
Regulation of Genes Involved in Heterocyst Differentiation in the Cyanobacterium Anabaena sp. Strain PCC 7120 by a Group 2 Sigma Factor SigC

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Mutant Construction

{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5'- 3') |
|---|---|
| sigC-5F | TAGAGCTCGCGGACTCACAGAAATGGTT |
| sigC-5R | TAGGATCCAATGGCGATATCAGGGTCT |
| sigC-3F | TAGGATCCTCGCAACCTTATCCGTGACT |
| sigC-3R | ATCTCGAGTTTGGCAGTCCAGTAGGTGA |
| sigG-5F | AGGAGCTCACGTCCATGATCAAACCAA |
| sigG-5R | TAGGATCCGTGGTTCGAGAGTTTGTCA |
| sigG-3F | ACGGATCCTTGCCGAAATCACAGGTGTA |
| sigG-3R | ATCTCGAGGGCGTGGGTATATTTGATG |
| RTrrn16SF2 | GCAAGTCGAACGGTCTCTTC |
| RTrrn16SR2 | GGTATTAGCCACCGTTTCCA |
| RTsigA-F | TTGTTGCTCGCTGATGATGG |
| RTsigA-R | TTCTTGCTTTGTGTCCGACG |
| RTsigB2-F | ACACCCACACAGAAGACACA |
| RTsigB2-R | TCTTTAGCGTCAATCAGCGAC |
| RTsigC-F | ACCTGGAGCCATAGAGACGA |
| RTsigC-R | CATCCACCGACAAATCACTG |
| RTsigD-F | AGCGTAGAAGAGTGGGCAAA |
| RTsigD-R | GGATACCACTAGCCGCAAGT |
| RTsigE-F | TGGCACGTTATCCACTGCTA |
| RTsigE-R | GTCGGATGTTGCCCTATTTG |
| RTsigF-F | TTGCGGGAACAATACAACCG |
| RTsigF-R | CCATCTTGCACGGGTACATC |
| RTsigG-F | TTGATGCGAGGTGTCCAGAA |
| RTsigG-R | GGTTTGATAGCGGCGCAATA |
| RTsigI-F | GATTCGGCGGCATTAAGTGT |
| RTsigI-R | GCTTCTTGGGAATCAGCCAG |
| RTsigJ-F | GGCAGCAAGTGAGTCCTCTA |
| RTsigJ-R | GCCGGTGTGTAATTGAACCA |
| RTfbp-F | CAACCTTATCCCGTCACGTC |
| RTfbp-R | GCGACGGGCAACTAATTTAC |
| RTtalB-F | GACCACCAATCCCTCTCTGA |
| RTtalB-R | AAGCGAGGGAGACAATTTGA |
| RTcoxB3-F | AAGGGCCGACAGCATTAGTA |
| RTcoxB3-R | ATACCCACGCCCATTGTTTA |
| RTcoxA2-F | CAACGCATTCATGACCAATC |
| RTcoxA2-R | AAGGTGGGTAAGCAGTCCAA |
| RThepA-F | CAGGAATTAGCTGGGTTGACA |
| RThepA-R | ATTGAAGGTAGCACGCATCC |
| RThepB-F | AAATTTATCGCGCCAACAAG |
| RThepB-R | CTCCGACACGATGCACTAAA |
| RTntcA-F | CAAGATAAGGCCCTAGCAAATG |
| RTntcA-R | TCCGACTTGTTTCCTGTCAAC |
| RT4160-F | TCATGACTAGCCAACCCACA |
| RT4160-R | TACTGCTTCCAGCACGCTTA |
2.3. DNA Microarray Analysis
2.4. Quantitative Reverse Transcription PCR (qRT-PCR)
2.5. Acetylene Reduction Assays
3. Results and Discussion
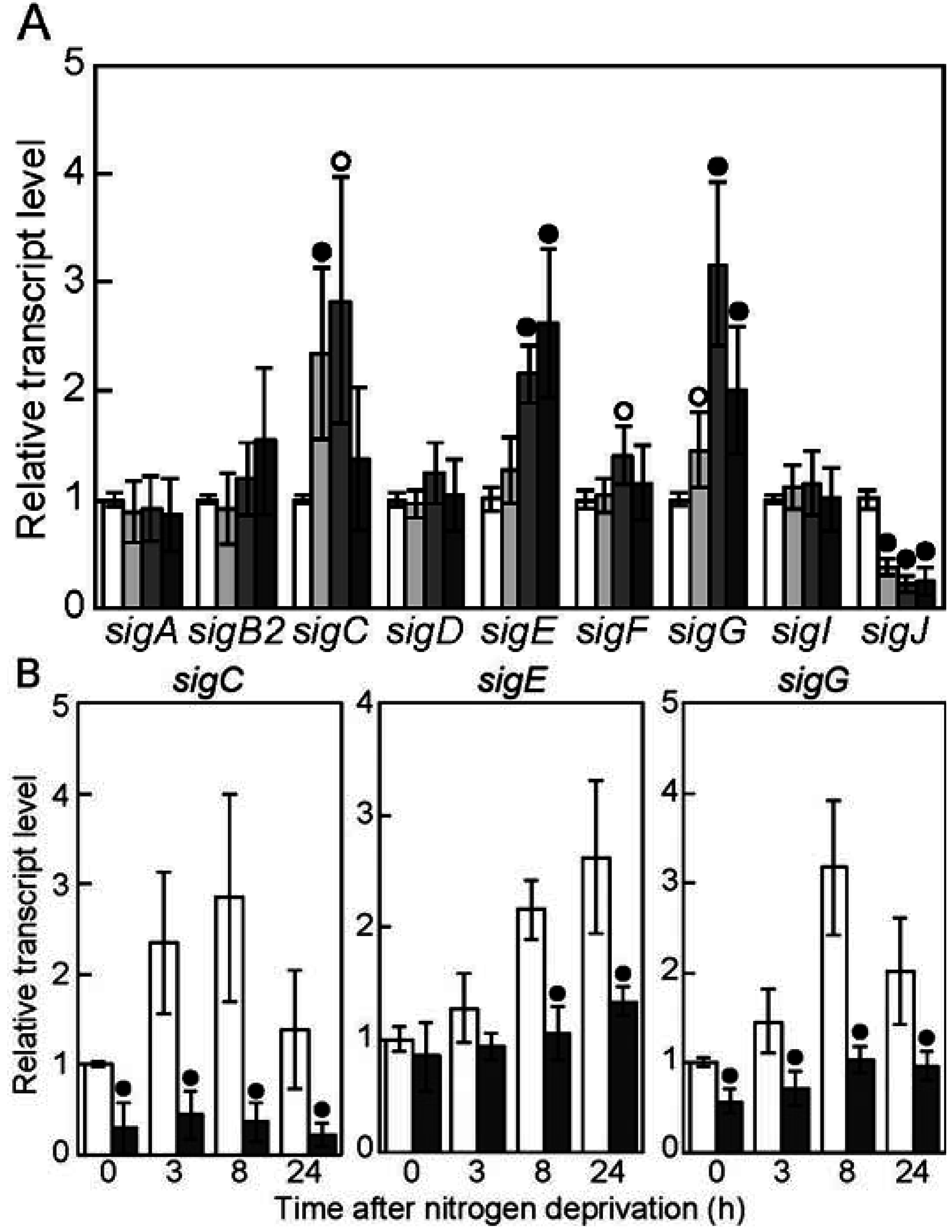
3.1. Nitrogen-Regulated Expression of Genes Encoding Sigma Factors
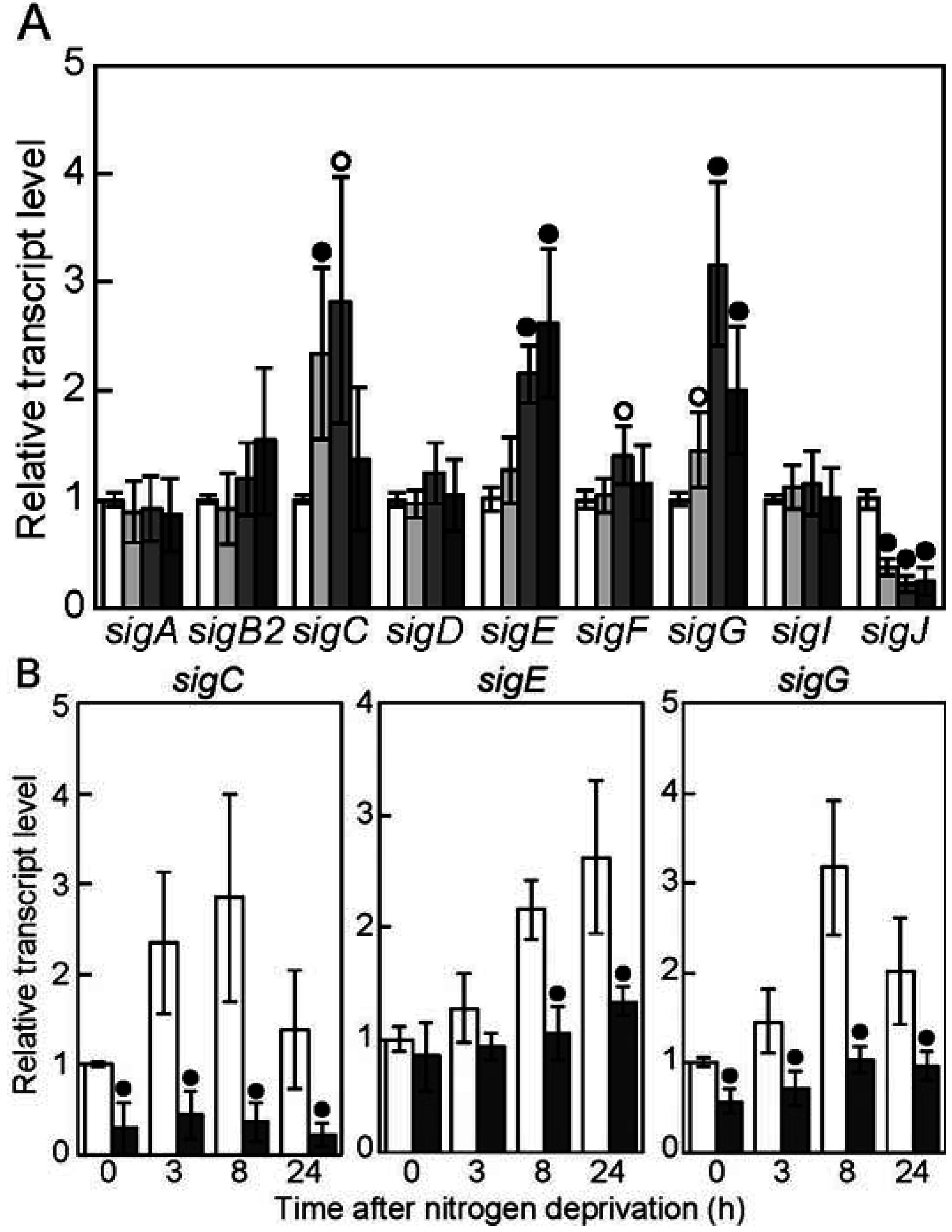
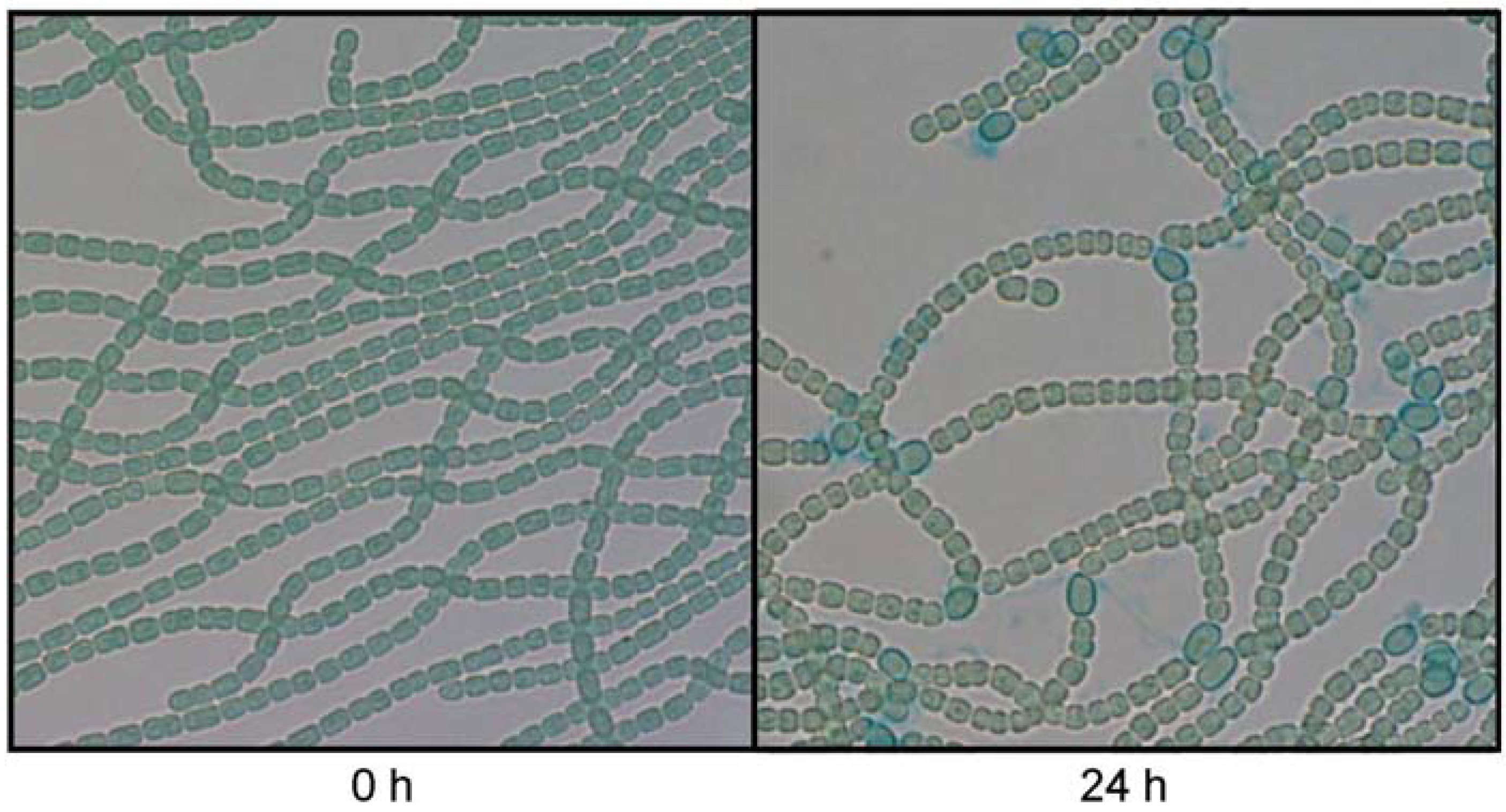
3.2. The sigC Gene Is Required for Normal Induction of Genes Involved in Heterocyst Differentiation
| ORF No. | Gene | Product | ΔsigC/WT a | p b |
|---|---|---|---|---|
| all0406 | - | Unknown protein | −1.16 | 3.6 × 10−2 |
| all0438 | - | Serine/threonine kinase with WD-40 repeat | −1.53 | 5.1 × 10−3 |
| asl0597 | - | Hypothetical protein | −1.01 | 1.4 × 10−2 |
| all0918 | - | Unknown protein | −1.19 | 4.5 × 10−2 |
| all0919 | - | Probable glycosyltransferase | −1.96 | 5.9 × 10−4 |
| all1101 | - | Ferrichrome iron receptor | −1.01 | 3.0 × 10−2 |
| alr1112 | - | Probable transglycosylase | −1.04 | 3.4 × 10−2 |
| alr1276 | - | Putative acetyl transferase | −1.14 | 2.7 × 10−3 |
| asr1405 | - | Hypothetical protein | −1.11 | 6.3 × 10−3 |
| asr1408 | nifZ | Iron-sulfur cofactor synthesis protein | −1.29 | 2.9 × 10−3 |
| all1424 | - | Unknown protein | −1.19 | 9.5 × 10−5 |
| asl1778 | - | Unknown protein | −1.54 | 2.9 × 10−5 |
| alr2323 | htpG | Heat shock protein | −1.04 | 4.3 × 10−2 |
| alr2405 | isiB | Flavodoxin | −1.50 | 8.7 × 10−3 |
| alr2514 | coxB2 | Cytochrome c oxidase subunit II | −1.37 | 2.2 × 10−2 |
| alr2515 | coxA2 | Cytochrome c oxidase subunit I | −1.62 | 1.4 × 10−3 |
| asr2523 | - | Unknown protein | −1.24 | 2.4 × 10−3 |
| all2563 | talB | Transaldolase | −1.13 | 1.9 × 10−2 |
| alr2582 | - | Hypothetical protein | −1.41 | 9.5 × 10−3 |
| all2637 | - | Unknown protein | −1.08 | 3.5 × 10−2 |
| all2655 | - | Unknown protein | −1.16 | 1.2 × 10−2 |
| alr2730 | - | Hypothetical protein | −1.12 | 1.3 × 10−2 |
| alr2731 | coxB3 | Cytochrome c oxidase subunit II | −1.11 | 2.0 × 10−3 |
| alr2818 | hetP | Heterocyst differentiation protein | −1.39 | 1.7 × 10−3 |
| alr2822 | - | Hypothetical protein | −1.48 | 3.9 × 10−4 |
| alr2823 | - | Hypothetical protein | −1.58 | 1.3 × 10−4 |
| alr2824 | - | Hypothetical protein | −2.24 | 4.9 × 10−4 |
| alr2825 | - | Glucose-1-P cytidylyltransferase | −1.38 | 2.1 × 10−4 |
| alr2826 | - | Hypothetical protein | −1.77 | 3.8 × 10−4 |
| alr2827 | - | Putative epimerase/dehydratase | −1.32 | 4.2 × 10−4 |
| alr2828 | - | Unknown protein | −1.92 | 7.3 × 10−4 |
| alr2829 | - | Unknown protein | −1.70 | 3.1 × 10−4 |
| alr2830 | rfbC | dTDP-4-dehydrorhamnose 3,5-epimerase | −1.40 | 3.1 × 10−4 |
| alr2831 | - | Probable NAD(P)-dependent oxidoreductase | −1.54 | 1.5 × 10−3 |
| alr2832 | - | Putative glycosyltransferase | −1.16 | 9.6 × 10−5 |
| alr2833 | - | Hypothetical protein | −1.44 | 3.7 × 10−3 |
| alr2834 | hepC | Similar to glycosyltransferase | −1.65 | 2.7 × 10−5 |
| alr2835 | hepA | ATP-binding protein of ABC transporter | −1.54 | 5.1 × 10−3 |
| alr2836 | - | Glycosyltransferase | −1.38 | 3.9 × 10−5 |
| alr2837 | - | Glycosyltransferase | −1.07 | 3.2 × 10−3 |
| alr2838 | - | Unknown protein | −1.00 | 2.7 × 10−2 |
| alr2839 | - | Glycosyltransferase | −1.13 | 5.6 × 10−3 |
| alr2841 | - | Unknown protein | −1.21 | 1.9 × 10−3 |
| alr2887 | - | Hypothetical protein | −1.00 | 2.0 × 10−2 |
| all3420 | - | Carboxyl-terminal processing protease | −2.65 | 1.5 × 10−3 |
| all3780 | - | Similar to kinesin light chain | −1.08 | 3.5 × 10−2 |
| all3999 | - | Unknown protein | −1.37 | 9.7 × 10−3 |
| all4000 | - | Photosystem II CP43 protein PsbC homolog | −1.57 | 1.8 × 10−2 |
| all4001 | isiA | Photosystem II chlorophyll a-binding protein | −2.59 | 1.4 × 10−2 |
| all4002 | - | Photosystem II CP43 protein PsbC homolog | −2.18 | 1.3 × 10−2 |
| all4003 | - | Photosystem II CP43 protein PsbC homolog | −1.87 | 1.1 × 10−2 |
| all4020 | talA | Transaldolase | −1.10 | 2.4 × 10−2 |
| all4021 | fbp | Fructose 1,6-bisphosphatase | −1.35 | 9.2 × 10−3 |
| all4160 | - | Probable glycosyltransferase | −1.12 | 1.4 × 10−3 |
| asl4206 | rps17 | 30S ribosomal protein S17 | −1.10 | 4.8 × 10−2 |
| alr4392 | ntcA | Nitrogen-responsive global transcriptional regulator | −1.24 | 6.4 × 10−3 |
| alr4984 | - | Unknown protein | −1.39 | 1.3 × 10−4 |
| alr5256 | - | Biotin acetyl-CoA carboxylase ligase | −1.05 | 5.4 × 10−4 |
| Strain | Nitrogenase activity a | |
|---|---|---|
| (µmol C2H4/mg chla/h) | ||
| 24 h | 48 h | |
| WT | 19.4 ± 7.0 | 17.4 ± 2.5 |
| DRsigCS | 6.0 ± 3.0 | 19.0 ± 5.5 |
| DRsigEK | 19.3 ± 6.3 | 11.0 ± 2.4 |
| ORF No. | Gene | Product | ΔsigG/WT a | p b |
|---|---|---|---|---|
| alr2313 | - | Unknown protein | −2.56 | 4.7 × 10−5 |
| all2564 | pyk1 | Pyruvate kinase | −1.74 | 1.3 × 10−2 |
| alr3281 | - | Hypothetical protein | −1.05 | 7.3 × 10−4 |
| alr3301 | - | Unknown protein | −1.34 | 5.6 × 10−3 |
| alr3445 | - | Hypothetical protein | −1.24 | 9.4 × 10−4 |
| alr3608 | - | Similar to endoglucanase | −1.12 | 3.0 × 10−3 |
| alr3816 | - | Unknown protein | −1.41 | 3.6 × 10−3 |
| alr3817 | - | Unknown protein | −1.09 | 6.6 × 10−3 |
| all3983 | - | Similar to surface layer protein | −1.79 | 5.7 × 10−3 |
| all4254 | - | Unknown protein | −1.07 | 1.6 × 10−4 |
| all4427 | - | Similar to phytanoyl-CoA hydroxylase | −1.03 | 6.5 × 10−4 |
| all4523 | - | Hypothetical protein | −1.32 | 2.3 × 10−3 |
| all4830 | - | Mannosyl transferase | −1.76 | 9.4 × 10−4 |
| alr5340 | - | Hypothetical protein | −1.38 | 4.1 × 10−3 |
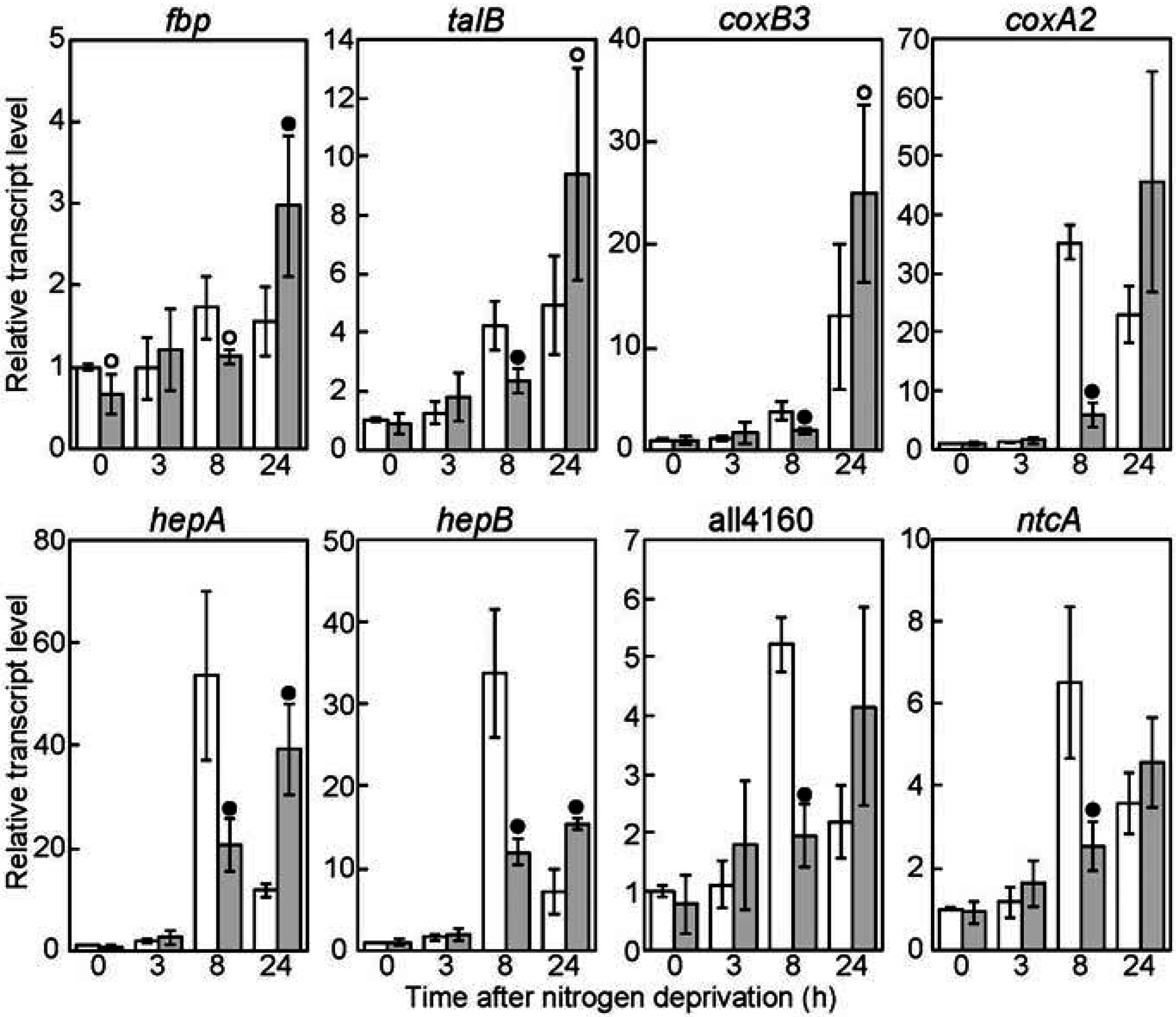
3.3. Genes Regulated by SigC Partially Overlap with Those Regulated by SigE
| ORF No. | Gene | Product | ΔsigE/WT a | p b |
|---|---|---|---|---|
| asl0046 | - | Hypothetical protein | −1.59 | 1.2 × 10−3 |
| all0178 | - | Flavoprotein | −1.00 | 1.5 × 10−3 |
| all0349 | - | Unknown protein | −1.24 | 2.0 × 10−3 |
| all0438 | - | Serine/threonine kinase with WD-40 repeat | −1.24 | 6.7 × 10−3 |
| all0916 | - | ABC transporter ATP-binding subunit | −1.05 | 4.0 × 10−2 |
| all0917 | - | ABC transporter permease protein | −1.22 | 6.6 × 10−4 |
| all0918 | - | Unknown protein | −1.12 | 6.2 × 10−4 |
| all0919 | - | Probable glycosyltransferase | −1.21 | 2.5 × 10−5 |
| alr1407 | nifV1 | Homocitrate synthase | −1.41 | 2.2 × 10−4 |
| asr1408 | nifZ | Iron-sulfur cofactor synthesis protein | −1.06 | 5.3 × 10−3 |
| all1425 | - | Unknown protein | −1.07 | 4.9 × 10−2 |
| all1456 | nifU | Nitrogen fixation protein | −1.00 | 4.5 × 10−2 |
| all1814 | - | Unknown protein | −1.03 | 1.3 × 10−2 |
| alr2514 | coxB2 | Cytochrome c oxidase subunit II | −1.40 | 4.4 × 10−2 |
| alr2515 | coxA2 | Cytochrome c oxidase subunit I | −1.62 | 2.8 × 10−2 |
| alr2516 | coxC2 | Cytochrome c oxidase subunit III | −1.59 | 2.4 × 10−2 |
| alr2517 | - | Hypothetical protein | −1.13 | 7.1 × 10−3 |
| asr2523 | - | Unknown protein | −1.36 | 1.8 × 10−4 |
| alr2524 | - | Unknown protein | −2.04 | 2.5 × 10−3 |
| all2563 | talB | Transaldolase | −1.23 | 1.1 × 10−4 |
| all2564 | pyk1 | Pyruvate kinase | −1.45 | 2.7 × 10−3 |
| all2566 | gap1 | Glyceraldehyde-3-phosphate dehydrogenase | −1.23 | 6.8 × 10−6 |
| all2567 | xfp | Phosphoketolase | −1.33 | 7.3 × 10−5 |
| all2571 | - | Unknown protein | −1.37 | 6.8 × 10−4 |
| alr2582 | - | Hypothetical protein | −1.73 | 1.3 × 10−3 |
| all2635 | - | Polyketide synthase type I | −1.19 | 4.0 × 10−2 |
| all2647 | - | Microcystin synthetase B | −1.16 | 2.6 × 10−2 |
| all2650 | - | ABC transporter ATP-binding protein | −1.39 | 3.4 × 10−2 |
| all2652 | - | Hypothetical protein | −1.03 | 2.8 × 10−2 |
| all2655 | - | Unknown protein | −1.45 | 3.7 × 10−2 |
| alr2729 | - | Hypothetical protein | −1.33 | 3.5 × 10−4 |
| alr2730 | - | Hypothetical protein | −1.12 | 4.5 × 10−5 |
| alr2818 | hetP | Heterocyst differentiation protein | −1.34 | 1.2 × 10−4 |
| alr2822 | - | Hypothetical protein | −1.42 | 3.6 × 10−3 |
| alr2823 | - | Hypothetical protein | −1.60 | 3.6 × 10−3 |
| alr2824 | - | Hypothetical protein | −1.93 | 1.2 × 10−4 |
| alr2825 | - | Glucose-1-P cytidylyltransferase | −1.49 | 3.0 × 10−3 |
| alr2826 | - | Hypothetical protein | −1.75 | 4.7 × 10−6 |
| alr2827 | - | Putative epimerase/dehydratase | −1.42 | 1.0 × 10−4 |
| alr2828 | - | Unknown protein | −1.87 | 4.4 × 10−5 |
| alr2829 | - | Unknown protein | −1.92 | 3.1 × 10−5 |
| alr2830 | rfbC | dTDP-4-dehydrorhamnose 3,5-epimerase | −1.69 | 3.9 × 10−5 |
| alr2831 | - | Probable NAD(P)-dependent oxidoreductase | −1.19 | 4.0 × 10−2 |
| alr2832 | - | Putative glycosyltransferase | −1.69 | 1.0 × 10−6 |
| alr2833 | - | Hypothetical protein | −1.61 | 1.2 × 10−5 |
| alr2834 | hepC | Similar to glycosyltransferase | −1.94 | 1.3 × 10−4 |
| alr2835 | hepA | ATP-binding protein of ABC transporter | −1.74 | 1.4 × 10−5 |
| alr2836 | - | Glycosyltransferase | −1.57 | 2.2 × 10−4 |
| alr2838 | - | Unknown protein | −1.18 | 9.3 × 10−4 |
| alr2839 | - | Glycosyltransferase | −1.17 | 4.9 × 10−4 |
| alr2840 | - | Glycosyltransferase | −1.38 | 1.9 × 10−4 |
| alr2841 | - | Unknown protein | −1.32 | 6.8 × 10−5 |
| alr2857 | - | Unknown protein | −1.22 | 2.6 × 10−3 |
| alr3195 | - | Probable glutathione S-transferase | −1.04 | 1.5 × 10−4 |
| alr3301 | - | Unknown protein | −1.05 | 1.8 × 10−4 |
| all3420 | - | Carboxyl-terminal processing protease | −1.40 | 3.0 × 10−3 |
| alr3698 | hepB | Heterocyst envelope polysaccharide synthesis protein | −1.35 | 4.5 × 10−4 |
| alr3699 | - | Similar to glycosyltransferase | −1.00 | 7.7 × 10−4 |
| alr3710 | devB | Membrane fusion protein of ABC transporter | −1.92 | 1.9 × 10−4 |
| alr3711 | devC | Substrate-binding protein of ABC transporter | −1.71 | 9.3 × 10−6 |
| all3773 | - | Serine/threonine kinase | −1.47 | 5.9 × 10−3 |
| all3780 | - | Similar to kinesin light chain | −1.80 | 1.4 × 10−3 |
| all3793 | - | Unknown protein | −1.25 | 3.9 × 10−3 |
| all4160 | - | Probable glycosyltransferase | −1.27 | 2.3 × 10−3 |
| alr4984 | - | Unknown protein | −1.60 | 1.4 × 10−3 |
| all4991 | desC | Delta-9 desaturase | −1.06 | 2.2 × 10−2 |
| all5341 | hglT | Heterocyst-specific glycolipid synthase | −1.65 | 6.0 × 10−3 |
| alr5358 | hetN | Ketoacyl reductase | −1.34 | 1.9 × 10−3 |
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flores, E.; Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol. 2010, 8, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Vioque, A.; Haas, F.; Hess, W.R.; Muro-Pastor, A.M. Dynamics of transcriptional start site selection during nitrogen stress-induced cell differentiation in Anabaena sp. PCC7120. Proc. Natl. Acad. Sci. USA 2011, 108, 20130–20135. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, B.L.; van Nieuwerburgh, F.; Head, S.R.; Golden, J.W. Directional RNA deep sequencing sheds new light on the transcriptional response of Anabaena sp. strain PCC 7120 to combined-nitrogen deprivation. BMC Genomics 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M.; Sato, N. Genome-wide expression analysis of the responses to nitrogen deprivation in the heterocyst-forming cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. 2003, 10, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-responsive response regulator facilitates heterocyst development in the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 2006, 59, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Muro-Pastor, A.M.; Valladares, A.; Flores, E. Cellular differentiation and the NtcA transcription factor in filamentous cyanobacteria. FEMS Microbiol. Rev. 2004, 28, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-X.; Jiang, Y.-L.; He, Y.-X.; Chen, Y.-F.; Teng, Y.-B.; Chen, Y.; Zhang, C.-C.; Zhou, C.-Z. Structural basis for the allosteric control of the global transcription factor NtcA by the nitrogen starvation signal 2-oxoglutarate. Proc. Natl. Acad. Sci. USA 2010, 107, 12487–12492. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, A.M.; Olmedo-Verd, E.; Flores, E. All4312, an NtcA-regulated two-component response regulator in Anabaena sp. strain PCC 7120. FEMS Microbiol. Lett. 2006, 256, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA directly regulates expression of hetR during heterocyst differentiation in the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2006, 188, 8520–8525. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Characterization of a gene controlling heterocyst differentiation in the cyanobacterium Anabaena 7120. Genes Dev. 1991, 5, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Expression of the Anabaena hetR gene from a copper-regulated promoter leads to heterocyst differentiation under repressing conditions. Proc. Natl. Acad. Sci. USA 2001, 98, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dong, Y.; Zhao, J. HetR homodimer is a DNA-binding protein required for heterocyst differentiation, and the DNA-binding activity is inhibited by PatS. Proc. Natl. Acad. Sci. USA 2004, 101, 4848–4853. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Ni, S.; Rivers, O.S.; Ushijima, B.; Feldmann, E.A; Cozy, L.M.; Kennedy, M.A.; Callahan, S.M. Expanding the direct HetR regulon in Anabaena sp. strain PCC 7120. J. Bacteriol. 2014, 196, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, B.L.; Johnson, D.; Golden, J.W. Deep sequencing of HetR-bound DNA reveals novel HetR targets in Anabaena sp. strain PCC7120. BMC Microbiol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Camargo, S.; Valladares, A.; Flores, E.; Herrero, A. Transcription activation by NtcA in the absence of consensus NtcA-binding sites in an anabaena heterocyst differentiation gene promoter. J. Bacteriol. 2012, 194, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, A.M.; Valladares, A.; Flores, E.; Herrero, A. Mutual dependence of the expression of the cell differentiation regulatory protein HetR and the global nitrogen regulator NtcA during heterocyst development. Mol. Microbiol. 2002, 44, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Wösten, M.M. Eubacterial sigma-factors. FEMS Microbiol. Rev. 1998, 22, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Nakamura, Y.; Wolk, C.P.; Kuritz, T.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Ishikawa, A.; Kawashima, K.; Kimura, T.; et al. Complete genomic sequence of the filamentous nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. 2001, 8, 205–213, 227–253. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, H.; Okamoto, S.; Tsumuraya, Y.; Ohmori, M. Group 3 sigma factor gene, sigJ, a key regulator of desiccation tolerance, regulates the synthesis of extracellular polysaccharide in cyanobacterium Anabaena sp. strain PCC 7120. DNA Res. 2007, 14, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Brahamsha, B.; Haselkorn, R. Identification of multiple RNA polymerase sigma factor homologs in the cyanobacterium Anabaena sp. strain PCC 7120: Cloning, expression, and inactivation of the sigB and sigC genes. J. Bacteriol. 1992, 174, 7273–7282. [Google Scholar] [PubMed]
- Mella-Herrera, R.A; Neunuebel, M.R.; Kumar, K.; Saha, S.K.; Golden, J.W. The sigE gene is required for normal expression of heterocyst-specific genes in Anabaena sp. strain PCC 7120. J. Bacteriol. 2011, 193, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.R.; Mella-Herrera, R.A; Golden, J.W. Sigma factor genes sigC, sigE, and sigG are upregulated in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 8392–8396. [Google Scholar] [CrossRef] [PubMed]
- Khudyakov, I.Y.; Golden, J.W. Identification and inactivation of three group 2 sigma factor genes in Anabaena sp. strain PCC 7120. J. Bacteriol. 2001, 183, 6667–6675. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-regulated response regulator protein, controls glycogen catabolism in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. J. Biol. Chem. 2011, 286, 38109–38114. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. The pknH gene restrictively expressed in heterocysts is required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2012, 158, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Wiest, D.R. Genome rearrangement and nitrogen fixation in Anabaena blocked by inactivation of xisA gene. Science 1988, 242, 1421–1423. [Google Scholar] [CrossRef] [PubMed]
- Black, T.A.; Cai, Y.; Wolk, C.P. Spatial expression and autoregulation of hetR, a gene involved in the control of heterocyst development in Anabaena. Mol. Microbiol. 1993, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 1998–2005. [Google Scholar] [PubMed]
- Pinto, F.L.; Thapper, A.; Sontheim, W.; Lindblad, P. Analysis of current and alternative phenol based RNA extraction methodologies for cyanobacteria. BMC Mol. Biol. 2009, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Mackinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar]
- Ehira, S. Transcriptional Regulation of Heterocyst Differentiation in Anabaena sp. strain PCC 7120. Russ. J. Plant Physiol. 2013, 60, 443–452. [Google Scholar] [CrossRef]
- Krasikov, V.; Aguirre von Wobeser, E.; Dekker, H.L.; Huisman, J.; Matthijs, H.C.P. Time-series resolution of gradual nitrogen starvation and its impact on photosynthesis in the cyanobacterium Synechocystis PCC 6803. Physiol. Plant. 2012, 145, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, A.M.; Herrero, A.; Flores, E. Nitrogen-regulated group 2 sigma factor from Synechocystis sp. strain PCC 6803 involved in survival under nitrogen stress. J. Bacteriol. 2001, 183, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Olmedo-Verd, E.; Muro-Pastor, A.M.; Flores, E.; Herrero, A. Localized induction of the ntcA regulatory gene in developing heterocysts of Anabaena sp. strain PCC 7120. J. Bacteriol. 2006, 188, 6694–6699. [Google Scholar] [CrossRef] [PubMed]
- Lechno-Yossef, S.; Fan, Q.; Wojciuch, E.; Wolk, C.P. Identification of ten Anabaena sp. genes that under aerobic conditions are required for growth on dinitrogen but not for growth on fixed nitrogen. J. Bacteriol. 2011, 193, 3482–3489. [Google Scholar] [CrossRef] [PubMed]
- Valladares, A.; Herrero, A.; Pils, D.; Schmetterer, G.; Flores, E. Cytochrome c oxidase genes required for nitrogenase activity and diazotrophic growth in Anabaena sp. PCC 7120. Mol. Microbiol. 2003, 47, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.M.; Haselkorn, R. Newly identified cytochrome c oxidase operon in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120 specifically induced in heterocysts. J. Bacteriol. 2002, 184, 2491–2499. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lechno-Yossef, S.; Gong, Y.; Fan, Q.; Wolk, C.P.; Xu, X. Predicted glycosyl transferase genes located outside the HEP island are required for formation of heterocyst envelope polysaccharide in Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 5372–5378. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, G.; Arnold, M.; Hannus, S.; Maldener, I. The DevBCA exporter is essential for envelope formation in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 1998, 27, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Awai, K.; Wolk, C.P. Identification of the glycosyl transferase required for synthesis of the principal glycolipid characteristic of heterocysts of Anabaena sp. strain PCC 7120. FEMS Microbiol. Lett. 2007, 266, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Callahan, S.M.; Buikema, W.J. The role of HetN in maintenance of the heterocyst pattern in Anabaena sp. PCC 7120. Mol. Microbiol. 2001, 40, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Imamura, S.; Tanaka, K.; Shirai, M.; Asayama, M. Growth phase-dependent activation of nitrogen-related genes by a control network of group 1 and group 2 sigma factors in a cyanobacterium. J. Biol. Chem. 2006, 281, 2668–2675. [Google Scholar] [CrossRef] [PubMed]
- Kafri, R.; Springer, M.; Pilpel, Y. Genetic redundancy: New tricks for old genes. Cell 2009, 136, 389–392. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ehira, S.; Miyazaki, S. Regulation of Genes Involved in Heterocyst Differentiation in the Cyanobacterium Anabaena sp. Strain PCC 7120 by a Group 2 Sigma Factor SigC. Life 2015, 5, 587-603. https://doi.org/10.3390/life5010587
Ehira S, Miyazaki S. Regulation of Genes Involved in Heterocyst Differentiation in the Cyanobacterium Anabaena sp. Strain PCC 7120 by a Group 2 Sigma Factor SigC. Life. 2015; 5(1):587-603. https://doi.org/10.3390/life5010587
Chicago/Turabian StyleEhira, Shigeki, and Shogo Miyazaki. 2015. "Regulation of Genes Involved in Heterocyst Differentiation in the Cyanobacterium Anabaena sp. Strain PCC 7120 by a Group 2 Sigma Factor SigC" Life 5, no. 1: 587-603. https://doi.org/10.3390/life5010587
APA StyleEhira, S., & Miyazaki, S. (2015). Regulation of Genes Involved in Heterocyst Differentiation in the Cyanobacterium Anabaena sp. Strain PCC 7120 by a Group 2 Sigma Factor SigC. Life, 5(1), 587-603. https://doi.org/10.3390/life5010587