Toxicity and Biological Effects of Beauveria brongniartii Fe0 Nanoparticles against Spodoptera litura (Fabricius)



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Cultures
2.2. Fungal Inoculum
2.3. Preparation of Fe0NPs
2.4. Characterization of Fe0NPs
2.5. Concentration Mortality Responses of Spodoptera litura to B. brongniartii Fe0NPs
2.6. Influence of B. brongniartii Fe0NPs on Feeding and Growth of Spodoptera litura
2.7. Effects of B. brongniartii Fe0NPs on Activities of Detoxifying Enzymes
2.8. Data Analysis
3. Results
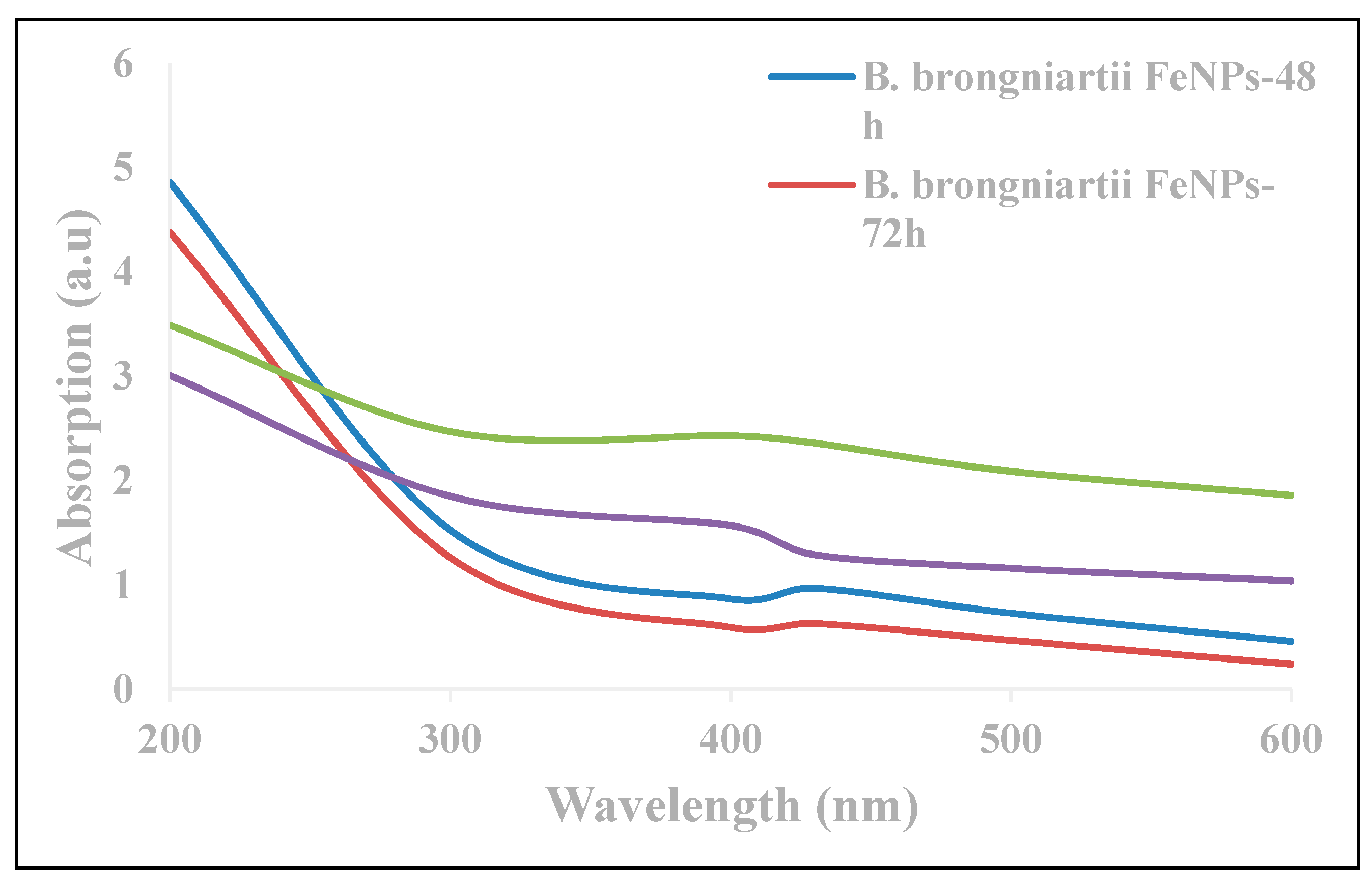
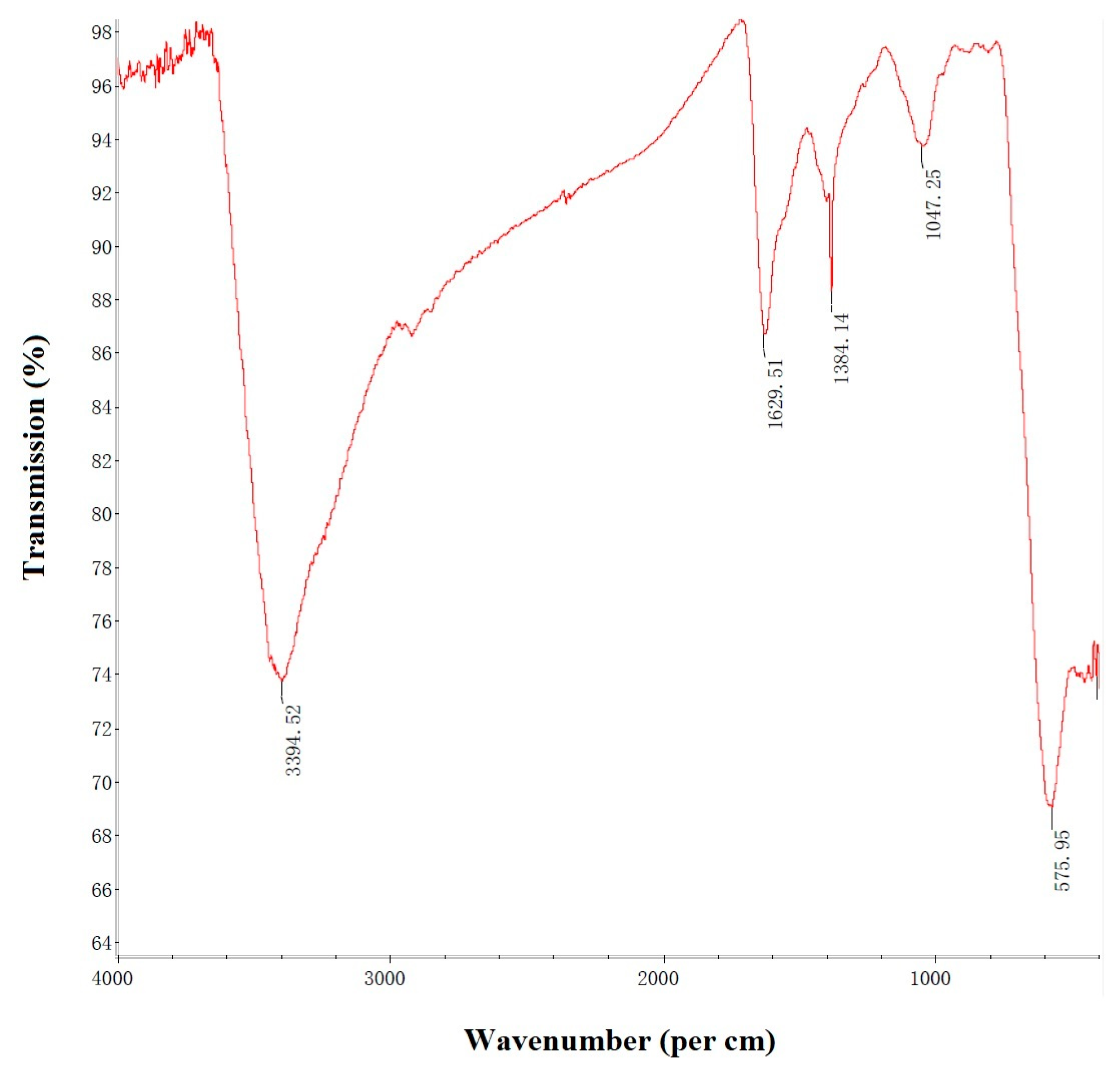
3.1. Characterization of B. brongniartii Fe0 Nanoparticles
3.2. Dose Mortality Responses of S. litura to B. brongniartii Fe0NPs
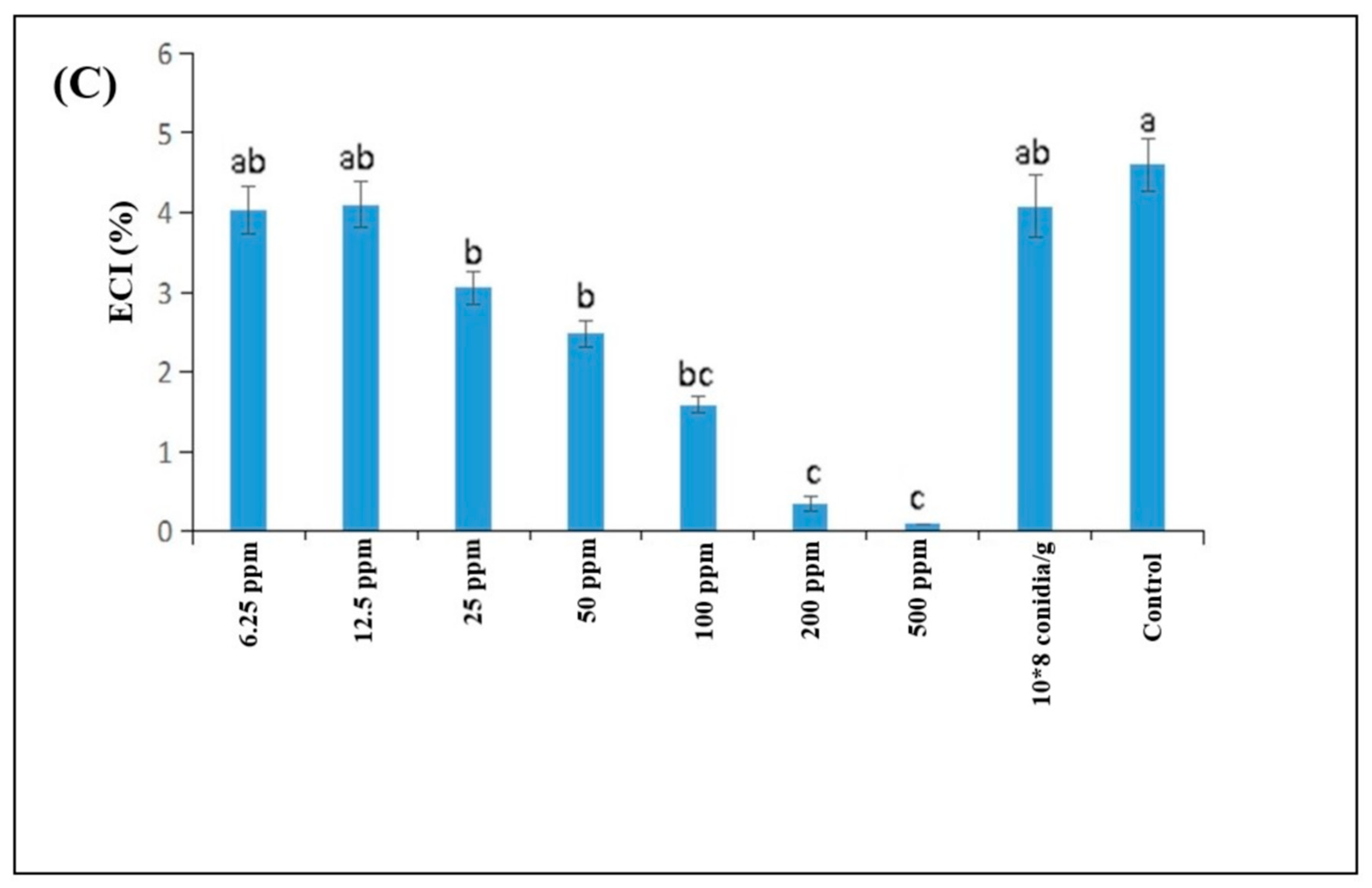
3.3. Influence of B. brongniartii Fe0NPs on Feeding and Growth of Spodoptera litura
3.4. Effects of B. brongniartii Fe0NPs on Activities of Detoxifying Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Matsuura, H.; Naito, A. Studies on the cold-hardiness and overwintering of Spodoptera litura F. (Lepidoptera: Noctuidae) VI. Possible overwintering areas predicted from meteorological data in Japan. Appl. Entomol. Zool. 1997, 32, 167–177. [Google Scholar] [CrossRef][Green Version]
- Sahayara, K.; Gabriel, P.M. Screening the relative toxicity of some plant extracts to Spodoptera litura fab. (Insecta: Lepidoptera: Noctuidae) of groundnut. Fresenius Environ. Bull. 1998, 7, 557–560. [Google Scholar]
- Ahmad, M.; Arif, M.I.; Ahmad, M. Occurrence of insecticide resistance in field populations of Spodoptera litura (Lepidoptera: Noctuidae) in Pakistan. Crop Prot. 2007, 26, 809–817. [Google Scholar] [CrossRef]
- Ahmad, M.; Mehmood, R. Monitoring of resistance to new chemistry insecticides in Spodoptera litura (Lepidoptera, Noctuidae) in Pakistan. J. Econ. Entomol. 2015, 108, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Ishtiaq, M.; Saleem, M.A.; Razaq, M. Monitoring of resistance in Spodoptera exigua (Lepidoptera: Noctuidae) from four districts of the Southern Punjab, Pakistan to four conventional and six new chemistry insecticides. Crop Prot. 2012, 33, 13–20. [Google Scholar] [CrossRef]
- Driver, F.; Milner, R.J.; Trueman, J.W.H. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycol. Res. 2000, 104, 134–150. [Google Scholar] [CrossRef]
- Sundh, I.; Goettel, M.S. Regulating biocontrol agents: A historical perspective and a critical examination comparing microbial and macrobial agents. Biocontrol 2013, 58, 575–593. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Sharma, S.; Raliya, R. Nanotechnology: Interdisciplinary science of applications. Afr. J. Biotechnol. 2013, 12, 219–226. [Google Scholar]
- Goswami, A.; Roy, I.; Sengupta, S.; Debnath, N. Novel applications of solid and liquid formulations of nanoparticles against insect pests and pathogens. Thin Solid Films 2010, 519, 1252–1257. [Google Scholar] [CrossRef]
- Banu, A.N.; Balasubramanian, C. Myco-synthesis of silver nanoparticles using Beauveria bassiana against dengue vector, Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2014, 113, 2869–2877. [Google Scholar] [CrossRef]
- Banu, A.N.; Balasubramanian, C. Optimization and synthesis of silver nanoparticles using Isaria fumosorosea against human vector mosquitoes. Parasitol. Res. 2014, 113, 3843–3851. [Google Scholar] [CrossRef] [PubMed]
- Amerasan, D.; Nataraj, T.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Nicoletti, M.; Benelli, G. Myco-synthesis of silver nanoparticles using Metarhizium anisopliae against the rural malaria vector Anopheles culicifacies Giles (Diptera: Culicidae). J. Pest Sci. 2016, 89, 249–256. [Google Scholar] [CrossRef]
- Wang, X.S.; Xu, J.; Wang, X.M.; Qiu, B.L.; Cuthbertson, A.G.S.; Du, C.L.; Wu, J.H.; Ali, S. Isaria fumosorosea-based-zero-valent iron nanoparticles affect the growth and survival of sweet potato whitefly, Bemisia tabaci (Gennadius). Pest Manag. Sci. 2019, 75, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- David, W.; Ellaby, S.; Taylor, G. Rearing Spodoptera exempta on semi synthetic diets and on growing maize. Entomol. Exp. Appl. 1975, 19, 226–236. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Media composition influences on growth, enzyme activity and virulence of the entomopathogen hyphomycete Isaria fumosorosea. Entomol. Exp. Appl. 2009, 131, 30–38. [Google Scholar] [CrossRef]
- Ali, S.; Zhang, C.; Wang, Z.; Wang, X.M.; Wu, J.H.; Cuthbertson, A.G.; Shao, Z.; Qiu, B.L. Toxicological and biochemical basis of synergism between the entomopathogenic fungus Lecanicillium muscarium and the insecticide matrine against Bemisia tabaci (Gennadius). Sci. Rep. 2017, 7, 46558. [Google Scholar] [CrossRef]
- Wu, J.; Yu, X.; Wang, X.; Tang, L.; Ali, S. Matrine enhances the pathogenicity of Beauveria brongniartii against Spodoptera litura (Lepidoptera: Noctuidae). Front. Microbiol. 2019, 10, 1812. [Google Scholar] [CrossRef]
- Koul, O.; Shankar, J.S.; Mehta, N.; Taneja, S.C.; Tripathi, A.K.; Dhar, K.L. Bio-efficacy of crude extracts of Aglaia species (Meliaceae) and some active fractions against lepidopteran larvae. J. Appl. Entomol. 1997, 121, 245–248. [Google Scholar] [CrossRef]
- Rizwan, M.H.; Hu, Q.B.; Hu, M.Y.; Zhong, G.H.; Weng, Q.F. Study of destruxin B and tea saponin, their interaction and synergism activities with Bacillus thuringiensis kurstaki against Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2009, 44, 419–428. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J. Pest Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein—Dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase-improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [PubMed]
- Simon, L.M.; Fatrai, Z.; Jonas, D.E.; Matkovics, B. Study of metabolism enzymes during the development of Phaseolus vulgaris L. Biochem. Physiol. Pflanz. 1974, 166, 387–392. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione-S-transferase. Methods Enzymol. 1981, 77, 398–405. [Google Scholar]
- SAS Institute. SAS User’s Guide; Statistics SAS Institute: Cary, NC, USA, 2000. [Google Scholar]
- Sabbour, M.M.; El-Aziz, S.E.A. Efficacy of some nano-imidacloprid against red flour beetle Tribolium castaneum and confused flour beetle, Tribolium confusum (Coleoptera:Tenebrionidae) under laboratory and store conditions. Bull. Environ. Pharmacol. Life Sci. 2015, 4, 54–59. [Google Scholar]
- Rai, M.; Ingle, A. Role of nanotechnology in agriculture with special reference to management of insect pests. Appl. Microbiol. Biotechnol. 2012, 94, 287–293. [Google Scholar] [CrossRef]
- Gajbhiye, M.; Kesharwani, J.; Ingle, A.; Gade, A.; Rai, M. Fungus mediated synthesis of silver nanoparticles and its activity against pathogenic fungi in combination of fluconazole. Nanomedicine 2009, 5, 282–286. [Google Scholar]
- Mukherjee, P.; Ahmad, A.; Mandal, D.; Senapati, S.; Sainkar, S.R.; Khan, M.I.; Ramani, R.; Parischa, R.; Ajayakumar, P.V.; Alam, M.; et al. Bioreduction of AuCl4-ions by the fungus, Verticillium sp. and surface trapping of the gold nanoparticles formed. Angew. Chem. Int. Ed. 2001, 40, 3585–3588. [Google Scholar] [CrossRef]
- Vigneshwaran, N.; Ashtaputrea, N.M.; Varadarajana, P.V.; Nachanea, R.P.; Paralikara, K.M.; Balasubramanyaa, R.H. Biological synthesis of silver nanoparticles using the fungus Aspergillus Flavus. Mater. Lett. 2007, 61, 1413–1418. [Google Scholar] [CrossRef]
- Lengke, F.M.; Fleet, E.M.; Southam, G. Biosynthesis of silver nanoparticles by filamentous cyanobacteria from a silver (I) nitrate complex. Langmuir 2007, 23, 2694–2699. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.H.; Raj, L.F.A.A.; Namasivayam, S.K.R.; Bharani, R.S.A. Improved pesticidal activity of fungal metabolite from Nomuraea rileyi with chitosan nanoparticles. In Book of Abstracts International Conference on Advanced Nanomaterials and Emerging Engineering Technologies (ICANMEET) Chennai, India; IEEE: New York, NY, USA, 2013; pp. 387–390. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Production and regulation of extracellular proteases from the entomopathogenic fungus Isaria fumosoroseus (Cordycipitaceae; Hypocreales) in the presence of diamondback moth, Plutella xylostella cuticle. Biocontrol Sci. Technol. 2009, 19, 523–535. [Google Scholar] [CrossRef]
- Ali, S.; Wu, J.H.; Huang, Z.; Ren, S.X. Production and regulation of extracellular chitinase from the entomopathogenic fungus Isaria fumosorosea. Biocontrol Sci. Technol. 2010, 20, 723–738. [Google Scholar] [CrossRef]
- Jamei, M.R.; Khosravi, M.R.; Anvaripour, B. Soil remediation using nano zero-valent iron synthesized by an ultrasonic method. Iran. J. Oil Gas Sci. Technol. 2012, 1, 1–12. [Google Scholar]
- Brandt, C.R.; Adang, M.J.; Spence, K.D. The peritrophic membrane: Ultrastructural analysis and function as a mechanical barrier to microbial infection in Orgyia pseudotsugata. J. Invertebr. Pathol. 1978, 32, 12–24. [Google Scholar] [CrossRef]
- Shahabuddin, M.; Toyoshima, T.; Aikawa, M.; Kaslow, D.C. Transmission-blocking activity of a chitinase inhibitor and activation of malarial parasite chitinase by mosquito protease. Proc. Natl. Acad. Sci. USA 1993, 90, 4266–4270. [Google Scholar] [CrossRef]
- Chang, M.C.; Shu, H.Y.; Hsieh, W.P.; Wang, M.C. Using nanoscale zero-valent iron for the remediation of polycyclic aromatic hydrocarbons contaminated soil. J. Air Waste Manag. Assoc. 2012, 55, 1200–1207. [Google Scholar] [CrossRef]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef]
- Tian, J.; Diao, H.L.; Liang, L.; Hao, C.; Arthurs, S.; Ma, R.Y. Pathogenicity of Isaria fumosorosea to Bemisia tabaci, with some observations on the fungal infection process and host immune response. J. Invertebr. Pathol. 2015, 130, 147–153. [Google Scholar] [CrossRef]
- Jia, M.; Cao, G.C.; Li, Y.B.; Tu, X.B.; Wang, G.J.; Nong, X.Q.; Whitman, D.W.; Zhang, Z.H. Biochemical basis of synergism between pathogenic fungus Metarhizium anisopliae and insecticide chlorantraniliprole in Locusta migratoria (Meyen). Sci. Rep. 2016, 6, 28424. [Google Scholar] [CrossRef]
- Wu, J.J.; Li, J.Y.; Zhang, C.; Yu, X.T.; Cuthbertson, A.G.S.; Ali, S. Biological impact and enzyme activities of Spodoptera litura (Lepidoptera: Noctuidae) in response to synergistic action of Matrine and Beauveria brongniartii. Front. Physiol. 2020, 1, 584405. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Treatment Description | Concentration |
|---|---|---|
| T1 | B. brongniartii Fe0NPs | 6.25 ppm of diet |
| T2 | B. brongniartii Fe0NPs | 12.5 ppm of diet |
| T3 | B. brongniartii Fe0NPs | 25 ppm of diet |
| T4 | B. brongniartii Fe0NPs | 50 ppm of diet |
| T5 | B. brongniartii Fe0NPs | 100 ppm of diett |
| T6 | B. brongniartii Fe0NPs | 200 ppm of diet |
| T7 | B. brongniartii Fe0NPs | 500 ppm of diet |
| T8 | B. brongniartii conidial suspension | 108 conidia/g of diet |
| T9 | Control (ddH2O) | 0 |
| Treatment Time (Days) | LC50 (ppm) | 95% Fiducial Limit | Slope ± S.E | χ2 (df = 4) | p |
|---|---|---|---|---|---|
| 7 | 58 | 36–95 | 1.63 ± 0.13 | 3.92 | 0.870 |
| Concentration (ppm) | LT50 (Days) | 95% Fiducial Limit | Slope | χ2 (df = 4) | p |
|---|---|---|---|---|---|
| 200 | 5.10 | 3.72–6.99 | 2.32 ± 0.24 | 4.77 | 0.869 |
| 500 | 2.29 | 1.72–3.06 | 2.63 ± 0.19 | 3.92 | 0.964 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhang, K.; Cuthbertson, A.G.S.; Du, C.; Ali, S. Toxicity and Biological Effects of Beauveria brongniartii Fe0 Nanoparticles against Spodoptera litura (Fabricius). Insects 2020, 11, 895. https://doi.org/10.3390/insects11120895
Xu J, Zhang K, Cuthbertson AGS, Du C, Ali S. Toxicity and Biological Effects of Beauveria brongniartii Fe0 Nanoparticles against Spodoptera litura (Fabricius). Insects. 2020; 11(12):895. https://doi.org/10.3390/insects11120895
Chicago/Turabian StyleXu, Jing, Kaihui Zhang, Andrew G. S. Cuthbertson, Cailian Du, and Shaukat Ali. 2020. "Toxicity and Biological Effects of Beauveria brongniartii Fe0 Nanoparticles against Spodoptera litura (Fabricius)" Insects 11, no. 12: 895. https://doi.org/10.3390/insects11120895
APA StyleXu, J., Zhang, K., Cuthbertson, A. G. S., Du, C., & Ali, S. (2020). Toxicity and Biological Effects of Beauveria brongniartii Fe0 Nanoparticles against Spodoptera litura (Fabricius). Insects, 11(12), 895. https://doi.org/10.3390/insects11120895