Side Effects of Sulfur Dust on the European Grapevine Moth Lobesia botrana and the Predatory Mite Kampimodromus aberrans in Vineyards

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Bioassays
2.1.1. Lobesia botrana Used in Bioassays
2.1.2. Application of Sulfur Dust on Grapevine Berry Surfaces
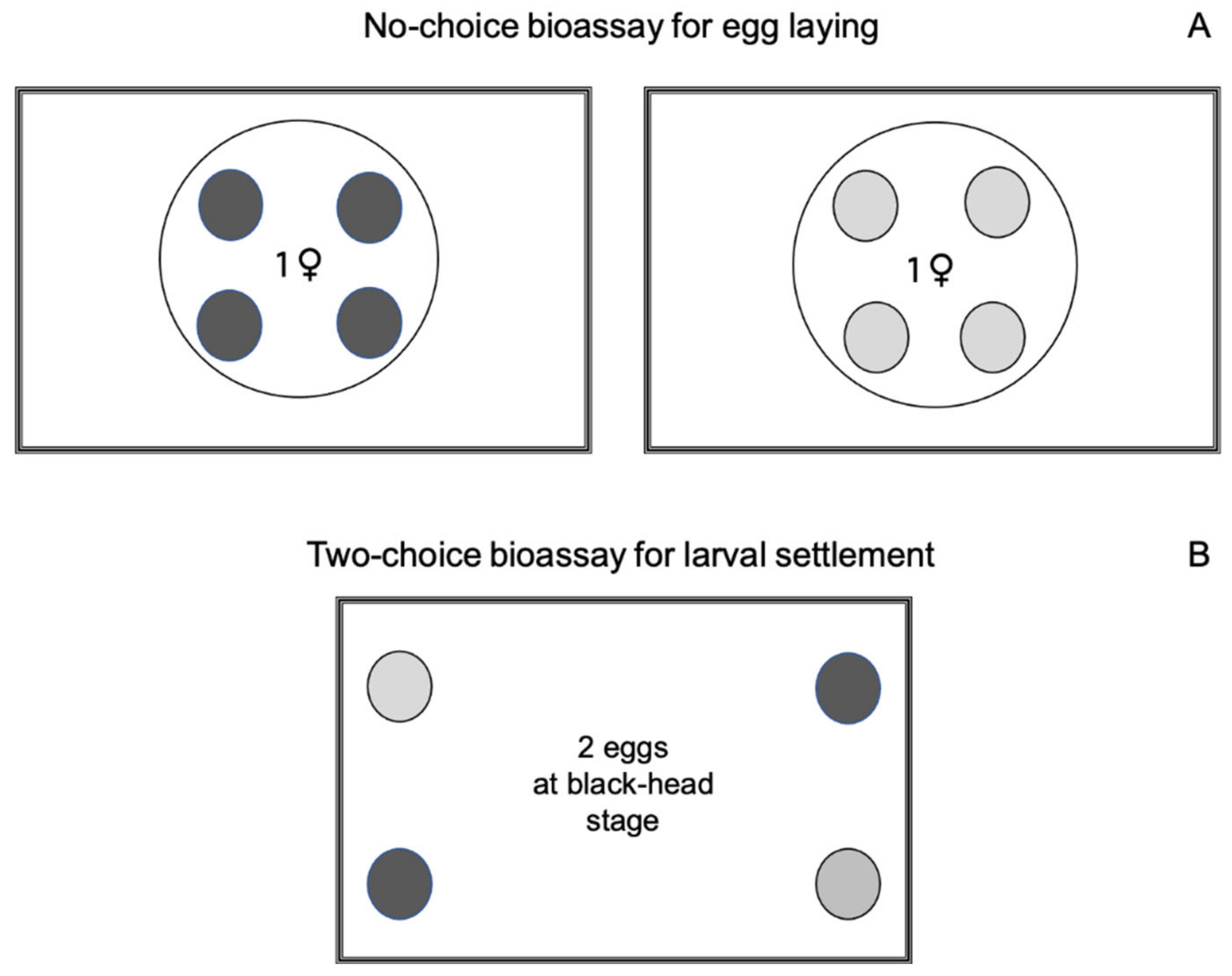
2.1.3. Influence of Sulfur Dust on Female Egg Laying
2.1.4. Influence of Sulfur Dust on Egg Hatching
2.1.5. Influence of Sulfur Dust on Larval Settlement Preference
2.2. Field Bioassay on Egg-Laying Preference
2.3. Field Trials
2.3.1. Efficacy against Lobesia botrana
2.3.2. Side Effects on Phytoseiid Mites
2.4. Statistical Analyses
3. Results
3.1. Laboratory Bioassays on Lobesia botrana
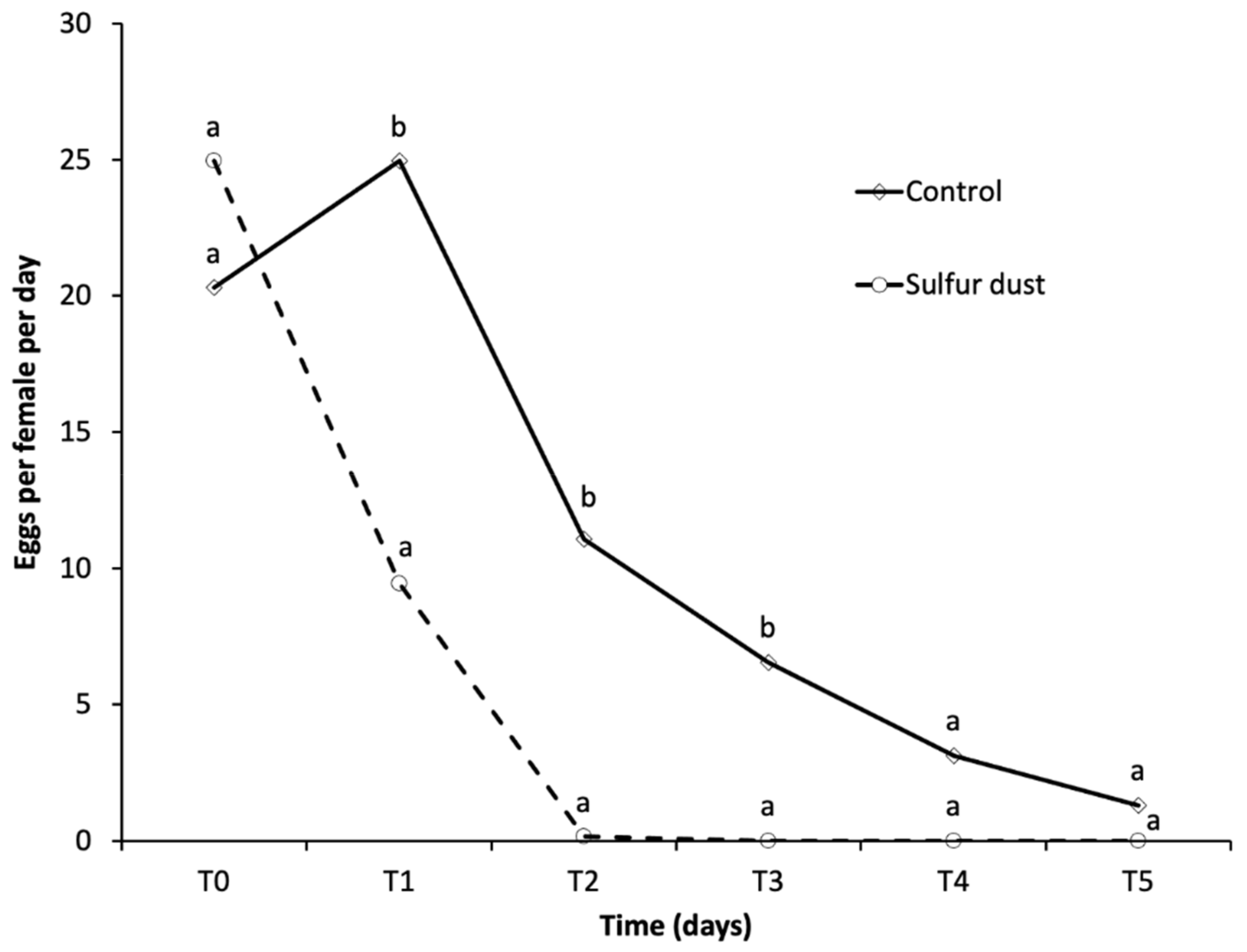
3.1.1. Influence of Sulfur Dust on Female Egg Laying
3.1.2. Influence of Sulfur Dust on Egg Hatching
3.1.3. Influence of Sulfur Dust on Larval Settlement Preference
3.2. Field Bioassay on Egg-Laying Preference of Lobesia botrana
3.3. Field Trials
3.3.1. Efficacy against the Second Generation of Lobesia botrana
3.3.2. Efficacy against the Third Generation of Lobesia botrana
3.3.3. Side Effects on Kampimodromus aberrans
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilligan, T.M.; Epstein, M.E.; Passoa, S.C.; Powell, J.A.; Sage, O.C.; Brown, J.W. Discovery of Lobesia botrana ([Denis & Schiffermuller]) in California: An invasive species new to North America (Lepidoptera: Tortricidae). Proc. Entomol. Soc. Wash. 2011, 113, 14–30. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A.; Varela, L.G. Grape berry moths in Western European vineyards and their recent movement into the New World. In Arthropod Management in Vineyards: Pest, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaac, R., Eds.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 339–360. [Google Scholar] [CrossRef]
- Pavan, F.; Floreani, C.; Barro, P.; Zandigiacomo, P.; Dalla Montà, L. Occurrence of two different development patterns in Lobesia botrana (Lepidoptera: Tortricidae) larvae during the second generation. Agric. For. Entomol. 2013, 14, 398–406. [Google Scholar] [CrossRef]
- Fermaud, M.; Giboulot, A. Influence of Lobesia botrana larvae on field severity of Botrytis rot of grape berries. Plant Dis. 1992, 76, 404–409. [Google Scholar] [CrossRef]
- Pavan, F.; Girolami, V.; Sacilotto, G. Second generation of grape berry moths, Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) and Eupoecilia ambiguella (Hb.) (Lep., Cochylidae): Spatial and frequency distributions of larvae, weight loss and economic injury level. J. Appl. Entomol. 1998, 122, 361–368. [Google Scholar] [CrossRef]
- Moschos, T. Yield loss quantification and economic injury level estimation for the carpophagous generations of the European grapevine moth Lobesia botrana Den. et Schiff. (Lepidoptera: Tortricidae). Int. J. Pest. Manag. 2006, 52, 141–147. [Google Scholar] [CrossRef]
- Pavan, F.; Bigot, G.; Cargnus, E.; Zandigiacomo, P. Influence of the carpophagous generations of the European grapevine moth Lobesia botrana on grape bunch rots. Phytoparasitica 2014, 42, 61–69. [Google Scholar] [CrossRef]
- Civolani, S.; Boselli, M.; Butturini, A.; Chicca, M.; Fano, E.A.; Cassinelli, S. Assessment of insecticide resistance of Lobesia botrana (Lepidoptera: Tortricidae) in Emilia-Romagna region. J. Econ. Entomol. 2014, 107, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Benelli, G. Towards pesticide-free farming? Sharing needs and knowledge promotes Integrated Pest Management. Environ. Sci. Pollut. Res. 2018, 25, 13439–13445. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A. Semiochemical strategies for tortricid moth control in apple orchards and vineyards in Italy. J. Chem. Ecol. 2016, 42, 571–583. [Google Scholar] [CrossRef]
- Lucchi, A.; Ladurner, E.; Iodice, A.; Savino, F.; Ricciardi, R.; Cosci, F.; Conte, G.; Benelli, G. Eco-friendly pheromone dispensers—A green route to manage the European grapevine moth? Environ. Sci. Pollut. Res. 2018, 25, 9426–9442. [Google Scholar] [CrossRef]
- Ifoulis, A.A.; Savopoulou-Soultani, M. Biological control of Lobesia botrana (Lepidoptera: Tortricidae) larvae by using different formulations of Bacillus thuringiensis in 11 vine cultivars under filed conditions. J. Econ. Entomol. 2004, 97, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Boselli, M.; Scannavini, M.; Melandri, M. Confronto fra strategie di difesa contro la tignoletta della vite. Inf. Agrar. 2000, 56, 61–65. (In Italian) [Google Scholar]
- Fermaud, M. Cultivar susceptibility of grape berry clusters to larvae of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 1998, 91, 974–980. [Google Scholar] [CrossRef]
- Moreau, J.; Benrey, B.; Thiéry, D. Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2006, 96, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Sharon, R.; Zahavi, T.; Soroker, V.; Harari, A.R. The effect of grapevine cultivars on Lobesia botrana (Lepidoptera: Tortricidae) population levels. J. Pest. Sci. 2009, 82, 187–193. [Google Scholar] [CrossRef]
- Pavan, F.; Stefanelli, G.; Villani, A.; Cargnus, E. Influence of grapevine cultivar on the second generations of Lobesia botrana and Eupoecilia ambiguella. Insects 2018, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Rid, M.; Markheiser, A.; Hoffmann, C.; Gross, J. Waxy bloom on grape berry surface is one important factor for oviposition of European grapevine moths. J. Pest. Sci. 2018, 91, 1225–1239. [Google Scholar] [CrossRef]
- Vartholomaiou, A.N.; Navrozidis, E.I.; Payne, C.C.; Salpiggidis, G.A. Agronomic techniques to control Lobesia botrana. Phytoparasitica 2008, 36, 264–271. [Google Scholar] [CrossRef]
- Pavan, F.; Cargnus, E.; Kiaeianmoosavi, S.; Bigot, G.; Tacoli, F.; Zandigiacomo, P. Bunch-zone leaf removal of grapevines to prevent damage by Lobesia botrana and grey mould. Bull. Insectol. 2016, 69, 107–115. [Google Scholar]
- Kiaeian Moosavi, F.; Cargnus, E.; Pavan, F.; Zandigiacomo, P. Effects of grapevine bunch exposure to sunlight on berry surface temperature and Lobesia botrana (Lepidoptera: Tortricidae) egg laying, hatching and larval settlement. Agric. For. Entomol. 2018, 20, 420–432. [Google Scholar] [CrossRef]
- Tacoli, F.; Cargnus, E.; Kiaeian Moosavi, F.; Zandigiacomo, P.; Pavan, F. Efficacy and mode of action of kaolin and its interaction with bunch-zone leaf removal against Lobesia botrana on grapevines. J. Pest. Sci. 2019, 92, 465–475. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Kreiter, S.; Tixier, M.-S.; Candolfi, M.P. Management of phytophagous mites in European vineyards. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 191–217. [Google Scholar] [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourasso, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef]
- Tixier, M.S.; Baldassar, A.; Duso, C.; Kreiter, S. Phytoseiidae in European grape (Vitis vinifera L.): Bio-ecological aspects and keys to species (Acari: Mesostigmata). Zootaxa 2013, 3721, 101–142. [Google Scholar] [CrossRef] [PubMed]
- Pozzebon, A.; Borgo, M.; Duso, C. The effects of fungicides on non-target mites can be mediated by plant pathogens. Chemosphere 2010, 79, 8–17. [Google Scholar] [CrossRef]
- Pozzebon, A.; Tirello, P.; Moret, R.; Pederiva, M.; Duso, C. A fundamental step in IPM on grapevine: Evaluating the side effects of pesticides on predatory mites. Insects 2015, 6, 847–857. [Google Scholar] [CrossRef]
- Tirello, P.; Pozzebon, A.; Duso, C. The effect of insecticides on the non-target predatory mite Kampimodromus aberrans: Laboratory studies. Chemosphere 2013, 93, 1139–1144. [Google Scholar] [CrossRef]
- Tixier, M.-S. Predatory mites (Acari: Phytoseiidae) in agro-ecosystems and conservation biological control: A review and explorative approach for forecasting plant-predatory mite interactions and mite dispersal. Front. Ecol. Evol. 2018, 6, 192. [Google Scholar] [CrossRef]
- Bencivelli, A.; Rapparini, G. The activity of chemicals of known and of recent formulation against grapevine powdery mildew. Inf. Fitopatol. 1975, 25, 19–21. [Google Scholar]
- Touzeau, J. Les actions secondaires du soufre sur les maladies et ravageurs de la vigne. In Proceedings of the International Symposium Elemental Sulphur in Agriculture, Acropolis, Nice, France, 25–27 March 1987; pp. 167–177. [Google Scholar]
- Michelatti, G.; Pinoggi, G.; Schreiber, G.; Mozzone, G.C. Esperienze di lotta razionale contro l’eriofide del nocciolo (Phytocoptella avellanae Nal.) condotte nell’arco di un quinquennio in Piemonte. Acta Hortic. 1994, 351, 575–581. [Google Scholar] [CrossRef]
- Goebel, O.; Vergnet, C.; Heller, J.J. Lutte contre l’érinose et l’acariose de la vigne: Intérêt des traitements de début de saison au soufre. Phytoma 2001, 537, 39–41. (In French) [Google Scholar]
- Ahmed, M.; Hoque, A.K.M.R.; Mamun, M.S.A. Efficacy of different sulphur formulations against Red Spider Mite (Oligonychus coffeae Nietner) of tea in Bangladesh. Int. J. Sust. Agric. Technol. 2011, 7, 29–33. [Google Scholar]
- Haddadi, A.; Mirfakhraei, S.; Aramideh, S. Effects of abamectin, volk oil, detergent and sulfur in control of grape erineum mite, Colomerus vitis Pagenstecher (Acari: Eriophyidae) in vineyards of West-Azerbaijan Province, Iran. Ann. Biol. 2019, 35, 94–104. [Google Scholar]
- Jepsen, S.J.; Rosenheim, J.A.; Bench, M.E. The effect of sulfur on biological control of the grape leafhopper, Erythroneura elegantula, by the egg parasitoid Anagrus erythroneurae. BioControl 2007, 52, 721–732. [Google Scholar] [CrossRef]
- Cerutti, F.; Roux, O.; Delucchi, V. L’énigme de la nuisibilité de la cicadelle de la vigne au Tessin. Mitt. Schweiz. Entomol. Ges. 1989, 62, 247–252. (In French) [Google Scholar]
- Schwartz, A. Occurrence of natural enemies of phytophagous mites on grapevine leaves following application of fungicides for disease control. S. Afr. J. Enol. Vitic. 1993, 14, 16–17. [Google Scholar] [CrossRef]
- Hanna, R.; Zalom, F.G.; Wilson, L.T.; Leavitt, G.M. Sulfur can suppress mite predators in vineyards. Calif. Agric. 1997, 51, 19–21. [Google Scholar] [CrossRef]
- Hassan, S.A.; Bigler, F.; Bogenschütz, H.; Boller, E.; Brun, J.; Calis, J.N.M.; Coremans-Pelseneer, J.; Duso, C.; Grove, A.; Heimbach, U.; et al. Results of the sixth joint pesticide testing programme of the IOBC/WPRS-Working Group «Pesticides and Beneficial Organisms». Entomophaga 1994, 39, 107–119. [Google Scholar] [CrossRef]
- Bernard, M.B.; Cole, P.; Kobelt, A.; Horne, P.A.; Altmann, J.; Wratten, S.D.; Yen, A.L. Reducing the impact of pesticides on biological control in Australian vineyards: Pesticide mortality and fecundity effects on an indicator species, the predatory mite Euseius victoriensis (Acari: Phytoseiidae). J. Econ. Entomol. 2010, 103, 2061–2071. [Google Scholar] [CrossRef]
- Gadino, A.N.; Walton, V.M.; Dreves, A.J. Impact of vineyard pesticides on a beneficial arthropod, Typhlodromus pyri (Acari: Phytoseiidae), in laboratory bioassays. J. Econ. Entomol. 2011, 104, 970–977. [Google Scholar] [CrossRef]
- Zappalà, L.; Siscaro, G.; Biondi, A.; Mollá, O.; González-Cabera, J.; Urbaneja, A. Efficacy of sulphur on Tuta absoluta and its side effects on the predator Nesidiocoris tenuis. J. Appl. Entomol. 2012, 136, 401–409. [Google Scholar] [CrossRef]
- Zakher, A.G.; Abdel-Aziz, M.A.; Afsah, A.F.E.; Farha, H.F. Response of tomato plants to some agricultural and chemical treatments on fruit yield and its quality relation to Tuta absoluta (Meyrick) and Bemisia tabaci (Genn.) infestation. Arab Univ. J. Agric. Sci. 2016, 24, 609–622. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth stages of the grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp vinifera). Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Rapagnani, M.R.; Caffarelli, V.; Barlattani, M.; Minelli, F. Descrizione di un allevamento, in laboratorio, della tignoletta dell’uva Lobesia botrana Den. e Schiff. (Lepidoptera—Tortricidae) su un nuovo alimento semi-sintetico. Boll. Ist. Ent. G. Grandi Univ. Bologna 1990, 44, 57–64. (In Italian) [Google Scholar]
- Maher, N.; Thiéry, D. A bioassay to evaluate the activity of chemical stimuli from grape berries on the oviposition of Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2004, 94, 27–33. [Google Scholar] [CrossRef]
- Tacoli, F.; Pavan, F.; Cargnus, E.; Tilatti, E.; Pozzebon, A.; Zandigiacomo, P. Efficacy and mode of action of kaolin in the control of Empoasca vitis and Zygina rhamni (Hemiptera: Cicadellidae) in vineyards. J. Econ. Entomol. 2017, 110, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Hoy, M.A.; Standow, K.A. Resistance to sulfur in a vineyard spider mite predator. Calif. Agric. 1981, 35, 8–10. [Google Scholar]
- Papaioannou-Souliotis, P.; Markoyiannaki-Printziou, D.; Tsagkarakou, A.; Rumbos, I.; Adamopoulos, I. Effects of different fungicides and insecticides on populations of Phytoseius finitimus (Ribaga) in vineyard in four regions of Greece. Redia 1998, 81, 17–35. [Google Scholar]
- Rumbos, I.C.; Papaioannou-Souliotis, P.; Markoyiannaki-Printziou, D.; Adamopoulos, I.C. Promotion of integrated pest control system in viticulture in Greece with respect to predatory mites. IOBC WPRS Bull. 2000, 23, 125–126. [Google Scholar]
- Costello, M.J. Impact of sulfur on density of Tetranychus pacificus (Acari: Tetranychidae) and Galendromus occidentalis (Acari: Phytoseiidae) in a central California vineyard. Exp. Appl. Acarol. 2007, 42, 197–208. [Google Scholar] [CrossRef]
- Gázquez, J.C.; López, J.C.; Baeza, E.J.; Pérez-Parra, J.J.; Pérez, C.; Meca, D.E.; Navarro, S. Influence of the sulphur application method on pests, diseases and natural enemies in a greenhouse pepper crop. Acta Hortic. 2011, 893, 1309–1316. [Google Scholar] [CrossRef]
- Tacoli, F.; Cargnus, E.; Pozzebon, A.; Duso, C.; Tirello, P.; Pavan, F. Side effects of kaolin and bunch-zone leaf removal on predatory mite population (Acari: Phytoseiidae) occurring in vineyards. J. Econ. Entomol. 2019, 112, 1292–1298. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source of Variation | Levels | Mean ± SE | F | Degrees of Freedom | p |
|---|---|---|---|---|---|
| Trial | 2017 | 8.83 ± 2.64 a | 27.59 | 2, 24 | <0.0001 |
| 2018 | 30.50 ± 7.56 b | ||||
| 2020 | 5.75 ± 1.48 a | ||||
| Treatment | Control | 24.67 ± 6.38 c | 16.39 | 2, 24 | <0.0001 |
| Sulfur dust | 15.25 ± 6.75 b | ||||
| Bacillus thuringiensis | 7.17 ± 2.01 a | ||||
| Replicate | 1.61 | 3, 24 | 0.21 | ||
| Trial × treatment | 0.99 | 4, 24 | 0.43 |
| Source of Variation | Levels | Mean ± SE | F | Degrees of Freedom | p |
|---|---|---|---|---|---|
| Trial | 2017 | 7.00 ± 1.41 | 242.52 | 1, 21 | <0.0001 |
| 2018 | 35.50 ± 3.73 | ||||
| Treatment | Control | 35.25 ± 8.17 c | 22.76 | 3, 21 | <0.0001 |
| Sulfur dust | 21.75 ± 6.45 b | ||||
| Kaolin | 15.25 ± 4.16 ab | ||||
| Bacillus thuringiensis | 12.75 ± 3.65 a | ||||
| Replicate | 0.69 | 3, 21 | 0.57 | ||
| Trial × treatment | 1.90 | 3, 21 | 0.16 |
| Source of Variation | Levels | Mean ± SE | F | Degrees of Freedom | p |
|---|---|---|---|---|---|
| Time | 10 June | 9.46 ± 1.42 a | 19.89 | 2, 18 | <0.0001 |
| 22 June | 14.66 ± 1.82 b | ||||
| 29 June | 14.14 ± 1.96 b | ||||
| Treatment | Control | 13.09 ± 1.80 | 0.18 | 2, 9 | 0.84 |
| Sulfur dust | 12.82 ± 1.80 | ||||
| Bacillus thuringiensis | 12.35 ± 2.00 | ||||
| Time × Treatment | 1.37 | 4, 18 | 0.282 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tacoli, F.; Cargnus, E.; Zandigiacomo, P.; Pavan, F. Side Effects of Sulfur Dust on the European Grapevine Moth Lobesia botrana and the Predatory Mite Kampimodromus aberrans in Vineyards. Insects 2020, 11, 825. https://doi.org/10.3390/insects11110825
Tacoli F, Cargnus E, Zandigiacomo P, Pavan F. Side Effects of Sulfur Dust on the European Grapevine Moth Lobesia botrana and the Predatory Mite Kampimodromus aberrans in Vineyards. Insects. 2020; 11(11):825. https://doi.org/10.3390/insects11110825
Chicago/Turabian StyleTacoli, Federico, Elena Cargnus, Pietro Zandigiacomo, and Francesco Pavan. 2020. "Side Effects of Sulfur Dust on the European Grapevine Moth Lobesia botrana and the Predatory Mite Kampimodromus aberrans in Vineyards" Insects 11, no. 11: 825. https://doi.org/10.3390/insects11110825
APA StyleTacoli, F., Cargnus, E., Zandigiacomo, P., & Pavan, F. (2020). Side Effects of Sulfur Dust on the European Grapevine Moth Lobesia botrana and the Predatory Mite Kampimodromus aberrans in Vineyards. Insects, 11(11), 825. https://doi.org/10.3390/insects11110825