
Opinion: The Key Steps in the Origin of Life to the Formation of the Eukaryotic Cell



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Some Guiding Principles
1.1.1. Steady and Substantial Fluxes
1.1.2. QWERTY and the First Metabolism
1.1.3. Pre-Adaptation
1.1.4. The Likely Complexity of Life’s Evolutionary Path
1.1.5. The Application of Guiding Principles
1.2. A Short History of Our Current Understanding
1.3. The Definition of Life
2. The Emergence of the Modern Synthesis
3. Venue—What Was the Nature of the Cradles of Life?
3.1. Mid-Ocean Ridge Hydrothermal Vent Systems
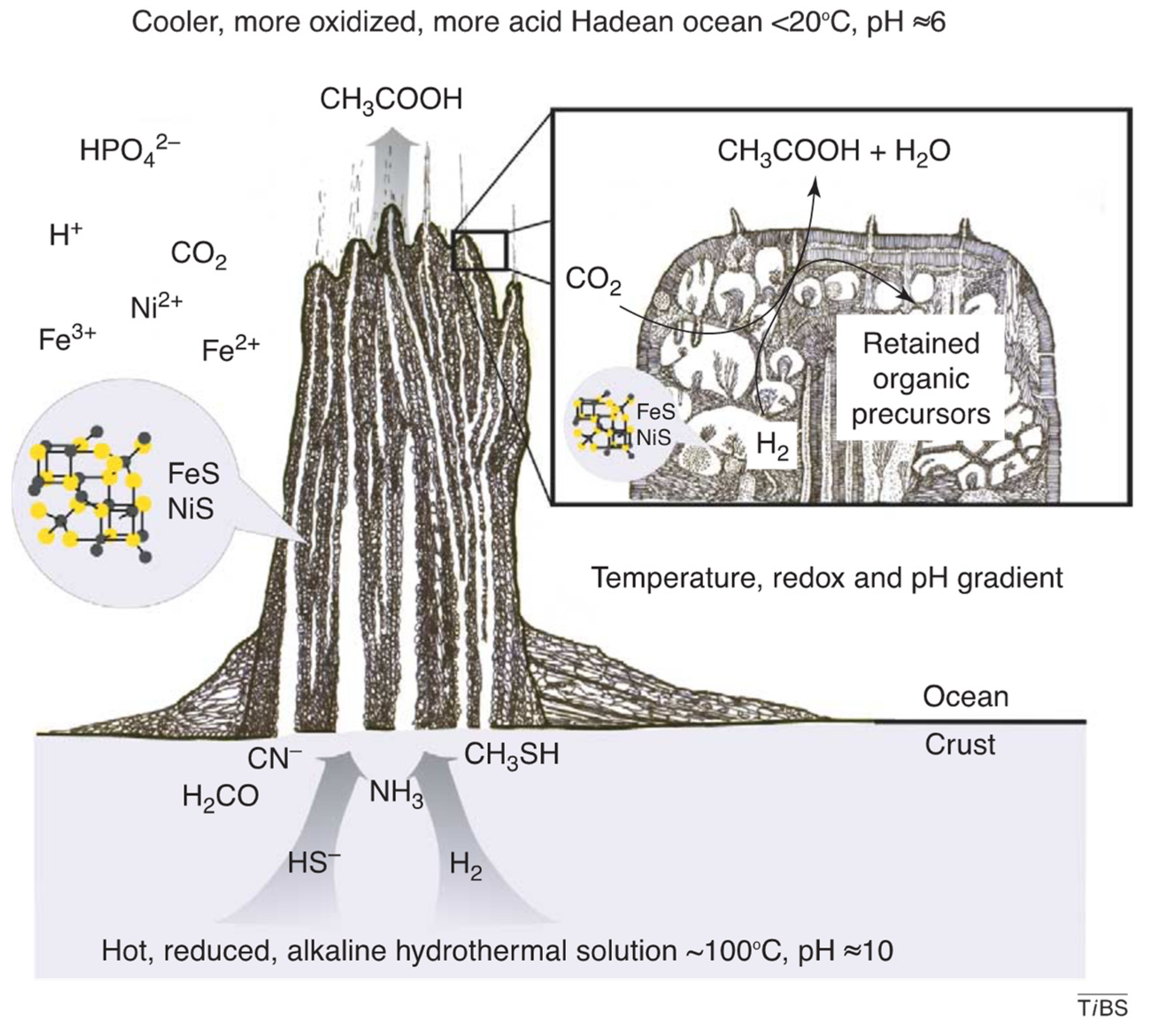
3.2. Alkaline Hydrothermal Vent Systems
4. Stages in the Development of Life
4.1. Pre-Biotic Stage
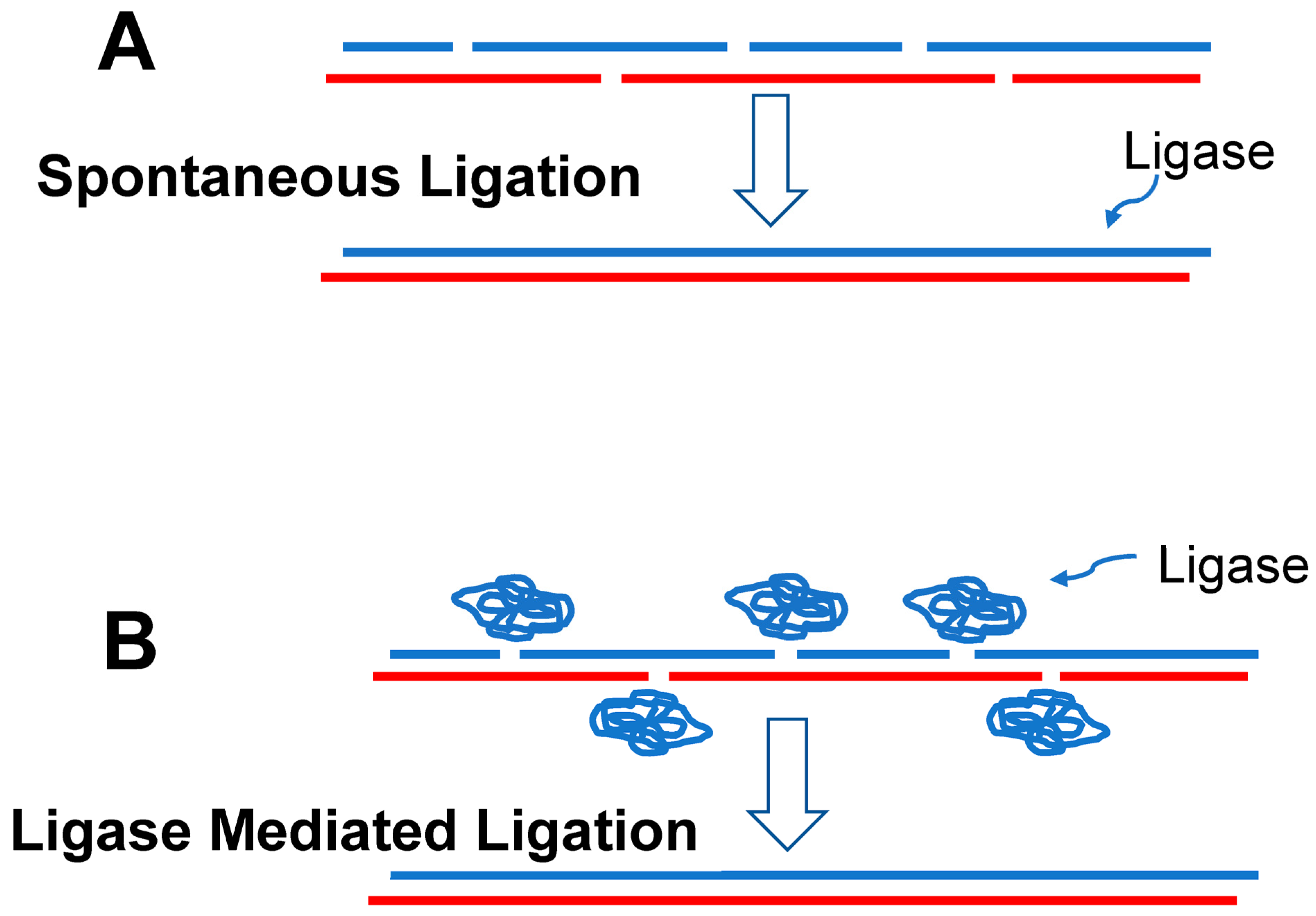
4.1.1. Initial RAFs
4.1.2. Proteinoids
4.1.3. Polynucleotides
4.1.4. Energy Storage Molecules
4.2. RNA and First Life
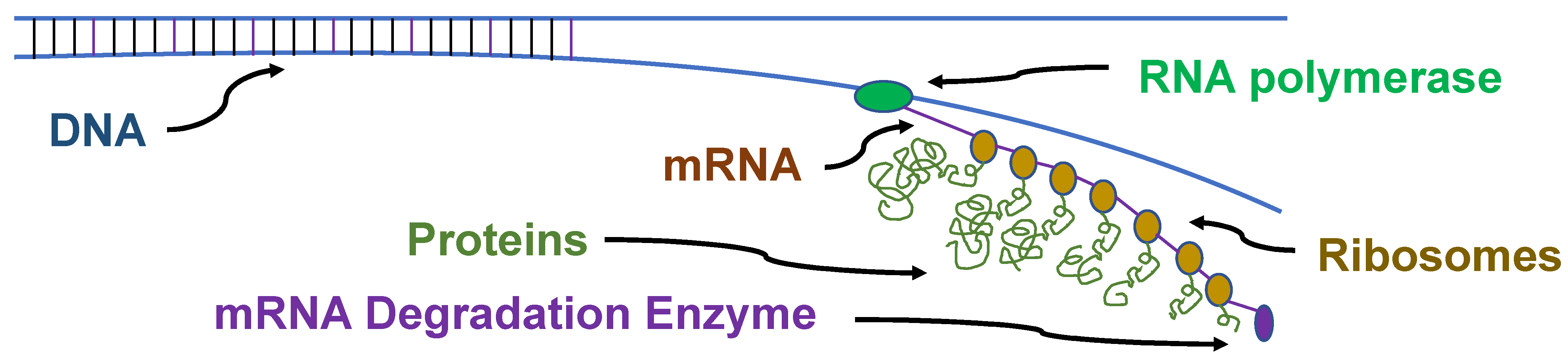
4.3. Protein Synthesis
4.4. Exploiting the Natural Proton Gradient to Augment ATP Production
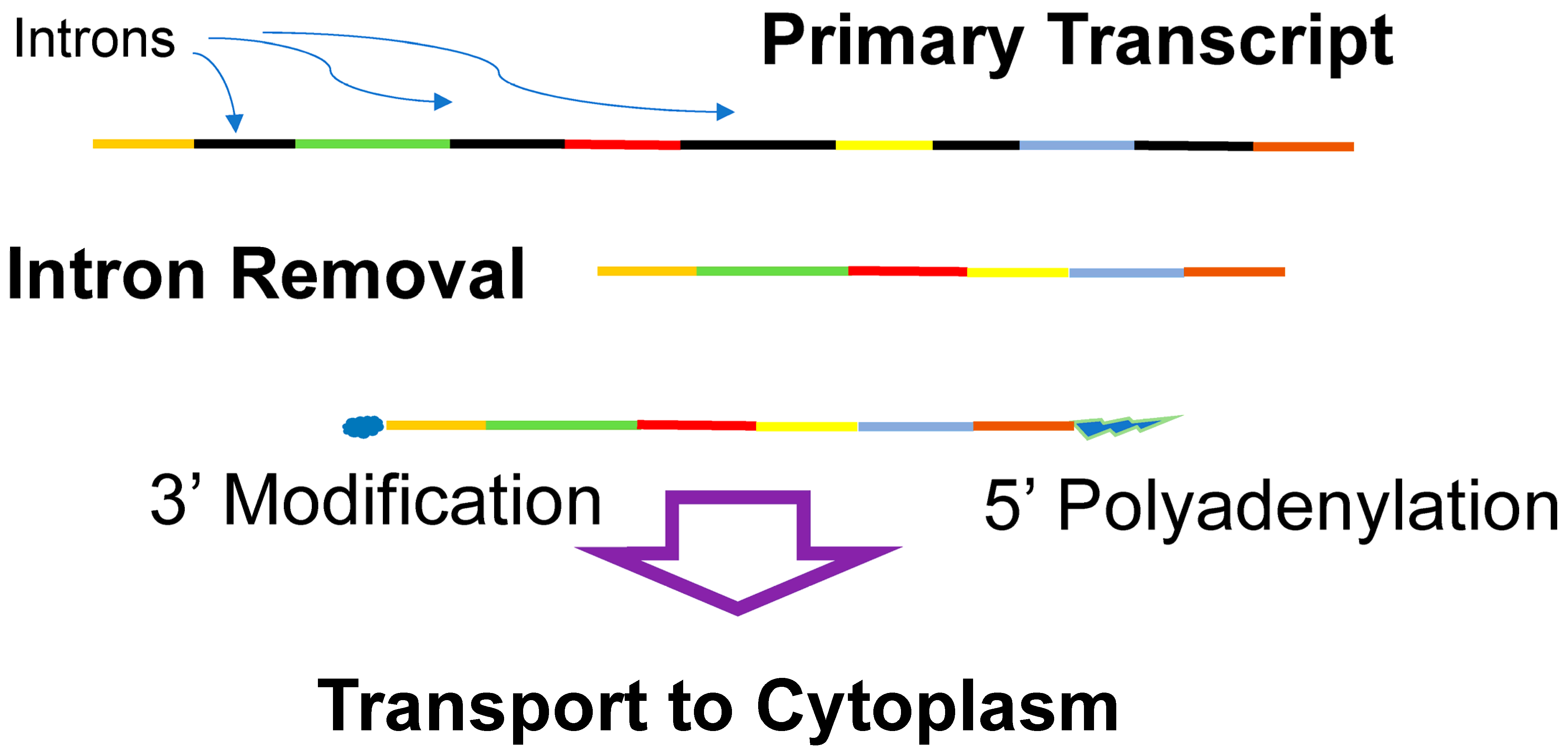
4.5. DNA
5. Cellularization and Life beyond the Vent
5.1. At Least Two Cellularization Events
5.2. The Diversification of Electron Donors and the Origin of Photosynthesis
6. Prokaryotic Cells versus Eukaryotic Cells
7. Moving beyond a Gene-Centric View
8. The Origin of Eukaryotic Cells
9. The Deep Evolutionary Heritage of the Eukaryotic Cell
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- David, P.A. Clio and the Economics of QWERTY. Am. Econ. Rev. 1985, 75, 332–337. [Google Scholar]
- David, P.A. Path Dependence and the Quest for Historical Economics: One More Chorus of the Ballad of QWERTY; Discussion Papers in Economic and Social History; University of Oxford: Oxford, UK, 1997. [Google Scholar]
- Neil, M.K. Rerun the Tape of History and QWERTY Always Wins. Res. Policy 2013, 42, 1175–1185. [Google Scholar]
- Noyes, J. The QWERTY Keyboard: A Review. Int. J. Man-Mach. Stud. 1983, 18, 265–281. [Google Scholar] [CrossRef]
- Schopf, J.W. The Evolution of the Earliest Cells. Sci. Am. 1978, 239, 110–139. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthesis CO2 Assimilation in leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Schönheit, P.; Buckel, W.; Martin, W.F. On the Origin of Heterotrophy. Trends Microbiol. 2016, 24, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F. Older than Genes: The Acetyl CoA Pathway and Origins. Front. Microbiol. 2020, 11, 817. [Google Scholar] [CrossRef]
- Preiner, M.; Igarashi, K.; Muchowska, B.K.; Yu, M.; Varma, S.J.; Kleinermanns, K.; Nobu, M.K.; Kamagata, Y.; Tüysüz, H.; Moran, J.; et al. A hydrogen-dependent geochemical analogue of primordial carbon and energy metabolism. Nat. Ecol. Evol. 2020, 4, 534–542. [Google Scholar] [CrossRef]
- Marshall, C.; Schultze, H.-P. Relative Importance of Molecular, Neontological, and Paleontological Data in the Understanding the Biology of the Vertebrate Invasion of Land. J. Mol. Evol. 1992, 35, 93–101. [Google Scholar] [CrossRef]
- Long, J.A.; Gordon, M.S. The Greatest Step in Vertebrate History: A Paleobiological Review of the Fish-Tetrapod Transition. Physiol. Biochem. Zool. 2004, 77, 700–719. [Google Scholar] [CrossRef]
- Capel, E.; Cleal, C.J.; Xue, J.; Monnet, C.; Servais, T.; Cascales-Miñana, B. The Silurian–Devonian terrestrial revolution: Diversity patterns and sampling bias of the vascular plant macrofossil record. Earth-Sci. Rev. 2022, 231, 104085. [Google Scholar] [CrossRef]
- Strullu-Derrien, C. Fossil filamentous microorganisms associated with plants in early terrestrial environments. Curr. Opin. Plant Biol. 2018, 44, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, J.A.; Garwood, R.J. Terrestrial Invertebrates in the Rhynie chert Ecosystem. Phil. Trans. R. Soc. B 2017, 373, 20160493. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.R. Explaining the Cambrian “Explosion” of Animals. Annu. Rev. Earth Planet. Sci. 2006, 34, 355–384. [Google Scholar] [CrossRef]
- Marshall, C.R.; Valentine, J.W. The Importance of Preadapted Genomes in the Origin of the Animal Bodyplans and the Cambrian Explosion. Evolution 2010, 64, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Valentine, J.W.; Marshall, C.R. Fossil and Transcriptomic Perspectives on the Origins and Success of Metazoan Multicellularity. Evol. Transit. Multicell. Life 2015, 2, 31–46. [Google Scholar] [CrossRef]
- Cole, D.B.; Mills, D.B.; Ewin, D.H.; Sperling, E.A.; Porter, S.M.; Reinhard, C.T.; Planavsky, N.J. On the co-evolution of surface oxygen levels and animals. Geobiology 2020, 18, 260–281. [Google Scholar] [CrossRef] [PubMed]
- Brunk, C.F.; Martin, W.F. Archaeal Histone Contributions to the Origin of Eukaryotes. Trends Microbiol. 2019, 27, 703–714. [Google Scholar] [CrossRef]
- Martin, W.; Garg, S.; Zimorski, V. Endosymbiotic Theories for Eukaryote Origin. Phil. Trans. R. Soc. B 2015, 370, 20140330. [Google Scholar] [CrossRef]
- Martin, W.F. Symbiogenesis, gradualism, and mitochondrial energy in eukaryote evolution. Period. Biol. 2017, 119, 141–158. [Google Scholar] [CrossRef]
- Gavette, J.V.; Stoop, M.; Hud, N.V.; Krishnamurth, R. RNA–DNA Chimeras in the Context of an RNA World Transition to an RNA/DNA World. Angew. Chem. Int. Ed. 2016, 55, 13204–13209. [Google Scholar] [CrossRef] [PubMed]
- Holler, S.; Bartlett, S.; Löffler, R.J.; Casiraghi, F.; Diaz, C.I.S.; Cartwright, J.H.; Hanczyc, M.M. Hybrid organic–inorganic structures trigger the formation of primitive cell-like compartments. Proc. Natl. Acad. Sci. USA 2023, 120, e2300491120. [Google Scholar] [CrossRef] [PubMed]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Evolution of Photosynthesis. Annu. Rev. Plant Biol. 2011, 62, 515–548. [Google Scholar] [CrossRef]
- Wills, P.R.; Carter, C.W. Insuperable problems of the genetic code initially emerging in an RNA world. BioSystems 2018, 164, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Wills, P.R.; Carter, C.W. Impedance Matching and the Choice Between Alternative Pathways for the Origin of Genetic Coding. Int. J. Mol. Sci. 2020, 21, 7392. [Google Scholar] [CrossRef]
- Carter, C.W.; Wills, P.R. Reciprocally-Coupled Gating: Strange Loops in Bioenergetics, Genetics, and Catalysis. Biomolecules 2021, 11, 265. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D. Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions. Life 2021, 11, 134. [Google Scholar] [CrossRef]
- Sasselov, D.D.; Grotzinger, J.P.; Sutherland, J.D. The origin of life as a planetary phenomenon. Sci. Adv. 2020, 6, eaax3419. [Google Scholar] [CrossRef]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef]
- Xu, J.; Chmela, V.; Green, N.; Russell, D.A.; Janicki, M.J.; Góra, R.W.; Szabla, R.; Bond, A.D.; Sutherland, J.D. Selective Prebiotic Formation of RNA Pyrimidine and DNA Purine Nucleosides. Nature 2020, 582, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.D. Studies on the origin of life—The end of the beginning. Nat. Rev. Chem. 2017, 1, 0012. [Google Scholar] [CrossRef]
- Brunk, C.F.; Marshall, C.R. ‘Whole Organism’, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life. Life 2021, 11, 690. [Google Scholar] [CrossRef]
- Marshall, M. The Genesis Quest: The Geniuses and Eccentrics on a Journey to Uncover the Origin of Life on Earth; University of Chicago Press: Chicago, IL, USA, 2020; 368p. [Google Scholar]
- Russell, M.J. The “Water Problem” (sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life 2021, 11, 429. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Brack, A.; Fairén, A.G.; Schulte, M.D. Setting the Geological Scene for the Origin of Life and Continuing Open Questions about its Emergence. Front. Astron. Space Sci. 2023, 9, 1095701. [Google Scholar] [CrossRef] [PubMed]
- Schwander, L.; Brabender, M.; Mrnjavac, N.; Wimmer, J.L.E.; Preiner, M.; Martin, W.F. Serpentinization as the Source of Energy, Electrons, Organics, Catalysts, Nutrients and pH Gradients for the Origin of LUCA and Life. Front. Microbiol. 2023, 14, 1257597. [Google Scholar] [CrossRef] [PubMed]
- Lazcano, A. Historical Development of Origins Research. Cold Spring Harb. Perspect. Biol. 2010, 2, a002089. [Google Scholar] [CrossRef]
- Oparin, A.I. The Origin of Life, 1st ed.; Macmillan: New York, NY, USA, 1938. [Google Scholar]
- Haldane, J.B.S. The Origin of Life. In The Rationalist Annual; The Rationalist Press Association: London, UK, 1929; Volume 3, pp. 3–10. [Google Scholar]
- Bernal, J.D. The Problem of Stages in Biopoesis. In Aspects of the Origin of Life; Florkin, M., Ed.; Pergamon Press: New York, NY, USA, 1960; pp. 30–45. [Google Scholar]
- Mereschkowsky, C. Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen. Biol. Centralbl. 1910, 30, 278–288. [Google Scholar]
- Haeckel, E. Natürliche Schöpfungs-Geschichte. Gemeinverständliche Wissenschaftliche Vorträge über die Entwickelungslehre Zehnte verbesserte Auflage. Zweiter Theil: Allgemeine Stammesgeschichte; Georg Reimer Verlag: Berlin, Germany, 1902. [Google Scholar]
- Miller, S.L.; Urey, H.C. Organic Compound Synthesis on the Primitive Earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef]
- Cech, T.R. The Chemistry of Self-Splicing RNA and RNA Enzymes. Science 1987, 236, 1532–1539. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. the AT3-60 Shipboard Party An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30° N. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef]
- Kelley, D.S. From the Mantle to Microbes the Lost City Hydrothermal Field. Oceanography 2005, 18, 33–45. [Google Scholar] [CrossRef]
- Camprubí, E.; de Leeuw, J.W.; House, C.H.; Raulin, F.; Russell, M.J.; Spang, A.; Tirumalai, M.R.; Westall, F. The Emergence of Life. Space Sci. Rev. 2019, 215, 56. [Google Scholar] [CrossRef]
- Nutman, A.P.; Mojzsis, S.J.; Friend, C.R.L. Recognition of ≥3850 Ma water-lain sediments in West Greenland and their significance for the early Archaean Earth. Geochim. Cosmochim. Acta 1997, 61, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Dalrymple, G.B. The Age of the Earth; Stanford University Press: Stanford, CA, USA, 1991. [Google Scholar]
- Cavalazzi, B.; Lemelle, L.; Simionovici, A.; Cady, S.L.; Russell, M.J.; Bailo, E.; Canteri, R.; Enrico, E.; Manceau, A.; Maris, A.; et al. Cellular remains in a ~3.42-billion-year-old subseafloor hydrothermal environment. Sci. Adv. 2021, 7, eabf3963. [Google Scholar] [CrossRef] [PubMed]
- Voytek, M.A. About Life Detection. 2023. Available online: https://astrobiology.nasa.gov (accessed on 15 October 2023).
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favorured Races in the Struggle for Life, 1st ed.; John Murray: London, UK, 1859. [Google Scholar]
- Huxley, J. EVOLUTION. The Modern Synthesis; George Allen and Unwin Ltd.: London, UK, 1942. [Google Scholar]
- Mendel, G.J. Versuche über Pflanzeb-Hybriden. Verh. Naturforschenden Ver. Brünn 1865, 4, 3–47. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Mayr, E. Systematics and the Origin of Species; Harvard University Press: Cambridge, MA, USA, 1942. [Google Scholar]
- Fairbank, D.J. Mendel and Darwin: Untangling a persistent enigma. Heredity 2020, 124, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Kolb, V.M. Defining Life: Multiple Perspectives. In Handbook of Astrobiology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2018; Chapter 2.1; pp. 57–64. [Google Scholar] [CrossRef]
- Pross, A.; Pascal, R. On the Emergence of Autonomous Chemical Systems through Dissipation Kinetics. Life 2023, 13, 2171. [Google Scholar] [CrossRef]
- Bowler, P. Evolution: The History of an Idea; University of California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- Koonin, E.V. Darwinian evolution in the light of genomics. Nucleic Acids Res. 2009, 37, 1011–1034. [Google Scholar] [CrossRef]
- Berry, A.; Browne, J. Mendel and Darwin. Proc. Natl. Acad. Sci. USA 2022, 119, e2122144119. [Google Scholar] [CrossRef]
- Avery, O.T.; MacLeod, C.M.; McCarty, M. Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types. J. Exp. Med. 1944, 79, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Crick, F.H. On Protein Synthesis. In Symposia of the Society for Experimental Biology, Number XII: The Biological Replication of Macromolecules; Sanders, F.K., Ed.; Cambridge University Press: Cambridge, UK, 1958; pp. 138–163. [Google Scholar]
- Temin, H.M. Homology between RNA from Rous Sarcoma Virus and DNA from Rous Sarcoma Virus-Infected Cells. Proc. Natl. Acad. Sci. USA 1964, 52, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, D. Viral RNA-dependent DNA Polymerase. Nature 1970, 226, 1209–1211. [Google Scholar] [CrossRef] [PubMed]
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral Gene transfer and the Nature of Bactrial Innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Qiu, H.; Han, W.; Wang, Y.; Xu, D.; Zhang, X.; Bhattacharya, D.; Ye, N. Phytoplankton pangenome reveals extensive prokaryotic horizontal gene transfer of diverse functions. Sci. Adv. 2020, 6, eaba0111. [Google Scholar] [CrossRef]
- Etten, J.V.; Bhattacharya, D. Horizontal Gene Transfer in Eukaryotes: Not if, but How Much? Trends Genet. 2020, 36, 915–925. [Google Scholar] [CrossRef]
- Martin, W.F. Physiology, phylogeny, and the energetic roots of life. Period. Biol. 2016, 118, 343–352. [Google Scholar] [CrossRef][Green Version]
- Shapiro, J.; Noble, D. What prevents mainstream evolutionists teaching the whole truth. Progr. Biophys. Mol. Biol. 2021, 165, 140–152. [Google Scholar] [CrossRef]
- Szostak, J.W. On the Origin of Life. Medicina 2016, 76, 199–203. [Google Scholar]
- Schrum, J.P.; Zhu, T.F.; Szostak, J.W. The Origins of Cellular Life. Cold Spring Harb. Perspect. Biol. 2010, 2, a002212. [Google Scholar] [CrossRef]
- Ritson, D.J.; Mojzsis Sutherland, J.D. Supply of Phosphate to Early Earth by Photogeochemistry after Meteoritic Weathering. Nat. Geosci. 2020, 13, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Bonfio, C.; Valer, L.; Scintilla, S.; Shah, S.; Evans, D.J.; Jin, L.; Szostak, J.W.; Sasselov, D.D.; John, D.; Sutherland, J.D.; et al. UV-Light-driven Prebiotic Synthesis of Iron–sulfur Clusters. Nat. Chem. 2017, 9, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.W.; Harada, K. Thermal copolymerization of amino acids to a product resembling protein. Science 1985, 128, 1214. [Google Scholar] [CrossRef]
- Fox, S.W. Molecular Evolution to the First Cells. Pure Appl. Chem. 1973, 34, 641–669. [Google Scholar] [CrossRef] [PubMed]
- Wegener, A.L. Die Entstehung der Kontinente. Geol. Rundsch. 1912, 3, 276–292. [Google Scholar] [CrossRef]
- Corliss, J.B.; Dymond, J.; Gordon, L.I.; Edmond, J.M.; von Herzen, R.P.; Ballard, R.D.; Green, K.; Williams, D.; Bainbridge, A.; Crane, K.; et al. Submarine Thermal Springs on the Galapagos Rift. Science 1979, 203, 1073–1083. [Google Scholar] [CrossRef]
- Ballard, R.D. Notes on a Major Oceanographic Find. Oceanus 1977, 20, 35–44. [Google Scholar]
- Corliss, J.B.; Baross, J.A.; Hoffman, S.E. An Hypothesis Concerning the Relationship between Submarine Hot Springs and the Origin of Life on Earth. Oceanol. Acta 1981, 4, 59–69. [Google Scholar]
- Baross, J.A.; Hoffman, S.E. Submarine Hydrothermal Vents and Associated Gradient Environments as Sites for the Origin and Evolution of Life. Orig. Life 1985, 15, 327–345. [Google Scholar] [CrossRef]
- Miller, S.L.; Bada, J.L. Submarine Hot Springs and the Origin of Life. Nature 1988, 334, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J.; Cairns-Smith, A.G.; Braterman, P.S. Submarine Hot Springs and the Origin of Life. Nature 1988, 336, 117. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Turner, D. In vitro Growth of iIron Sulphide Chimneys: Possible Culture Chambers for Origin-of-Life Experiments. Terra Nova 1989, 1, 238–241. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Frúh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, D.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A Serpentinite-Hosted Ecosystem: The Lost City Hydrothermal Field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J. The Importance of Being Alkaline. Science 2003, 302, 580–581. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a Source of Energy at the Origin of Life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Branscomb, E.; Russell, M.J. Turnstiles and bifurcators: The disequilibrium converting engines that put metabolism on the road. Biochim. Biophys. Acta 2018, 1827, 62–78. [Google Scholar] [CrossRef]
- Russell, M.J.; Barge, L.M.; Bhartia, R.; Bocanegra, D.; Bracher, P.J.; Branscomb, E.; Kidd, R.; McGlynn, S.; Meier, D.H.; Nitschke, W.; et al. The Drive to Life on Wet and Icy Worlds. Astrobiology 2014, 14, 308–343. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. Why the Submarine Alkaline Vent is the Most Reasonable Explanation for the Emergence of Life. BioEssays 2018, 41, 62–78. [Google Scholar] [CrossRef]
- Koonin, E.V.; Matrin, W. On the origin of genomes and cells within inorganic compartments. Trends Genet. 2005, 21, 647–654. [Google Scholar] [CrossRef]
- Russell, M.J.; Martin, W. The rocky roots of the acetyl-CoA pathway. Trends Biochem. Sci. 2004, 29, 358–363. [Google Scholar] [CrossRef]
- Martin, W.; Russell, M.J. On the Origin of Biochemistry at an Alkaline Hydrothermal Vent. Phil. Trans. R. Soc. B 2007, 362, 1887–1925. [Google Scholar] [CrossRef]
- Belthle, K.S.; Beyazay, T.; Ochoa-Hernández, C.; Miyazaki, R.; Foppa, L.; Martin, W.F.; Tüysüz, H. Effects of Silica Mo6dification (Mg, Al, Ca, Ti, and Zr) on Supported Cobalt Catalysts for H2-Dependent CO2 Reduction to Metabolic Intermediates. J. Am. Chem. Soc. 2022, 144, 21232–21243. [Google Scholar] [CrossRef] [PubMed]
- Beyazay, T.; Ochoa-Hernández, C.; Song, Y.; Belthle, K.S.; Martin, W.F.; Tüysüz, H. Influence of Composition of Nickel-Iron Nanoparticles for Abiotic CO2 Conversion to Early Prebiotic Organics. Angew. Chem. 2023, 135, e202218189. [Google Scholar] [CrossRef]
- Russell, M.J. Cobalt: A must-have element for life and livelihood. Proc. Natl. Acad. Sci. USA 2022, 119, e2121307119. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J. Green Rust: The Simple Organizing ‘Seed’ of All Life? Life 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Mrnjavac, N.; Wimmer, J.L.E.; Brabender, M.; Schwander, L.; Martin, W.F. The Moon-Forming Impact and the Autotrophic Origin of Life. ChemPlusChem 2023, 88, e202300270. [Google Scholar] [CrossRef] [PubMed]
- Beyazay, T.; Belthle, K.S.; Farès, C.; Preiner, M.; Moran, J.; Martin, W.F.; Tüysüz, H. Ambient Temperature CO2 Fixation to Pyruvate and Subsequently to Citramalate over Iron and Nickel Nanoparticles. Nat. Commun. 2023, 14, 570. [Google Scholar] [CrossRef]
- Hordijk, W.; Steel, M. Detecting autocatalytic, self-sustaining sets in chemical reaction systems. J. Theor. Biol. 2004, 227, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The Physiology and Habitat of the Last Universal Common Ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef]
- Xavier, J.C.; Hordijk, W.; Kauffman, S.; Steel, M.; Martin, W.F. Autocatalytic chemical networks at the origin of metabolism. Proc. R. Soc. B 2020, 287, 2377–2387. [Google Scholar] [CrossRef]
- Lane, N.; Allen, J.F.; Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 2010, 32, 217–280. [Google Scholar] [CrossRef]
- Kauffman, S.A. Approaches to the Origin of Life on Earth. Life 2011, 1, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Corning, P.A.; Kauffman, S.A.; Noble, D.; Shapiro, J.A.; Vane-Wright, R.I.; Pross, A. Evolution “On Purpose”: Teleonomy in Living Systems; The MIT Press: Cambridge, MA, USA, 2023. [Google Scholar]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Nonenzymatic Metabolic Reactions and Life’s Origins. Chem. Rev. 2020, 120, 7708–7744. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Pinna, S.; Kunz, C.; Halpern, A.; Harrison, S.A.; Jordan, S.F.; Ward, J.; Werner, F.; Lane, N. A Prebiotic Basis for ATP as the Universal Energy Currency. PLoS Biol. 2022, 20, e3001437. [Google Scholar] [CrossRef] [PubMed]
- Vasas, V.; Szathmáry, E.; Mauro Santos, M. Lack of Evolvability in Self-sustaining Autocatalytic Networks Constraints Metabolism-first Scenarios for the Origin of Life. Proc. Natl. Acad. Sci. USA 2010, 107, 1470–1475. [Google Scholar] [CrossRef] [PubMed]
- Zaug, A.J.; Cech, T.R. In Vitro Splicing of the Ribosomal RNA Precursor in Nuclei of Tetrahymena. Cell 1980, 19, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R. The RNA Worlds in Context. Cold Spring Harb. Perspect Biol. 2012, 4, a006742. [Google Scholar] [CrossRef]
- Bartel, D.P.; Szostak, J.W. Isolation of a New Ribozymes from a Large Pool of Random Sequences. Science 1993, 261, 1411–1418. [Google Scholar] [CrossRef]
- Johnston, W.K.; Unrau, P.J.; Lawrence, M.S.; Glasner, M.E.; Bartel, D.P. RNA-catalyzed RNA polymerization: Accurate and general RNA- templated primer extension. Science 2001, 292, 1319–1325. [Google Scholar] [CrossRef]
- Paul, N.; Joyce, G.F. A Self-replicating Ligase Ribozyme. Proc. Natl. Acad. Sci. USA 2002, 99, 12733–12740. [Google Scholar] [CrossRef] [PubMed]
- Noller, H.F.; Hoffarth, V.; Zimniak, L. Unusual Resistance of Peptidyl Transferase to Protein Extraction Procedures. Science 1992, 256, 1416–1419. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Ban, N.; Hansen, J.; Moore, P.B.; Stetiz, T.A. The Structural Basis of Ribosome Activity in Peptide Bond Synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef]
- Sousa, F.L.; Nelson-Sathi, S.; Martin, W.F. One step beyond a ribosome: The ancient anaerobic core. Biochim. Biophys. Acta 2016, 1857, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Halevy, I.; Alesker, M.; Schuster, E.; Popovitz-Biro, R.; Feldman, Y. A Key Role for Green Rust in the Precambrian Oceans and the Genesis of Iron Formations. Nat. Geosci. 2017, 10, 135–139. [Google Scholar] [CrossRef]
- Reuveni, S.; Ehrenberg, M.; Paulsson, J. Ribosomes are Optimized for Autocatalytic Production. Nature 2017, 547, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Novozhilov, A.S. Origin and Evolution of the Genetic Code: The Universal Enigma. Life 2009, 61, 99–111. [Google Scholar] [PubMed]
- Wolf, Y.I.; Koonin, E.V. On the origin of the translation system and the genetic code in the RNA world by means of natural selection, exaptation, and subfunctionalization. Biol. Direct 2007, 2, 14. [Google Scholar] [CrossRef]
- Koonin, E.V.; Novozhilo, A.S. Origin and Evolution of the Universal Genetic Code. Annu. Rev. Genet. 2017, 51, 45–62. [Google Scholar] [CrossRef]
- Gilbert, W. Origin of life: The RNA World. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Dill, K.A.; Agozzino, L. Driving forces in the origins of life. Open Biol. 2021, 11, 200324. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Coupling of Phosphorylation to Electron and Hydrogen Transfer by a Chemi-Osmotic Type of Mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.D. A Research Journey with ATP Synthase. J. Biol. Chem. 2002, 277, 39045–39061. [Google Scholar] [CrossRef] [PubMed]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial Electron Transport and Energy Conservation—The Foundation for Optimizing Bioelectrochemical Systems. Front. Microbiol. 2015, 6, 575. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Wolberg, A.; Kahwaji, C.I. Biochemistry, Electron Transport Chain; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bleichert, F.; Botchan, M.R.; Berger, J.M. Mechanisms for initiating cellular DNA replication. Science 2017, 355, eaah6317. [Google Scholar] [CrossRef] [PubMed]
- Leipe, D.D.; Aravind, L.; Koonin, E.V. Did DNA replication evolve twice independently? Nucleic Acids Res. 1999, 27, 3389–3401. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Koonin, E.V. Archaeology of Eukaryotic DNA Replication. Cold Spring Harb. Perspect Biol. 2013, 5, a012963. [Google Scholar] [CrossRef]
- Beeby, M.; Ferreira, J.L.; Tripp, P.; Albers, S.V.; Mitchell, D.R. Propulsive Nanomachines: The Convergent Evolution of Archaella, Flagella and Ailia. FEMS Microbiol. Rev. 2020, 44, 253–304. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Korenaga, J. A Wet Heterogeneous Mantle Creates a Habitable World in the Hadean. Nature 2022, 603, 86–90. [Google Scholar] [CrossRef]
- Woese, C.R. Bacterial Evolution. Microbiolocical Rev. 1987, 51, 221–271. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Gadd, G.M. Prokaryotic Metabolism and Physiology; Cambridge University Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Russell, M.J.; Arndt, N.T. Geodynamic and metabolic cycles in the Hadean. Biogeosciences 2005, 2, 97–111. [Google Scholar] [CrossRef]
- Harrison, T.M. The Hadean Crust: Evidence from >4 Ga Zircons. Annu. Rev. Earth Planet. Sci. 2009, 37, 479–505. [Google Scholar] [CrossRef]
- Oliver, T.; Kim, T.D.; Trinugroho, J.P.; Cordón-Preciado, V.; Wijayatilake, N.; Bhatia, A.; Rutherford, A.W.; Cardona, T. The Evolution and Evolvability of Photosystem II. Annu. Rev. Plant Biol. 2023, 74, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W.; Kudryavtsev, A.B.; Czaja, A.D.; Tripathi, A.B. Evidence of Archean life: Stromatolites and microfossils. Precambrian Res. 2007, 158, 141–155. [Google Scholar] [CrossRef]
- Sagan, L. On the origin of mitosing cells. J. Theor. Biol. 1967, 14, 255–274. [Google Scholar] [CrossRef] [PubMed]
- Margulis, L. Origin of Eukaryotic Cells; Freeman: San Francisco, CA, USA, 1981. [Google Scholar]
- Margulis, L. Symbiogenesis. A new principle of evolution rediscovery of Boris Mikhaylovich Kozo-Polyansky (1890–1957). Paleontol. J. 2010, 44, 1525–1539. [Google Scholar] [CrossRef]
- Doolittle, W.F. Uprooting the Tree of Life. Sci. Am. 2000, 282, 90–92. [Google Scholar] [CrossRef]
- Doolittle, W.F. Is Junk DNA Bunk? A Critique of ENCODE. Proc. Natl. Acad. Sci. USA 2013, 110, 5294–5300. [Google Scholar] [CrossRef]
- The ENCODE Project Consortium. The ENCODE (ENCyclopedia Of DNA Elements) Project. Science 2004, 306, 636–640. [Google Scholar] [CrossRef]
- Graur, D.; Zheng, Y.; Price, N.; Azevedo, R.B.; Zufall, R.A.; Elhaik, E. On the immortality of television sets: “Function” in the human genome according to the evolution-free gospel of ENCODE. Genome Biol. Evol. 2013, 5, 578–590. [Google Scholar] [CrossRef]
- Niu, D.-K.; Jiang, L. Can ENCODE tell us how much junk DNA we carry in our genome? Biochem. Biophy. Res. Comm. 2013, 430, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.; Amaral, P. RNA, the Epicenter of Genetic Information; CRC Press: Boca Raton, FL, USA, 2023. [Google Scholar] [CrossRef]
- Morillon, A. Long Non-Coding RNA. The Dark Side of the Genome; Elsevier Ltd.: Oxford, UK; ISTE Press: London, UK, 2018. [Google Scholar]
- Aebersold, R.; Agar, J.N.; Amster, I.J.; Baker, M.S.; Bertozzi, C.R.; Boja, E.S.; Costello, C.E.; Cravatt, B.F.; Fenselau, C.; Garcia, B.A.; et al. How Many Human Proteoforms are There? Nat. Chem. Biol. 2018, 14, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Noble, D. Physiology is rocking the foundations of evolutionary biology. Exp. Physiol. 2013, 98, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Laland, K.N.; Uller, T.; Feldman, M.W.; Sterelny, K.; Müller, G.B.; Moczek, A.; Jablonka, E.; Odling-Smee, J. The Extended Evolutionary Synthesis: Its Structure, Assumptions and Predictions. Proc. R. Soc. B 2015, 282, 20151019. [Google Scholar] [CrossRef] [PubMed]
- Rechavi, O.; Minevich, G.; Hobert, O. Transgenerational Inheritance of an Acquired Small RNA-Based Antiviral Response in C. elegans. Cell 2011, 147, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Noble, D.; Jablonka, E.; Joyner, M.J.; Müller, G.B.; Omholt, S.W. Evolution evolves: Physiology returns to centre stage. J. Physiol. 2014, 11, 2237–2244. [Google Scholar] [CrossRef]
- Martin, W.; Müller, M. The Hydrogen Hypothesis for the First Eukaryote. Nature 1998, 392, 37–41. [Google Scholar] [CrossRef]
- Toni Gabaldón, T. Origin and Early Evolution of the Eukaryotic Cell. Annu. Rev. Microbiol. 2021, 75, 631–647. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunk, C.F.; Marshall, C.R. Opinion: The Key Steps in the Origin of Life to the Formation of the Eukaryotic Cell. Life 2024, 14, 226. https://doi.org/10.3390/life14020226
Brunk CF, Marshall CR. Opinion: The Key Steps in the Origin of Life to the Formation of the Eukaryotic Cell. Life. 2024; 14(2):226. https://doi.org/10.3390/life14020226
Chicago/Turabian StyleBrunk, Clifford F., and Charles R. Marshall. 2024. "Opinion: The Key Steps in the Origin of Life to the Formation of the Eukaryotic Cell" Life 14, no. 2: 226. https://doi.org/10.3390/life14020226
APA StyleBrunk, C. F., & Marshall, C. R. (2024). Opinion: The Key Steps in the Origin of Life to the Formation of the Eukaryotic Cell. Life, 14(2), 226. https://doi.org/10.3390/life14020226