.; Wang, H. Transcription profiles reveal the regulatory synthesis of phenols during the development of lotus rhizome (Nelumbo nucifera Gaertn). Int. J. Mol. Sci. 2019, 20, 2735. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Kumar, L.; Futschik, M.E. Mfuzz: A software package for soft clustering of microarray data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef]
- Xu, H.X.; Hong, Y.; Zhang, M.Z.; Wang, Y.L.; Liu, S.S.; Wang, X.W. Transcriptional responses of invasive and indigenous whiteflies to different host plants reveal their disparate capacity of adaptation. Sci. Rep. 2015, 5, 10774. [Google Scholar] [CrossRef]
- Maramorosch, K.; Jensen, D.D. Harmful and beneficial effects of plant viruses in insects. Annu. Rev. Microbiol. 1963, 17, 495–530. [Google Scholar] [CrossRef]
- Rubinstein, G.; Czosnek, H. Long-term association of Tomato yellow leaf curl virus with its whitefly vector Bemisia tabaci: Effect on the insect transmission capacity, longevity and fecundity. J. Gen. Virol. 1997, 78, 2683–2689. [Google Scholar] [CrossRef]
- Pusag, J.C.A.; Hemayet Jahan, S.M.; Lee, K.S.; Lee, S.; Lee, K.Y. Upregulation of temperature susceptibility in Bemisia tabaci upon acquisition of Tomato yellow leaf curl virus (TYLCV). J. Insect Physiol. 2012, 58, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Chen, W.B.; Zheng, Y.; Hasegawa, D.K.; Ling, K.S.; Fei, Z.J.; Wintermantel, W.M. Transcriptome analysis of the whitefly, Bemisia tabaci MEAM1 during feeding on tomato infected with the crinivirus, Tomato chlorosis virus, identifies a temporal shift in gene expression and differential regulation of novel orphan genes. BMC Genom. 2017, 18, 370. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. AMPK and autophagy get connected. Embo J. 2011, 30, 634–635. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signaling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.R.; Dudek, H.; Tao, X. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Zhou, H.; Li, X.M.; Meinkoth, J.; Pittman, R.N. Akt regulates cell survival and apoptosis at a postmitochondrial level. J. Cell Biol. 2000, 151, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Rusten, T.E.; Lindmo, K.; Juhász, G.; Sass, M.; Seglen, P.O.; Brech, A.; Stenmark, H. Programmed autophagy in the Drosophila fat body is induced by ecdysone through regulation of the PI3K pathway. Dev. Cell 2004, 7, 179–192. [Google Scholar] [CrossRef]
- Berry, D.L.; Baehrecke, E.H. Growth arrest and autophagy are required for salivary gland cell degradation in Drosophila. Cell 2007, 131, 1137–1148. [Google Scholar] [CrossRef]
- Liu, H.J.; Ji, W.T. PI3K–Akt signaling and viral infection. Recent. Pat. Biotechnol. 2008, 2, 218–226. [Google Scholar]
- Guo, H.T.; Zhou, T.L.; Jiang, D.; Cuconati, A.; Xiao, G.H.; Block, T.M.; Guo, J.T. Regulation of hepatitis B virus replication by the phosphatidylinositol 3-kinase-akt signal transduction pathway. J. Virol. 2007, 81, 10072–10080. [Google Scholar] [CrossRef]
- Mannova, P.; Beretta, L. Activation of the N-Ras-PI3K-Akt-mTOR pathway by hepatitis C virus, control of cell survival and viral replication. J. Virol. 2005, 79, 8742–8749. [Google Scholar] [CrossRef] [PubMed]
- Fothergill, T.; McMillan. N.A.J. Papillomavirus virus–like particles activate the PI3-kinase pathway via alpha-6 beta-4 integrin upon binding. Virology 2006, 352, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Menges, C.W.; Baglia, L.A.; Lapoint, R.; McCance, D.J. Human papillomavirus type 16 E7 up-regulates Akt activity through the etinoblastoma protein. Cancer Res. 2006, 66, 5555–5559. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.M.; Liang, S.M.; Liang, C.M. VP1 of foot-and-mouth disease virus induces apoptosis via the Akt signaling pathway. J. Biol. Chem. 2004, 279, 52168–52174. [Google Scholar] [CrossRef]
- Surjit, M.; Liu, B.; Jameel, S.; Chow, V.T.K.; Lal, S.K. The SARS coronavirus nucleocapsid protein induces actin reorganization and apoptosis in COS-1 cells in the absence of growth factors. Biochem. J. 2004, 383, 13–18. [Google Scholar] [CrossRef]
- Qian, Y.; Kang, Z.; Liu, C.; Li, X. IL-17 signaling in host defense and inflammatory diseases. Cell. Mol. Immunol. 2010, 7, 328–333. [Google Scholar] [CrossRef]
- Wiehler, S.; Proud, D. Interleukin-17A modulates human airway epithelial responses to human rhinovirus infection. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L505–L515. [Google Scholar] [CrossRef]
- Crowe, C.R.; Chen, K.; Pociask, D.A.; Alcorn, J.F.; Krivich, C.; Enelow, R.I.; Ross, T.M.; Witztum, J.L.; Kolls, J.K. Critical role of IL-17RA in immunopathology of influenza infection. J. Immunol. 2009, 183, 5301–5310. [Google Scholar] [CrossRef]
- Rana, V.S.; Popli, S.; Saurav, G.K.; Raina, H.S.; Chaubey, R.; Ramamurthy, V.V.; Rajagopal, R.A. Bemisia tabaci midgut protein interacts with begomoviruses and plays a role in virus transmission. Cell. Microbiol. 2015, 18, 663–678. [Google Scholar] [CrossRef]
- Kanakala, S.; Ghanim, M. Implication of the whitefly Bemisia tabaci cyclophilin B protein in the transmission of Tomato yellow leaf curl virus. Front. Plant. Sci. 2016, 7, 1702. [Google Scholar] [CrossRef]
- Rana, V.S.; Popli, S.; Saurav, G.K.; Raina, H.S.; Jamwal, R.; Chaubey, R.; Ramamurthy, V.V.; Natarajan, K.; Rajagopal, R. Implication of the whitefly, Bemisia tabaci, collagen protein in begomoviruses acquisition and transmission. Phytopathology 2019, 108, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Götz, M.; Popovski, S.; Kollenberg, M.; Gorovits, R.; Brown, J.K.; Cicero, J.M.; Czosnek, H.; Winter, S.; Ghanim, M. Implication of Bemisia tabaci heat shock protein 70 in begomovirus-whitefly interactions. J. Virol. 2012, 86, 13241–13252. [Google Scholar] [CrossRef] [PubMed]
- Aliza, H.S.; Iris, S.; Murad, G.; Liu, S.S.; Henryk, C. The whitefly Bemisia tabaci knottin-1 gene is implicated in regulating the quantity of Tomato yellow leaf curl virus ingested and transmitted by the insect. Viruses 2016, 8, 205. [Google Scholar] [CrossRef]
- Zhao, J.; Chi, Y.; Zhang, X.J.; Wang, X.W.; Liu, S.S. Implication of whitefly vesicle associated membrane protein-associated protein B in the transmission of Tomato yellow leaf curl virus. Virology 2019, 535, 210–217. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Product Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| PCR TYLCV-detection-F | ATCGAAGCCCTGATGTTCCTCGTGG | 683 | 55 |
| TYLCV-detection-R | CAGAGCAGTTGATCATGTATTGTATG | ||
| qPCR TYLCV-qPCR-F | GAAGCGACCAGGCGATATAA | 192 | 60 |
| TYLCV-qPCR-R | GGAACATCAGGGCTTCGATA | ||
| β-Actin-qPCR-F | TCTTCCAGCCATCCTTCTTG | 174 | 60 |
| β-Actin-qPCR-R | CGGTGATTTCCTTCTGCATT | ||
| Bta04892-F | TCGGCAAGCCCTACATCATC | 145 | 60 |
| Bta04892-R | GGGCAGACCTCTTCAACGAT | ||
| Bta12976-F | CCGACCAATGTGAAGGGTCA | 77 | 60 |
| Bta12976-R | AGTTTGTTGGTGGCGAAGGA | ||
| Bta12500-F | CAACCCTGGTCACGTATCCT | 145 | 60 |
| Bta12500-R | ACTCCTTCAAATTCCCCCGC | ||
| Bta05614-F | CCGGCTTTTTCGACTATGCG | 77 | 60 |
| Bta05614-R | TGTCGGGGTCGAATCTGTTG | ||
| Bta02344-F | ACGGGTGAAGACGAGAAACC | 85 | 60 |
| Bta02344-R | ACTGATTTTCCTCGCCCTCG | ||
| Bta05747-F | GAGGCTGTCCCAGATCACAC | 146 | 60 |
| Bta05747-R | TTGGCCTTTGTAGCCACGAT | ||
| Bta04099-F | CGAGCTGTGGGACTACAAGG | 88 | 60 |
| Bta04099-R | CACCGTCTTTCGCTCGTAGA | ||
| Bta00151-F | AGGACATCAACGAGGCCAAG | 83 | 60 |
| Bta00151-R | ATTCTTCAGCTTGGACGGGG | ||
| Bta00553-F | ATTGATCGCTCGAAGCACCT | 81 | 60 |
| Bta00553-R | AACTCCTTGCTGGGATTCGG | ||
| Bta15430-F | ATTCCATCCGCGAGTGTGAG | 81 | 60 |
| Bta15430-R | TTCCATCGTTGTCACCCGTT |
| Gene ID | Accession No. | Gene Annotation | log2 (Fold Change) | |||
|---|---|---|---|---|---|---|
| 0 h vs. 2 h | 0 h vs. 6 h | 0 h vs. 12 h | 0 h vs. 48 h | |||
| Bta00780 | XP_021008519.1 | gamma-aminobutyric acid receptor subunit epsilon | −0.90 | 2.94 | 3.72 | 4.28 |
| Bta00927 | PSN49542.1 | V-type proton ATPase subunit H | - | 3.63 | 4.75 | 4.48 |
| Bta01800 | XP_018900560.1 | sodium-coupled monocarboxylate transporter 2-like | 0.13 | 1.89 | 1.61 | 2.33 |
| Bta04724 | XP_018908139.1 | solute carrier family 25 member 35-like | 0.05 | 0.97 | 1.20 | 1.31 |
| Bta04889 | XP_023109180.1 | vegetative cell wall protein gp1-like, partial | 0.34 | 3.47 | 3.83 | 3.68 |
| Bta04890 | XP_010186100.1 | uncharacterized protein DDB_G0271670-like, partial | 0.65 | 2.80 | 3.04 | 3.02 |
| Bta04892 | KFH08892.1 | chloride transporter, chloride channel (ClC) family protein | 0.61 | 3.19 | 3.56 | 3.56 |
| Bta04893 | XP_023109180.1 | vegetative cell wall protein gp1-like, partial | −0.29 | 3.76 | 4.12 | 4.05 |
| Bta04894 | XP_009899731.1 | mucin-2 | 0.58 | 1.51 | 2.15 | 2.11 |
| Bta04923 | XP_018636297.1 | chloride transporter, chloride channel (ClC) family protein | 0.79 | 2.57 | 2.88 | 2.71 |
| Bta04929 | XP_018918095.1 | uncharacterized protein LOC109044714 | 0.80 | 2.42 | 2.74 | 2.59 |
| Bta07312 | XP_018896664.1 | putative ammonium transporter 1 isoform X2 | 1.44 | 2.39 | 2.89 | 2.87 |
| Bta11245 | XP_018900402.1 | facilitated trehalose transporter Tret1-like isoform X2 | 0.36 | 0.89 | 1.25 | 1.25 |
| Bta12976 | XP_018901925.1 | facilitated trehalose transporter Tret1-like | 0.26 | 2.34 | 2.25 | 2.81 |
| Bta14126 | XP_011498176.1 | zinc finger CCCH domain-containing protein 13-like | 0.88 | 2.71 | 3.34 | 3.40 |
| Bta15384 | XP_018915842.1 | zinc transporter ZIP3-like | 1.49 | 2.96 | 2.80 | 2.86 |
| Gene ID | Accession No. | Gene Annotation | log2 (Fold Change) | |||
|---|---|---|---|---|---|---|
| 0 h vs. 2 h | 0 h vs. 6 h | 0 h vs. 12 h | 0 h vs. 48 h | |||
| Bta02344 | XP_017068262.1 | cell wall protein DAN4 | −0.28 | −0.33 | −1.28 | −1.63 |
| Bta02727 | XP_018907479.1 | adhesive plaque matrix protein–like | −0.64 | −0.40 | −1.00 | −1.26 |
| Bta02853 | XP_018908670.1 | extracellular signal–regulated kinase 2 | −0.34 | −0.84 | −0.68 | −1.13 |
| Bta06169 | XP_018896164.1 | uncharacterized protein LOC109029914 | −0.30 | −0.12 | −0.47 | −1.37 |
| Bta07157 | XP_018910204.1 | uncharacterized protein LOC109039249 | −0.18 | −0.96 | −2.37 | −1.78 |
| Bta07860 | XP_018897127.1 | uncharacterized protein LOC109030562 isoform X6 | −0.49 | −0.45 | −0.89 | −2.52 |
| Bta08110 | XP_018897436.1 | uncharacterized protein LOC109030770 | −0.26 | −0.61 | −0.92 | −1.21 |
| Bta11904 | XP_018913143.1 | uncharacterized protein LOC109041298 | −0.53 | −0.28 | −1.16 | −1.50 |
| Bta12179 | XP_018901010.1 | D(2) dopamine receptor isoform X2 | −0.94 | −0.53 | −1.15 | −1.66 |
| Bta14749 | XP_018914842.1 | uncharacterized protein LOC109042509 | −0.80 | −0.51 | −1.68 | −2.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Zhao, J.; Su, Y.-L. Transcriptome Analysis of Gene Expression Profiles of Tomato Yellow Leaf Curl Virus-Infected Whiteflies over Different Viral Acquisition Access Periods. Insects 2020, 11, 297. https://doi.org/10.3390/insects11050297
Li M, Zhao J, Su Y-L. Transcriptome Analysis of Gene Expression Profiles of Tomato Yellow Leaf Curl Virus-Infected Whiteflies over Different Viral Acquisition Access Periods. Insects. 2020; 11(5):297. https://doi.org/10.3390/insects11050297
Chicago/Turabian StyleLi, Meng, Jing Zhao, and Yun-Lin Su. 2020. "Transcriptome Analysis of Gene Expression Profiles of Tomato Yellow Leaf Curl Virus-Infected Whiteflies over Different Viral Acquisition Access Periods" Insects 11, no. 5: 297. https://doi.org/10.3390/insects11050297
APA StyleLi, M., Zhao, J., & Su, Y.-L. (2020). Transcriptome Analysis of Gene Expression Profiles of Tomato Yellow Leaf Curl Virus-Infected Whiteflies over Different Viral Acquisition Access Periods. Insects, 11(5), 297. https://doi.org/10.3390/insects11050297